

The Characterization of Subcutaneous Adipose Tissue in Sunit Sheep at Different Growth Stages: A Comprehensive Analysis of the Morphology, Fatty Acid Profile, and Metabolite Profile
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
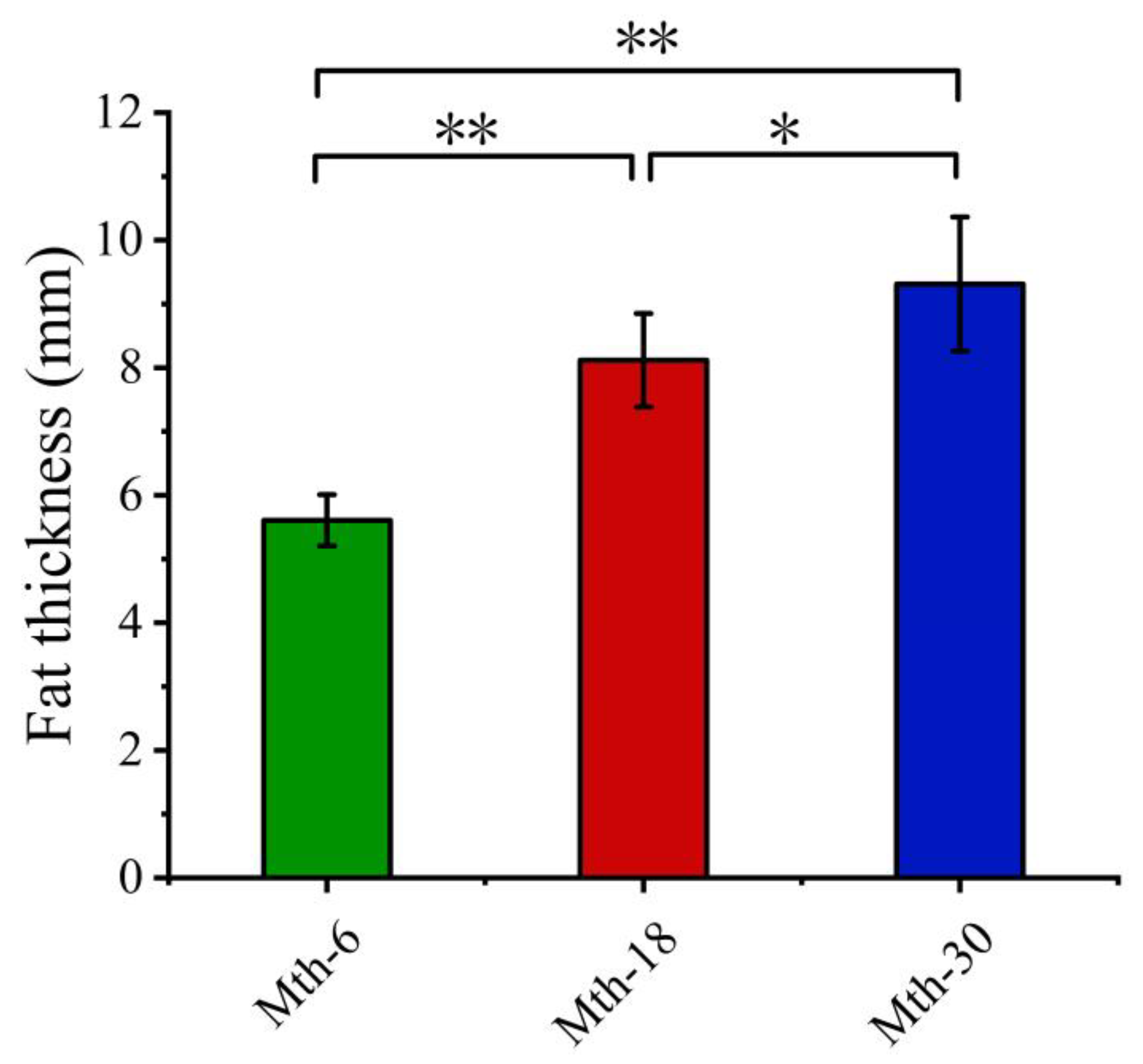
2.2. Fat Thickness Measurement
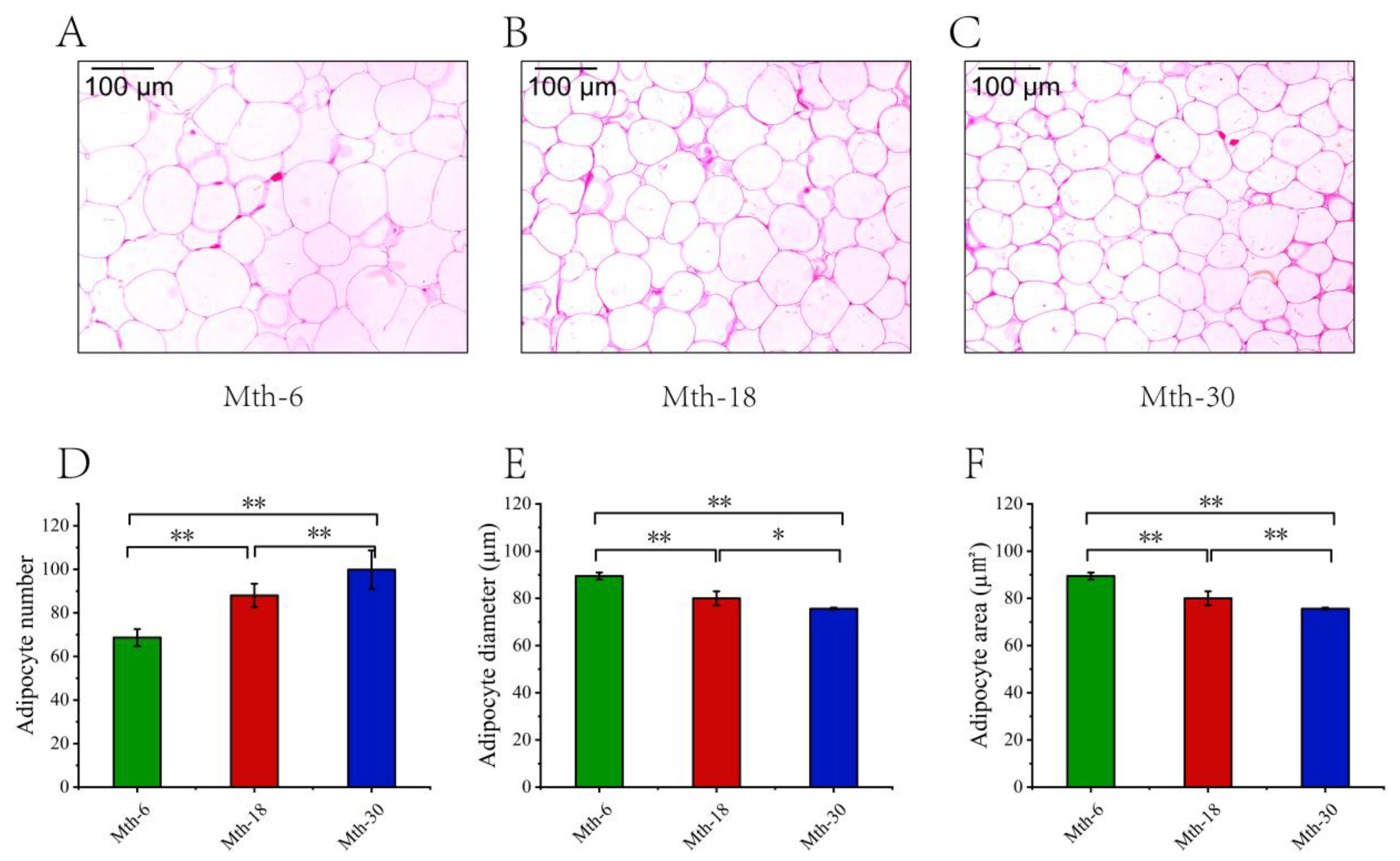
2.3. Histological Analysis
2.4. Extraction and Analysis of Fatty Acids
2.5. Extraction and Analysis of Metabolites
2.6. Statistical Analysis
3. Results
3.1. Fat Thickness and Adipocyte Morphology Analysis
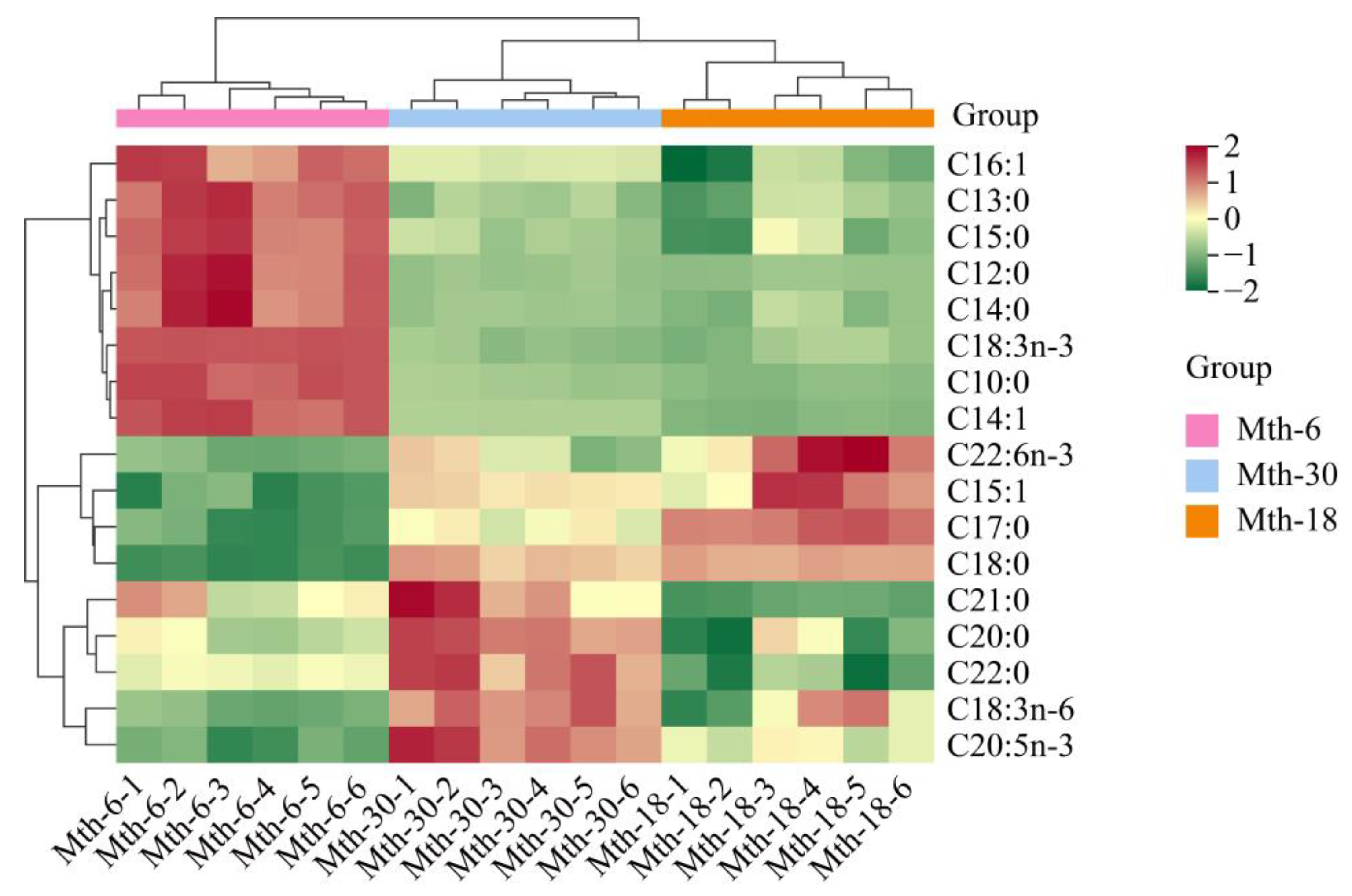
3.2. Fatty Acid Profile Analysis
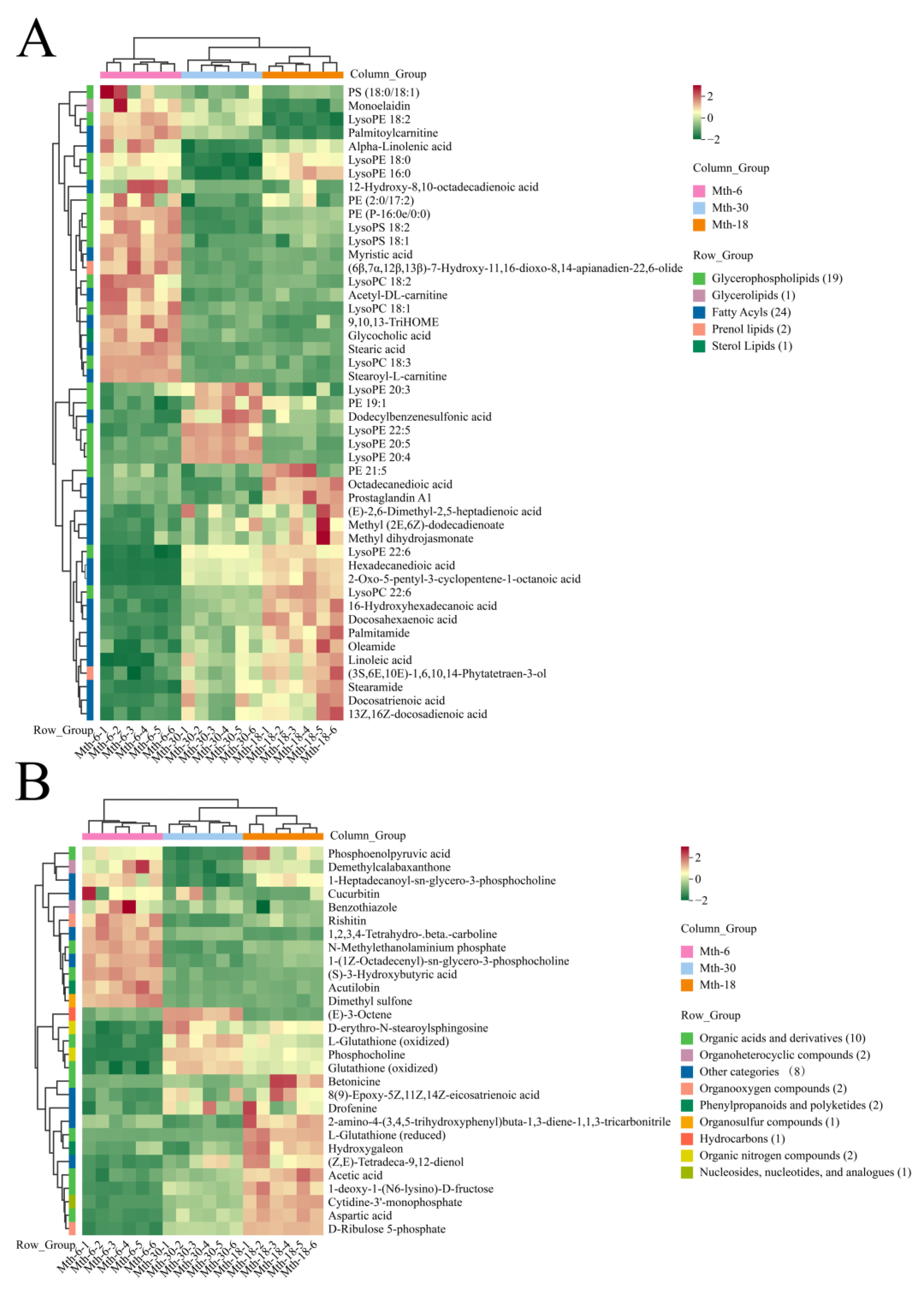
3.3. Metabolite Profile Analysis
3.3.1. Summary of Metabolite Profile Analysis
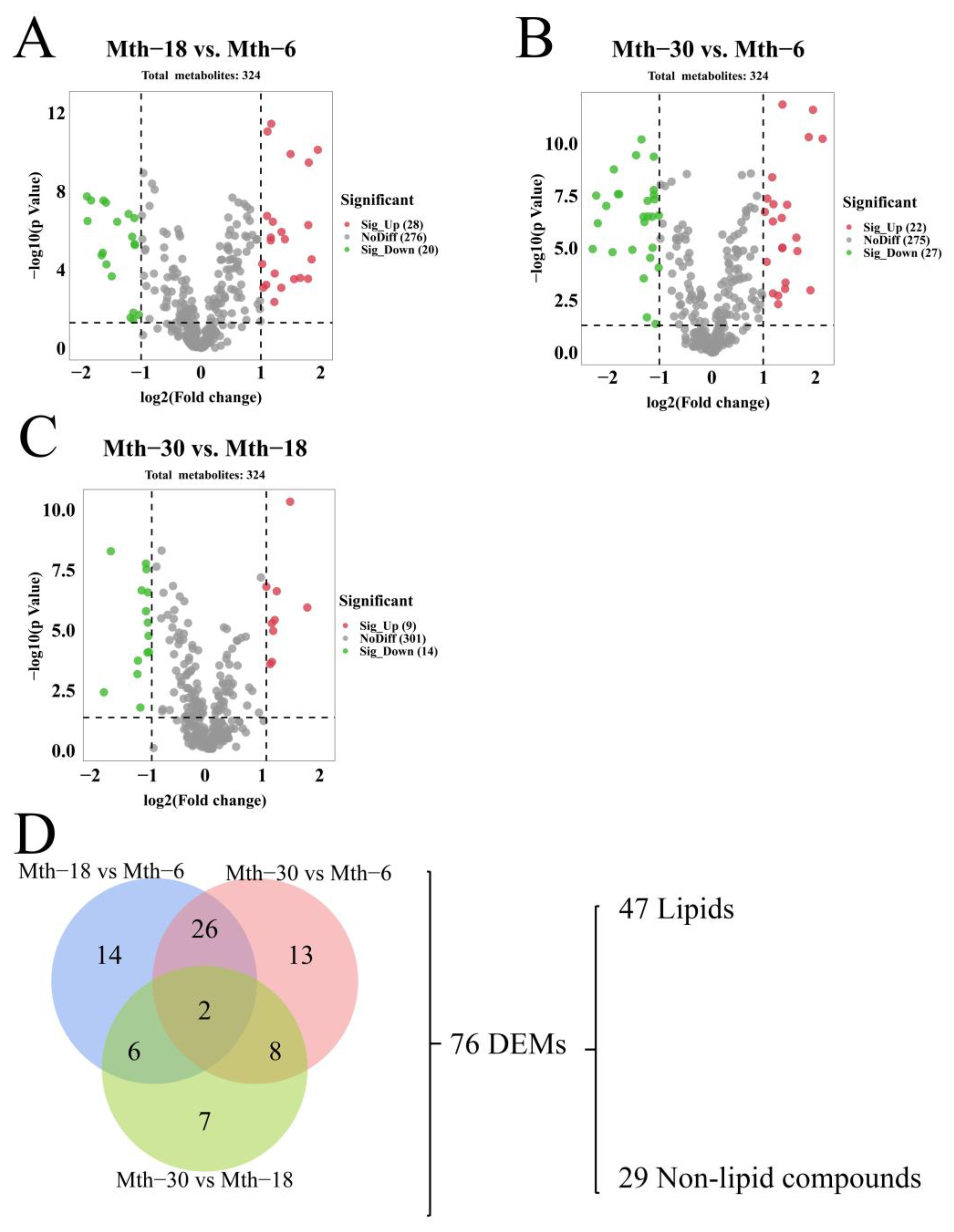
3.3.2. Differential Expression Analysis
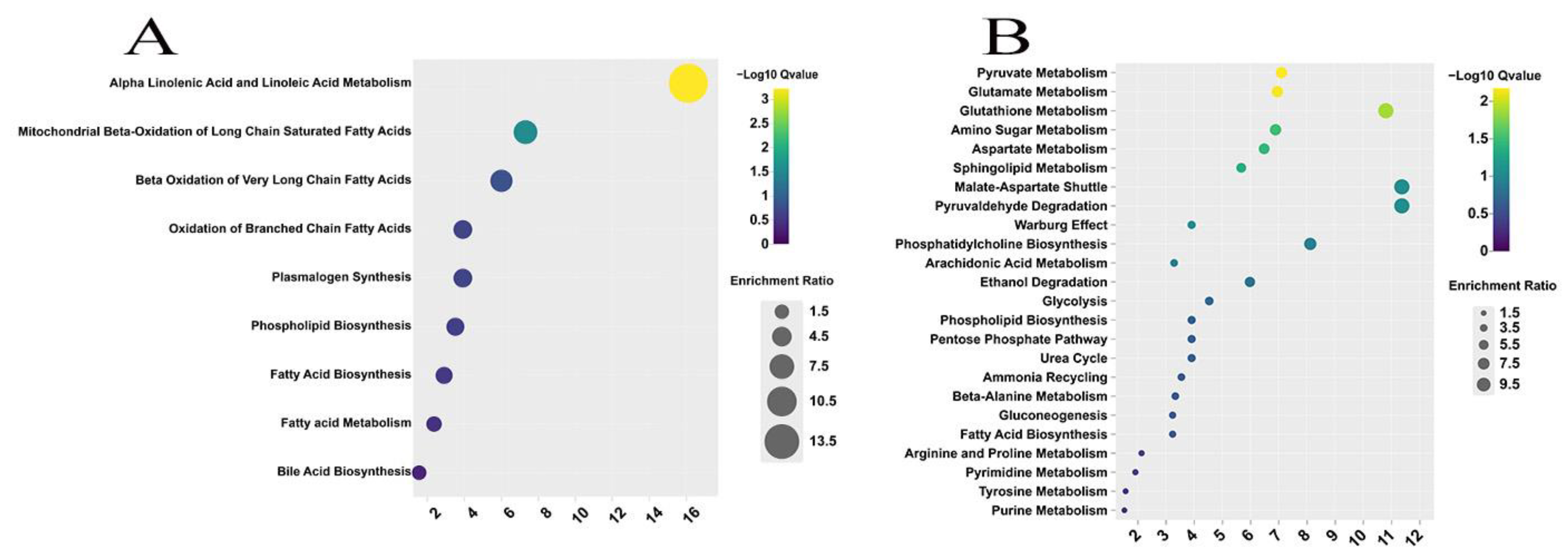
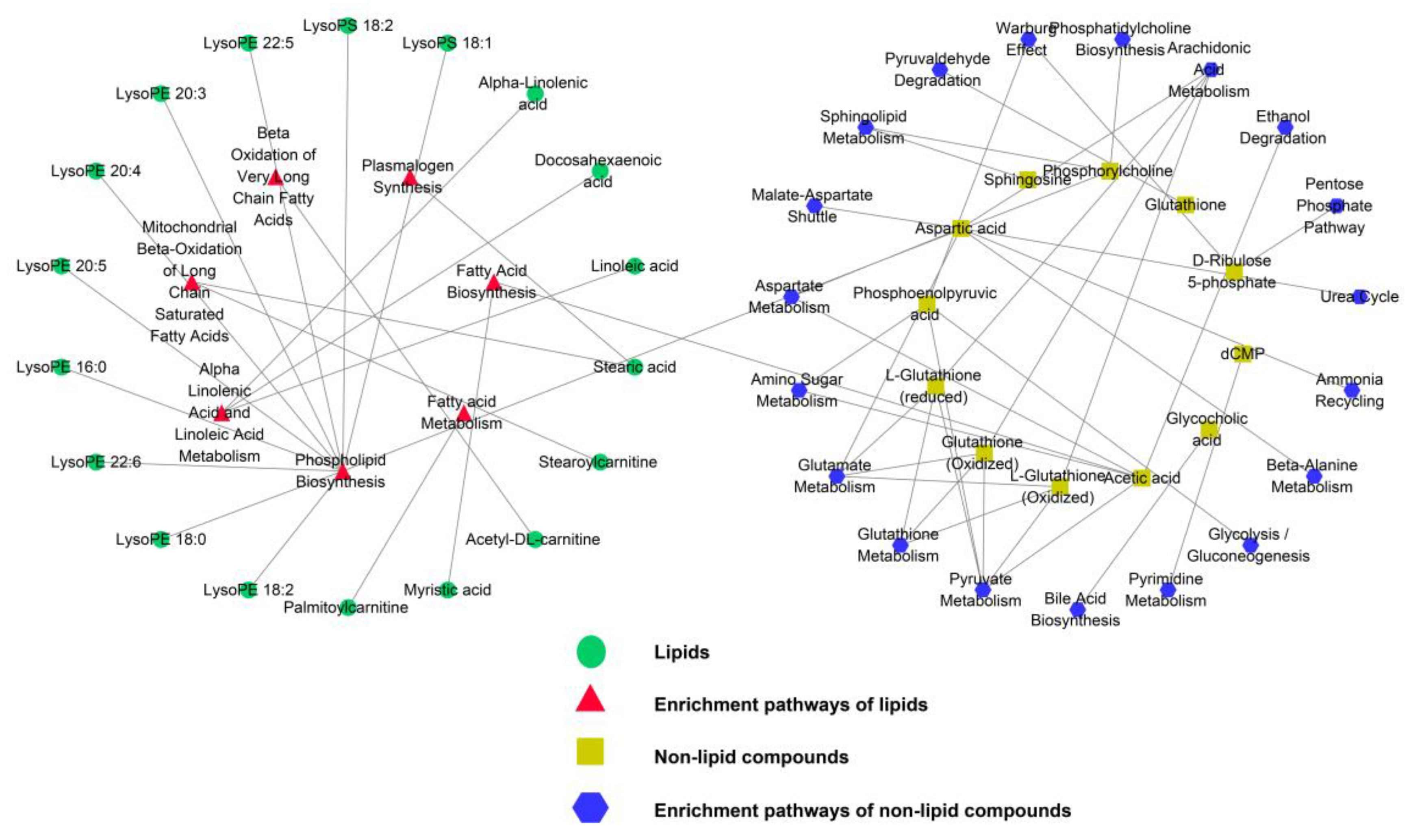
3.3.3. Enrichment Pathway Analysis of Differentially Expressed Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deng, K.; Ren, C.; Liu, Z.; Gao, X.; Fan, Y.; Zhang, G.; Zhang, Y.; Ma, E.S.; Wang, F.; You, P. Characterization of RUNX1T1, an Adipogenesis Regulator in Ovine Preadipocyte Differentiation. Int. J. Mol. Sci. 2018, 19, 1300. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wu, R.; Yun, Y.; Qin, X.; Huang, Y.; Chen, L.; Han, Y.; Wu, J.; Sha, L.; Borjigin, G. MicroRNA and circular RNA profiling in the deposited fat tissue of Sunite sheep. Front. Vet. Sci. 2022, 9, 954882. [Google Scholar] [CrossRef] [PubMed]
- Junior, F.F.; Pena, A.D.; Grandis, F.A.; Koritiaki, N.A.; Rego, F.C.D.; dos Santos, R.M.; Ribeiro, E.L.D. Subcutaneous fat thickness at slaughter in castrated and non-castrated Santa Ines and Dorper lambs and its influence on meat and carcass quality. Livest. Sci. 2021, 253, 104694. [Google Scholar] [CrossRef]
- Song, S.Z.; Wu, J.P.; Zhao, S.G.; Casper, D.P.; He, B.; Liu, T. The effect of energy restriction on fatty acid profiles of longissimus dorsi and tissue adipose depots in sheep1. J. Anim. Sci. 2017, 95, 3940–3948. [Google Scholar] [CrossRef] [PubMed]
- Ugnivenko, A.; Kruk, O.; Nosevych, D.; Antoniuk, T.; Kryzhova, Y.; Gruntovskyi, M.; Prokopenko, N.; Yemtcev, V.; Kharsika, I.; Nesterenko, N. The expressiveness of meat forms of cattle depending on the content of adipose tissue under the skin and between the muscles. Potravin. Slovak J. Food Sci. 2023, 17, 358–370. [Google Scholar] [CrossRef]
- Silva-Vignato, B.; Coutinho, L.L.; Poleti, M.D.; Cesar, A.S.M.; Moncau, C.T.; Regitano, L.C.A.; Balieiro, J.C.C. Gene co-expression networks associated with carcass traits reveal new pathways for muscle and fat deposition in Nelore cattle. BMC Genom. 2019, 20, 32. [Google Scholar] [CrossRef]
- Xu, Y.X.; Wang, B.; Jing, J.N.; Ma, R.; Luo, Y.H.; Li, X.; Yan, Z.; Liu, Y.J.; Gao, L.; Ren, Y.L.; et al. Whole-body adipose tissue multi-omic analyses in sheep reveal molecular mechanisms underlying local adaptation to extreme environments. Commun. Biol. 2023, 6, 159. [Google Scholar] [CrossRef]
- He, C.; Wang, Y.; Xu, Q.; Xiong, Y.; Zhu, J.; Lin, Y. Overexpression of Krueppel like factor 3 promotes subcutaneous adipocytes differentiation in goat Capra hircus. Anim. Sci. J. 2021, 92, e13514. [Google Scholar] [CrossRef]
- Joffin, N.; Paschoal, V.A.; Gliniak, C.M.; Crewe, C.; Scherer, P.E. Mitochondrial metabolism is a key regulator of the fibro-inflammatory and adipogenic stromal subpopulations in white adipose tissue. Cell Stem. Cell 2021, 28, 702–717. [Google Scholar] [CrossRef]
- Du, L.; Chang, T.; An, B.; Liang, M.; Deng, T.; Li, K.; Cao, S.; Du, Y.; Gao, X.; Xu, L.; et al. Transcriptomics and Lipid Metabolomics Analysis of Subcutaneous, Visceral, and Abdominal Adipose Tissues of Beef Cattle. Genes 2022, 14, 37. [Google Scholar] [CrossRef]
- Xiong, L.; Pei, J.; Bao, P.; Wang, X.; Guo, S.; Cao, M.; Kang, Y.; Yan, P.; Guo, X. The Effect of the Feeding System on Fat Deposition in Yak Subcutaneous Fat. Int. J. Mol. Sci. 2023, 24, 7381. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Pei, J.; Wu, X.; Kalwar, Q.; Yan, P.; Guo, X. Effect of Gender to Fat Deposition in Yaks Based on Transcriptomic and Metabolomics Analysis. Front. Cell Dev. Biol. 2021, 9, 653188. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Liufu, S.; Liu, X.; Ai, N.; Xu, X.; Li, X.; Yu, Z.; Yin, Y.; Liu, M.; Ma, H. Comprehensive analysis of transcriptomic and metabolomic profiles uncovered the age-induced dynamic development pattern of subcutaneous fat in Ningxiang pig. Gene 2023, 880, 147624. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, D.; Zhang, J.; He, M.; Li, Y.; Liu, R.; Li, M. Transcriptomic and lipidomic profiling of subcutaneous and visceral adipose tissues in 15 vertebrates. Sci. Data 2023, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wu, R.; Yun, Y.; Qin, X.; Chen, L.; Han, Y.; Wu, J.; Sha, L.; Borjigin, G. Transcriptome analysis of messenger RNA and long noncoding RNA related to different developmental stages of tail adipose tissues of sunite sheep. Food Sci. Nutr. 2021, 9, 5722–5734. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, R.; Ying, Y.; Zhang, Y.; Huang, Y.; Wu, H.; Lin, K. Non-genetic factors affecting the meat quality and flavor of Inner Mongolian lambs: A review. Front. Vet. Sci. 2022, 9, 1067880. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Wang, Z.; Zhao, M.; Wang, Y.; Zhang, J.; Li, Y.; Gegen, T.; Jia, Y. Study on quality characteristics of natural pastures in different steppe types. Feed. Res. 2022, 4, 75–78. [Google Scholar]
- Zhang, X.; Han, L.; Hou, S.; Raza, S.H.A.; Gui, L.; Sun, S.; Wang, Z.; Yang, B.; Yuan, Z.; Simal-Gandara, J.; et al. Metabolomics approach reveals high energy diet improves the quality and enhances the flavor of black Tibetan sheep meat by altering the composition of rumen microbiota. Front. Nutr. 2022, 9, 915558. [Google Scholar] [CrossRef]
- Zheng, Y.; He, J.; Yang, D.; Yuan, M.; Liu, S.; Dai, F.; Jia, Y.; Cheng, Y. Irisin reduces the abnormal reproductive and metabolic phenotypes of PCOS by regulating the activity of brown adipose tissue in micedagger. Biol. Reprod. 2022, 107, 1046–1058. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1959, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fang, J.; Qi, X.; Lin, M.; Zhong, Y.; Sun, L.; Cui, W. Combined Analysis of the Fruit Metabolome and Transcriptome Reveals Candidate Genes Involved in Flavonoid Biosynthesis in Actinidia arguta. Int. J. Mol. Sci. 2018, 19, 1471. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liang, C.; Manthari, R.K.; Yu, Y.; Zhang, J.; Wang, J.; Cao, J. Distribution characteristics and regulation of amino acids and fatty acids in muscle and adipose tissues of sheep grown in natural grazing environment. Anim. Sci. J. 2022, 93, e13769. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Zhou, J.; Degen, A.; Wang, W.; Guo, Y.; Kang, J.; Liu, P.; Ding, L.; Shang, Z.; Qiu, Q. Comparison between Tibetan and Small-tailed Han sheep in adipocyte phenotype, lipid metabolism and energy homoeostasis regulation of adipose tissues when consuming diets of different energy levels. Br. J. Nutr. 2020, 124, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Jiao, D.; Ji, K.; Liu, H.; Wang, W.; Wu, X.; Zhou, J.; Zhang, Y.; Zhou, H.; Hickford, J.G.H.; Degen, A.A.; et al. Transcriptome Analysis Reveals Genes Involved in Thermogenesis in Two Cold-Exposed Sheep Breeds. Genes 2021, 12, 375. [Google Scholar] [CrossRef] [PubMed]
- Fuller-Jackson, J.P.; Clarke, I.J.; Rao, A.; Henry, B.A. Exercise counteracts the homeostatic decrease in thermogenesis caused by caloric restriction in sheep. FASEB J. 2018, 32, 3859–3869. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef]
- Li, A.; Li, Y.Y.; Wuqie, Q.B.; Li, X.; Zhang, H.; Wang, Y.; Wang, Y.L.; Zhu, J.J.; Lin, Y.Q. Effect of ACADL on the differentiation of goat subcutaneous adipocyte. Anim. Biosci. 2023, 36, 829–839. [Google Scholar] [CrossRef]
- Murariu, O.C.; Murariu, F.; Frunză, G.; Ciobanu, M.M.; Boișteanu, P.C. Fatty Acid Indices and the Nutritional Properties of Karakul Sheep Meat. Nutrients 2023, 15, 1061. [Google Scholar] [CrossRef]
- Vitor, A.C.M.; Godinho, M.; Francisco, A.E.; Silva, J.; Almeida, J.; Fialho, L.; Soldado, D.; Jeronimo, E.; Scollan, N.D.; Huws, S.A.; et al. Nannochloropsis oceanica microalga feeding increases long-chain omega-3 polyunsaturated fatty acids in lamb meat. Meat Sci. 2023, 197, 109053. [Google Scholar] [CrossRef]
- Wang, B.; Yang, L.; Luo, Y.; Su, R.; Lin, S.; Zhao, L.; Jin, Y. Effects of feeding regimens on meat quality, fatty acid composition and metabolism as related to gene expression in Chinese Sunit sheep. Small Rumin. Res. 2018, 169, 127–133. [Google Scholar] [CrossRef]
- Mapiye, C.; Dugan, M.E.; Juarez, M.; Basarab, J.A.; Baron, V.S.; Turner, T.; Yang, X.; Aldai, N.; Aalhus, J.L. Influence of alpha-tocopherol supplementation on trans-18:1 and conjugated linoleic acid profiles in beef from steers fed a barley-based diet. Animal 2012, 6, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, T.; Anzani, C.; Ferri, M.; Marzocchi, S.; Caboni, M.F.; Monari, S.; Tassoni, A. Enzymatic Digestion of Calf Fleshing Meat By-Products: Antioxidant and Anti-Tyrosinase Activity of Protein Hydrolysates, and Identification of Fatty Acids. Foods 2021, 10, 755. [Google Scholar] [CrossRef] [PubMed]
- Lanzarini, F.; Pereira, F.A.; Camargo, J.D.; Oliveira, A.M.; Belaz, K.R.A.; Melendez-Perez, J.J.; Eberlin, M.N.; Brum, M.C.S.; Mesquita, F.S.; Sudano, M.J. ELOVL5 Participates in Embryonic Lipid Determination of Cellular Membranes and Cytoplasmic Droplets. Int. J. Mol Sci. 2021, 22, 1311. [Google Scholar] [CrossRef] [PubMed]
- Zhan, P.; Tian, H.L.; Sun, B.G.; Zhang, Y.; Chen, H. Quality Control of Mutton by Using Volatile Compound Fingerprinting Techniques and Chemometric Methods. J. Food Qual. 2017, 2017, 1–8. [Google Scholar] [CrossRef]
- Watkins, P.J.; Frank, D. Heptadecanoic acid as an indicator of BCFA content in sheep fat. Meat Sci. 2019, 151, 33–35. [Google Scholar] [CrossRef] [PubMed]
- Heier, C.; Knittelfelder, O.; Hofbauer, H.F.; Mende, W.; Pornbacher, I.; Schiller, L.; Schoiswohl, G.; Xie, H.; Gronke, S.; Shevchenko, A.; et al. Hormone-sensitive lipase couples intergenerational sterol metabolism to reproductive success. Elife 2021, 10, e63252. [Google Scholar] [CrossRef]
- Ma, W.P.; Yin, S.N.; Chen, J.P.; Geng, X.C.; Liu, M.F.; Li, H.H.; Liu, M.; Liu, H.B. Stimulating the Hematopoietic Effect of Simulated Digestive Product of Fucoidan from Sargassum fusiforme on Cyclophosphamide-Induced Hematopoietic Damage in Mice and Its Protective Mechanisms Based on Serum Lipidomics. Mar. Drugs 2022, 20, 201. [Google Scholar] [CrossRef]
- Cheng, K.; Liu, T.; Yang, C.; Yang, H.; Liu, D. Relationship between phospholipid molecules species and volatile compounds in grilled lambs during the heating process. Food Chem. X 2024, 21, 1–11. [Google Scholar] [CrossRef]
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef]
- Robichaud, P.P.; Poirier, S.J.; Boudreau, L.H.; Doiron, J.A.; Barnett, D.A.; Boilard, E.; Surette, M.E. On the cellular metabolism of the click chemistry probe 19-alkyne arachidonic acid. J. Lipid Res. 2016, 57, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bi, C.; Pang, W.; Liu, Y.; Yuan, Y.; Zhao, H.; Zhang, T.; Zhao, Y.; Li, Y. Plasma Metabolic Profiling Analysis of Gout Party on Acute Gout Arthritis Rats Based on UHPLC-Q-TOF/MS Combined with Multivariate Statistical Analysis. Int. J. Mol. Sci. 2019, 20, 5753. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Sun, Z.; Yu, Z.; Li, H.; Luo, H.; Wang, B. Transcriptome and targeted metabolome analysis provide insights into bile acids’ new roles and mechanisms on fat deposition and meat quality in lamb. Food Res. Int. 2022, 162, 111941. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Huang, S.; Yang, N.; Cao, A.; Zhao, L.; Zhang, J.; Zhao, G.; Ma, Q. Porcine bile acids promote the utilization of fat and vitamin A under low-fat diets. Front. Nutr. 2022, 9, 1005195. [Google Scholar] [CrossRef] [PubMed]
- Cheema, N.; Herbst, A.; McKenzie, D.; Aiken, J.M. Apoptosis and necrosis mediate skeletal muscle fiber loss in age-induced mitochondrial enzymatic abnormalities. Aging Cell 2015, 14, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Dolly, A.; Lecomte, T.; Tabchouri, N.; Caulet, M.; Michot, N.; Anon, B.; Chautard, R.; Desvignes, Y.; Ouaissi, M.; Fromont-Hankard, G.; et al. Pectoralis major muscle atrophy is associated with mitochondrial energy wasting in cachectic patients with gastrointestinal cancer. J. Cachexia Sarcopenia Muscle 2022, 13, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Machiela, E.; Liontis, T.; Dues, D.J.; Rudich, P.D.; Traa, A.; Wyman, L.; Kaufman, C.; Cooper, J.F.; Lew, L.; Nadarajan, S.; et al. Disruption of mitochondrial dynamics increases stress resistance through activation of multiple stress response pathways. FASEB J. 2020, 34, 8475–8492. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.S.; Zheng, Z.; Yang, C.R.; Zhang, X.R.; Cheng, L.; Su, G.H.; Bai, C.L.; Li, G.P. Comparative Transcriptome Analysis Provides Insights into the Polyunsaturated Fatty Acid Synthesis Regulation of Fat-1 Transgenic Sheep. Int. J. Mol. Sci. 2020, 21, 1121. [Google Scholar] [CrossRef]
- Xu, Y.; Han, J.; Dong, J.; Fan, X.; Cai, Y.; Li, J.; Wang, T.; Zhou, J.; Shang, J. Metabolomics Characterizes the Effects and Mechanisms of Quercetin in Nonalcoholic Fatty Liver Disease Development. Int. J. Mol. Sci. 2019, 20, 1220. [Google Scholar] [CrossRef]
- Shu, T.; Song, X.; Cui, X.; Ge, W.; Gao, R.; Wang, J. Fc Gamma Receptor IIb Expressed in Hepatocytes Promotes Lipid Accumulation and Gluconeogenesis. Int. J. Mol. Sci. 2018, 19, 2932. [Google Scholar] [CrossRef]
- Horikoshi, Y.; Yan, Y.; Terashvili, M.; Wells, C.; Horikoshi, H.; Fujita, S.; Bosnjak, Z.J.; Bai, X. Fatty Acid-Treated Induced Pluripotent Stem Cell-Derived Human Cardiomyocytes Exhibit Adult Cardiomyocyte-Like Energy Metabolism Phenotypes. Cells 2019, 8, 1095. [Google Scholar] [CrossRef] [PubMed]
- Marcocci, L.; Suzuki, Y.J. Metabolomics Studies to Assess Biological Functions of Vitamin E Nicotinate. Antioxidants 2019, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Terrell, J.A.; Chung, J.H.; Chen, C. Electrospun Microfibers Modulate Intracellular Amino Acids in Liver Cells via Integrin β1. Bioengineering 2021, 8, 88. [Google Scholar] [CrossRef] [PubMed]
- Jersin, R.Å.; Sri Priyanka Tallapragada, D.; Skartveit, L.; Bjune, M.S.; Muniandy, M.; Lee-Ødegård, S.; Heinonen, S.; Alvarez, M.; Birkeland, K.I.; André Drevon, C.; et al. Impaired Adipocyte SLC7A10 Promotes Lipid Storage in Association with Insulin Resistance and Altered BCAA Metabolism. J. Clin. Endocrinol. Metab. 2023, 108, 2217–2229. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.M.; Liu, G.Y.; Wang, H.L.; Wassie, T.; Wu, X. Maternal pyrimidine nucleoside supplementation regulates fatty acid, amino acid and glucose metabolism of neonatal piglets. Anim Nutr. 2022, 11, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Huertas, E.; Alcaide-Hidalgo, J.M. Characterisation of Endogenous Peptides Present in Virgin Olive Oil. Int. J. Mol. Sci. 2022, 23, 1712. [Google Scholar] [CrossRef] [PubMed]
- Asantewaa, G.; Tuttle, E.T.; Ward, N.P.; Kang, Y.P.; Kim, Y.; Kavanagh, M.E.; Girnius, N.; Chen, Y.; Duncan, R.; Rodriguez, K.; et al. Glutathione supports lipid abundance in vivo. bioRxiv 2023. bioRxiv:2023.02.10.524960. [Google Scholar]
- Oates, E.H.; Antoniewicz, M.R. Coordinated reprogramming of metabolism and cell function in adipocytes from proliferation to differentiation. Metab. Eng. 2022, 69, 221–230. [Google Scholar]
- Garcia-Galicia, I.A.; Arras-Acosta, J.A.; Huerta-Jimenez, M.; Renteria-Monterrubio, A.L.; Loya-Olguin, J.L.; Carrillo-Lopez, L.M.; Tirado-Gallegos, J.M.; Alarcon-Rojo, A.D. Natural Oregano Essential Oil May Replace Antibiotics in Lamb Diets: Effects on Meat Quality. Antibiotics 2020, 9, 248. [Google Scholar] [CrossRef]
- Feng, X.; Li, M.H.; Xia, J.; Deng Ba, D.J.; Ruan, L.Y.; Xing, Y.X.; Chen, C.; Wang, J.S.; Zhong, G.J. Tibetan Medical Formula Shi-Wei-Gan-Ning-Pill Protects Against Carbon Tetrachloride-Induced Liver Fibrosis—An NMR-Based Metabolic Profiling. Front. Pharmacol. 2018, 9, 965. [Google Scholar] [CrossRef]
- Sun, X.; Seidman, J.S.; Zhao, P.; Troutman, T.D.; Spann, N.J.; Que, X.; Zhou, F.; Liao, Z.; Pasillas, M.; Yang, X.; et al. Neutralization of Oxidized Phospholipids Ameliorates Non-alcoholic Steatohepatitis. Cell Metab. 2020, 31, 189–206.e8. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mth-6 | Mth-18 | Mth-30 | |
|---|---|---|---|
| C8:0 | 8.16 ± 0.36 a | 8.68 ± 0.86 a | 8.24 ± 0.28 a |
| C10:0 | 139.62 ± 3.75 a | 86.83 ± 1.51 b | 91.44 ± 1.83 b |
| C12:0 | 333.80 ± 59.03 a | 75.69 ± 8.48 b | 78.94 ± 7.70 b |
| C13:0 | 24.80 ± 1.81 a | 13.74 ± 2.79 b | 14.08 ± 1.19 b |
| C14:0 | 3406.00 ± 399.28 a | 1913.76 ± 206.22 b | 1941.31 ± 43.94 b |
| C15:0 | 520.03 ± 25.69 a | 357.80 ± 54.98 b | 376.36 ± 15.74 b |
| C16:0 | 13,629.55 ± 147.70 a | 13,150.13 ± 686.79 a | 1294.02 ± 175.82 a |
| C17:0 | 911.50 ± 16.76 a | 1031.60 ± 10.67 b | 972.83 ± 13.35 c |
| C18:0 | 9358.51 ± 186.43 a | 13,501.52 ± 132.74 b | 13,230.81 ± 441.67 b |
| C20:0 | 36.82 ± 1.90 ab | 34.29 ± 4.94 a | 43.14 ± 1.77 b |
| C21:0 | 57.00 ± 6.2 ab | 44.82 ± 1.42 a | 63.22 ± 8.83 b |
| C22:0 | 15.50 ± 0.75 a | 7.36 ± 4.99 b | 24.66 ± 4.40 c |
| C23:0 | 5.98 ± 0.77 a | 5.85 ± 0.41 a | 5.58 ± 0.33 a |
| C24:0 | 47.40 ± 2.74 a | 51.91 ± 5.18 a | 48.21 ± 4.49 a |
| C14:1 | 130.72 ± 9.47 a | 30.26 ± 2.64 b | 45.35 ± 0.40 c |
| C15:1 | 25.03 ± 6.02 a | 58.22 ± 14.75 b | 49.93 ± 2.00 b |
| C16:1 | 1454.34 ± 76.38 a | 1090.31 ± 127.17 b | 1226.24 ± 8.20 b |
| C17:1 | 18.18 ± 1.19 a | 23.04 ± 0.7.84 a | 17.01 ± 2.04 a |
| 9t-C18:1 | 20.81 ± 3.65 a | 24.79 ± 4.89 a | 27.27 ± 2.92 a |
| 9c-C18:1 | 20,112.40 ± 944.72 a | 19,495.23 ± 405.04 a | 20,003.87 ± 555.85 a |
| C20:1 | 443.73 ± 10.67 a | 456.78 ± 61.51 a | 448.31 ± 1.22 a |
| C22:1 | 9.34 ± 0.97 a | 10.55 ± 1.76 a | 8.57 ± 0.05 a |
| C24:1 | 14.06 ± 1.30 a | 12.44 ± 2.14 a | 12.51 ± 1.00 a |
| 9t,12 t-C18:2 | 190.81 ± 3.82 a | 182.79 ± 5.74 a | 187.92 ± 1.75 a |
| 9c,12 c-C18:2 | 726.44 ± 18.40 a | 711.98 ± 53.82 a | 673.75 ± 19.29 a |
| C18:3 n-6 | 13.80 ± 0.37 a | 15.22 ± 2.33 ab | 17.18 ± 0.61 b |
| C18:3 n-3 | 828.49 ± 3.23 a | 472.33 ± 38.33 b | 465.65 ± 22.34 b |
| C20:2 | 7.20 ± 0.40 a | 6.22 ± 0.15 a | 7.51 ± 1.95 a |
| C20:3 n-6 | 5.05 ± 1.36 a | 7.08 ± 2.98 a | 4.16 ± 0.46 a |
| C20:3 n-3 | 39.22 ± 1.06 a | 42.04 ± 0.47 a | 42.53 ± 7.56 a |
| C20:4 n-6 | 7.21 ± 0.56 a | 7.49 ± 0.56 a | 7.86 ± 0.89 a |
| C22:2 n-6 | 5.26 ± 0.08 a | 5.48 ± 0.81 a | 5.35 ± 0.12 a |
| C20:5 n-3 | 6.12 ± 1.00 a | 9.37 ± 1.01 b | 13.56 ± 1.63 c |
| C22:6 n-3 | 12.76 ± 0.56 a | 19.18 ± 3.30 b | 15.08 ± 2.33 ab |
| SFA | 28,492.68 ± 268.60 a | 30,283.97 ± 801.37 b | 29,861.85 ± 590.09 b |
| MUFA | 22,228.62 ± 965.56 a | 21,201.61 ± 478.54 a | 21,839.05 ± 563.52 a |
| PUFA | 1842.35 ± 21.89 a | 1479.19 ± 69.20 b | 1440.55 ± 53.10 b |
| PUFA/SFA | 0.06 ± 0.00 a | 0.05 ± 0.00 b | 0.05 ± 0.00 b |
| n-6 | 948.56 ± 19.09 a | 930.04 ± 60.65 a | 896.22 ± 21.27 a |
| n-3 | 886.58 ± 3.93 a | 542.92 ± 40.81 b | 536.82 ± 33.43 b |
| n-6/n-3 | 1.07 ± 0.02 a | 1.71 ± 0.05 b | 1.67 ± 0.07 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; He, X.; Yun, Y.; Chen, L.; Huang, Y.; Wu, Q.; Qin, X.; Wu, H.; Wu, J.; Sha, R.; et al. The Characterization of Subcutaneous Adipose Tissue in Sunit Sheep at Different Growth Stages: A Comprehensive Analysis of the Morphology, Fatty Acid Profile, and Metabolite Profile. Foods 2024, 13, 544. https://doi.org/10.3390/foods13040544
Han Y, He X, Yun Y, Chen L, Huang Y, Wu Q, Qin X, Wu H, Wu J, Sha R, et al. The Characterization of Subcutaneous Adipose Tissue in Sunit Sheep at Different Growth Stages: A Comprehensive Analysis of the Morphology, Fatty Acid Profile, and Metabolite Profile. Foods. 2024; 13(4):544. https://doi.org/10.3390/foods13040544
Chicago/Turabian StyleHan, Yunfei, Xige He, Yueying Yun, Lu Chen, Yajuan Huang, Qiong Wu, Xia Qin, Haiyan Wu, Jindi Wu, Rina Sha, and et al. 2024. "The Characterization of Subcutaneous Adipose Tissue in Sunit Sheep at Different Growth Stages: A Comprehensive Analysis of the Morphology, Fatty Acid Profile, and Metabolite Profile" Foods 13, no. 4: 544. https://doi.org/10.3390/foods13040544