
Use of Anthracophyllum discolor and Stereum hirsutum as a Suitable Strategy for Delignification and Phenolic Removal of Olive Mill Solid Waste

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Olive Mill Solid Waste (OMSW)
2.3. Qualitative Detection of Lignocellulolytic Enzymes
2.4. Mycelial Growth Assays
2.5. OMSW Pretreatment Assays
2.6. Enzymatic Activity Quantification
2.7. Microstructure Analysis
3. Results and Discussion
3.1. Olive Mill Solid Waste Characterization
3.2. Qualitative Detection of Lignocellulolytic Enzymes
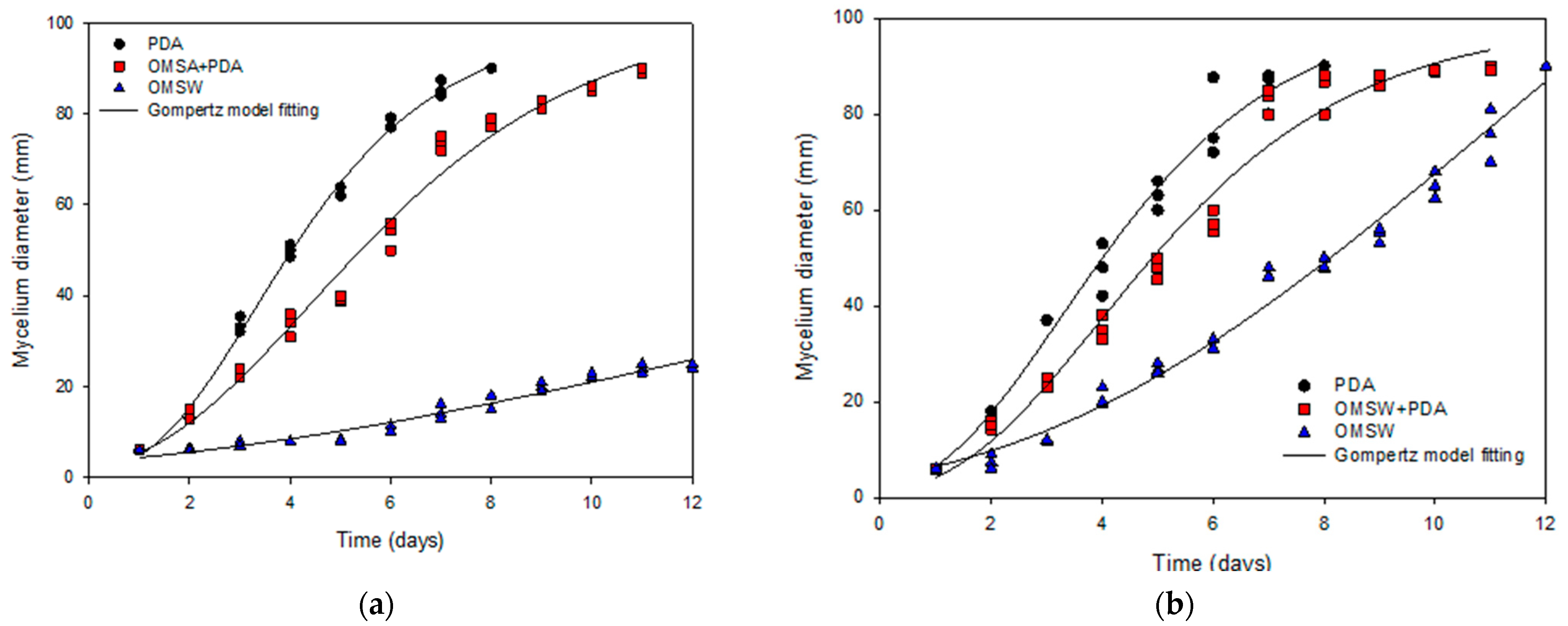
3.3. Mycelial Growth Assays
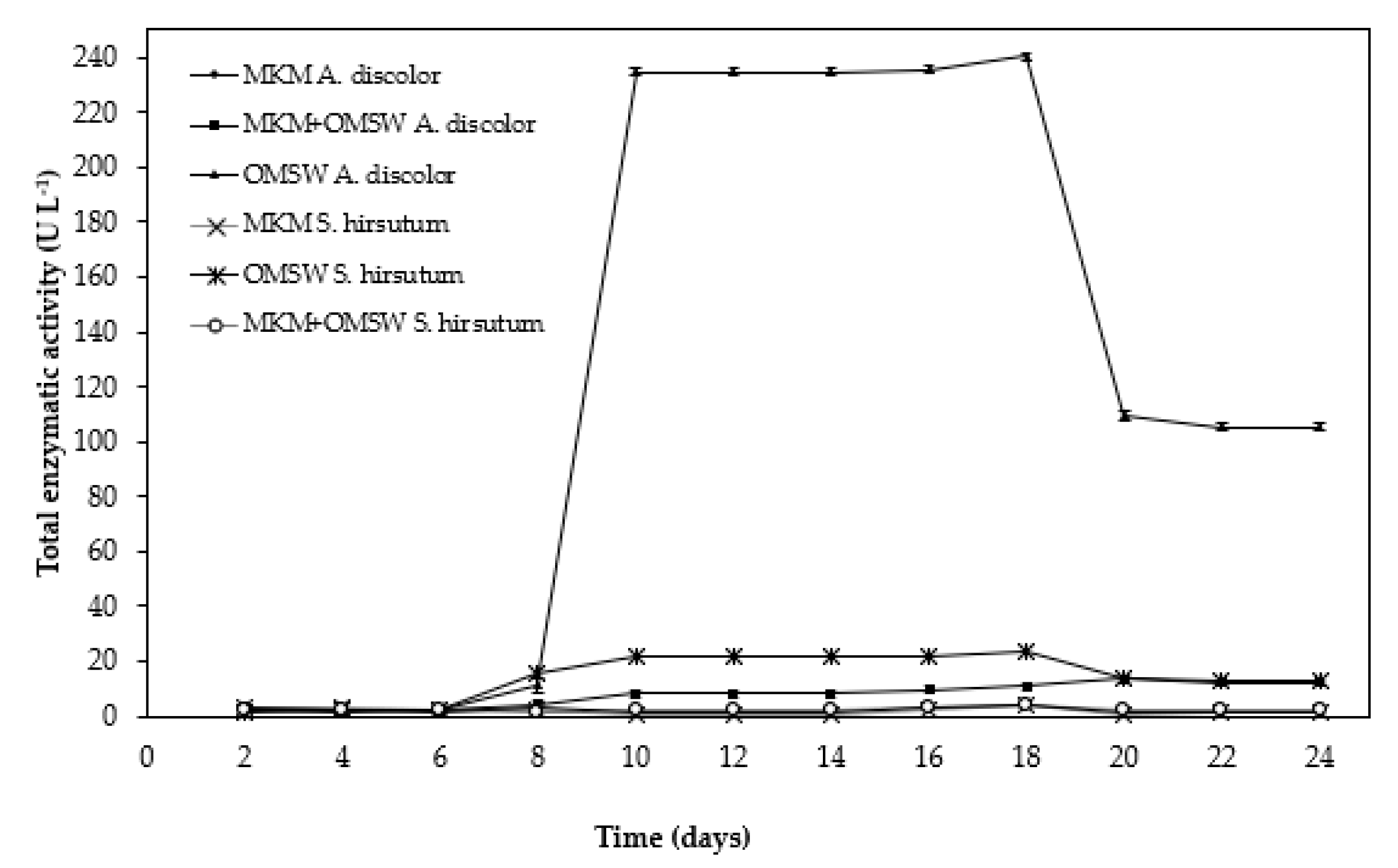
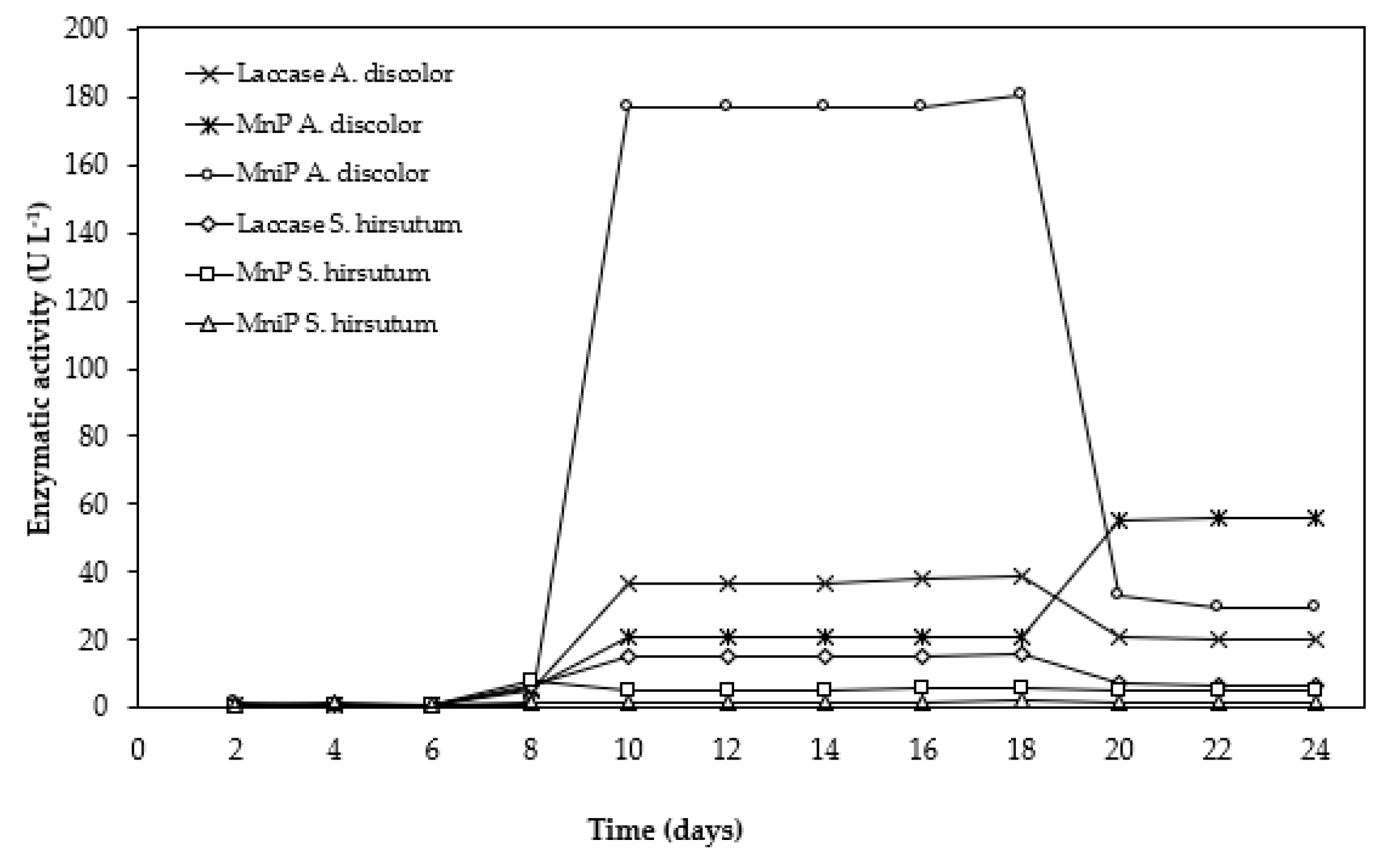
3.4. Enzymatic Extract Production Using OMSW
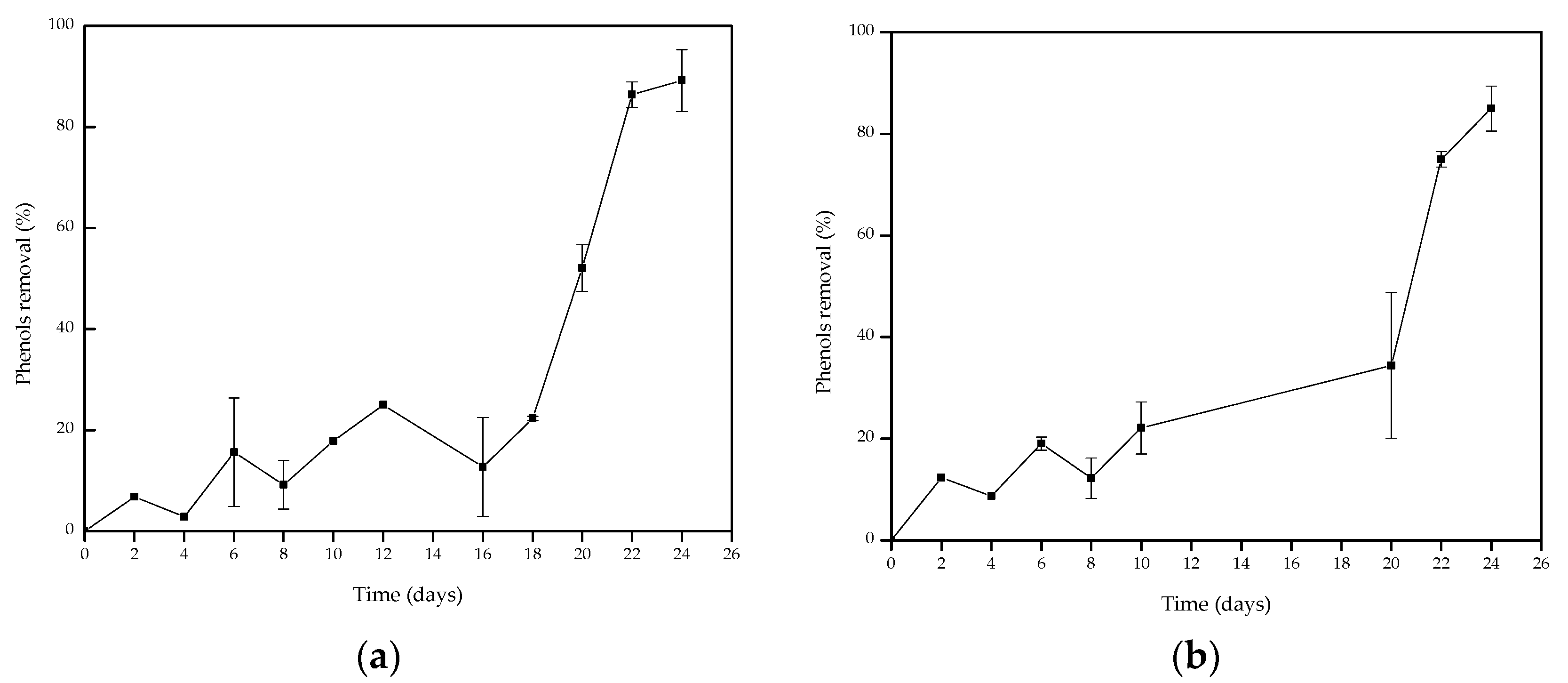
3.5. OMSW Phenol Removal Using A. discolor and S. hirsutum
3.6. Microstructure Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hermosilla, E.; Schalchli, H.; Mutis, A.; Diez, M.C. Combined effect of enzyme inducers and nitrate on selective lignin degradation in wheat straw by Ganoderma lobatum. Environ. Sci. Pollut. Res. 2017, 24, 21984–21996. [Google Scholar] [CrossRef] [PubMed]
- Al-Mallahi, J.; Furuichi, T.; Ishii, K. Appropriate conditions for applying NaOH-pretreated two-phase olive milling waste for codigestion with food waste to enhance biogas production. Waste Manag. 2016, 48, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Li, Y. Fungal pretreatment of lignocellulosic biomass. Biotechnol. Adv. 2012, 30, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Parenti, A.; Muguerza, E.; Redin Iroz, A.; Omarini, A.; Conde, E.; Alfaro, M.; Castanera, R.; Santoyo, F.; Ramírez, L.; Pisabarro, A.G. Induction of laccase activity in the white rot fungus Pleurotus ostreatus using water polluted with wheat straw extracts. Bioresour. Technol. 2013, 133, 142–149. [Google Scholar] [CrossRef] [PubMed]
- van Kuijk, S.J.A.; del Río, J.C.; Rencoret, J.; Gutiérrez, A.; Sonnenberg, A.S.M.; Baars, J.J.P.; Hendriks, W.H.; Cone, J.W. Selective ligninolysis of wheat straw and wood chips by the white-rot fungus Lentinula edodes and its influence on in vitro rumen degradability. J. Anim. Sci. Biotechnol. 2016, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouches, E.; Herpoël-Gimbert, I.; Steyer, J.P.; Carrere, H. Improvement of anaerobic degradation by white-rot fungi pretreatment of lignocellulosic biomass: A review. Renew. Sustain. Energy Rev. 2016, 59, 179–198. [Google Scholar] [CrossRef]
- Hendriks, A.T.W.M.; Zeeman, G. Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresour. Technol. 2009, 100, 10–18. [Google Scholar] [CrossRef]
- Taherzadeh, M.; Karimi, K. Pretreatment of Lignocellulosic Wastes to Improve Ethanol and Biogas Production: A Review. Int. J. Mol. Sci. 2008, 9, 1621–1651. [Google Scholar] [CrossRef] [Green Version]
- Bilal, M.; Iqbal, H.M.N. Ligninolysis Potential of Ligninolytic Enzymes: A Green and Sustainable Approach to Bio-transform Lignocellulosic Biomass into High-Value Entities. In Alternative Energy Resources; Pathak, P., Srivastava, R.R., Eds.; Springer International Publishing: Cham, Switzerland, 2020; Volume 99, pp. 151–171. ISBN1 978-3-030-57922-7. ISBN2 978-3-030-57922-4. [Google Scholar]
- Vivekanand, V.; Dwivedi, P.; Sharma, A.; Sabharwal, N.; Singh, R.P. Enhanced delignification of mixed wood pulp by Aspergillus fumigatus laccase mediator system. World J. Microbiol. Biotechnol. 2008, 24, 2799–2804. [Google Scholar] [CrossRef]
- Wan, C.; Li, Y. Effectiveness of microbial pretreatment by Ceriporiopsis subvermispora on different biomass feedstocks. Bioresour. Technol. 2011, 102, 7507–7512. [Google Scholar] [CrossRef]
- Salvachúa, D.; Prieto, A.; López-Abelairas, M.; Lu-Chau, T.; Martínez, Á.T.; Martínez, M.J. Fungal pretreatment: An alternative in second-generation ethanol from wheat straw. Bioresour. Technol. 2011, 102, 7500–7506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, T.; Ikeda, K.; Kawagishi, H.; Hirai, H. Improvement of saccharide yield from wood by simultaneous enzymatic delignification and saccharification using a ligninolytic enzyme and cellulase. J. Biosci. Bioeng. 2021, 132, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Suryadi, H.; Judono, J.J.; Putri, M.R.; Eclessia, A.D.; Ulhaq, J.M.; Agustina, D.N.; Sumiati, T. Biodelignification of lignocellulose using ligninolytic enzymes from white-rot fungi. Heliyon 2022, 8, e08865. [Google Scholar] [CrossRef] [PubMed]
- Shirkavand, E.; Baroutian, S.; Gapes, D.J.; Young, B.R. Pretreatment of radiata pine using two white rot fungal strains Stereum hirsutum and Trametes versicolor. Energy Convers. Manag. 2017, 142, 13–19. [Google Scholar] [CrossRef]
- Ntougias, S.; Baldrian, P.; Ehaliotis, C.; Nerud, F.; Merhautová, V.; Zervakis, G.I. Olive mill wastewater biodegradation potential of white-rot fungi—Mode of action of fungal culture extracts and effects of ligninolytic enzymes. Bioresour. Technol. 2015, 189, 121–130. [Google Scholar] [CrossRef]
- Kissi, M.; Mountadar, M.; Assobhei, O.; Gargiulo, E.; Palmieri, G.; Giardina, P.; Sannia, G. Roles of two white-rot basidiomycete fungi in decolorisation and detoxification of olive mill waste water. Appl. Microbiol. Biotechnol. 2001, 57, 221–226. [Google Scholar] [CrossRef]
- Cui, T.; Yuan, B.; Guo, H.; Tian, H.; Wang, W.; Ma, Y.; Li, C.; Fei, Q. Enhanced lignin biodegradation by consortium of white rot fungi: Microbial synergistic effects and product mapping. Biotechnol. Biofuels 2021, 14, 162. [Google Scholar] [CrossRef]
- Hermosilla, E.; Rubilar, O.; Schalchli, H.; da Silva, A.S.; Ferreira-Leitao, V.; Diez, M.C. Sequential white-rot and brown-rot fungal pretreatment of wheat straw as a promising alternative for complementary mild treatments. Waste Manag. 2018, 79, 240–250. [Google Scholar] [CrossRef]
- Serrano, A.; Villa-Gomez, D.; Fermoso, F.G.; Alonso-Fariñas, B. Is anaerobic digestion a feasible alternative to the combustion of olive mill solid waste in terms of energy production? A critical review. Biofuels Bioprod. Biorefining 2021, 15, 150–162. [Google Scholar] [CrossRef]
- Sounni, F.; Elgnaoui, Y.; El Bari, H.; Merzouki, M.; Benlemlih, M. Effect of mixture ratio and organic loading rate during anaerobic co-digestion of olive mill wastewater and agro-industrial wastes. Biomass Convers. Biorefinery 2021, 11, 1–7. [Google Scholar] [CrossRef]
- Genethliou, C.; Kornaros, M.; Dailianis, S. Biodegradation of olive mill wastewater phenolic compounds in a thermophilic anaerobic upflow packed bed reactor and assessment of their toxicity in digester effluents. J. Environ. Manag. 2020, 255, 109882. [Google Scholar] [CrossRef] [PubMed]
- Caroca, E.; Serrano, A.; Borja, R.; Jiménez, A.; Carvajal, A.; Braga, A.F.M.; Rodriguez-Gutierrez, G.; Fermoso, F.G. Influence of phenols and furans released during thermal pretreatment of olive mill solid waste on its anaerobic digestion. Waste Manag. 2021, 120, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.; Fermoso, F.G.; Alonso-Fariñas, B.; Rodríguez-Gutierrez, G.; Fernandez-Bolaños, J.; Borja, R. Olive mill solid waste biorefinery: High-temperature thermal pre-treatment for phenol recovery and biomethanization. J. Clean. Prod. 2017, 148, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Prior, Á.; Trujillo-Reyes, Á.; Serrano, A.; Rodríguez-Gutiérrez, G.; Reinhard, C.; Fermoso, F.G. Biogas Potential of the Side Streams Obtained in a Novel Phenolic Extraction System from Olive Mill Solid Waste. Molecules 2020, 25, 5438. [Google Scholar] [CrossRef]
- Sampedro, I.; Cajthaml, T.; Marinari, S.; Stazi, S.R.; Grego, S.; Petruccioli, M.; Federici, F.; D’Annibale, A. Immobilized Inocula of White-Rot Fungi Accelerate both Detoxification and Organic Matter Transformation in Two-Phase Dry Olive-Mill Residue. J. Agric. Food Chem. 2009, 57, 5452–5460. [Google Scholar] [CrossRef] [Green Version]
- Zerva, A.; Zervakis, G.I.; Christakopoulos, P.; Topakas, E. Degradation of olive mill wastewater by the induced extracellular ligninolytic enzymes of two wood-rot fungi. J. Environ. Manag. 2017, 203, 791–798. [Google Scholar] [CrossRef]
- Schalchli, H.; Hormazábal, E.; Rubilar, O.; Briceño, G.; Mutis, A.; Zocolo, G.J.; Diez, M.C. Production of ligninolytic enzymes and some diffusible antifungal compounds by white-rot fungi using potato solid wastes as the sole nutrient source. J. Appl. Microbiol. 2017, 123, 886–895. [Google Scholar] [CrossRef]
- Rubilar, O.; Tortella, G.; Cea, M.; Acevedo, F.; Bustamante, M.; Gianfreda, L.; Diez, M.C. Bioremediation of a Chilean Andisol contaminated with pentachlorophenol (PCP) by solid substrate cultures of white-rot fungi. Biodegradation 2011, 22, 31–41. [Google Scholar] [CrossRef]
- Tortella, G.R.; Rubilar, O.; Gianfreda, L.; Valenzuela, E.; Diez, M.C. Enzymatic characterization of Chilean native wood-rotting fungi for potential use in the bioremediation of polluted environments with chlorophenols. World J. Microbiol. Biotechnol. 2008, 24, 2805–2818. [Google Scholar] [CrossRef] [Green Version]
- Rossman, A.Y. The impact of invasive fungi on agricultural ecosystems in the United States. Biol. Invasions 2009, 11, 97–107. [Google Scholar] [CrossRef]
- García, A.; Rodríguez-Juan, E.; Rodríguez-Gutiérrez, G.; Rios, J.J.; Fernández-Bolaños, J. Extraction of phenolic compounds from virgin olive oil by deep eutectic solvents (DESs). Food Chem. 2016, 197, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Fregapane, G.; Salvador, M. Fate and Prediction of Phenolic Secoiridoid Compounds throughout the Different Stages of the Virgin Olive Oil Making Process. Antioxidants 2017, 6, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, E.W.; Baird, R.B. Standard Methods for the Examination of Water and Wastewater; American Public Health Association, American Water Works Association, Water Environment Federation: Washington, DC, USA, 2012.
- Ochoa-Velasco, C.E.; Navarro-Cruz, A.R.; Vera-López, O.; Palou, E.; Avila-Sosa, R. Growth modeling to control (in vitro) Fusarium verticillioides and Rhizopus stolonifer with thymol and carvacrol. Rev. Argent. Microbiol. 2018, 50, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, F.; Pizzul, L.; Castillo, M.d.P.; Rubilar, O.; Lienqueo, M.E.; Tortella, G.; Diez, M.C. A practical culture technique for enhanced production of manganese peroxidase by Anthracophyllum discolor Sp4. Braz. Arch. Biol. Technol. 2011, 54, 1175–1186. [Google Scholar] [CrossRef] [Green Version]
- Wariishi, H.; Valli, K.; Gold, M.H. Manganese(II) oxidation by manganese peroxidase from the basidiomycete Phanerochaete chrysosporium. Kinetic mechanism and role of chelators. J. Biol. Chem. 1992, 267, 23688–23695. [Google Scholar] [CrossRef]
- Kaouachi, A.; Amane, M.; Jaafari, S. El Biovalorisation of Two-Phase Olive Mill Waste by Composting Process. Int. J. Sci. Res. 2015, 6, 5. [Google Scholar]
- Fernández-Rodríguez, M.J.; de la Lama-Calvente, D.; Jiménez-Rodríguez, A.; Borja, R.; Rincón-Llorente, B. Influence of the cell wall of Chlamydomonas reinhardtii on anaerobic digestion yield and on its anaerobic co-digestion with a carbon-rich substrate. Process Saf. Environ. Prot. 2019, 128, 167–175. [Google Scholar] [CrossRef]
- Messineo, A.; Maniscalco, M.P.; Volpe, R. Biomethane recovery from olive mill residues through anaerobic digestion: A review of the state of the art technology. Sci. Total Environ. 2020, 703, 135508. [Google Scholar] [CrossRef]
- Bustamante, M.; González, M.E.; Cartes, A.; Diez, M.C. Effect of soya lecithin on the enzymatic system of the white-rot fungi Anthracophyllum discolor. J. Ind. Microbiol. Biotechnol. 2011, 38, 189–197. [Google Scholar] [CrossRef]
- Jain, M.S.; Paul, S.; Kalamdhad, A.S. Recalcitrant carbon for composting of fibrous aquatic waste: Degradation kinetics, spectroscopic study and effect on physico-chemical and nutritional properties. J. Environ. Manag. 2019, 251, 109568. [Google Scholar] [CrossRef]
- Gill, P.; Arora, D. Effect of culture conditions on manganese peroxidase production and activity by some white rot fungi. J. Ind. Microbiol. Biotechnol. 2003, 30, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Papinutti, V.L.; Diorio, L.A.; Forchiassin, F. Production of laccase and manganese peroxidase by Fomes sclerodermeus grown on wheat bran. J. Ind. Microbiol. Biotechnol. 2003, 30, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Tortella, G.R.; Diez, M.C.; Durán, N. Fungal Diversity and Use in Decomposition of Environmental Pollutants. Crit. Rev. Microbiol. 2005, 31, 197–212. [Google Scholar] [CrossRef]
- Koranda, M.; Kaiser, C.; Fuchslueger, L.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S.; Richter, A. Fungal and bacterial utilization of organic substrates depends on substrate complexity and N availability. FEMS Microbiol. Ecol. 2014, 87, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Del Río, J.C.; Rencoret, J.; Prinsen, P.; Martínez, Á.T.; Ralph, J.; Gutiérrez, A. Structural Characterization of Wheat Straw Lignin as Revealed by Analytical Pyrolysis, 2D-NMR, and Reductive Cleavage Methods. J. Agric. Food Chem. 2012, 60, 5922–5935. [Google Scholar] [CrossRef] [Green Version]
- Chmelová, D.; Legerská, B.; Kunstová, J.; Ondrejovič, M.; Miertuš, S. The production of laccases by white-rot fungi under solid-state fermentation conditions. World J. Microbiol. Biotechnol. 2022, 38, 21. [Google Scholar] [CrossRef]
- Elisashvili, V.; Kachlishvili, E.; Penninckx, M. EVect of growth substrate, method of fermentation, and nitrogen source on lignocellulose-degrading enzymes production by white-rot basidiomycetes. J. Ind. Microbiol. Biotechnol. 2008, 35, 1531–1538. [Google Scholar] [CrossRef]
- González Bautista, E.; Gutierrez, E.; Dupuy, N.; Gaime-Perraud, I.; Ziarelli, F.; Farnet da Silva, A.-M. Pre-treatment of a sugarcane bagasse-based substrate prior to saccharification: Effect of coffee pulp and urea on laccase and cellulase activities of Pycnoporus sanguineus. J. Environ. Manag. 2019, 239, 178–186. [Google Scholar] [CrossRef]
- Torres-Mancera, M.T.; Cordova-López, J.; Rodríguez-Serrano, G.; Roussos, S.; Ramírez-Coronel, M.A.; Favela-Torres, E.; Saucedo-Castañeda, G. Enzymatic Extraction of Hydroxycinnamic Acids from Coffee Pulp. Food Technol. Biotechnol. 2011, 49, 369–373. [Google Scholar]
- Tien, M.; Myer, S.B. Selection and characterization of mutants of Phanerochaete chrysosporium Exhibiting Ligninolytic activity under nutrient-rich conditions. Appl. Environ. Microbiol. 1990, 56, 2540–2544. [Google Scholar] [CrossRef] [Green Version]
- Xiaoping, X.; Xianghua, W.E.N.; Yanan, B.A.I.; Yi, Q. Effects of culture conditions on ligninolytic enzymes and protease production by Phanerochaete chrysosporium in air. J. Environ. Sci. 2008, 20, 94–100. [Google Scholar]
- Rodríguez-Couto, S. Industrial and environmental applications of white-rot fungi. Mycosphere 2017, 8, 456–466. [Google Scholar] [CrossRef]
- Wu, F.; Ozaki, H.; Terashima, Y.; Imada, T.; Ohkouchi, Y. Activities of ligninolytic enzymes of the white rot fungus, phanerochaete chrysosporium and its recalcitrant substance degradability. Wat. Sci. Tech. 1996, 34, 69–78. [Google Scholar] [CrossRef]
- Gassara, F.; Ajila, C.M.; Brar, S.K.; Verma, M.; Tyagi, R.D.; Valero, J.R. Liquid state fermentation of apple pomace sludge for the production of ligninolytic enzymes and liberation of polyphenolic compounds. Process Biochem. 2012, 47, 999–1004. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, K. Lignin-Degrading Enzyme from the Hymenomycete Phanerochaete chrysosporium Burds. Science 1983, 221, 661–663. [Google Scholar] [CrossRef] [Green Version]
- Ansari, Z.; Karimi, A.; Ebrahimi, S.; Emami, E. Improvement in ligninolytic activity of Phanerochaete chrysosporium cultures by glucose oxidase. Biochem. Eng. J. 2016, 105, 332–338. [Google Scholar] [CrossRef]
- Knežević, A.; Stajić, M.; Vukojević, J.; Milovanović, I. The effect of trace elements on wheat straw degradation by Trametes gibbosa. Int. Biodeterior. Biodegrad. 2014, 96, 152–156. [Google Scholar] [CrossRef]
- Reina, R.; Liers, C.; Ocampo, J.A.; García-Romera, I.; Aranda, E. Solid state fermentation of olive mill residues by wood- and dung-dwelling Agaricomycetes: Effects on peroxidase production, biomass development and phenol phytotoxicity. Chemosphere 2013, 93, 1406–1412. [Google Scholar] [CrossRef]
- Fenice, M.; D’Annibale, A. Submerged and solid-state production of laccase and Mn-peroxidase by Panus tigrinus on olive mill wastewater-based media. J. Biotechnol. 2003, 100, 77–85. [Google Scholar] [CrossRef]
- Mishra, V.; Jana, A.K.; Jana, M.M.; Gupta, A. Enhancement in multiple lignolytic enzymes production for optimized lignin degradation and selectivity in fungal pretreatment of sweet sorghum bagasse. Bioresour. Technol. 2017, 236, 49–59. [Google Scholar] [CrossRef]
- Atilano-Camino, M.M.; Álvarez-Valencia, L.H.; García-González, A.; García-Reyes, R.B. Improving laccase production from Trametes versicolor using lignocellulosic residues as cosubstrates and evaluation of enzymes for blue wastewater biodegradation. J. Environ. Manag. 2020, 275, 111231. [Google Scholar] [CrossRef]
- Liang, S.; McDonald, A.G. Chemical and Thermal Characterization of Potato Peel Waste and Its Fermentation Residue as Potential Resources for Biofuel and Bioproducts Production. J. Agric. Food Chem. 2014, 62, 8421–8429. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.; Fermoso, F.G.; Alonso-Fariñas, B.; Rodríguez-Gutierrez, G.; Fernandez-Bolaños, J.; Borja, R. Phenols recovery after steam explosion of Olive Mill Solid Waste and its influence on a subsequent biomethanization process. Bioresour. Technol. 2017, 243, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linares, A.; Caba, J.M.; Ligero, F.; De la Rubia, T.; Martínez, J. Detoxification of semisolid olive-mill wastes and pine-chip mixtures using Phanerochaete flavido-alba. Chemosphere 2003, 51, 887–891. [Google Scholar] [CrossRef]
- Sampedro, I.; Marinari, S.; D’Annibale, A.; Grego, S.; Ocampo, J.A.; García-Romera, I. Organic matter evolution and partial detoxification in two-phase olive mill waste colonized by white-rot fungi. Int. Biodeterior. Biodegrad. 2007, 60, 116–125. [Google Scholar] [CrossRef]
- Khdair, A.; Abu-Rumman, G. Sustainable Environmental Management and Valorization Options for Olive Mill Byproducts in the Middle East and North Africa (MENA) Region. Processes 2020, 8, 671. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
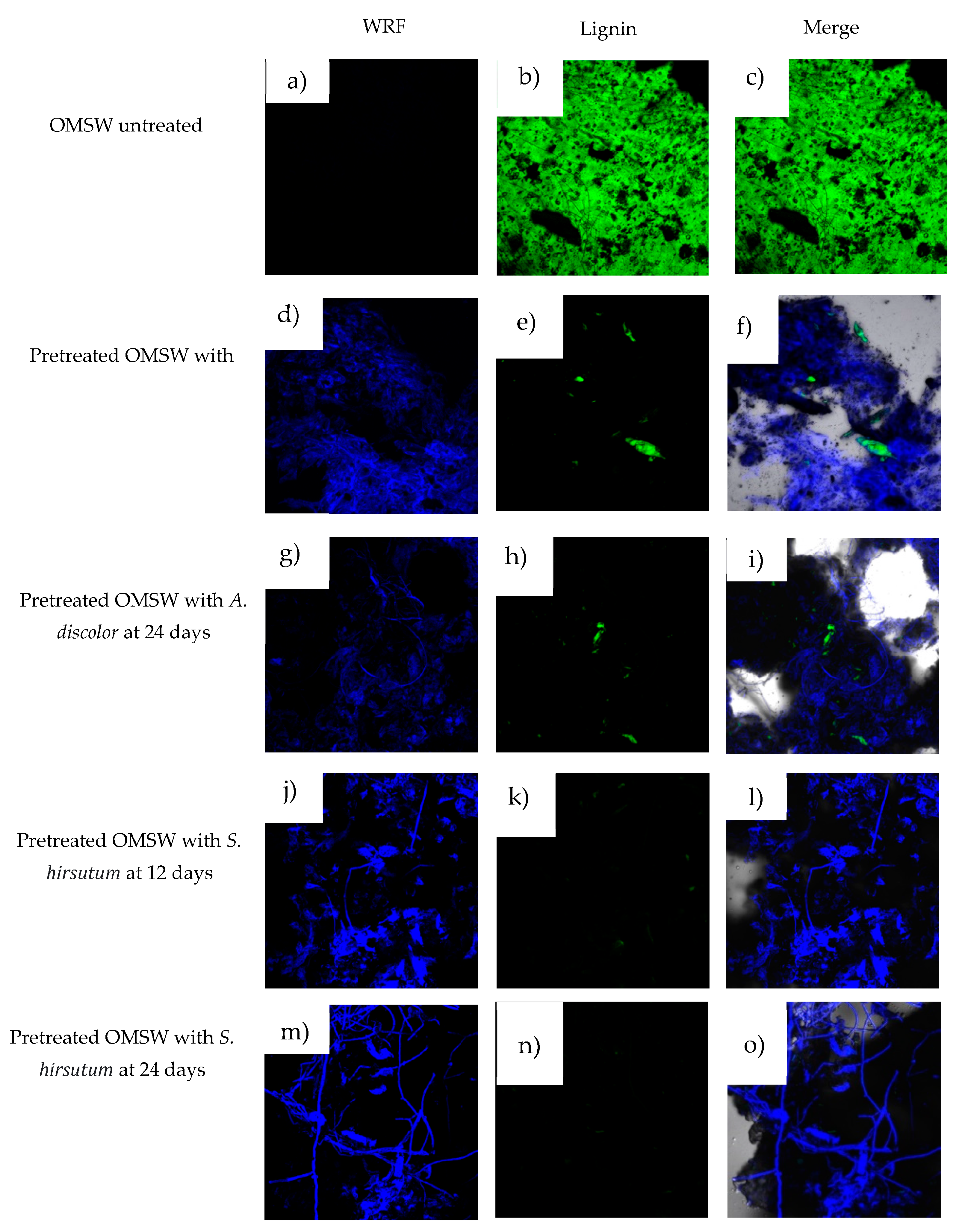{kind=link}
| Strain | Culture Media |
|---|---|
| S. hirsutum | 100 mL MKM |
| A. discolor | 100 mL MKM |
| S. hirsutum | 100 mL MKM + 30 g OMSW |
| A. discolor | 100 mL MKM + 30 g OMSW |
| S. hirsutum | 100 mL distilled water + 30 g OMSW |
| A. discolor | 100 mL distilled water + 30 g OMSW |
| Characteristics | Value |
|---|---|
| α-cellulose (% w/w) | 35.2 ± 3 |
| Lignin (% w/w) | 33.4 ± 4 |
| Hemicellulose (% w/w) | 45.5 ± 8 |
| Elemental analysis (C/N) (% w/w) | 48.4/0.84 |
| Ash (% w/w) | 2.96 |
| Total nitrogen (mg/kg) | 134.2 |
| Nitrate (mg/kg) | <1.0 |
| Nitrite (mg/kg) | <0.6 |
| Total copper (mg/kg) | 8.5 |
| Total iron (mg/kg) | <140.0 |
| Total manganese (mg/kg) | <13.0 |
| Chemical oxygen demand (mg O2/L) | 87.9 |
| Total solids (% w/w) | 38.5 |
| Volatile solids (% of TS) | 89.7 |
| pH | 5.08 |
| Total polyphenols (mg GA/100 g) | 149.0 |
| Antioxidant capacity (mg Trolox/L) | 214.4 |
| Gallic acid hexoside (mg/100 g) | 0.9 |
| Gallic acid protocatechuic acid hexoside (mg/100 g) | 0.5 |
| Hexoside ferulic acid (mg/100 g) | 0.3 |
| Cautaric acid (mg/100 g) | 1.0 |
| Caftaric acid (mg/100 g) | 0.9 |
| Catechin (mg/100 g) | 1.6 |
| Epicatechin (mg/100 g) | 1.9 |
| Quercetin-3-rutinoside (mg/100 g) | 1.3 |
| Quercetin-3-hexoside (mg/100 g) | 1.7 |
| Quercetin-3-glucuronide (mg/100 g) | 2.0 |
| Kaempferol-3-glucoside (mg/100 g) | 3.5 |
| Kaempferol-3-hexoside (mg/100 g) | 10.2 |
| Lipids (% w/w) | 13.23 |
| Strain | PDA+RBBR (Decoloration) | PDA+ABTS (Coloration) |
|---|---|---|
| A. discolor | ++++ | ++++ |
| S. hirsutum | ++ | ++++ |
| Culture Conditions | Maximum Mycelial Diameter (mm) (Amax) | Mycelial Growth Rate (mm/day) (rmax) | Lag Phase (d) (λ) | e | R2 |
|---|---|---|---|---|---|
| PDA medium | |||||
| A. discolor | 100 ± 2 | 17.9 ± 0.5 | 1.2 ± 0.1 | 2.7183 | 0.9968 |
| S. hirsutum | 102 ± 5 | 16.8 ± 1 | 1.0 ± 0.2 | 2.7183 | 0.9838 |
| OMSW | |||||
| A. discolor | 78 ± 39 | 2.5 ± 0.5 | 1.6 ± 1.9 | 2.7183 | 0.9585 |
| S. hirsutum | 212 ± 46 | 9.7 ± 0.7 | 3.0 ± 0.6 | 2.7183 | 0.9857 |
| PDA and OMSW | |||||
| A. discolor | 103 ± 4 | 12.1 ± 0.5 | 1.3 ± 0.1 | 2.7183 | 0.9864 |
| S. hirsutum | 100 ± 3 | 14.3 ± 0.9 | 1.4 ± 0.2 | 2.7183 | 0.9793 |
| Strain | Phenolic Compounds Removal (%) | Reaction Time (days) | Specific Conditions | OMSW (g)/Water (mL) Proportion | References | |
|---|---|---|---|---|---|---|
| Temperature (°C) | Others | |||||
| Phanerochaete flavido-alba | 70 | 60 | 30 | Aeration: sterile O2 (3 L/min for 1 min) every 24 h | 1:0.3 | [66] |
| P. radiata | 95.8 | 140 | 30 | - | 1:0.1 | [67] |
| C. rigida | 89.2 | |||||
| P. cinnabarinus | 88.7 | |||||
| Phlebiasp. | 85 | 30 | 28 | Immobilization in Polyurethane sponge | 1:0.1 | [26] |
| Panus tigrinus | 36 | |||||
| Phlebiasp. | 43 | |||||
| A. auricula-judae | 60 | 28 | 24 | - | 1:4.5 | [60] |
| B. adusta | 80 | |||||
| C. radians | 75 | |||||
| A discolor | 90 | 24 | 25 | - | 1:3.3 | This work |
| S. hirsutum | 85 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benavides, V.; Pinto-Ibieta, F.; Serrano, A.; Rubilar, O.; Ciudad, G. Use of Anthracophyllum discolor and Stereum hirsutum as a Suitable Strategy for Delignification and Phenolic Removal of Olive Mill Solid Waste. Foods 2022, 11, 1587. https://doi.org/10.3390/foods11111587
Benavides V, Pinto-Ibieta F, Serrano A, Rubilar O, Ciudad G. Use of Anthracophyllum discolor and Stereum hirsutum as a Suitable Strategy for Delignification and Phenolic Removal of Olive Mill Solid Waste. Foods. 2022; 11(11):1587. https://doi.org/10.3390/foods11111587
Chicago/Turabian StyleBenavides, Viviana, Fernanda Pinto-Ibieta, Antonio Serrano, Olga Rubilar, and Gustavo Ciudad. 2022. "Use of Anthracophyllum discolor and Stereum hirsutum as a Suitable Strategy for Delignification and Phenolic Removal of Olive Mill Solid Waste" Foods 11, no. 11: 1587. https://doi.org/10.3390/foods11111587