Spatial and Temporal Dynamics of Contact Zones Between Chromosomal Races of House Mice, Mus musculus domesticus, on Madeira Island

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Material and Methods
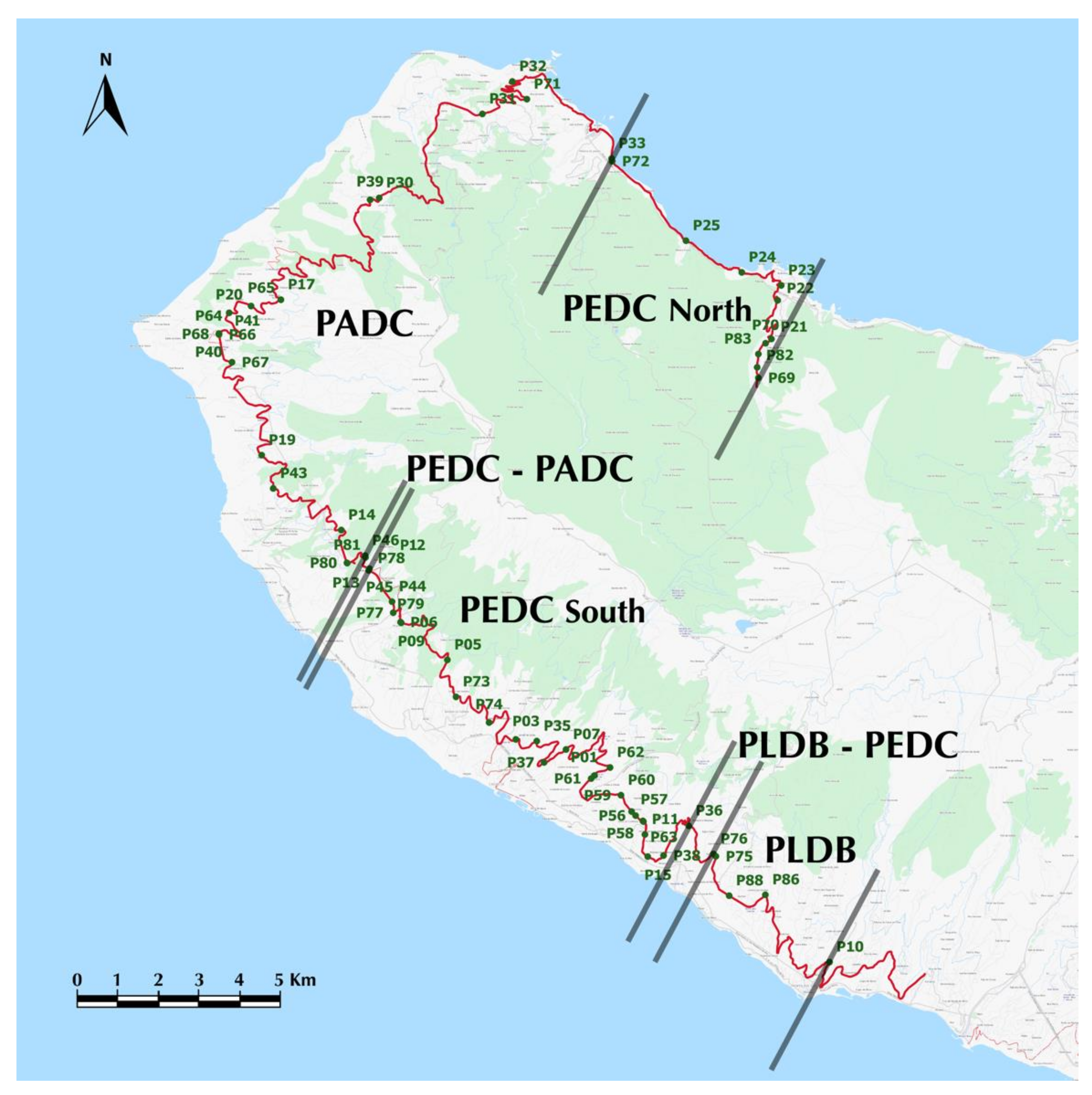
2.1. Study Area and Sampling of Mice
2.2. Chromosome Analysis
2.3. Temporal Analysis
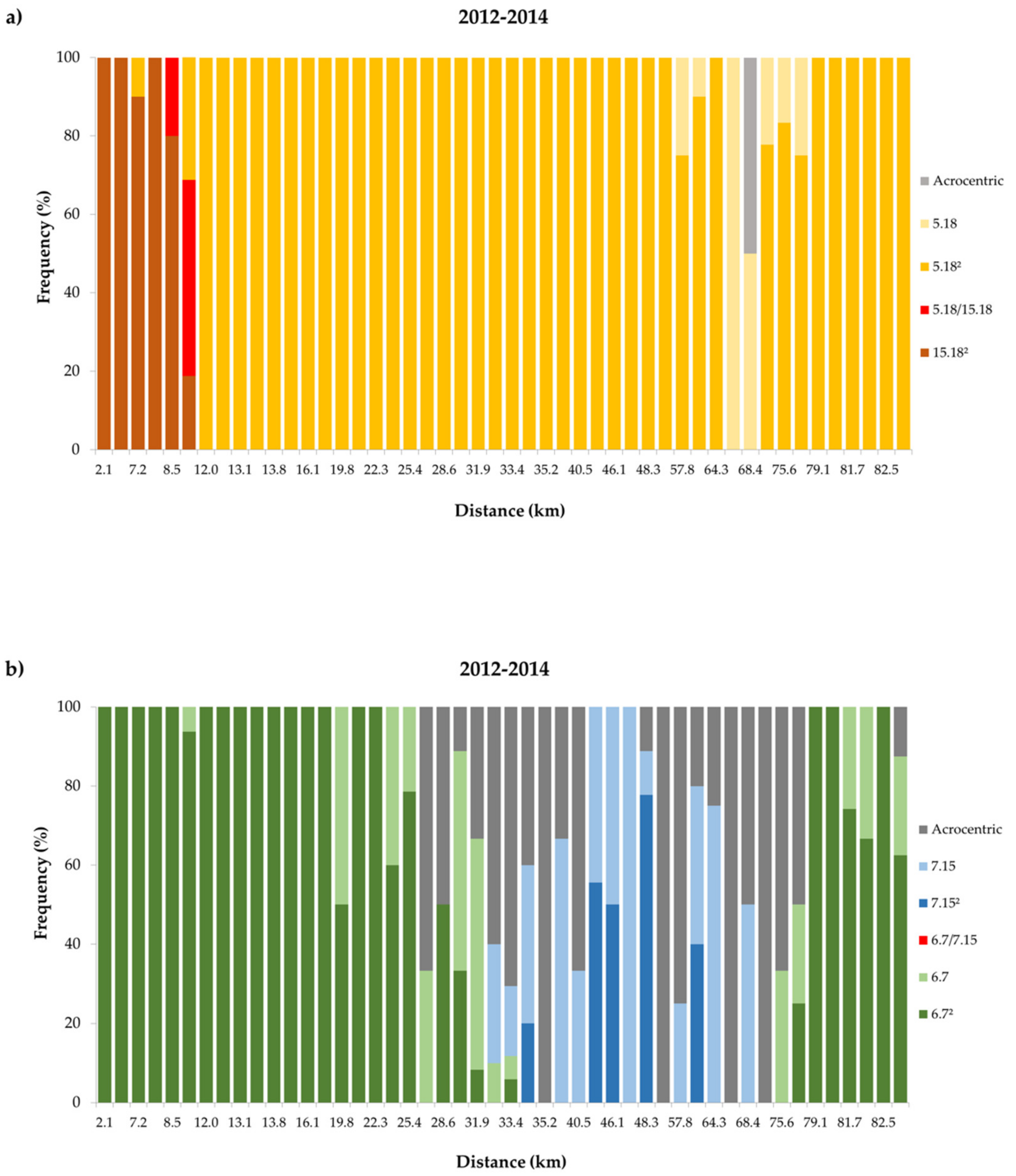
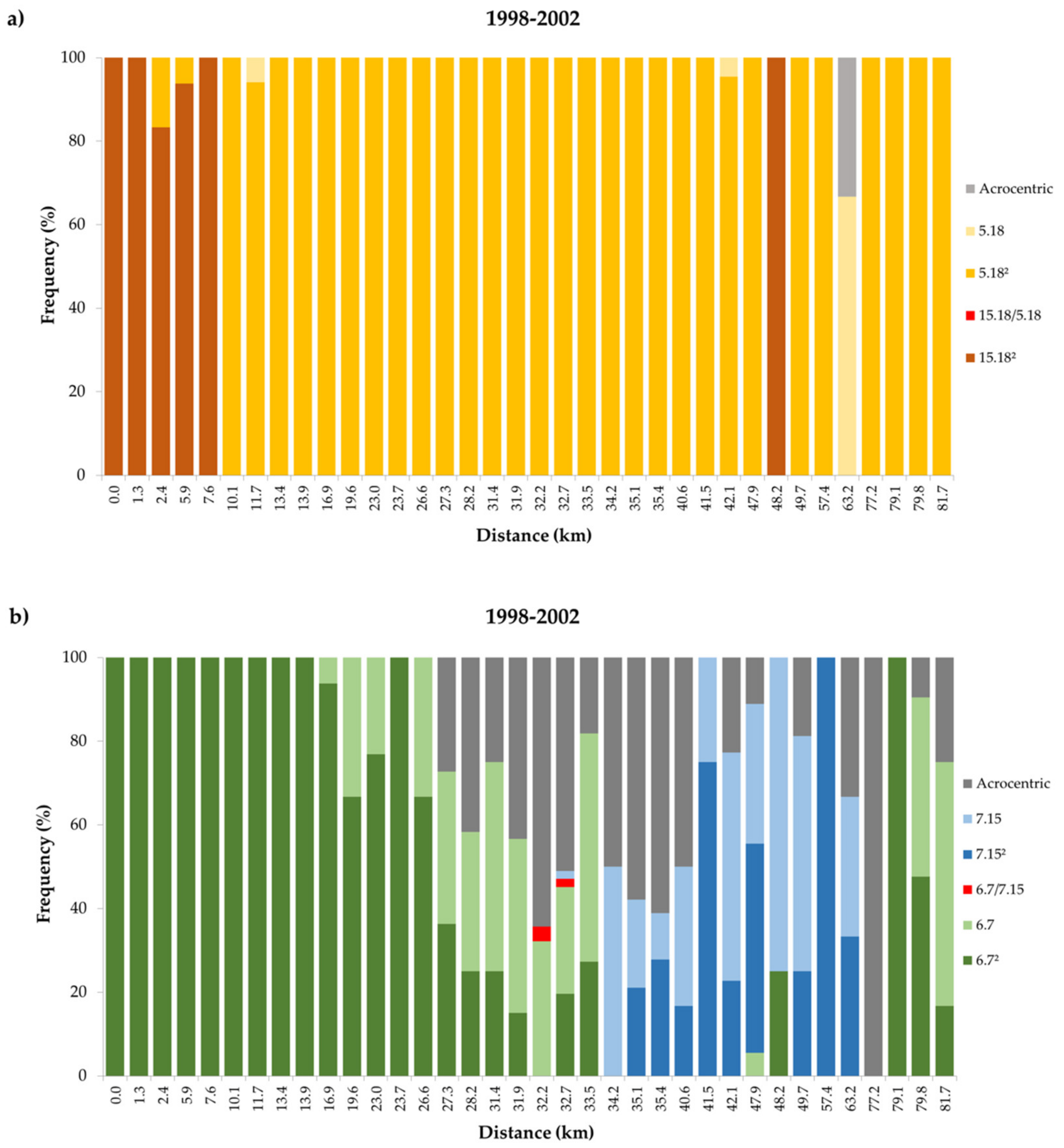
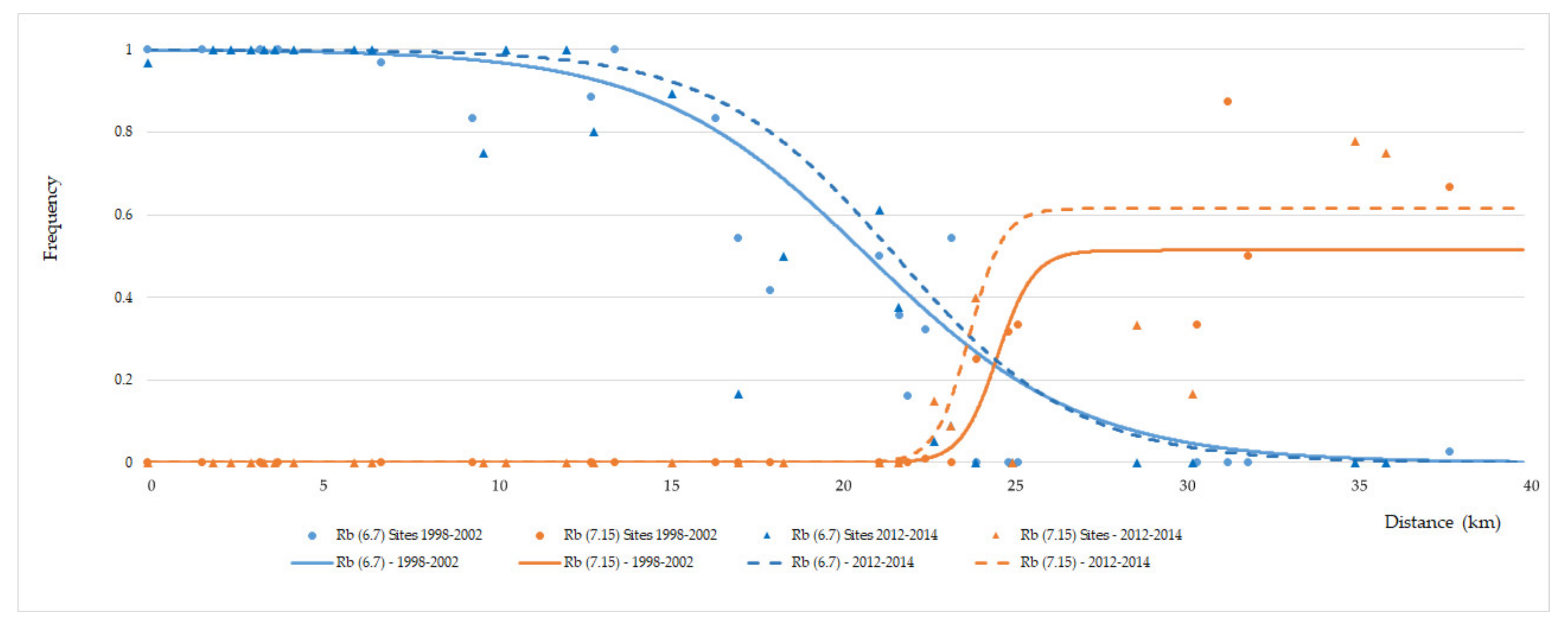
3. Results
3.1. Current Chromosomal Variation in Western Madeira
3.2. Temporal Variation in the Distribution of Madeira Rb Races
4. Discussion
4.1. Current Distribution of Estreito da Calheta (PEDC) and Achadas da Cruz (PADC) Races
4.2. Dynamics of the Contact Zone Between Estreito da Calheta (PEDC) and Achadas da Cruz (PADC) Races
4.3. Contact Zone Between Races Estreito da Calheta (PEDC) and Lugar de Baixo (PLDB)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gropp, A.; Tettenborn, U.; von Lehemann, E. Chromosomenuntersuchungen bei der Tabakmaus (Mus poschiavinus) und bei den Hybriden mit der Laboratoriummaus. Experientia 1969, 25, 875–876. [Google Scholar] [CrossRef] [PubMed]
- Piálek, J.; Hauffe, H.C.; Searle, J.B. Chromosomal variation in the house mouse: A review. Biol. J. Linn. Soc. 2005, 84, 535–563. [Google Scholar] [CrossRef] [Green Version]
- Hauffe, H.C.; Giménez, M.D.; Searle, J.B. Chromosomal hybrid zones in the house mouse. In Evolution of the House Mouse; Macholán, M., Baird, S.J., Munclinger, P., Piálek, J., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 407–430. [Google Scholar]
- Britton-Davidian, J.; Catalan, J.; Ramalhinho, M.G.; Auffray, J.C.; Nunes, A.C.; Gazave, E.; Searle, J.B.; Mathias, M.L. Chromosomal phylogeny of Robertsonian races of the house mouse on the island of Madeira: Testing between alternative mutational processes. Genet. Res. 2005, 86, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Gündüz, İ.; Pollock, C.L.; Giménez, M.D.; Förster, D.W.; White, T.A.; Sans-Fuentes, M.A.; Hauffe, H.C.; Ventura, J.; López-Fuster, M.J.; Searle, J.B. Staggered chromosomal hybrid zones in the house mouse: Relevance to reticulate evolution and speciation. Genes 2010, 1, 193–209. [Google Scholar] [CrossRef]
- Brünner, H.; Hausser, J. Genetic and karyotypic structure of a hybrid zone between the chromosomal races Cordon and Valais in the common shrew Sorex araneus. Hereditas 1996, 125, 147–158. [Google Scholar] [CrossRef]
- Franchini, P.; Castiglia, R.; Capanna, E. Reproductive isolation between chromosomal races of the house mouse Mus musculus domesticus in a parapatric contact area revealed by an analysis of multiple unlinked loci. J. Evol. Biol. 2008, 21, 502–513. [Google Scholar] [CrossRef]
- Franchini, P.; Colangelo, P.; Solano, E.; Capanna, E.; Verheyen, E.; Castiglia, R. Reduced gene flow at pericentromeric loci in a hybrid zone involving chromosomal races of the house mouse Mus musculus domesticus. Evolution 2010, 64, 2020–2032. [Google Scholar]
- Giménez, M.D.; Förster, D.W.; Jones, E.P.; Jóhannesdóttir, F.; Gabriel, S.I.; Panithanarak, T.; Scascitelli, M.; Merico, V.; Garagna, S.; Searle, J.B.; et al. A half century of studies on a chromosomal hybrid zone of the house mouse. J. Hered. 2017, 108, 25–35. [Google Scholar] [CrossRef] [Green Version]
- White, M.J.D. Chain processes in chromosomal speciation. Syst. Zool. 1978, 27, 285–298. [Google Scholar] [CrossRef]
- Rieseberg, L.H. Chromosomal rearrangements and speciation. Trends Ecol. Evol. 2001, 16, 351–358. [Google Scholar] [CrossRef]
- Panithanarak, T.; Hauffe, H.C.; Dallas, J.F.; Glover, A.; Ward, R.G.; Searle, J.B. Linkage-dependent gene flow in a house mouse chromosomal hybrid zone. Evolution 2004, 58, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Ganem, G.; Searle, J.B. Behavioural discrimination among chromosomal races of the house mouse (Mus musculus domesticus). J. Evol. Biol. 1996, 9, 817–831. [Google Scholar] [CrossRef]
- Castiglia, R.; Capanna, E. Contact zones between chromosomal races of Mus musculus domesticus. 1. Temporal analysis of a hybrid zone between the CD chromosomal race (2n = 22) and populations with the standard karyotype. Heredity 1999, 83, 319–326. [Google Scholar] [CrossRef]
- Chatti, N.; Ganem, G.; Benzekri, K.; Catalan, J.; Britton-Davidian, J.; Saïd, K. Microgeographical distribution of two chromosomal races of house mice in Tunisia: Pattern and origin of habitat partitioning. Proc. R. Soc. Lond. B 1999, 266, 1561–1569. [Google Scholar] [CrossRef]
- Ratkiewicz, M.; Banaszek, A.; Jadwiszczak, W.; Chetnicki, W.; Fedyk, S. Genetic diversity, stability of population structure and barriers to gene flow in a hybrid zone between two Sorex araneus chromosome races. Mammalia 2003, 68, 275–283. [Google Scholar] [CrossRef]
- Nunes, A.C.; Britton-Davidian, J.; Catalan, J.; Ramalhinho, M.G.; Capela, R.; Mathias, M.L.; Ganem, G. Influence of physical environmental characteristics and anthropogenic factors on the position and structure of a contact zone between two chromosomal races of the house mouse on the island of Madeira (North Atlantic, Portugal). J. Biogeogr. 2005, 32, 2123–2134. [Google Scholar] [CrossRef]
- Ganem, G.; Litel, C.; Lenormand, T. Variation in mate preference across a house mouse hybrid zone. Heredity 2008, 100, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Nosil, P. Ernst Mayr and the integration of geographic and ecological factors in speciation. Biol. J. Linn. Soc. 2008, 95, 26–46. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.C.; Mathias, M.L.; Ganem, G. Odor preference in house mice: Influences of habitat heterogeneity and chromosomal incompatibility. Behav. Ecol. 2009, 20, 1252–1261. [Google Scholar] [CrossRef] [Green Version]
- Hiadlovská, Z.; Strnadová, M.; Macholán, M.; Bímová, B.V. Is water really a barrier for the house mouse? A comparative study of two mouse subspecies. Folia Zool. 2012, 61, 319–329. [Google Scholar] [CrossRef]
- Wierzbicki, H.; Moska, M.; Strzala, T.; Macierzynska, A. Do aquatic barriers reduce male-mediated gene flow in a hybrid zone of the common shrew (Sorex araneus)? Hereditas 2011, 148, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Britton-Davidian, J.; Catalan, J.; Ramalhinho, M.G.; Ganem, G.; Auffray, J.C.; Capela, R.; Biscoito, M.; Searle, J.B.; Mathias, M.L. Rapid chromosomal evolution in island mice. Nature 2000, 403, 158. [Google Scholar] [CrossRef]
- Dureje, L.; Macholán, M.; Baird, S.J.E.; Piálek, J. The mouse hybrid zone in central Europe: From morphology to molecules. Folia Zool. 2012, 61, 308–318. [Google Scholar] [CrossRef]
- Chambers, L.K.; Singleton, G.R.; van Wensveen, M. Spatial heterogeneity in wild populations of house mice (Mus domesticus) on the Darling Downs, south-eastern Queensland. Wildl. Res. 1996, 23, 23–38. [Google Scholar] [CrossRef]
- Dias, P.C.; Verheyen, G.R.; Raymond, M. Source–sink populations in Mediterranean blue tits: Evidence using single-locus minisatellite probes. J. Evol. Biol. 1996, 9, 965–978. [Google Scholar] [CrossRef]
- Hauffe, H.C.; Panithanarak, T.; Dallas, J.F.; Piálek, J.; Gündüz, İ.; Searle, J.B. The tobacco mouse and its relatives: A ‘tail’ of coat colours, chromosomes, hybridization and speciation. Cytogenet. Genome Res. 2004, 105, 395–405. [Google Scholar] [CrossRef]
- Ramalhinho, M.G.; Braz, C.; Catalan, J.; Mathias, M.L.; Britton-Davidian, J. AgNOR variability among Robertsonian races of the house mouse from the island of Madeira: Implications for patterns of Rb fusion formation and genetic differentiation. Biol. J. Linn. Soc. 2005, 84, 585–591. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.R.; Elder, F.F.B. Yeast stimulation of bone marrow mitosis for cytogenetic preparations. Cytogenet. Cell Genet. 1980, 26, 36–40. [Google Scholar] [CrossRef]
- Seabright, M. A rapid banding technique for human chromosomes. Lancet 1971, 2, 971–972. [Google Scholar] [CrossRef]
- Cowell, J.K. A photographic representation of the variability of G-banded structure of the chromosomes of the mouse karyotype. Chromosoma 1984, 89, 294–320. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Fel-Clair, F.; Lenormand, T.; Catalan, J.; Grobert, J.; Orth, A.; Boursot, P.; Viroux, M.C.; Britton-Davidian, J. Genomic incompatibilities in the hybrid zone between house mice in Denmark: Evidence from steep and non-coincident chromosomal clines for Robertsonian fusions. Genet. Res. 1996, 67, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Sites, J.W.; Moritz, C. Chromosomal evolution and speciation revisited. Syst. Zool. 1987, 36, 153–174. [Google Scholar] [CrossRef]
- Bidau, C.J.; Giménez, M.D.; Palmer, C.L.; Searle, J.B. The effects of Robertsonian fusions on chiasma frequency and distribution in the house mouse (Mus musculus domesticus) from a hybrid zone in northern Scotland. Heredity 2001, 87, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Gazave, E.; Catalan, J.; Ramalhinho, M.G.; Mathias, M.L.; Nunes, A.C.; Dumas, D.; Britton-Davidian, J.; Auffray, J.C. The non-random occurrence of Robertsonian fusion in the house mouse. Genet. Res. 2003, 81, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Sans-Fuentes, M.A.; Muñoz-Muñoz, F.; Ventura, J.; López-Fuster, M.J. Rb (7.17) a rare Robertsonian fusion in wild populations of the house mouse. Genet. Res. 2007, 89, 207–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adolph, S.; Klein, J. Genetic variation of wild mouse populations in southern Germany. I. Cytogenetic study. Genet. Res. 1983, 41, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Jiggins, C.D.; Mallet, J. Bimodal hybrid zones and speciation. Trends Ecol. Evol. 2000, 15, 250–255. [Google Scholar] [CrossRef]
- Castiglia, R.; Annesi, F.; Capanna, E. Contact zones between chromosomal races of Mus musculus domesticus. 3. Molecular and chromosomal evidence of restricted gene flow between the CD race (2n = 22) and the ACR race (2n = 24). Heredity 2002, 89, 219–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, N.H. The dynamics of hybrid zones. Heredity 1979, 43, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Barton, N.H.; Hewitt, G.M. Analysis of hybrid zones. Annu. Rev. Ecol. Syst. 1985, 16, 113–148. [Google Scholar] [CrossRef]
- Barton, N.H.; Hewitt, G.M. Adaptation, speciation and hybrid zones. Nature 1989, 341, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Barton, N.H. Chromosomal speciation and molecular divergence-accelerated evolution in rearranged chromosomes. Science 2003, 300, 321–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capilla, L.; Medarde, N.; Alemany-Schmidt, A.; Oliver-Bonet, M.; Ventura, J.; Ruiz-Herrera, A. Genetic recombination variation in wild Robertsonian mice: On the role of chromosomal fusions and Prdm9 allelic background. Proc. R. Soc. B 2014, 281, 20140297. [Google Scholar] [CrossRef] [Green Version]
- Dumas, D.; Britton-Davidian, J. Chromosomal rearrangements and evolution of recombination: Comparison of chiasma distribution patterns in standard and Robertsonian populations of the house mouse. Genetics 2002, 162, 1355–1366. [Google Scholar]
- Giménez, M.D.; White, T.A.; Hauffe, H.C.; Panithanarak, T.; Searle, J.B. Understanding the basis of diminished gene flow between hybridizing chromosome races of the house mouse. Evolution 2013, 67, 1446–1462. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.C.; Catalan, J.; Lopez, J.; Ramalhinho, M.G.; Mathias, M.L.; Britton-Davidian, J. Fertility assessment in hybrids between monobrachially homologous Rb races of the house mouse from the island of Madeira: Implications for modes of chromosomal evolution. Heredity 2011, 106, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Donahue, M.J.; Holyoak, M.; Feng, C. Patterns of dispersal and dynamics among habitat patches varying in quality. Am. Nat. 2003, 162, 302–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.M. Source-sink dynamics in a temporally heterogeneous environment. Ecology 2004, 85, 2037–2045. [Google Scholar] [CrossRef]
- Chmátal, L.; Gabriel, S.I.; Mitsainas, G.P.; Martínez-Vargas, J.; Ventura, J.; Searle, J.B.; Schultz, R.M.; Lampson, M.A. Centromere strength provides the cell biological basis for meiotic drive and karyotype evolution in mice. Curr. Biol. 2014, 24, 2295–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medarde, N.; López-Fuster, M.J.; Muñoz-Muñoz, F.; Ventura, J. Spatio-temporal variation in the structure of a chromosomal polymorphism zone in the house mouse. Heredity 2012, 109, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pocock, M.J.; Hauffe, H.C.; Searle, J.B. Dispersal in house mice. Biol. J. Linn. Soc. 2005, 84, 565–583. [Google Scholar] [CrossRef] [Green Version]
- Górecki, A.; Meczeva, R.; Pis, T.; Gerasimov, S.; Walkowa, W. Geographical variation of thermoregulation in wild populations of Mus musculus and Mus spretus. Acta Theriol. 1990, 35, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Mathias, M.L.; Nunes, A.C.; Marques, C.C.; Auffray, J.C.; Britton-Davidian, J.; Ganem, G.; Gündüz, İ.; Ramalhinho, M.G.; Searle, J.B.; Speakman, J. Effects of climate on oxygen consumption and energy intake of chromosomally divergent populations of the house mouse (Mus musculus domesticus) from the island of Madeira (North Atlantic, Portugal). Funct. Ecol. 2006, 20, 330–339. [Google Scholar] [CrossRef]
- McNab, B.K. Minimizing energy expenditure facilitates vertebrate persistence on oceanic islands. Ecol. Lett. 2002, 5, 693–704. [Google Scholar] [CrossRef]
- Teeter, K.C.; Payseur, B.A.; Harris, L.W.; Bakewell, M.A.; Thibodeau, L.M.; O’Brien, J.E.; Krenz, J.G.; Sans-Fuentes, M.A.; Nachman, M.W.; Tucker, P.K. Genome-wide patterns of gene flow across a house mouse hybrid zone. Genome Res. 2008, 18, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Turner, L.M.; Harr, B. Genome-wide mapping in a house mouse hybrid zone reveals hybrid sterility loci and Dobzhansky-Muller interactions. eLife 2014, 3, e02504. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clines | Centre | Confidence Interval | Width | Confidence Interval |
|---|---|---|---|---|
| Rb 6.7 1998–2002 | 19.07 | 18.37 - 19.76 | 12.48 | 10.70 - 14.95 |
| Rb 6.7 2012–2014 | 18.26 | 17.22 - 19.30 | 10.61 | 8.73 - 13.52 |
| Rb 7.15 1998–2002 | 24.71 | 24.16 - 25.26 | 2.09 | 1.49 - 3.52 |
| Rb 7.15 2012–2014 | 23.89 | 23.45 - 24.33 | 1.96 | 1.19 - 5.52 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapisso, J.T.; Gabriel, S.I.; Cerveira, A.M.; Britton-Davidian, J.; Ganem, G.; Searle, J.B.; Ramalhinho, M.d.G.; Mathias, M.d.L. Spatial and Temporal Dynamics of Contact Zones Between Chromosomal Races of House Mice, Mus musculus domesticus, on Madeira Island. Genes 2020, 11, 748. https://doi.org/10.3390/genes11070748
Tapisso JT, Gabriel SI, Cerveira AM, Britton-Davidian J, Ganem G, Searle JB, Ramalhinho MdG, Mathias MdL. Spatial and Temporal Dynamics of Contact Zones Between Chromosomal Races of House Mice, Mus musculus domesticus, on Madeira Island. Genes. 2020; 11(7):748. https://doi.org/10.3390/genes11070748
Chicago/Turabian StyleTapisso, Joaquim T., Sofia I. Gabriel, Ana Mota Cerveira, Janice Britton-Davidian, Guila Ganem, Jeremy B. Searle, Maria da Graça Ramalhinho, and Maria da Luz Mathias. 2020. "Spatial and Temporal Dynamics of Contact Zones Between Chromosomal Races of House Mice, Mus musculus domesticus, on Madeira Island" Genes 11, no. 7: 748. https://doi.org/10.3390/genes11070748