
Effects of Germination on the Structure, Functional Properties, and In Vitro Digestibility of a Black Bean (Glycine max (L.) Merr.) Protein Isolate
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Main Reagents
2.2. Instruments
2.3. Germination Treatment and Preparation of Black Bean Protein Isolate
2.4. Determination of Total Protein Content, Soluble Protein Content, and Protein Solubility in Germinated Black Bean Powder
2.5. Amino Acid Composition Analysis of the Germinated Black Soya Bean Isolated Protein
2.6. Protein Structure Determination
2.6.1. Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.6.2. Fourier Infrared Spectrum Analysis
2.6.3. Fluorescence Spectrometry
2.6.4. Ultraviolet Spectrometric Measurements
2.6.5. Particle Size Distribution and ζ Potential
2.7. Turbidity
2.8. Foaming Ability Measurements
2.9. Emulsifying Ability Measurements
2.10. Polarized Microscope Observations
2.11. In Vitro Digestibility Determination
2.12. Statistical Analysis
3. Results
3.1. Total Protein and Soluble Protein Contents of Germinated Black Beans
3.2. Amino Acid Composition and Content of the Germinated Black Bean Protein Isolate
3.3. Effect of Different Germination Times on Isolated Protein Structure
3.3.1. SDS-PAGE Analysis
3.3.2. Spectral Analysis
3.3.3. Particle Size and ζ-Potential
3.4. Solubility and Turbidity of the Germinated Black Bean Protein Isolate
3.5. Emulsification and Foamability
3.6. Polarized Microscope Observations
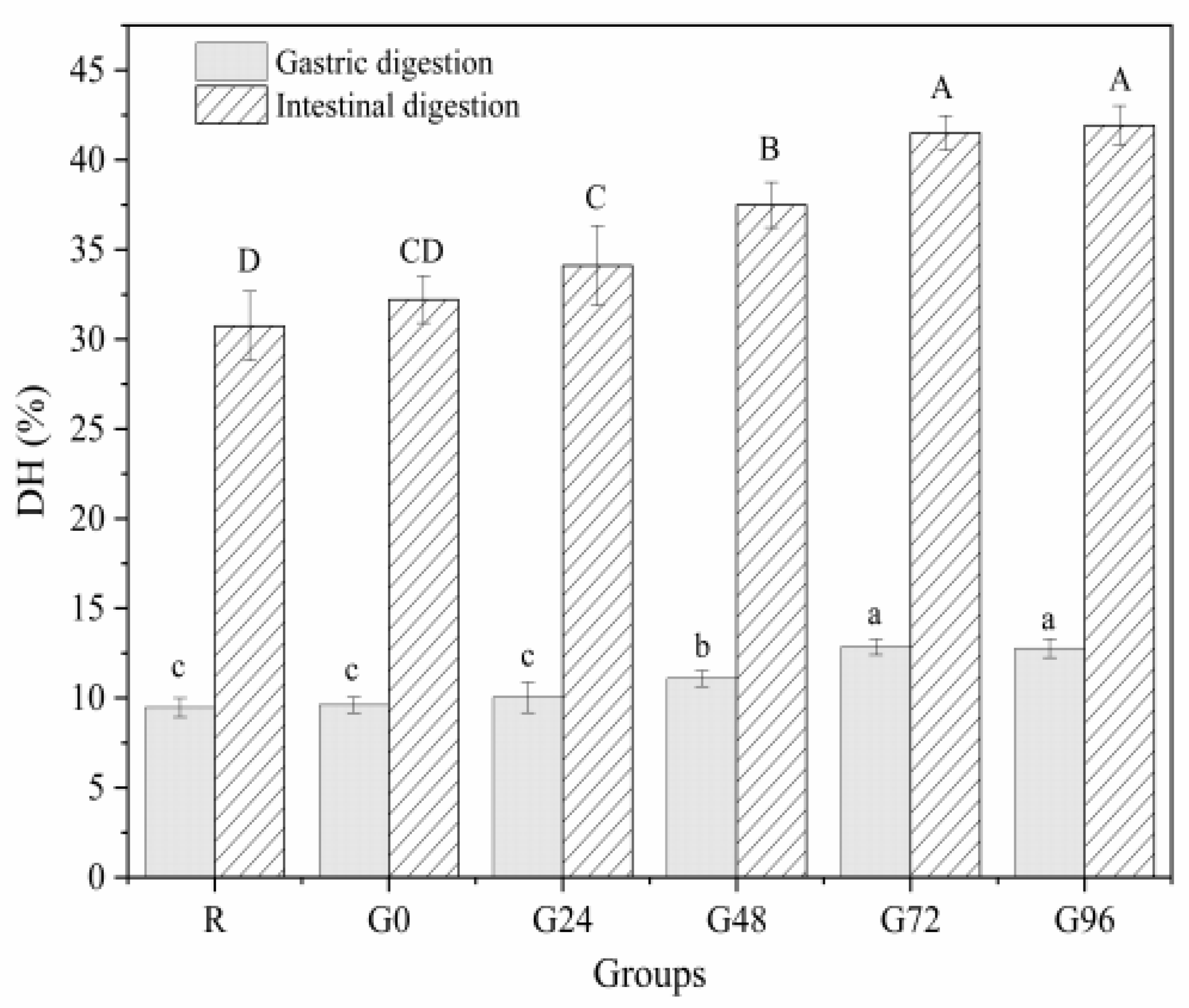
3.7. In Vitro Digestibility
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Piipponen, J.; Jalava, M.; de Leeuw, J.; Rizayeva, A.; Godde, C.; Cramer, G.; Herrero, M.; Kummu, M. Global trends in grassland carrying capacity and relative stocking density of livestock. Glob. Chang. Biol. 2022, 28, 3902–3919. [Google Scholar] [CrossRef]
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef]
- Munialo, C.D.; Vriesekoop, F. Plant-based foods as meat and fat substitutes. Food Sci. Nutr. 2023, 11, 4898–4911. [Google Scholar] [CrossRef]
- Monteiro, C.A.; Cannon, G.; Moubarac, J.C.; Martins, A.P.B.; Martins, C.A.; Garzillo, J.; Canella, D.S.; Baraldi, L.G.; Barciotte, M.; Louzada, M.L.D.C.; et al. Dietary guidelines to nourish humanity and the planet in the twenty-first century. A blueprint from Brazil. Public Health Nutr. 2015, 18, 2311–2322. [Google Scholar] [CrossRef]
- Melzer, H.M.; Brantsæter, A.L.; Trolle, E.; Eneroth, H.; Fogelholm, M.; Ydersbond, T.A.; Birgisdottir, B.E. Environmental Sustainability Perspectives of the Nordic Diet. Nutrients 2019, 11, 2248. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef]
- Langyan, S.; Yadava, P.; Khan, F.N.; Dar, Z.A.; Singh, R.; Kumar, A. Sustaining protein nutrition through plant-based Foods. Front. Nutr. 2022, 8, 772573. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Zong, G.; Willett, W.C.; Zock, P.L.; Wanders, A.J.; Hu, F.B.; Sun, Q. Associations of monounsaturated fatty acids from plant and animal sources with total and cause-specific mortality in two US prospective cohort studies. Circ. Res. 2019, 124, 1266–1275. [Google Scholar] [CrossRef]
- Drulyte, D.; Orlien, V. The effect of processing on digestion of legume proteins. Foods 2019, 8, 224. [Google Scholar] [CrossRef]
- Kumar, M.; Suhag, R.; Hasan, M.; Dhumal, S.; Radha; Pandiselvam, R.; Senapathy, M.; Sampathrajan, V.; Punia, S.; Sayed, A.A.S.; et al. Black soybean (Glycine max (L.) Merr.): Paving the way toward new nutraceutical. Crit. Rev. Food Sci. Nutr. 2023, 63, 6208–6234. [Google Scholar] [CrossRef]
- Hernández, D.F.; Mojica, L.; Berhow, M.A.; Brownstein, K.; Cervantes, E.L.; de Mejia, E.G. Black and pinto beans (Phaseolus vulgaris L.) unique mexican varieties exhibit antioxidant and anti-inflammatory potential. Food Res. Int. 2023, 169, 112816. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, J.; Li, Y.; Wang, Z.; Liang, J.; Wang, R.; Chen, Y.; Ma, W.; Qi, B.; Zhang, M. Effects of ultrasound on the structure and physical properties of black bean protein isolates. Food Res. Int. 2014, 62, 595–601. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, H.; Zhang, S.; Li, J.; Zhu, X.; Jin, H.; Xu, J. Improvement of protein emulsion stability through glycosylated black bean protein covalent interaction with (−)-epigallocatechin-3-gallate. RSC Adv. 2021, 11, 2546–2555. [Google Scholar] [CrossRef]
- Jin, H.; Zhao, Q.; Feng, H.; Wang, Y.; Wang, J.; Liu, Y.; Han, D.; Xu, J. Changes on the structural and physicochemical properties of conjugates prepared by the maillard reaction of black bean protein isolates and glucose with ultrasound pretreatment. Polymers 2019, 11, 848. [Google Scholar] [CrossRef]
- Rodríguez-España, M.; Figueroa-Hernández, C.Y.; Figueroa-Cárdenas, J.D.D.; Rayas-Duarte, P.; Hernández-Estrada, Z.J. Effects of germination and lactic acid fermentation on nutritional and rheological properties of sorghum: A graphical review. Curr. Res. Food Sci. 2022, 5, 807–812. [Google Scholar] [CrossRef]
- Lemmens, E.; Moroni, A.V.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.A.; den Broeck, H.C.; Brouns, F.J.P.H.; Brier, N.; et al. Impact of cereal seed sprouting on it’s nutritional and technological properties: A critical review. Compr. Rev. Food Sci. Food Saf. 2018, 18, 305–328. [Google Scholar] [CrossRef]
- Guajardo-Flores, D.; Garcia-Patino, M.; Serna-Guerrero, D.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Characterization and quantification of saponins and flavonoids in sprouts, seed coats and cotyledons of germinated black beans. Food Chem. 2012, 134, 1312–1319. [Google Scholar] [CrossRef]
- Guajardo-Flores, D.; Serna-Saldívar, S.O.; Gutiérrez-Uribe, J.A. Evaluation of the antioxidant and antiproliferative activities of extracted saponins and flavonols from germinated black beans (Phaseolus vulgaris L.). Food Chem. 2013, 141, 1497–1503. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, J.; Li, J.; Sun, H.; Liu, Y. Physicochemical and antioxidative characteristics of black bean protein hydrolysates obtained from different enzymes. Food Hydrocoll. 2019, 97, 105222. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, S.; Feng, Y.; Wang, H.; Liu, Y.; Wang, C. Modification of the structural and functional characteristics of mung bean globin polyphenol complexes: Exploration under heat treatment conditions. Foods 2023, 12, 2091. [Google Scholar] [CrossRef]
- Mokni Ghribi, A.; Maklouf Gafsi, I.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of enzymatic hydrolysis on conformational and functional properties of chickpea protein isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Mujoo, R.; Trinh, D.T.; Ng, P.K.W. Characterization of storage proteins in different soybean varieties and their relationship to tofu yield and texture. Food Chem. 2003, 82, 265–273. [Google Scholar] [CrossRef]
- Sun, R.; Liu, X.; Yu, Y.; Miao, J.; Leng, K.; Gao, H. Preparation process optimization, structural characterization and in vitro digestion stability analysis of Antarctic krill (Euphausia superba) peptides-zinc chelate. Food Chem. 2021, 340, 128056. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, C.; Gao, X.; Chen, Y.; Kumar Santhanam, R.; Wang, C.; Xu, L.; Chen, H. Interaction characterization of preheated soy protein isolate with cyanidin-3-O-glucoside and their effects on the stability of black soybean seed coat anthocyanins extracts. Food Chem. 2019, 271, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.G.; Taylor, J.A.; Sibilo, M.Q.; Miura, K.; Mallory, K.L.; Mann, C.; Karch, C.; Beck, Z.; Matyas, G.R.; Long, C.A.; et al. Orientation of antigen display on self-assembling protein nanoparticles influences immunogenicity. Vaccines 2021, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Ding, J.; Andrade, J.; Rababah, T.M.; Almajwal, A.; Abulmeaty, M.M.; Feng, H. Modifying the physicochemical properties of pea protein by pH-shifting and ultrasound combined treatments. Ultrason. Sonochem. 2017, 38, 835–842. [Google Scholar] [CrossRef]
- Zhao, X.; Fan, X.; Shao, X.; Cheng, M.; Wang, C.; Jiang, H.; Zhang, X.; Yuan, C. Modifying the physicochemical properties, solubility and foaming capacity of milk proteins by ultrasound-assisted alkaline pH-shifting treatment. Ultrason. Sonochem. 2022, 88, 106089. [Google Scholar] [CrossRef]
- Wang, Q.; Pan, M.-H.; Chiou, Y.-S.; Li, Z.; Ding, B. Surface characteristics and emulsifying properties of whey protein/nanoliposome complexes. Food Chem. 2022, 384, 132510. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, S.; Zou, L.; Wang, B.; Bian, X.; Zhu, P.; Ren, L.; Shi, Y.; Zhang, N. Characterization of structural and functional properties of soybean 11S globulin during the renaturation after the guanidine hydrochloride denaturation. Food Hydrocoll. 2022, 130, 107715. [Google Scholar] [CrossRef]
- Jiang, P.; Liu, N.; Xiu, Y.; Wang, W.; Wang, C.; Zhang, D.; Li, Z. Identification and analysis of antioxidant peptides from sorghum (Sorghum bicolor L. Moench) on the basis of in vitro simulated gastrointestinal digestion. Food Funct. 2022, 13, 9635–9644. [Google Scholar] [CrossRef]
- Wang, K.; Gao, Y.; Zhao, J.; Wu, Y.; Sun, J.; Niu, G.; Zuo, F.; Zheng, X. Effects of in vitro digestion on protein degradation, phenolic compound release, and bioactivity of black bean tempeh. Front. Nutr. 2022, 9, 1017765. [Google Scholar] [CrossRef]
- Xing, L.-J.; Hu, Y.-Y.; Hu, H.-Y.; Ge, Q.-F.; Zhou, G.-H.; Zhang, W.-G. Purification and identification of antioxidative peptides from dry-cured Xuanwei ham. Food Chem. 2016, 194, 951–958. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Berhow, M.A.; Mandarino, J.M.G.; Chang, Y.K.; Mejia, E.G.D. Effect of time and temperature on bioactive compounds in germinated Brazilian soybean cultivar BRS 258. Food Res. Int. 2010, 43, 1856–1865. [Google Scholar] [CrossRef]
- De Souza Rocha, T.; Hernandez, L.M.R.; Mojica, L.; Johnson, M.H.; Chang, Y.K.; González de Mejía, E. Germination of phaseolus vulgaris and alcalase hydrolysis of its proteins produced bioactive peptides capable of improving markers related to type-2 diabetes in vitro. Food Res. Int. 2015, 76, 150–159. [Google Scholar] [CrossRef]
- Concha, D.D.R.M.; Martínez, J.E.B.; Velázquez, T.G.G.; Martínez, C.J.; Ruiz, J.C.R. Impact of germination time on protein solubility and anti-inflammatory properties of Pisum sativum L grains. Food Chem.-X 2022, 13, 100219. [Google Scholar] [CrossRef]
- Amir, R.; Galili, G.; Cohen, H. The metabolic roles of free amino acids during seed development. Plant Sci. 2018, 275, 11–18. [Google Scholar] [CrossRef]
- Bueno, D.B.; da Silva Júnior, S.I.; Seriani Chiarotto, A.B.; Cardoso, T.M.; Neto, J.A.; Lopes dos Reis, G.C.; Glória, M.B.A.; Tavano, O.L. The germination of soybeans increases the water-soluble components and could generate innovations in soy-based foods. LWT 2020, 117, 108599. [Google Scholar] [CrossRef]
- Gonzalez-Montoya, M.; Hernandez-Ledesma, B.; Silvan, J.M.; Mora-Escobedo, R.; Martinez-Villaluenga, C. Peptides derived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells proliferation and inflammation. Food Chem. 2018, 242, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Oyedeji, A.B.; Mellem, J.J.; Ijabadeniyi, O.A. Improvement of some quality attributes of soymilk through optimization of selected soybean sprouting parameters using response surface methodology. CYTA J. Food 2018, 16, 230–237. [Google Scholar] [CrossRef]
- Kim, H.Y.; Hwang, I.G.; Kim, T.M.; Woo, K.S.; Park, D.S.; Kim, J.H.; Kim, D.J.; Lee, J.; Lee, Y.R.; Jeong, H.S. Chemical and functional components in different parts of rough rice (Oryza sativa L.) before and after germination. Food Chem. 2012, 134, 288–293. [Google Scholar] [CrossRef]
- Klose, C.; Arendt, E.K. Proteins in Oats; their synthesis and changes during germination: A Review. Crit. Rev. Food Sci. Nutr. 2012, 52, 629–639. [Google Scholar] [CrossRef]
- Tada, Y.; Kashimura, T. Proteomic analysis of salt-responsive proteins in the mangrove plant, Bruguiera gymnorhiza. Plant Cell Physiol. 2009, 50, 439–446. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, G. Nutritionally Essential Amino Acids. Adv Nutr. 2018, 9, 849–851. [Google Scholar] [CrossRef]
- Luo, M. Chemical and biochemical perspectives of protein lysine methylation. Chem. Rev. 2018, 118, 6656–6705. [Google Scholar] [CrossRef]
- Li, M.; Qi, Y.; Mu, L.; Li, Z.; Zhao, Q.; Sun, J.; Jiang, Q. Effects of processing method on chemical compositions and nutritional quality of ready-to-eat sea cucumber (Apostichopus japonicus). Food Sci. Nutr. 2019, 7, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, S.; Ye, D.; Yu, M.; Lei, J. Effects of Germination on the content of amino acids and anti-nutritional factors of sorghum grain. Sci. Technol. Food Ind. 2022, 43, 87–92. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Andersen, L.L.; Otte, J.; Nielsen, H.H.; Jessen, F.; Jacobsen, C. Antioxidant activity of cod (Gadus morhua) protein hydrolysates: Fractionation and characterisation of peptide fractions. Food Chem. 2016, 204, 409–419. [Google Scholar] [CrossRef]
- Shazly, A.B.; He, Z.; El-Aziz, M.A.; Zeng, M.; Zhang, S.; Qin, F.; Chen, J. Fractionation and identification of novel antioxidant peptides from buffalo and bovine casein hydrolysates. Food Chem. 2017, 232, 753–762. [Google Scholar] [CrossRef]
- Wu, R.; Huang, J.; Huan, R.; Chen, L.; Yi, C.; Liu, D.; Wang, M.; Liu, C.; He, H. New insights into the structure-activity relationships of antioxidative peptide PMRGGGGYHY. Food Chem. 2021, 337, 127678. [Google Scholar] [CrossRef]
- Nwachukwu, I.D.; Aluko, R.E. Physicochemical and emulsification properties of flaxseed (Linum usitatissimum) albumin and globulin fractions. Food Chem. 2018, 255, 216–225. [Google Scholar] [CrossRef]
- Sandoval-Oliveros, M.R.; Paredes-López, O. Isolation and characterization of proteins from chia seeds (Salvia hispanica L.). J. Agric. Food Chem. 2012, 61, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.-T.; Qian, Z.-J.; Lee, B.; Heo, S.-J.; Kim, K.-N.; Jeon, Y.-J.; Park, W.S.; Choi, I.-W.; Jang, C.H.; Ko, S.-C.; et al. Fucoxanthin derivatives from Sargassum siliquastrum inhibit matrix metalloproteinases by suppressing NF-κB and MAPKs in human fibrosarcoma cells. Algae 2014, 29, 355–366. [Google Scholar] [CrossRef]
- Wang, K.; Han, L.; Hong, H.; Pan, J.; Liu, H.; Luo, Y. Purification and identification of novel antioxidant peptides from silver carp muscle hydrolysate after simulated gastrointestinal digestion and transepithelial transport. Food Chem. 2021, 342, 128275. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, F.; Gardner, C.D. Dietary protein and amino acids in vegetarian diets—A review. Nutrients 2019, 11, 2661. [Google Scholar] [CrossRef]
- Teraishi, M.; Takahashi, M.; Hajika, M.; Matsunaga, R.; Uematsu, Y.; Ishimoto, M. Suppression of soybean β-conglycinin genes by a dominant gene, Scg-1. Theor. Appl. Genet. 2001, 103, 1266–1272. [Google Scholar] [CrossRef]
- Wang, B.; Teng, D.; Yu, C.; Yao, L.; Ma, X.; Wu, T. Increased sulfur-containing amino acid content and altered conformational characteristics of soybean proteins by rebalancing 11S and 7S compositions. Front. Plant Sci. 2022, 13, 828153. [Google Scholar] [CrossRef]
- López-Monterrubio, D.I.; Lobato-Calleros, C.; Alvarez-Ramirez, J.; Vernon-Carter, E.J. Huauzontle (Chenopodium nuttalliae Saff.) protein: Composition, structure, physicochemical and functional properties. Food Hydrocoll. 2020, 108, 106043. [Google Scholar] [CrossRef]
- Siano, F.; Sorrentino, G.; Riccardi, M.; De Cunzo, F.; Orefice, G.; Volpe, M.G. Chemical, nutritional, and spectroscopic characterization of typical ecotypes of Mediterranean area beans. Eur. Food Res. Technol. 2017, 244, 795–804. [Google Scholar] [CrossRef]
- Sui, X.; Sun, H.; Qi, B.; Zhang, M.; Li, Y.; Jiang, L. Functional and conformational changes to soy proteins accompanying anthocyanins: Focus on covalent and non-covalent interactions. Food Chem. 2018, 245, 871–878. [Google Scholar] [CrossRef]
- Hunt, N.G.; Gregoret, L.M.; Cohen, F.E. The origins of protein secondary structure. Effects of packing density and hydrogen bonding studied by a fast conformational search. J. Mol. Biol. 1994, 241, 214–225. [Google Scholar] [CrossRef]
- Carbonaro, M.; Nucara, A. Secondary structure of food proteins by Fourier transform spectroscopy in the mid-infrared region. Amino Acids 2009, 38, 679–690. [Google Scholar] [CrossRef]
- Yang, X.; Su, Y.; Li, L. Study of soybean gel induced by Lactobacillus plantarum: Protein structure and intermolecular interaction. LWT 2020, 119, 108794. [Google Scholar] [CrossRef]
- Yu, J.; Sun, B.; Zhang, S.; Liu, X.; Xie, P. The effect of different induction methods on the structure and physicochemical properties of glycosylated soybean isolate gels. Foods 2022, 11, 3595. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, C.; Zhao, J.; Ma, T.; He, Z.; Huang, M.; Wang, Y. Modification of structure and functionalities of ginkgo seed proteins by pH-shifting treatment. Food Chem. 2021, 358, 129862. [Google Scholar] [CrossRef]
- Leu, M.; Marciniak, A.; Chamberland, J.; Pouliot, Y.; Bazinet, L.; Doyen, A. Effect of skim milk treated with high hydrostatic pressure on permeate flux and fouling during ultrafiltration. J. Dairy Sci. 2017, 100, 7071–7082. [Google Scholar] [CrossRef]
- Chen, X.; Zou, Y.; Han, M.; Pan, L.; Xing, T.; Xu, X.; Zhou, G. Solubilisation of myosin in a solution of low ionic strength l -histidine: Significance of the imidazole ring. Food Chem. 2016, 196, 42–49. [Google Scholar] [CrossRef]
- Yan, X.; Jia, Y.; Man, H.; Sun, S.; Huang, Y.; Qi, B.; Li, Y. Tracking the driving forces for the unfolding and folding of kidney bean protein isolates: Revealing mechanisms of dynamic changes in structure and function. Food Chem. 2023, 402, 134230. [Google Scholar] [CrossRef]
- Wong, B.T.; Day, L.; Augustin, M.A. Deamidated wheat protein–dextran Maillard conjugates: Effect of size and location of polysaccharide conjugated on steric stabilization of emulsions at acidic pH. Food Hydrocoll. 2011, 25, 1424–1432. [Google Scholar] [CrossRef]
- Yang, H.; Li, X.; Gao, J.; Tong, P.; Yang, A.; Chen, H. Germination-assisted enzymatic hydrolysis can improve the quality of soybean protein. J. Food Sci. 2017, 82, 1814–1819. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Mourad, F.K.; Zhang, X.; Ahn, D.U.; Cai, Z.; Jin, Y. Phase separation behavior and characterization of ovalbumin and propylene glycol alginate complex coacervates. Food Hydrocoll. 2020, 108, 105978. [Google Scholar] [CrossRef]
- Jambrak, A.R.; Mason, T.J.; Lelas, V.; Herceg, Z.; Herceg, I.L. Effect of ultrasound treatment on solubility and foaming properties of whey protein suspensions. J. Food Eng. 2008, 86, 281–287. [Google Scholar] [CrossRef]
- O’Sullivan, J.; Murray, B.; Flynn, C.; Norton, I. The effect of ultrasound treatment on the structural, physical and emulsifying properties of animal and vegetable proteins. Food Hydrocoll. 2016, 53, 141–154. [Google Scholar] [CrossRef]
- Fei, W.; Wen, B.Z.; Rui, J.Y.; Xiao, H.; Wei, Z. Extraction and functional properties of quinoa protein isolates. Food Sci. Technol. 2018, 43, 228–234. [Google Scholar] [CrossRef]
- Akio, K.; Shuryo, N. Hydrophobicity determined by a fluorescence probe method and its correlation with surface properties of proteins. Acta Biochim. Biophys. Sin. 1980, 624, 13–20. [Google Scholar] [CrossRef]
- Agyare, K.K.; Addo, K.; Xiong, Y.L. Emulsifying and foaming properties of transglutaminase-treated wheat gluten hydrolysate as influenced by pH, temperature and salt. Food Hydrocoll. 2009, 23, 72–81. [Google Scholar] [CrossRef]
- Setia, R.; Dai, Z.; Nickerson, M.T.; Sopiwnyk, E.; Malcolmson, L.; Ai, Y. Impacts of short-term germination on the chemical compositions, technological characteristics and nutritional quality of yellow pea and faba bean flours. Food Res. Int. 2019, 122, 263–272. [Google Scholar] [CrossRef]
- Di, Y.; Li, X.; Chang, X.W.; Gu, R.J.; Duan, X.; Liu, F.G.; Liu, X.B.; Wang, Y.T. Impact of germination on structural, functional properties and in vitro protein digestibility of sesame (Sesamum indicum L.) protein. LWT 2022, 154, 112651. [Google Scholar] [CrossRef]
- Bautista-Exposito, S.; Vandenberg, A.; Penas, E.; Frias, J.; Martinez-Villaluenga, C. Lentil and fava bean with contrasting germination kinetics: A focus on digestion of proteins and bioactivity of resistant peptides. Front. Plant Sci. 2021, 12, 754287. [Google Scholar] [CrossRef]
- Devi, C.B.; Kushwaha, A.; Kumar, A. Sprouting characteristics and associated changes in nutritional composition of cowpea (Vigna unguiculata). J. Food Sci. Technol. 2015, 52, 6821–6827. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid Species | Amino Acid Content (g/100 g) | ||||||
|---|---|---|---|---|---|---|---|
| R | G0 | G24 | G48 | G72 | G96 | ||
| Hydrophobic Amino Acid | Ala | 3.793 ± 0.015 e | 3.775 ± 0.011 e | 3.898 ± 0.014 d | 4.072 ± 0.015 c | 4.267 ± 0.009 a | 4.105 ± 0.014 b |
| * Val | 4.421 ± 0.008 e | 4.369 ± 0.010 f | 4.586 ± 0.022 d | 4.703 ± 0.012 b | 4.908 ± 0.017 a | 4.663 ± 0.025 c | |
| * Ile | 1.152 ± 0.019 de | 1.138 ± 0.013 e | 1.370 ± 0.021 a | 1.312 ± 0.005 b | 1.246 ± 0.007 c | 1.174 ± 0.016 d | |
| * Leu | 4.024 ± 0.016 f | 4.078 ± 0.023 e | 4.189 ± 0.010 d | 4.287 ± 0.009 c | 4.534 ± 0.016 a | 4.334 ± 0.021 b | |
| Pro | 7.284 ± 0.024 f | 7.325 ± 0.006 e | 7.541 ± 0.011 d | 7.738 ± 0.017 c | 8.211 ± 0.007 a | 7.874 ± 0.011 b | |
| * Phe | 4.091 ± 0.014 f | 4.127 ± 0.012 e | 4.294 ± 0.013 d | 4.441 ± 0.022 c | 4.659 ± 0.018 a | 4.571 ± 0.008 b | |
| * Met | 3.836 ± 0.021 e | 4.361 ± 0.010 d | 4.393 ± 0.018 c | 4.431 ± 0.010 b | 4.400 ± 0.005 c | 4.659 ± 0.020 a | |
| Total Hydrophobic Amino Acids | 28.601 ± 0.012 f | 29.173 ± 0.025 e | 30.271 ± 0.014 d | 30.984 ± 0.015 c | 32.225 ± 0.016 a | 31.380 ± 0.020 b | |
| Polar Amino Acid | * Thr | 3.511 ± 0.007 f | 3.584 ± 0.011 e | 3.663 ± 0.014 d | 3.800 ± 0.015 c | 3.943 ± 0.006 a | 3.871 ± 0.011 b |
| Ser | 4.767 ± 0.016 f | 5.160 ± 0.012 e | 5.213 ± 0.018 d | 5.367 ± 0.019 c | 5.498 ± 0.005 b | 5.536 ± 0.008 a | |
| Gly | 3.547 ± 0.012 f | 3.856 ± 0.013 e | 3.909 ± 0.009 d | 3.987 ± 0.012 c | 4.072 ± 0.009 b | 4.118 ± 0.007 a | |
| Tyr | 3.504 ± 0.009 e | 3.440 ± 0.012 f | 3.550 ± 0.011 d | 3.653 ± 0.011 c | 3.908 ± 0.002 a | 3.709 ± 0.010 b | |
| Total Polar Amino Acid | 15.329 ± 0.101 f | 16.040 ± 0.020 e | 16.335 ± 0.017 d | 16.807 ± 0.012 c | 17.421 ± 0.011 a | 17.234 ± 0.011 b | |
| Acidic Amino Acid | Asp | 11.760 ± 0.008 d | 11.449 ± 0.018 f | 11.620 ± 0.013 e | 12.009 ± 0.014 c | 12.684 ± 0.011 a | 12.258 ± 0.020 b |
| Glu | 15.925 ± 0.013 e | 19.816 ± 0.007 c | 19.896 ± 0.015 b | 19.821 ± 0.005 c | 19.323 ± 0.007 d | 21.136 ± 0.015 a | |
| Total Acidic Amino Acid | 27.685 ± 0.009 f | 31.265 ± 0.014 e | 31.516 ± 0.012 d | 31.830 ± 0.009 c | 32.007 ± 0.009 b | 33.394 ± 0.019 a | |
| Basic Amino Acid | * Lys | 4.956 ± 0.011 e | 5.712 ± 0.011 d | 5.739 ± 0.012 c | 5.760 ± 0.002 b | 5.743 ± 0.011 bc | 6.123 ± 0.009 a |
| * His | 2.063 ± 0.011 d | 2.336 ± 0.008 b | 2.277 ± 0.012 c | 2.355 ± 0.009 b | 2.286 ± 0.013 c | 2.498 ± 0.012 a | |
| Arg | 5.604 ± 0.010 e | 7.030 ± 0.009 b | 6.937 ± 0.015 c | 6.950 ± 0.009 c | 6.604 ± 0.006 d | 7.398 ± 0.011 a | |
| Total Basic Amino Acid | 12.623 ± 0.012 e | 15.078 ± 0.011 b | 14.953 ± 0.012 c | 15.065 ± 0.021 b | 14.633 ± 0.018 d | 16.019 ± 0.015 a | |
| EAA | 28.054 ± 0.011 f | 29.705 ± 0.008 e | 30.511 ± 0.010 d | 31.089 ± 0.017 c | 31.719 ± 0.011 b | 31.893 ± 0.014 a | |
| TAA | 84.238 ± 0.019 f | 91.556 ± 0.015 e | 93.075 ± 0.011 d | 94.686 ± 0.012 c | 96.25 ± 0.009 b | 98.027 ± 0.012 a | |
| Protein | Subunit | Relative Content of Isolated Protein Subunits at Different Germination Times/% | |||||
|---|---|---|---|---|---|---|---|
| R | G0 | G24 | G48 | G72 | G96 | ||
| 7S Globulin | a′ | 3.69 | 4.00 | 2.93 | 2.71 | 2.24 | 2.11 |
| a | 3.75 | 3.92 | 3.68 | 3.41 | 2.66 | 2.44 | |
| β | 3.37 | 3.46 | 3.64 | 3.79 | 3.50 | 3.43 | |
| a′ + a + β | 10.80 | 11.37 | 10.24 | 9.90 | 8.40 | 7.98 | |
| 11S Globulin | Acid | 19.83 | 20.63 | 20.04 | 19.39 | 17.48 | 16.50 |
| Basic | 10.69 | 11.21 | 10.87 | 11.19 | 12.77 | 12.83 | |
| Acid + Basic | 30.52 | 31.84 | 30.91 | 30.58 | 30.25 | 29.33 | |
| 7S + 11S | 41.32 | 43.22 | 41.15 | 40.48 | 38.65 | 37.31 | |
| 11S/7S | 2.82 | 2.80 | 3.02 | 3.09 | 3.60 | 3.67 | |
| β-Sheet (%) | Random Coil (%) | α-Helix (%) | β-Turn (%) | |
|---|---|---|---|---|
| R | 35.91 ± 0.27 a | 17.22 ± 0.13 a | 15.81 ± 0.62 c | 24.87 ± 0.18 ab |
| G0 | 36.21 ± 0.79 a | 15.98 ± 2.03 a | 15.67 ± 0.86 c | 26.12 ± 2.35 a |
| G24 | 33.05 ± 2.69 b | 15.66 ± 1.80 a | 23.45 ± 1.43 b | 25.53 ± 2.48 ab |
| G48 | 30.87 ± 1.72 bc | 15.42 ± 1.54 a | 27.01 ± 2.08 a | 22.47 ± 1.48 bc |
| G72 | 29.79 ± 0.68 c | 16.59 ± 0.28 a | 29.05 ± 0.63 a | 19.94 ± 0.51 c |
| G96 | 28.99 ± 0.69 c | 17.33 ± 2.32 a | 28.94 ± 1.08 a | 20.17 ± 1.85 c |
| Groups | R | G0 | G24 | G48 | G72 | G96 |
|---|---|---|---|---|---|---|
| Particle size (nm) | 17.12 ± 2.89 | 14.96 ± 2.91 | 15.63 ± 2.94 | 16.47 ± 2.82 | 15.03 ± 2.00 | 11.62 ± 1.75 |
| ζ-potential (mV) | −22.33 ± 0.57 b | −23.23 ± 1.39 ab | −22.97 ± 0.90 ab | −23.17 ± 0.40 ab | −24.47 ± 0.65 a | −24.13 ± 0.75 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-H.; Tai, Z.-J.; Song, X.-J.; Li, Z.-J.; Zhang, D.-J. Effects of Germination on the Structure, Functional Properties, and In Vitro Digestibility of a Black Bean (Glycine max (L.) Merr.) Protein Isolate. Foods 2024, 13, 488. https://doi.org/10.3390/foods13030488
Wang X-H, Tai Z-J, Song X-J, Li Z-J, Zhang D-J. Effects of Germination on the Structure, Functional Properties, and In Vitro Digestibility of a Black Bean (Glycine max (L.) Merr.) Protein Isolate. Foods. 2024; 13(3):488. https://doi.org/10.3390/foods13030488
Chicago/Turabian StyleWang, Xin-Hui, Zhen-Jia Tai, Xue-Jian Song, Zhi-Jiang Li, and Dong-Jie Zhang. 2024. "Effects of Germination on the Structure, Functional Properties, and In Vitro Digestibility of a Black Bean (Glycine max (L.) Merr.) Protein Isolate" Foods 13, no. 3: 488. https://doi.org/10.3390/foods13030488