Bioaccessibility of Bioactive Compounds of ‘Fresh Garlic’ and ‘Black Garlic’ through In Vitro Gastrointestinal Digestion

 ,
,  , , ,
, , ,  and
and
Abstract
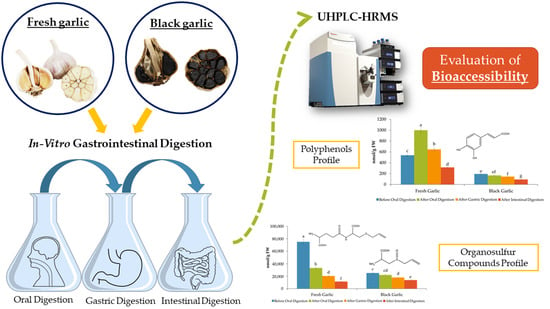
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Materials and Sample Preparation
2.3. In Vitro Gastrointestinal Digestion
2.4. Phenolic and Organosulfur Compounds Extraction and Analysis
2.5. Bioaccessibility of (Poly)Phenols and Organosulfur Compounds
2.6. Statistical Analysis
3. Results and Discussion
3.1. Bioaccessibility of Phenolic Compounds after Simulated Gastrointestinal Digestion
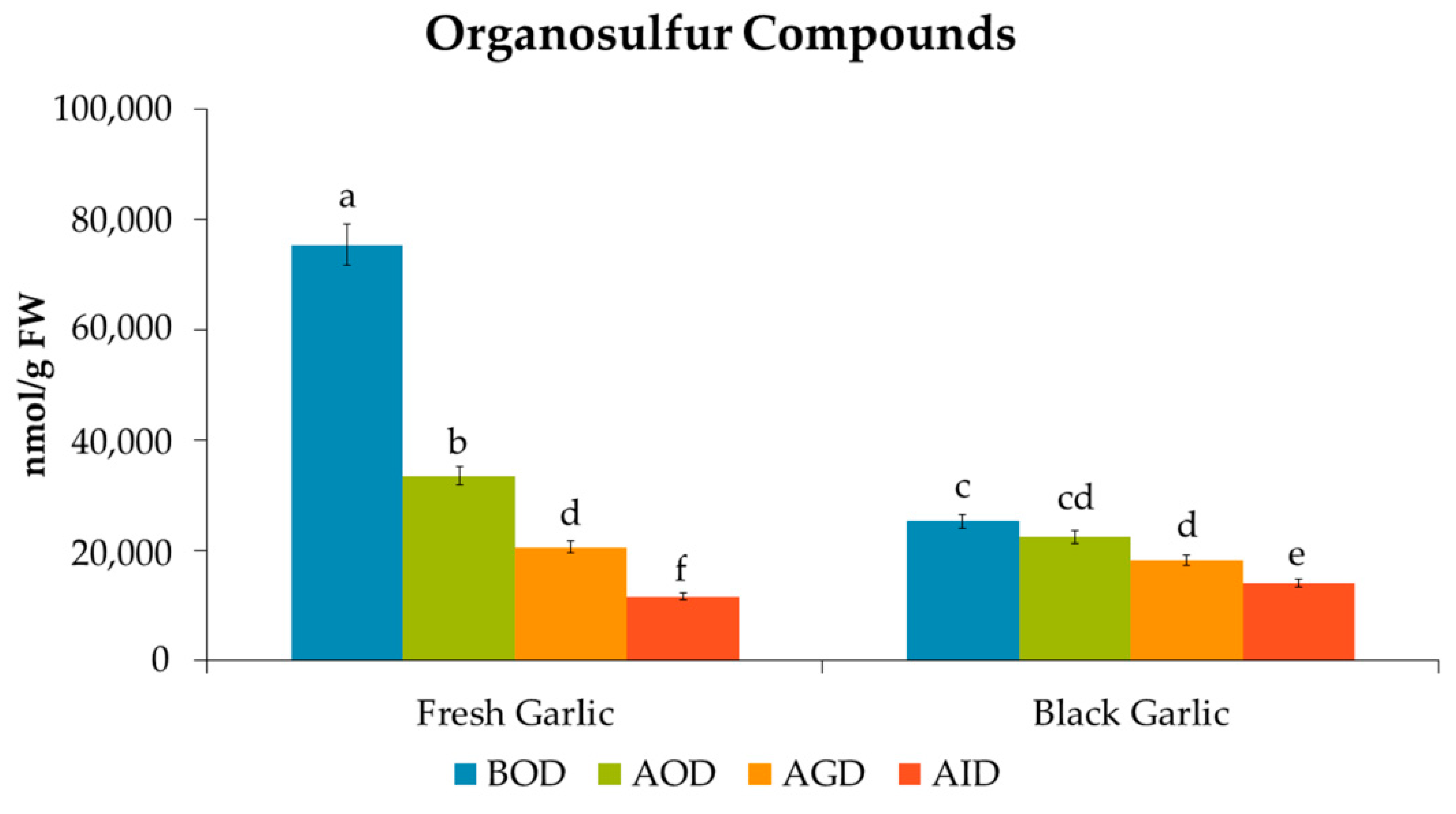
3.2. Bioaccessibility of Organosulfur Compounds after Simulated Gastrointestinal Digestion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Metro, D.; Tardugno, R.; Papa, M.; Bisignano, C.; Manasseri, L.; Calabrese, G.; Gervasi, T.; Dugo, G.; Cicero, N. Adherence to the Mediterranean diet in a Sicilian student population. Nat. Prod. Res. 2017, 32, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Metro, D.; Papa, M.; Manasseri, L.; Gervasi, T.; Campone, L.; Pellizzeri, V.; Tardugno, R.; Dugo, G. Mediterranean diet in a Sicilian student population. Second part: Breakfast and its nutritional profile. Nat. Prod. Res. 2018, 34, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, H.L.; Ross, S.A.; Milner, J.A. Garlic and onions: Their cancer prevention properties. Cancer Prev. Res. 2015, 8, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Li, Y.; Yang, J.; Pu, X.; Du, J.; Yang, X.; Yang, T.; Yang, S. Therapeutic Role of Functional Components in Alliums for Preventive Chronic Disease in Human Being. Evidence-Based Complement. Altern. Med. 2017, 2017, 1–13. [Google Scholar] [CrossRef]
- Quesada, I.; de Paola, M.; Torres-Palazzolo, C.; Camargo, A.; Ferder, L.; Manucha, W.; Castro, C. Effect of Garlic’s Active Constituents in Inflammation, Obesity and Cardiovascular Disease. Curr. Hypertens. Rep. 2020, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.; Ruan, J.; Zhuang, X.; Zhang, X.; Li, Z. Phytochemicals of garlic: Promising candidates for cancer therapy. Biomed. Pharmacother. 2020, 123, 109730. [Google Scholar] [CrossRef]
- Choudhary, P.R.; Jani, R.D.; Sharma, M.S. Effect of Raw Crushed Garlic (Allium sativum L.) on Components of Metabolic Syndrome. J. Diet. Suppl. 2018, 15, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K. Garlic and aging: New insights into an old remedy. Ageing Res. Rev. 2003, 2, 39–56. [Google Scholar] [CrossRef]
- Zhai, B.; Zhang, C.; Sheng, Y.; Zhao, C.; He, X.; Xu, W.; Huang, K.; Luo, Y. Hypoglycemic and hypolipidemic effect of S-allyl-cysteine sulfoxide (alliin) in DIO mice. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kimura, S.; Tung, Y.C.; Pan, M.H.; Su, N.W.; Lai, Y.J.; Cheng, K.C. Black garlic: A critical review of its production, bioactivity, and application. J. Food Drug Anal. 2017, 25, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.H.; Kang, D. Physicochemical properties, biological activity, health benefits, and general limitations of aged black garlic: A review. Molecules 2017, 22, 919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colín-González, A.L.; Santana, R.A.; Silva-Islas, C.A.; Chánez-Cárdenas, M.E.; Santamaría, A.; Maldonado, P.D. The antioxidant mechanisms underlying the aged garlic extract- and S-allylcysteine-induced protection. Oxid. Med. Cell. Longev. 2012, 2012, 1–16. [Google Scholar] [CrossRef]
- Martinez-Casas, L.; Lage-Yusty, M.; Lopez-Hernandez, J. Changes in aromatic profile, sugars and bioactive compounds when purple garlic is transformed into black garlic. J. Agric. Food Chem. 2017, 65, 10804–10811. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.A.; Sutherland, W.H.F.; McCormick, M.P.; Yeoman, D.J.; De Jong, S.A. Aged garlic extract improves endothelial function in men with coronary artery disease. Phyther. Res. 2005, 19, 314–319. [Google Scholar] [CrossRef]
- Tanaka, S.; Haruma, K.; Yoshihara, M.; Kajiyama, G.; Kira, K.; Amagase, H.; Chayama, K. Aged Garlic Extract Has Potential Suppressive Effect on Colorectal Adenomas in Humans. J. Nutr. 2006, 136, 821S–826S. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, K.; Kinninger, A.; Cherukuri, L.; Birudaraju, D.; Nakanishi, R.; Almeida, S.; Jayawardena, E.; Shekar, C.; Flores, F.; Hamal, S.; et al. Aged garlic extract reduces low attenuation plaque in coronary arteries of patients with diabetes: A randomized, double-blind, placebo-controlled study. Exp. Ther. Med. 2019, 19, 1457–1461. [Google Scholar] [CrossRef] [Green Version]
- Wlosinska, M.; Nilsson, A.C.; Hlebowicz, J.; Hauggaard, A.; Kjellin, M.; Fakhro, M.; Lindstedt, S. The effect of aged garlic extract on the atherosclerotic process-A randomized double-blind placebo-controlled trial. BMC Complement. Med. Ther. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Lingua, M.S.; Theumer, M.G.; Kruzynski, P.; Wunderlin, D.A.; Baroni, M.V. Bioaccessibility of polyphenols and antioxidant properties of the white grape by simulated digestion and Caco-2 cell assays: Comparative study with its winemaking product. Food Res. Int. 2019, 122, 496–505. [Google Scholar] [CrossRef]
- Castaldo, L.; Narváez, A.; Izzo, L.; Graziani, G.; Ritieni, A. In vitro bioaccessibility and antioxidant activity of coffee silverskin polyphenolic extract and characterization of bioactive compounds using UHPLC-Q-Orbitrap HRMS. Molecules 2020, 25, 2132. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A. Influence of Food Matrix on the Bioaccessibility of Fruit Polyphenolic Compounds. J. Agric. Food Chem. 2020, 68, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.G.S.; Rebellato, A.P.; dos Santos Caramês, E.T.; Greiner, R.; Pallone, J.A.L. In vitro digestion effect on mineral bioaccessibility and antioxidant bioactive compounds of plant-based beverages. Food Res. Int. 2020, 130, 108993. [Google Scholar] [CrossRef] [PubMed]
- García-Hernández, J.; Hernández-Pérez, M.; Peinado, I.; Andrés, A.; Heredia, A. Tomato-antioxidants enhance viability of L. reuteri under gastrointestinal conditions while the probiotic negatively affects bioaccessibility of lycopene and phenols. J. Funct. Foods 2018, 43, 1–7. [Google Scholar] [CrossRef]
- Torres-Palazzolo, C.; Ramirez, D.; Locatelli, D.; Manucha, W.; Castro, C.; Camargo, A. Bioaccessibility and permeability of bioactive compounds in raw and cooked garlic. J. Food Compos. Anal. 2018, 70, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Ortega, A.; Pereira-Caro, G.; Luis Ordóñez, J.; Manuel Muñoz-Redondo, J.; Moreno-Rojas, R.; Pérez-Aparicio, J.; Manuel Moreno-Rojas, J. Changes in the antioxidant activity and metabolite profile of three onion varieties during the elaboration of ‘black onion’. Food Chem. 2020, 311, 1–9. [Google Scholar] [CrossRef]
- Juániz, I.; Ludwig, I.A.; Bresciani, L.; Dall’Asta, M.; Mena, P.; Del Rio, D.; Cid, C.; de Peña, M.P. Bioaccessibility of (poly)phenolic compounds of raw and cooked cardoon (Cynara cardunculus L.) after simulated gastrointestinal digestion and fermentation by human colonic microbiota. J. Funct. Foods 2017, 32, 195–207. [Google Scholar] [CrossRef]
- Moreno-Rojas, J.M.; Moreno-Ortega, A.; Ordóñez, J.L.; Moreno-Rojas, R.; Pérez-Aparicio, J.; Pereira-Caro, G. Development and validation of UHPLC-HRMS methodology for the determination of flavonoids, amino acids and organosulfur compounds in black onion, a novel derived product from fresh shallot onions (Allium cepa var. aggregatum). LWT 2018, 97, 376–383. [Google Scholar] [CrossRef]
- Ortega, N.; Macià, A.; Romero, M.P.; Reguant, J.; Motilva, M.J. Matrix composition effect on the digestibility of carob flour phenols by an in vitro digestion model. Food Chem. 2011, 124, 65–71. [Google Scholar] [CrossRef]
- Gomes, T.M.; Toaldo, I.M.; Haas, I.C. da S.; Burin, V.M.; Caliari, V.; Luna, A.S.; de Gois, J.S.; Bordignon-Luiz, M.T. Differential contribution of grape peel, pulp, and seed to bioaccessibility of micronutrients and major polyphenolic compounds of red and white grapes through simulated human digestion. J. Funct. Foods 2019, 52, 699–708. [Google Scholar] [CrossRef]
- Kim, J.S.; Kang, O.J.; Gweon, O.C. Comparison of phenolic acids and flavonoids in black garlic at different thermal processing steps. J. Funct. Foods 2013, 5, 80–86. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. Soymilk phenolic compounds, isoflavones and antioxidant activity as affected by in vitro gastrointestinal digestion. Food Chem. 2013, 136, 206–212. [Google Scholar] [CrossRef]
- Lucas-González, R.; Viuda-Martos, M.; Pérez Álvarez, J.A.; Fernández-López, J. Changes in bioaccessibility, polyphenol profile and antioxidant potential of flours obtained from persimmon fruit (Diospyros kaki) co-products during in vitro gastrointestinal digestion. Food Chem. 2018, 256, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Blancas-Benitez, F.J.; Mercado-Mercado, G.; Quirós-Sauceda, A.E.; Montalvo-González, E.; González-Aguilar, G.A.; Sáyago-Ayerdi, S.G. Bioaccessibility of polyphenols associated with dietary fiber and in vitro kinetics release of polyphenols in Mexican “Ataulfo” mango (Mangifera indica L.) by-products. Food Funct. 2015, 6, 859–868. [Google Scholar] [CrossRef]
- Alminger, M.; Aura, A.M.; Bohn, T.C.; Dufour, S.N.E.L.; Gomes, A.; Karakaya, S.; Martínez-Cuesta, M.C.; McDougall, G.J.; Requena, T.; Santos, C.N. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Yang, Y.; Gao, J.; Feng, J.; Shang, Y.; Wei, Z. Phenolics and antioxidant activity of bamboo leaves soup as affected by in vitro digestion. Food Chem. Toxicol. 2020, 135, 110941. [Google Scholar] [CrossRef]
- Goncharov, N.; Orekhov, A.N.; Voitenko, N.; Ukolov, A.; Jenkins, R.; Avdonin, P. Organosulfur Compounds as Nutraceuticals. In Nutraceuticals: Efficacy, Safety and Toxicity; Elsevier Inc.: San Diego, CA, USA, 2016; pp. 555–568. ISBN 9780128021477. [Google Scholar]
- Verhagen, H.; Hageman, G.J.; Rauma, A.-L.; Versluis-de Haan, G.; van Herwijnen, M.H.M.; de Groot, J.; Törrönen, R.; Mykkänen, H. Biomonitoring the intake of garlic via urinary excretion of allyl mercapturic acid. Br. J. Nutr. 2001, 86, S111–S114. [Google Scholar] [CrossRef] [Green Version]
- Trio, P.Z.; You, S.; He, X.; He, J.; Sakao, K.; Hou, D.-X. Chemopreventive functions and molecular mechanisms of garlic organosulfur compounds. Food Funct. 2014, 5, 833. [Google Scholar] [CrossRef]
- Fujii, T.; Matsutomo, T.; Kodera, Y. Changes of S-Allylmercaptocysteine and γ-Glutamyl- S-allylmercaptocysteine Contents and Their Putative Production Mechanisms in Garlic Extract during the Aging Process. J. Agric. Food Chem. 2018, 66, 10506–10512. [Google Scholar] [CrossRef]
- Molina-Calle, M.; de Medina, V.S.; Priego-Capote, F.; de Castro, M.D.L. Establishing compositional differences between fresh and black garlic by a metabolomics approach based on LC–QTOF MS/MS analysis. J. Food Compos. Anal. 2017, 62, 155–163. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, D.S.; Kyung, K.H. Factors influencing the stability of garlic thiosulfinates. Food Sci. Biotechnol. 2014, 23, 1593–1600. [Google Scholar] [CrossRef]
- Lee, B.C.; Gladyshev, V.N. The biological significance of methionine sulfoxide stereochemistry. Free Radic. Biol. Med. 2011, 50, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Xing, F.; Liu, Y.; Lv, Y.; Wang, X.; Ling, M.T.; Gao, H.; Ouyang, S.; Yang, M.; Zhu, J.; et al. Garlic-derived compound S-allylmercaptocysteine inhibits hepatocarcinogenesis through targeting LRP6/Wnt pathway. Acta Pharm. Sin. B 2018, 8, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; So, K.F.; Wong, N.K.; Xiao, J. Anti-cancer activities of S-allylmercaptocysteine from aged garlic. Chin. J. Nat. Med. 2019, 17, 43–49. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | BOD | AOD | % Recovery | AGD | % Recovery | AID | % Recovery-Bioaccessibility | p-Value |
|---|---|---|---|---|---|---|---|---|
| Fresh Garlic | ||||||||
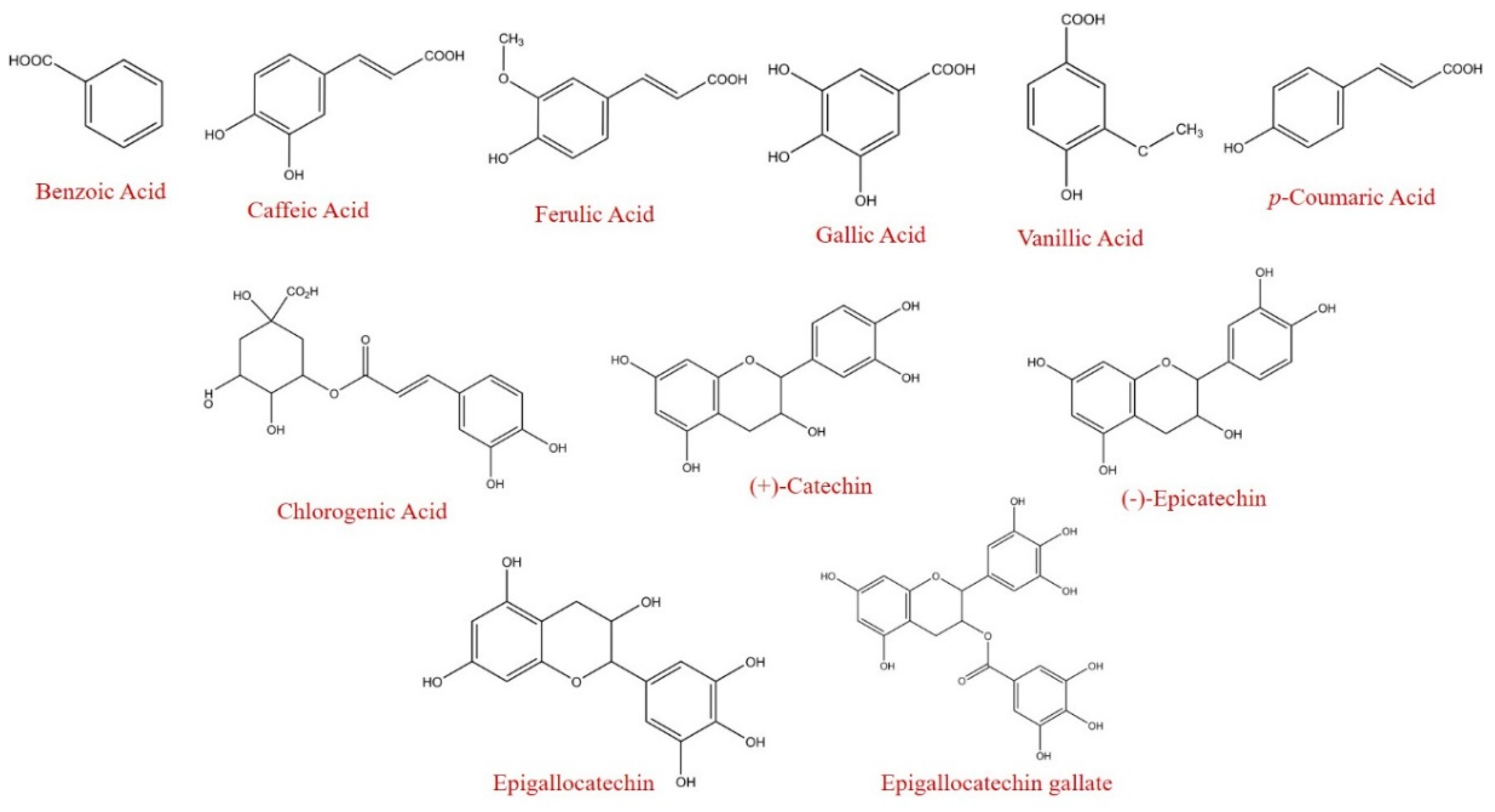
| Benzoic Acid | nq | 162 ab | - | 141 b | - | 173 a | - | *** |
| Vanillic acid | 18.3 a | 15.5 a | 84.7 | 11.7 b | 63.9 | 12.7 b | 69.4 | *** |
| Gallic acid | 58.7 b | 105 a | 180 | 119 a | 203 | 14.8 c | 25.2 | *** |
| Caffeic acid | 455 b | 712 a | 156 | 370 b | 81.3 | 112 c | 24.6 | *** |
| Ferulic Acid | 6.8 a | 5.6 b | 82.4 | 3.8 c | 55.9 | 3.5 c | 51.5 | *** |
| (+)-Catechin | nq | nq | - | 0.01 b | - | 0.11 a | - | *** |
| (−)-Epicatechin | nq | 0.092 a | - | 0.04 b | - | 0.03 c | - | *** |
| Epigallocatechin | 0.19 c | 0.39 b | 203 | 0.44 a | 230.0 | 0.07 d | 35.3 | *** |
| Chlorogenic acid | nq | 0.054 a | - | 0.04 b | - | 0.02 c | - | *** |
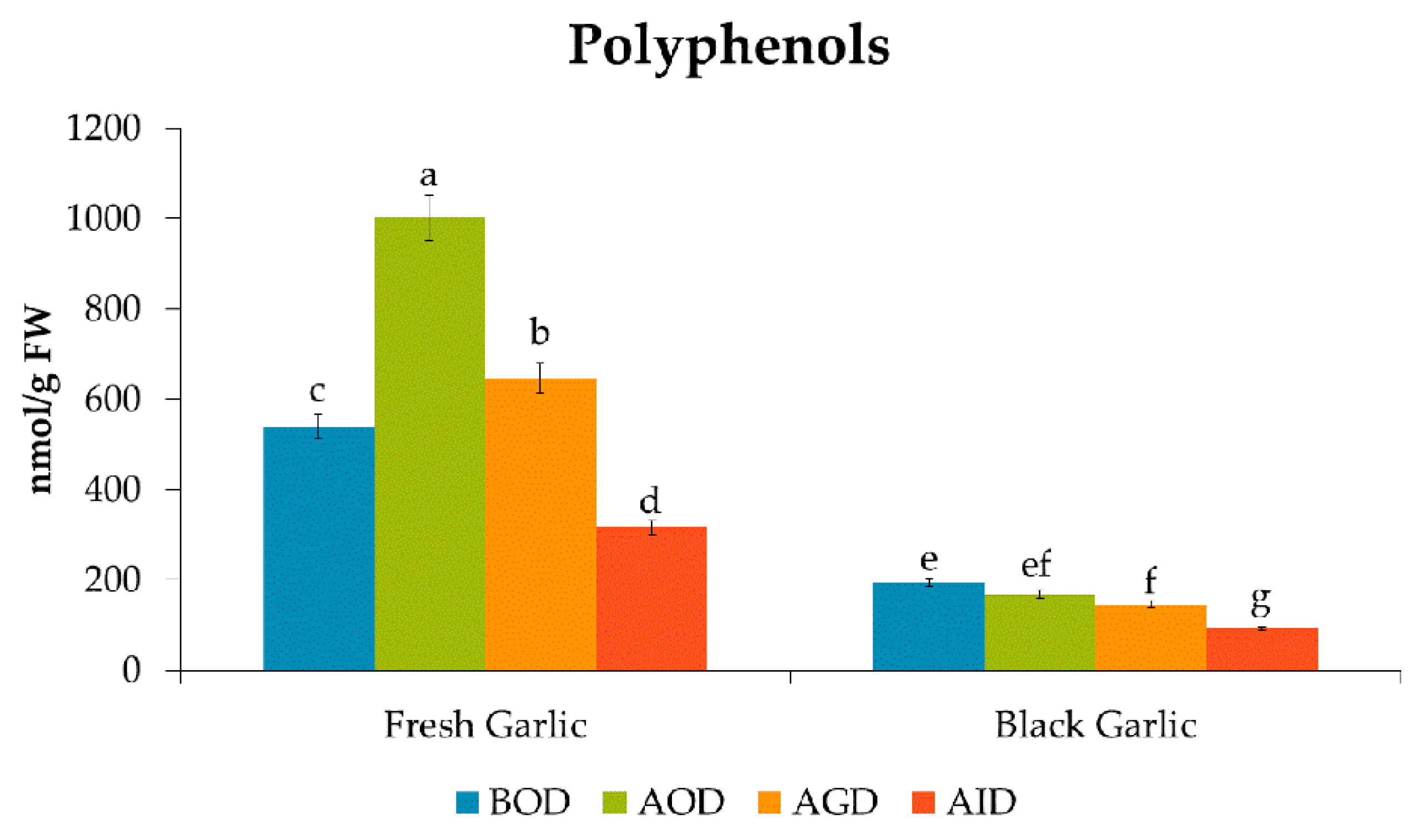
| Total | 539 c | 1001 a | 185.9 | 646 b | 120.0 | 316 d | 58.6 | *** |
| Black Garlic | ||||||||
| Gallic Acid | 14.7 a | 11.2 b | 76.7 | nq | - | nq | - | *** |
| Caffeic Acid | 142 a | 131 ab | 92.3 | 113 b | 79.6 | 92 c | 64.8 | *** |
| Coumaric Acid | 37.1 a | 25.3 c | 68.2 | 33.1 b | 89.4 | nq | - | *** |
| Epigallocatechin gallate | 1.2 a | 0.95 b | 79.4 | nq | - | nq | - | *** |
| Total | 195 a | 168 b | 86.2 | 146 c | 74.9 | 92 d | 47.2 | *** |
| Organosulfur Compounds. | BOD | AOD | % Recovery | AGD | % Recovery | AID | % Recovery-Bioaccessibility | p-Value |
|---|---|---|---|---|---|---|---|---|
| Fresh Garlic | ||||||||
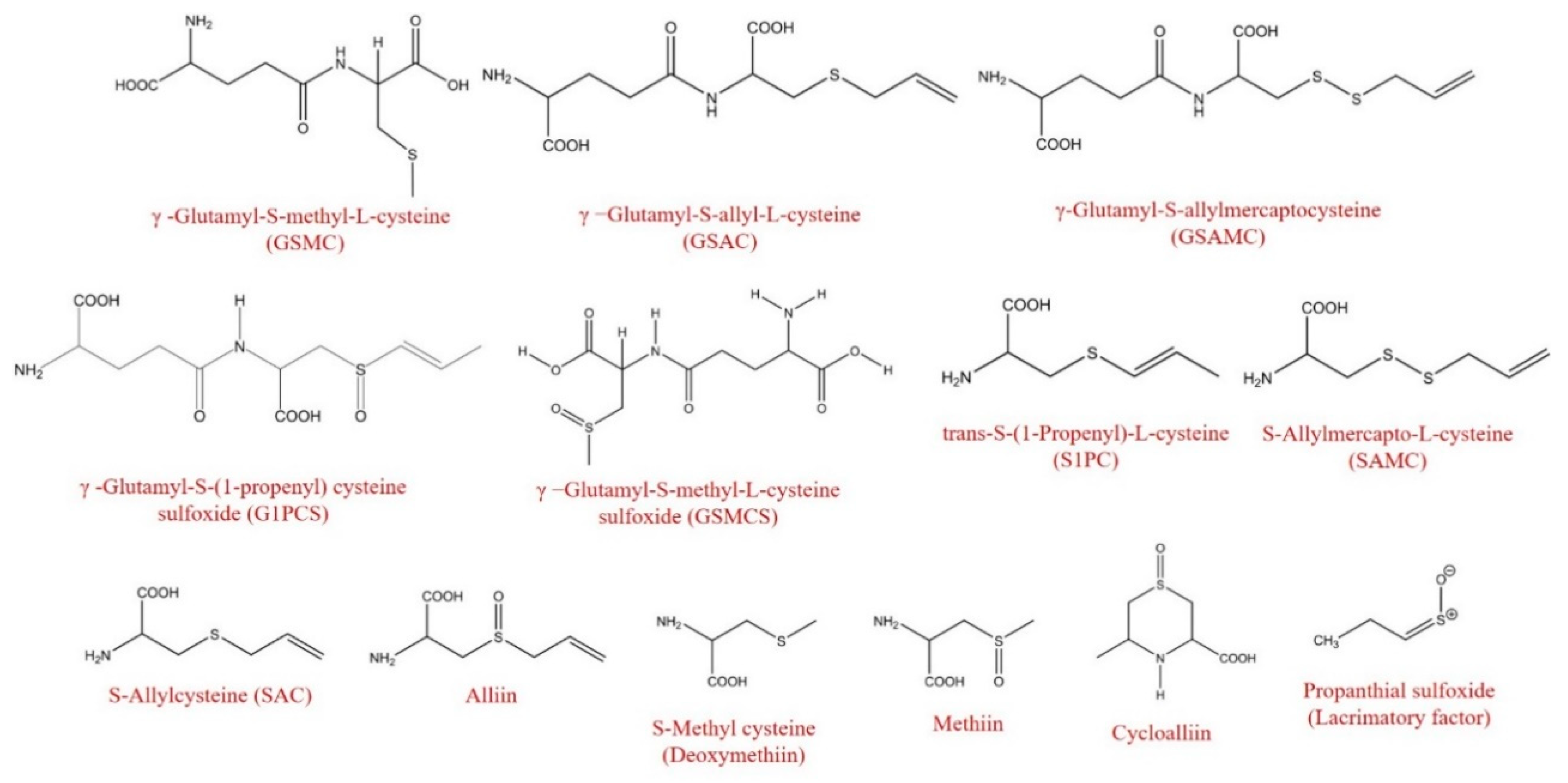
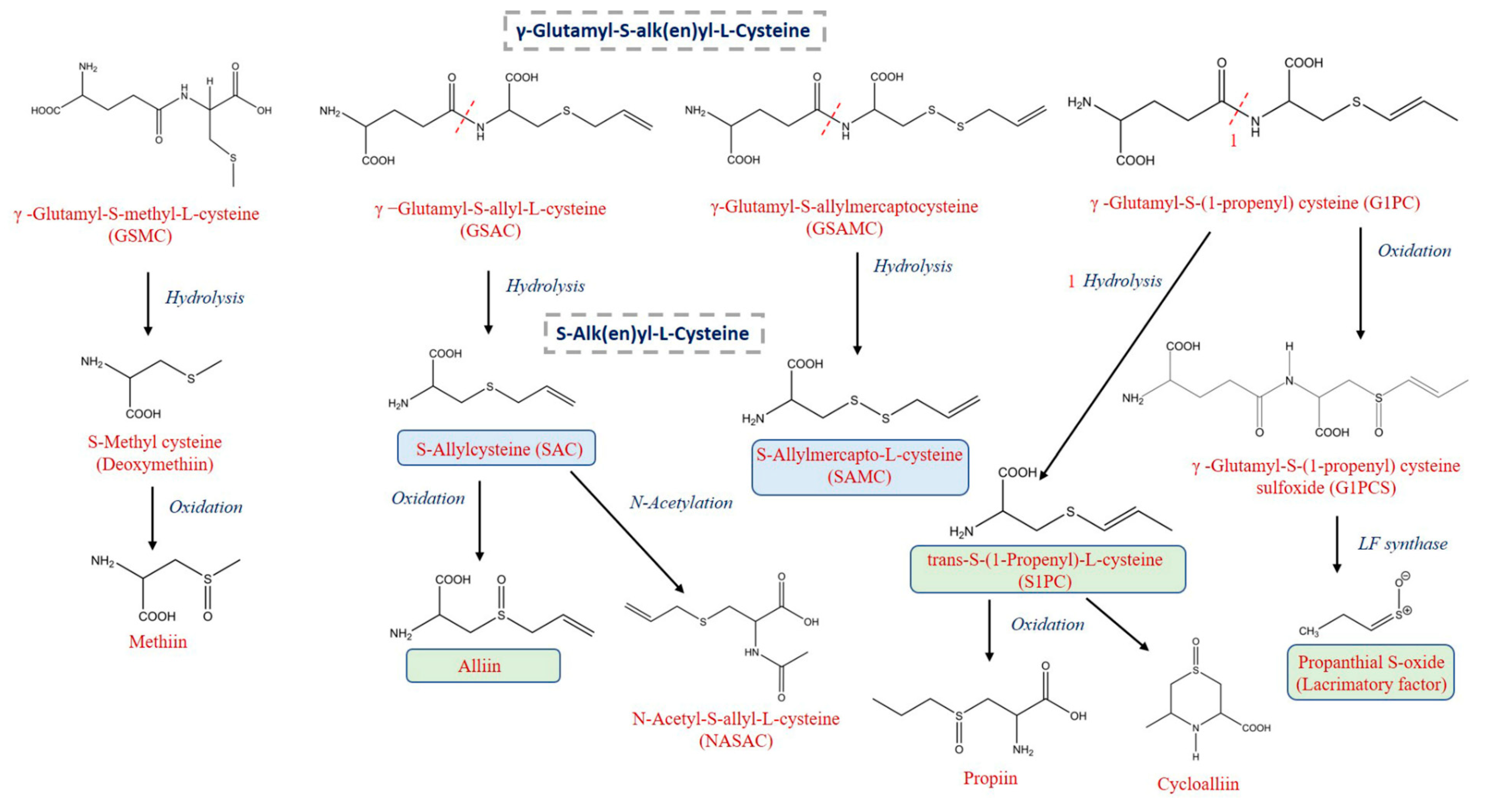
| γ-Glutamyl-S-Alk(en)yl-L-Cysteine Derivatives (GSAk) | ||||||||
| γ-Glutamyl-S-(2-carboxypropyl) cysteine-glycine | 287 a | 253 a | 88.2 | 163 b | 56.9 | 71 c | 24.6 | *** |
| γ-Glutamyl-S-(S-1-propenyl) cysteine-glycine | 159.0 a | 65.6 b | 41.3 | 45.5 c | 28.6 | 11.3 d | 7.1 | *** |
| γ-Glutamyl-S-(S-methyl) cysteine-glycine | 10.3 a | 4.0 b | 39.0 | 2.2 c | 21.3 | 0.8 d | 8.0 | *** |
| γ-Glutamyl-S-methyl-L-cysteine (GSMC) | 1580 a | 1490 a | 94.3 | 899 b | 56.9 | 598 c | 37.9 | *** |
| γ-Glutamyl-S-(propenyl) cysteine (GS1PC) | 6.7 a | 3.5 b | 52.3 | 2.7 b | 40.3 | 1.5 c | 22.8 | *** |
| γ-Glutamyl-S-allyl-L-cysteine (GSAC) | 13,714 a | 8837 b | 64.4 | 5849 c | 42.7 | 3328 d | 24.3 | *** |
| γ-Glutamyl-S-allylmercaptocysteine (GSAMC) | 343 a | 158 b | 46.0 | 97 c | 28.1 | 35 d | 10.2 | *** |
| γ-Glutamyl-cysteine | 36.1 a | 19.3 b | 53.5 | 10.6 c | 29.3 | 4.4 d | 12.2 | *** |
| γ-Glutamyl-S-methyl-L-cysteine sulfoxide (GSMCS) | 17.0 a | 14.5 b | 85.3 | 13.0 b | 76.1 | 13.0 b | 76.6 | *** |
| γ-Glutamyl-S-(1-propenyl) cysteine sulfoxide (G1PCS) | 1311 a | 1142 a | 87.1 | 706 b | 53.8 | 526 b | 40.1 | *** |
| Total GSAk Derivatives | 17,464 a | 11,987 b | 68.6 | 7788 c | 44.6 | 4589 d | 26.3 | *** |
| S-Alk(en)yl-L-Cysteine Derivatives (SACs) | ||||||||
| S-Methylcysteine (Deoxymethiin) | 534 a | 270 b | 50.7 | 118 c | 22.1 | 52 d | 9.8 | *** |
| S-Allylcysteine (SAC) | 1144 a | 1017 a | 88.9 | 384 c | 33.6 | 696 b | 60.8 | *** |
| S-(2-Carboxypropyl) cysteine | 88 a | 50 b | 57.0 | 34 c | 38.4 | 21 d | 24.1 | *** |
| S-allylglutathione (SAG) | 2.7 a | 1.4 b | 52.9 | 0.8 c | 29.2 | 0.4 d | 14.2 | *** |
| trans-S-(1-Propenyl)-L-cysteine (S1PC) | 2909 a | 1868 b | 64.2 | 1216 c | 41.8 | 662 d | 22.8 | *** |
| S-Allylmercapto-L-cysteine (SAMC) | 164 c | 256 b | 156.4 | 296 b | 181.1 | 539 a | 329.3 | *** |
| S-Allylsulfenic acid (Lacrimatory factor) | 1517 a | 887 b | 58.5 | 624 c | 41.2 | 291 d | 19.2 | *** |
| Alliin | 41,706 a | 11,621 b | 27.9 | 5937 c | 14.2 | 2207 c | 5.3 | *** |
| S-Methyl-l-cysteine sulfoxide (Methiin) | 4425 a | 2651 b | 59.9 | 2176 c | 49.2 | 1668 d | 37.7 | *** |
| S-Propyl-L-cysteine sulfoxide (Propiin) | 26.89 a | 3.7 b | 13.7 | 1.9 bc | 7.2 | 0.9 c | 3.2 | *** |
| Cycloalliin | 5251 a | 2741 b | 52.2 | 1855 c | 35.3 | 771 d | 14.7 | *** |
| Methionine sulfoxide | 14.5 a | 8.4 c | 57.9 | 8.7 c | 60.0 | 10.5 b | 72.4 | *** |
| N-Acetyl-S-allyl-L-cysteine (NASAC) | 60.3 b | 69.1 a | 114.7 | 37.5 c | 62.2 | 10.4 d | 17.2 | *** |
| Total SACs Derivatives | 57,842 a | 21,444 | 37.1 | 12,689 c | 22.0 | 6929 d | 12.0 | *** |
| Total OS Compounds | 75,306 a | 33,431 b | 44.4 | 20,477 c | 27.2 | 11,518 d | 15.3 | *** |
| Black Garlic | ||||||||
| γ-Glutamyl-S-Alk(en)yl-L-Cysteine (GSAk) | ||||||||
| γ-Glutamyl-S-(S-1-propenyl) cysteine-glycine | 119 a | 120 a | 100.9 | 120 a | 101.1 | nd | - | *** |
| γ-Glutamyl-S-methyl-L-cysteine (GSMC) | 199 a | 206 a | 103.7 | 200 a | 100.7 | nd | - | *** |
| γ-Glutamyl-S-allyl-L-cysteine (GSAC) | 4393 a | 3900 ab | 88.8 | 3597 b | 81.9 | 934 c | 21.3 | *** |
| γ-Glutamyl-S-allylmercaptocysteine (GSAMC) | 504 a | 510 a | 101.2 | 560 a | 111.3 | 287 b | 57.1 | *** |
| γ-Glutamyl-S-methyl-L-cysteine sulfoxide (GSMCS) | 380 | 384 | 101.1 | 340 | 89.4 | 338 | 88.9 | ns |
| γ-Glutamyl-S-(1-propenyl) cysteine sulfoxide (G1PCS) | 3452 a | 3346 a | 96.9 | 2645 b | 76.6 | 2305 b | 66.8 | *** |
| Total GSAk Derivatives | 9046 a | 8466 a | 93.6 | 7463 b | 82.5 | 3864 c | 42.7 | *** |
| S-Alk(en)yl-L-Cysteine (SACs) | ||||||||
| S-methyl cysteine (Deoxymethiin) | 168 b | 166 b | 98.7 | 218 a | 129.8 | nd | - | *** |
| S-Allyl-L-cysteine (SAC) | 7683 a | 6189 b | 80.5 | 4066 c | 52.9 | 2803 d | 36.5 | *** |
| S-(2-carboxypropyl) cysteine | 115 b | 116 b | 100.7 | 149 a | 130.3 | nd | - | *** |
| S-allylmercapto-L-cysteine (SAMC) | 408 ab | 358 b | 87.7 | 224 c | 54.9 | 434 a | 106.4 | *** |
| S-allylglutathione | 46.6 a | 43.6 a | 93.6 | nd | - | nd | - | |
| trans-S-(1-Propenyl)-L-cysteine (S1PC) | 205 c | 198 c | 96.6 | 243 b | 118.5 | 310 a | 151.2 | *** |
| S-allylsulfenic acid (Lacrimatory factor) | 935 a | 818 ab | 87.5 | 648 b | 69.4 | 895 a | 95.7 | ** |
| S-(S-propyl) cysteine | 229 b | 355 a | 155.3 | nd | - | nd | - | *** |
| Alliin | 5611 a | 4735 ab | 84.4 | 4423 b | 78.8 | 4302 b | 76.7 | ** |
| S-methyl-cysteine sulfoxide (Methiin) | 243 a | 255 a | 104.9 | 235 a | 96.6 | nd | - | *** |
| Methionine sulfoxide | 502 b | 535 b | 106.6 | 411 c | 81.9 | 1314 a | 262.0 | *** |
| Total SACs Derivatives | 16,145 a | 13,768 b | 85.3 | 10,617 c | 65.8 | 10,059 c | 62.3 | *** |
| Total OS Compounds | 25,191 a | 22,234 b | 88.3 | 18,080 c | 71.8 | 13,923 d | 55.3 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Ortega, A.; Pereira-Caro, G.; Ordóñez, J.L.; Moreno-Rojas, R.; Ortíz-Somovilla, V.; Moreno-Rojas, J.M. Bioaccessibility of Bioactive Compounds of ‘Fresh Garlic’ and ‘Black Garlic’ through In Vitro Gastrointestinal Digestion. Foods 2020, 9, 1582. https://doi.org/10.3390/foods9111582
Moreno-Ortega A, Pereira-Caro G, Ordóñez JL, Moreno-Rojas R, Ortíz-Somovilla V, Moreno-Rojas JM. Bioaccessibility of Bioactive Compounds of ‘Fresh Garlic’ and ‘Black Garlic’ through In Vitro Gastrointestinal Digestion. Foods. 2020; 9(11):1582. https://doi.org/10.3390/foods9111582
Chicago/Turabian StyleMoreno-Ortega, Alicia, Gema Pereira-Caro, José Luis Ordóñez, Rafael Moreno-Rojas, Víctor Ortíz-Somovilla, and José Manuel Moreno-Rojas. 2020. "Bioaccessibility of Bioactive Compounds of ‘Fresh Garlic’ and ‘Black Garlic’ through In Vitro Gastrointestinal Digestion" Foods 9, no. 11: 1582. https://doi.org/10.3390/foods9111582