
Survival of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) and Herpes Simplex Virus 1 (HSV-1) on Foods Stored at Refrigerated Temperature


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Food Sources and Sample Preparation
2.3. Virus Inoculation on Foods
2.4. Plaque Assay
2.5. Viral Genome Extraction, Quantitative Polymerase Chain Reaction (qPCR), and Reverse Transcription PCR (RT-PCR)
2.6. Statistical Analysis
3. Results
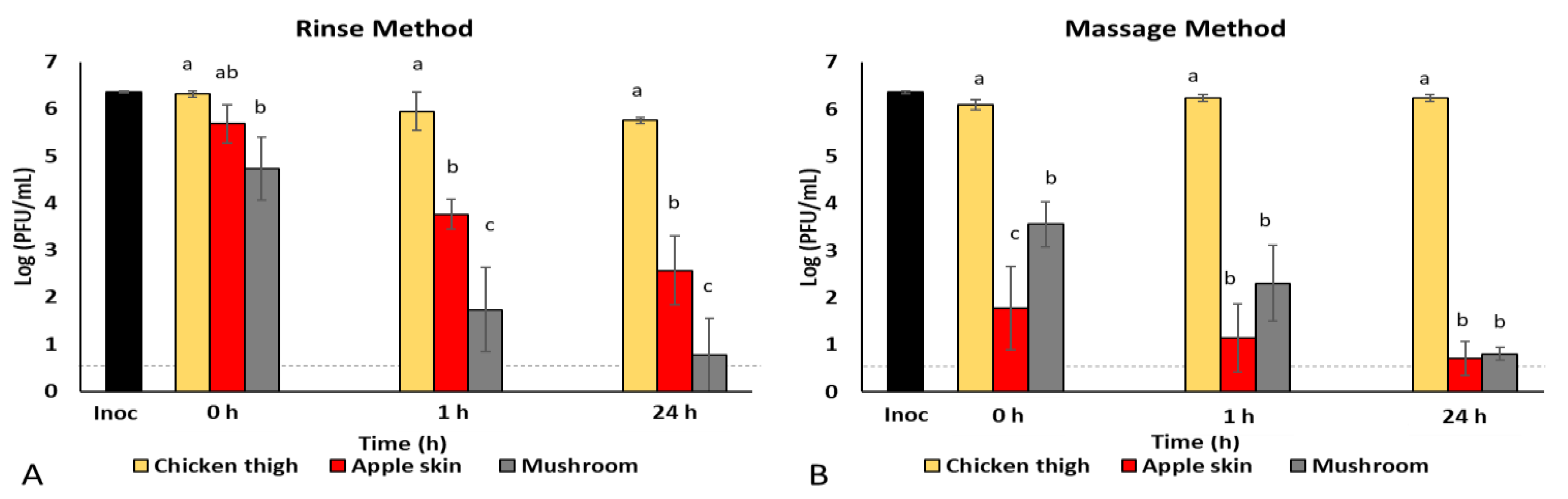
3.1. Rinse vs. Massage Method Comparison for Virus Recovery from Foods
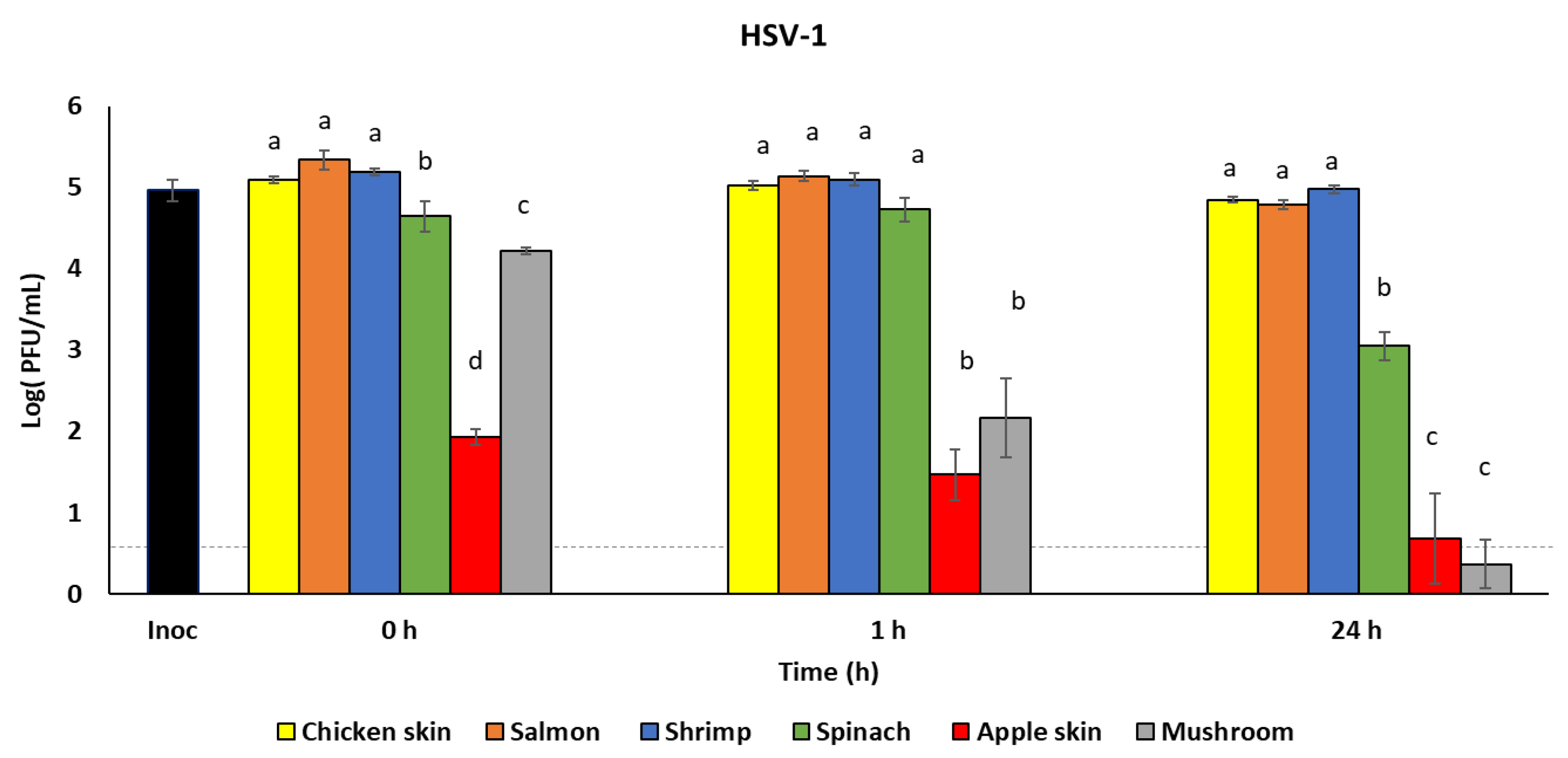
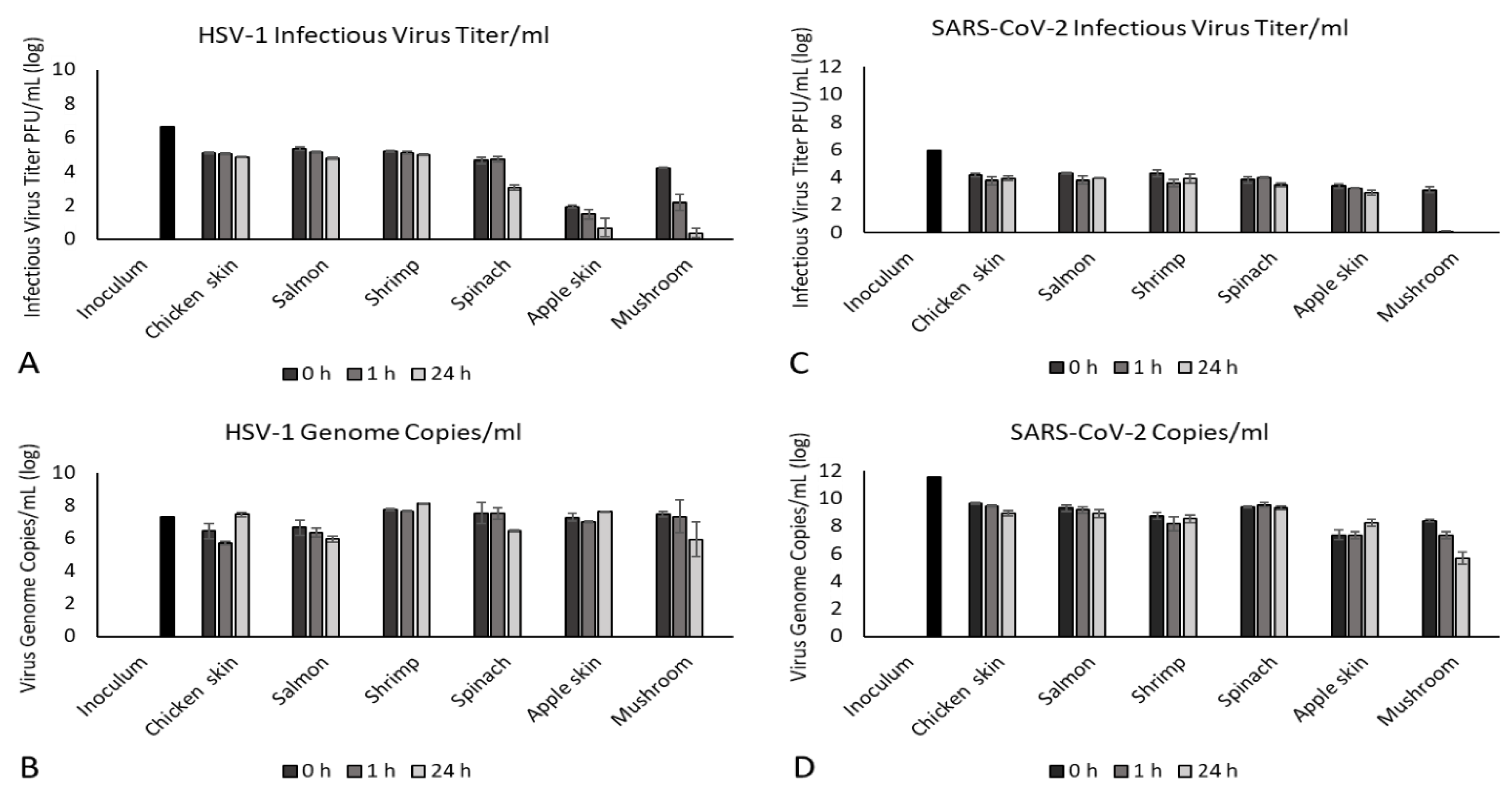
3.2. Survival of HSV-1 on Foods
3.3. Survival of SARS-CoV-2 on Foods
3.4. Relation of Infectious Virus Titer to Viral Genome Copy Number
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhand, R.; Li, J. Coughs and Sneezes: Their Role in Transmission of Respiratory Viral Infections, Including SARS-CoV-2. Am. J. Respir. Crit. Care Med. 2020, 202, 651–659. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Ji, M.; Pei, F.; Zhao, Q.; Zhou, Y.; Hong, Y.; Han, S.; Wang, J.; Wang, Q.; et al. Transmission Routes Analysis of SARS-CoV-2: A Systematic Review and Case Report. Front. Cell Dev. Biol. 2020, 8, 618. [Google Scholar] [CrossRef]
- Hamner, L.; Dubbel, P.; Capron, I.; Ross, A.; Jordan, A.; Lee, J.; Lynn, J.; Ball, A.; Narwal, S.; Russell, S.; et al. High SARS-CoV-2 Attack Rate Following Exposure at a Choir Practice—Skagit County, Washington, March 2020. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 606–610. [Google Scholar] [CrossRef]
- Groves, L.M.; Usagawa, L.; Elm, J.; Low, E.; Manuzak, A.; Quint, J.; Center, K.E.; Buff, A.M.; Kemble, S.K. Community Transmission of SARS-CoV-2 at Three Fitness Facilities—Hawaii, June–July 2020. MMWR. Morb. Mortal. Wkly. Rep. 2021, 70, 316–320. [Google Scholar] [CrossRef]
- Kang, M.; Wei, J.; Yuan, J.; Guo, J.; Zhang, Y.; Hang, J.; Qu, Y.; Qian, H.; Zhuang, Y.; Chen, X.; et al. Probable Evidence of Fecal Aerosol Transmission of SARS-CoV-2 in a High-Rise Building. Ann. Intern. Med. 2020, 173, 974–980. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Yuan, S.; Kok, K.-H.; To, K.K.-W.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.-Y.; Poon, R.W.-S.; et al. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: A study of a family cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Van Doremalen, N.; Bushmaker, T.; Lloyd-Smith, J.O.; De Wit, E.; Munster, V.J.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Chin, A.W.H.; Chu, J.T.S.; A Perera, M.R.; Hui, K.P.Y.; Yen, H.-L.; Chan, M.C.W.; Peiris, M.; Poon, L.L.M. Stability of SARS-CoV-2 in different environmental conditions. Lancet Microbe 2020, 1, e10. [Google Scholar] [CrossRef]
- Anelich, L.E.C.M.; Lues, R.; Farber, J.M.; Parreira, V.R. SARS-CoV-2 and Risk to Food Safety. Front. Nutr. 2020, 7, 580551. [Google Scholar] [CrossRef]
- Whitworth, J. China Reports Further Food-Related Coronavirus Findings. Available online: https://www.foodsafetynews.com/2020/11/china-reports-further-food-related-coronavirus-findings/ (accessed on 17 March 2021).
- Associated Press. Chinese City Reports Coronavirus Found on Ice Cream. Available online: https://apnews.com/article/beijing-tianjin-coronavirus-pandemic-wuhan-china-a75ec51c11338190c483c78d65c3348d (accessed on 17 March 2021).
- Pang, X.; Ren, L.; Wu, S.; Ma, W.; Yang, J.; Di, L.; Li, J.; Xiao, Y.; Kang, L.; Du, S.; et al. Cold-chain food contamination as the possible origin of COVID-19 resurgence in Beijing. Natl. Sci. Rev. 2020, 7, 1861–1864. [Google Scholar] [CrossRef]
- Oakenfull, R.J.; Wilson, A.J. Qualitative Risk Assessment: What Is the Risk of Food or Food Contact Materials Being a Source or Transmission Route of SARS-CoV-2 for UK Consumers. Available online: https://www.food.gov.uk/research/research-projects/qualitative-risk-assessment-on-the-risk-of-food-or-food-contact-materials-as-a-transmission-route-for-sars-cov-2 (accessed on 17 March 2021).
- Food and Drug Administration (FDA). Food Safety and the Coronavirus Disease 2019 (COVID-19). Available online: https://www.fda.gov/food/food-safety-during-emergencies/food-safety-and-coronavirus-disease-2019-covid-19 (accessed on 17 March 2021).
- World Health Organization (WHO). COVID-19 and Food Safety: Guidance for Food Businesses: Interim Guidance. Available online: https://www.who.int/publications/i/item/covid-19-and-food-safety-guidance-for-food-businesses (accessed on 17 March 2021).
- European Food Safety Authority (EFSA). Coronavirus: No Evidence That Food Is a Source or Transmission Route. Available online: https://www.efsa.europa.eu/en/news/coronavirus-no-evidence-food-source-or-transmission-route (accessed on 17 March 2021).
- Dyal, J.W.; Grant, M.P.; Broadwater, K.; Bjork, A.; Waltenburg, M.A.; Gibbins, J.D.; Hale, C.; Silver, M.; Fischer, M.; Steinberg, J.; et al. COVID-19 Among Workers in Meat and Poultry Processing Facilities—19 States, April 2020. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 69. [Google Scholar] [CrossRef]
- Waltenburg, M.A.; Victoroff, T.; Rose, C.E.; Butterfield, M.; Jervis, R.H.; Fedak, K.M.; Gabel, J.A.; Feldpausch, A.; Dunne, E.M.; Austin, C.; et al. Update: COVID-19 Among Workers in Meat and Poultry Processing Facilities—United States, April–May 2020. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Donahue, M.; Sreenivasan, N.; Stover, D.; Rajasingham, A.; Watson, J.; Bealle, A.; Ritchison, N.; Safranek, T.; Waltenburg, M.A.; Buss, B.; et al. Notes from the Field: Characteristics of Meat Processing Facility Workers with Confirmed SARS-CoV-2 Infection—Nebraska, April–May 2020. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 1020–1022. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, J.; Kennedy, E.D.; Basler, C.; Grant, M.P.; Jacobs, J.R.; Ortbahn, D.; Osburn, J.; Saydah, S.; Tomasi, S.; Clayton, J.L. COVID-19 Outbreak Among Employees at a Meat Processing Facility—South Dakota, March–April 2020. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 1015–1019. [Google Scholar] [CrossRef]
- Food & Environment Reporting Network (FERN). Mapping Covid-19 Outbreaks in the Food System. Available online: https://thefern.org/2020/04/mapping-covid-19-in-meat-and-food-processing-plants/ (accessed on 11 March 2021).
- He, X.; Lau, E.H.Y.; Wu, P.; Deng, X.; Wang, J.; Hao, X.; Lau, Y.C.; Wong, J.Y.; Guan, Y.; Tan, X.; et al. Temporal dynamics in viral shedding and transmissibility of COVID-19. Nat. Med. 2020, 26, 672–675. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Yao, L.; Wei, T.; Tian, F.; Jin, D.Y.; Chen, L.; Wang, M. Presumed Asymptomatic Carrier Transmission of COVID-19. JAMA 2020, 323, 1406–1407. [Google Scholar] [CrossRef] [Green Version]
- Arons, M.M.; Hatfield, K.M.; Reddy, S.C.; Kimball, A.; James, A.; Jacobs, J.R.; Taylor, J.; Spicer, K.; Bardossy, A.C.; Oakley, L.P.; et al. Presymptomatic SARS-CoV-2 Infections and Transmission in a Skilled Nursing Facility. N. Engl. J. Med. 2020, 382, 2081–2090. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, G.; Huang, Y.-W. Modeling the load of SARS-CoV-2 virus in human expelled particles during coughing and speaking. PLOS ONE 2020, 15, e0241539. [Google Scholar] [CrossRef] [PubMed]
- Ming, Z.; Han, S.; Deng, K.; Reyes, E.; Ha, Y.; Kim, S.; Zhao, Y.; Dobritsa, A.; Wu, M.; Zhang, D.; et al. Prevalence of SARS-CoV-2 Contamination on Food Plant Surfaces as Determined by Environmental Monitoring. J. Food Prot. 2021, 84, 352–358. [Google Scholar] [CrossRef]
- Meng, X.-J.; Liang, T. SARS-CoV-2 infection in the gastrointestinal tract: Fecal-oral route of transmission for COVID-19? Gastroenterology 2021. [Google Scholar] [CrossRef]
- Jiao, L.; Li, H.; Xu, J.; Yang, M.; Ma, C.; Li, J.; Zhao, S.; Wang, H.; Yang, Y.; Yu, W. The gastrointestinal tract is an alternative route for SARS-CoV-2 infection in a nonhuman primate model. Gastroenterology 2020. [Google Scholar] [CrossRef]
- Bardell, D. Survival of Herpes Simplex Virus Type 1 on Some Common Foods Routinely Touched before Consumption. J. Food Prot. 1997, 60, 1259–1261. [Google Scholar] [CrossRef]
- Sanglay, G.C.; Li, J.; Uribe, R.M.; Lee, K. Electron-Beam Inactivation of a Norovirus Surrogate in Fresh Produce and Model Systems. J. Food Prot. 2011, 74, 1155–1160. [Google Scholar] [CrossRef]
- A Boxman, I.L.; Tilburg, J.J.H.C.; Loeke, N.A.J.M.T.; Vennema, H.; Jonker, K.; De Boer, E.; Koopmans, M. Detection of noroviruses in shellfish in the Netherlands. Int. J. Food Microbiol. 2006, 108, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Yanez, A.A.; Harrell, T.; Sriranganathan, H.J.; Ives, A.M.; Bertke, A.S. Neurotrophic Factors NGF, GDNF and NTN Selectively Modulate HSV1 and HSV2 Lytic Infection and Reactivation in Primary Adult Sensory and Autonomic Neurons. Pathogens 2017, 6, 5. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). CDC 2019-Novel Coronavirus (2019-nCoV) Real-Time RT-PCR Diagnostic Panel. Available online: https://www.fda.gov/media/134922/download (accessed on 11 March 2021).
- Le Guyader, F.S.; Loisy, F.; Atmar, R.L.; Hutson, A.M.; Estes, M.K.; Ruvoën-Clouet, N.; Pommepuy, M.; Le Pendu, J. Norwalk Virus–specific Binding to Oyster Digestive Tissues. Emerg. Infect. Dis. 2006, 12, 931–936. [Google Scholar] [CrossRef]
- Di Girolamo, R.; Liston, J.; Matches, J. Ionic bonding, the mechanism of viral uptake by shellfish mucus. Appl. Environ. Microbiol. 1977, 33, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Hirneisen, K.A.; Kniel, K.E. Norovirus Attachment. Food Protection Trends 2013, 33, 290–299. [Google Scholar]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Smith, E.C.; Case, J.B.; Feng, J.Y.; Jordan, R.; et al. Coronavirus Susceptibility to the Antiviral Remdesivir (GS-5734) Is Mediated by the Viral Polymerase and the Proofreading Exoribonuclease. mBio 2018, 9, e00221-18. [Google Scholar] [CrossRef] [Green Version]
- Peck, K.M.; Lauring, A.S. Complexities of Viral Mutation Rates. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Zhong, P.; Xu, J.; Yang, D.; Shen, Y.; Wang, L.; Feng, Y.; Du, C.; Song, Y.; Wu, C.; Hu, X.; et al. COVID-19-associated gastrointestinal and liver injury: Clinical features and potential mechanisms. Signal Transduct. Target. Ther. 2020, 5, 1–8. [Google Scholar] [CrossRef]
- Cheung, K.S.; Hung, I.F.N.; Chan, P.P.Y.; Lung, K.C.; Tso, E.; Liu, R.; Ng, Y.Y.; Chu, M.Y.; Chung, T.W.H.; Tam, A.R.; et al. Gastrointestinal Manifestations of SARS-CoV-2 Infection and Virus Load in Fecal Samples From a Hong Kong Cohort: Systematic Review and Meta-analysis. Gastroenterology 2020, 159, 81–95. [Google Scholar] [CrossRef]
- Chan, K.-H.; Sridhar, S.; Zhang, R.; Chu, H.; Fung, A.-F.; Chan, G.; Chan, J.-W.; To, K.-W.; Hung, I.-N.; Cheng, V.-C.; et al. Factors affecting stability and infectivity of SARS-CoV-2. J. Hosp. Infect. 2020, 106, 226–231. [Google Scholar] [CrossRef]
- Dressman, J.B.; Berardi, R.R.; Dermentzoglou, L.C.; Russell, T.L.; Schmaltz, S.P.; Barnett, J.L.; Jarvenpaa, K.M. Upper Gastrointestinal (GI) pH in Young, Healthy Men and Women. Pharm. Res. 1990, 7, 756–761. [Google Scholar] [CrossRef]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS Coronavirus Spike Receptor-Binding Domain Complexed with Receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; Van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Pastorino, B.; Touret, F.; Gilles, M.; De Lamballerie, X.; Charrel, R.N. Prolonged Infectivity of SARS-CoV-2 in Fomites. Emerg. Infect. Dis. 2020, 26, 2256–2257. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization (ISO). Microbiology of the Food Chain—Horizontal Method for Determination of Hepatitis A Virus and Norovirus Using Real-Time RT-PCR—Part 1: Method for Quantification. Available online: https://www.iso.org/standard/65681.html (accessed on 13 April 2021).
- Mormann, S.; Dabisch, M.; Becker, B. Effects of Technological Processes on the Tenacity and Inactivation of Norovirus Genogroup II in Experimentally Contaminated Foods. Appl. Environ. Microbiol. 2009, 76, 536–545. [Google Scholar] [CrossRef] [Green Version]
- Fisher, D.; Reilly, A.; Zheng, A.K.E.; Cook, A.R.; Anderson, D. Seeding of outbreaks of COVID-19 by contaminated fresh and frozen food. BioRxiv 2020. [Google Scholar] [CrossRef]
- Mullis, L.; Saif, L.J.; Zhang, Y.; Zhang, X.; Azevedo, M.S. Stability of bovine coronavirus on lettuce surfaces under household refrigeration conditions. Food Microbiol. 2012, 30, 180–186. [Google Scholar] [CrossRef]
- Konowalchuk, J.; I Speirs, J. Antiviral effect of apple beverages. Appl. Environ. Microbiol. 1978, 36, 798–801. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.S.; Su, X.; D’Souza, D.H. Antiviral effects of grape seed extract against feline calicivirus, murine norovirus, and hepatitis A virus in model food systems and under gastric conditions. Food Microbiol. 2015, 52, 1–10. [Google Scholar] [CrossRef]
- Suárez, B.; Álvarez, Á.L.; García, Y.D.; del Barrio, G.; Lobo, A.P.; Parra, F. Phenolic profiles, antioxidant activity and in vitro antiviral properties of apple pomace. Food Chem. 2010, 120, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Blondin-Brosseau, M.; Harlow, J.; Doctor, T.; Nasheri, N. Examining the persistence of human Coronavirus 229E on fresh produce. Food Microbiol. 2021, 98, 103780. [Google Scholar] [CrossRef]
- Pradeep, P.; Manju, V.; Ahsan, M.F. Antiviral Potency of Mushroom Constituents. In Medicinal Mushrooms; Agrawal, D., Dhanasekaran, M., Eds.; Springer: Singapore, 2019; pp. 275–297. [Google Scholar]
- Dasgupta, A.; Acharya, K. Mushrooms: An emerging resource for therapeutic terpenoids. 3 Biotech 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Stals, A.; Baert, L.; Jasson, V.; Van Coillie, E.; Uyttendaele, M. Screening of Fruit Products for Norovirus and the Difficulty of Interpreting Positive PCR Results. J. Food Prot. 2011, 74, 425–431. [Google Scholar] [CrossRef]
- Koopmans, M.; Duizer, E. Foodborne viruses: An emerging problem. Int. J. Food Microbiol. 2004, 90, 23–41. [Google Scholar] [CrossRef]
- Batéjat, C.; Grassin, Q.; Manuguerra, J.-C.; Leclercq, I. Heat inactivation of the severe acute respiratory syndrome coronavirus 2. J. Biosaf. Biosecurity 2021, 3, 1–3. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Concentrations (log PFU/mL) over Time (Hours Post-Inoculation) | ||||||
|---|---|---|---|---|---|---|
| Rinse | Massage | |||||
| Inoculum | 8.4 ± 0.4 * | - | - | - | - | - |
| 0 h | 1 h | 24 h | 0 h | 1 h | 24 h | |
| Chicken thigh | 6.3 ± 0.1 a | 6.0 ± 0.7 a | 5.8 ± 0.1 a | 6.1 ± 0.2 A | 6.2 ± 0.1 A | 6.2 ± 0.1 A |
| Apple skin | 5.7 ± 0.7 a | 3.8 ± 0.5 b | 2.6 ± 1.3 b | 1.8 ± 1.5 A | 1.1 ± 1.2 A | 0.7 ± 0.6 A |
| Mushroom | 4.7 ± 1.2 a | 1.7 ± 1.6 b | 0.8 ± 1.3 b | 3.6 ± 0.8 A | 2.3 ± 1.4 AB | 0.8 ± 0.2 B |
| Virus Concentrations (log PFU/mL) over Time (Hours Post-Inoculation) | |||
|---|---|---|---|
| Inoculum | 6.7 ± 0.2 * | - | - |
| 0 h | 1 h | 24 h | |
| Chicken skin | 5.1 ± 0.1 a | 5.0 ± 0.1 ab | 4.9 ± 0.1 b |
| Salmon | 5.3 ± 0.2 a | 5.1 ± 0.1 ab | 4.8 ± 0.1 b |
| Shrimp | 5.2 ± 0.1 a | 5.1 ± 0.1 a | 5.0 ± 0.1 a |
| Spinach | 4.7 ± 0.3 a | 4.7 ± 0.3 a | 3.1 ± 0.3 b |
| Apple skin | 1.9 ± 0.2 a | 1.5 ± 0.5 a | 0.7 ± 1.0 a |
| Mushroom | 4.2 ± 0.1 a | 2.2 ± 0.8 b | 0.4 ± 0.5 c |
| Virus Concentrations (log PFU/mL) over Time (Hours Post-Inoculation) | |||
|---|---|---|---|
| Inoculum | 5.9 ± 0.2 * | - | - |
| 0 h | 1 h | 24 h | |
| Chicken skin | 4.2 ± 0.2 a | 3.7 ± 0.5 a | 3.9 ± 0.3 a |
| Salmon | 4.3 ± 0.1 a | 3.8 ± 0.5 a | 3.9 ± 0.1 a |
| Shrimp | 4.3 ± 0.4 a | 3.6 ± 0.4 a | 3.9 ± 0.6 a |
| Spinach | 3.8 ± 0.4 a | 4.0 ± 0.1 a | 3.4 ± 0.2 a |
| Apple skin | 3.4 ± 0.3 a | 3.2 ± 0.0 ab | 2.9 ± 0.3 b |
| Mushroom | 3.1 ± 0.4 a | 0.1 ± 0.1 b | 0.0 ± 0.0 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhakal, J.; Jia, M.; Joyce, J.D.; Moore, G.A.; Ovissipour, R.; Bertke, A.S. Survival of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) and Herpes Simplex Virus 1 (HSV-1) on Foods Stored at Refrigerated Temperature. Foods 2021, 10, 1005. https://doi.org/10.3390/foods10051005
Dhakal J, Jia M, Joyce JD, Moore GA, Ovissipour R, Bertke AS. Survival of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) and Herpes Simplex Virus 1 (HSV-1) on Foods Stored at Refrigerated Temperature. Foods. 2021; 10(5):1005. https://doi.org/10.3390/foods10051005
Chicago/Turabian StyleDhakal, Janak, Mo Jia, Jonathan D. Joyce, Greyson A. Moore, Reza Ovissipour, and Andrea S. Bertke. 2021. "Survival of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) and Herpes Simplex Virus 1 (HSV-1) on Foods Stored at Refrigerated Temperature" Foods 10, no. 5: 1005. https://doi.org/10.3390/foods10051005