
Postharvest Dehydration Temperature Modulates the Transcriptomic Programme and Flavonoid Profile of Grape Berries

Abstract
:
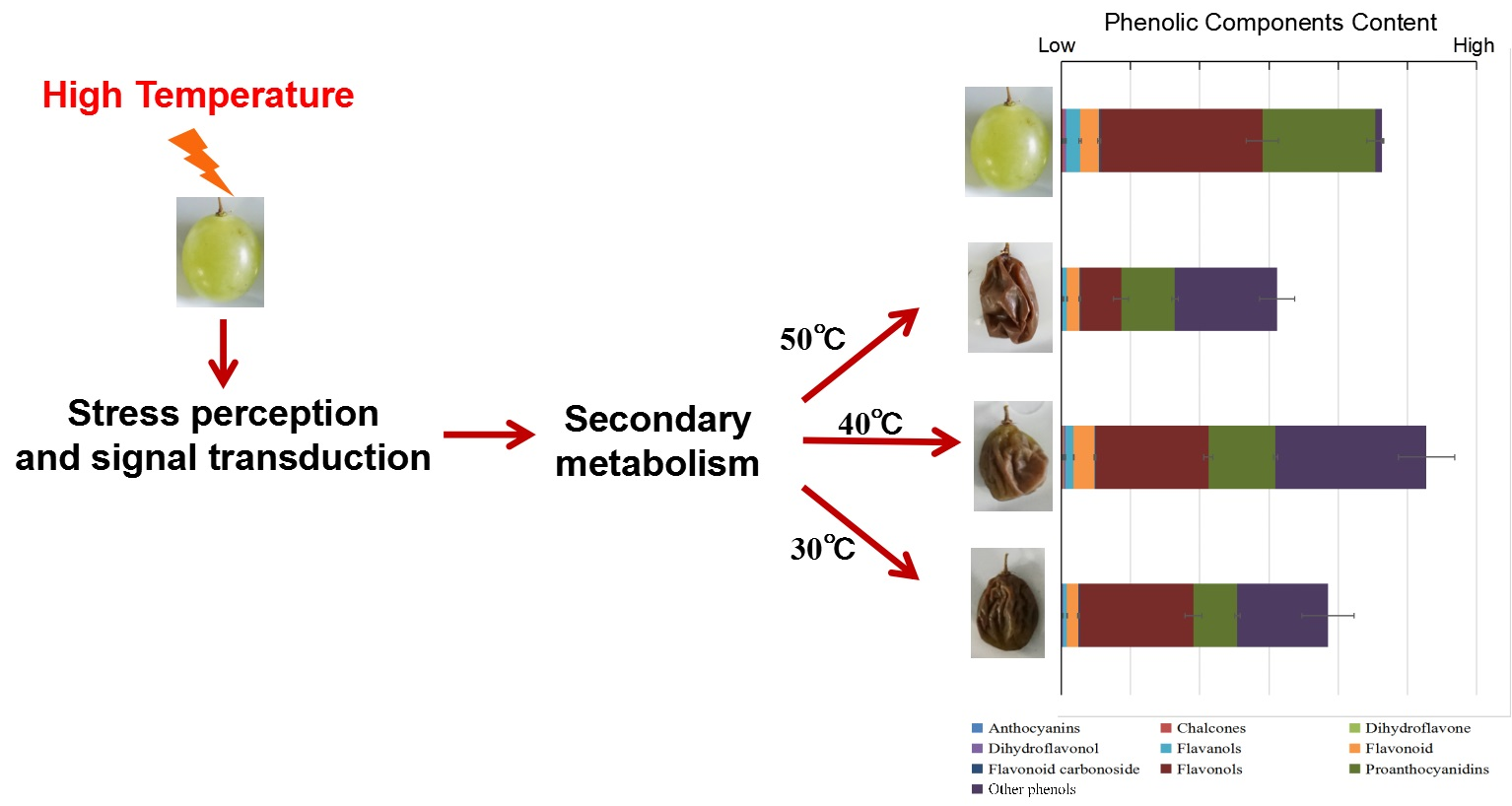{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials Collection and Experimental Layout
2.2. Metabolic Profile Analysis
2.3. ESI-Q TRAP-MS/MS for Targeted Flavonoids Detection and Analysis
2.4. RNA Library Construction and Sequencing
2.5. Transcriptome Analysis
2.6. qRT-PCR Analysis
2.7. Determination of Sugar Components and Organic Acid Components
2.8. Determination of NADP+ and NADPH Content
2.9. Statistical Analysis
3. Results
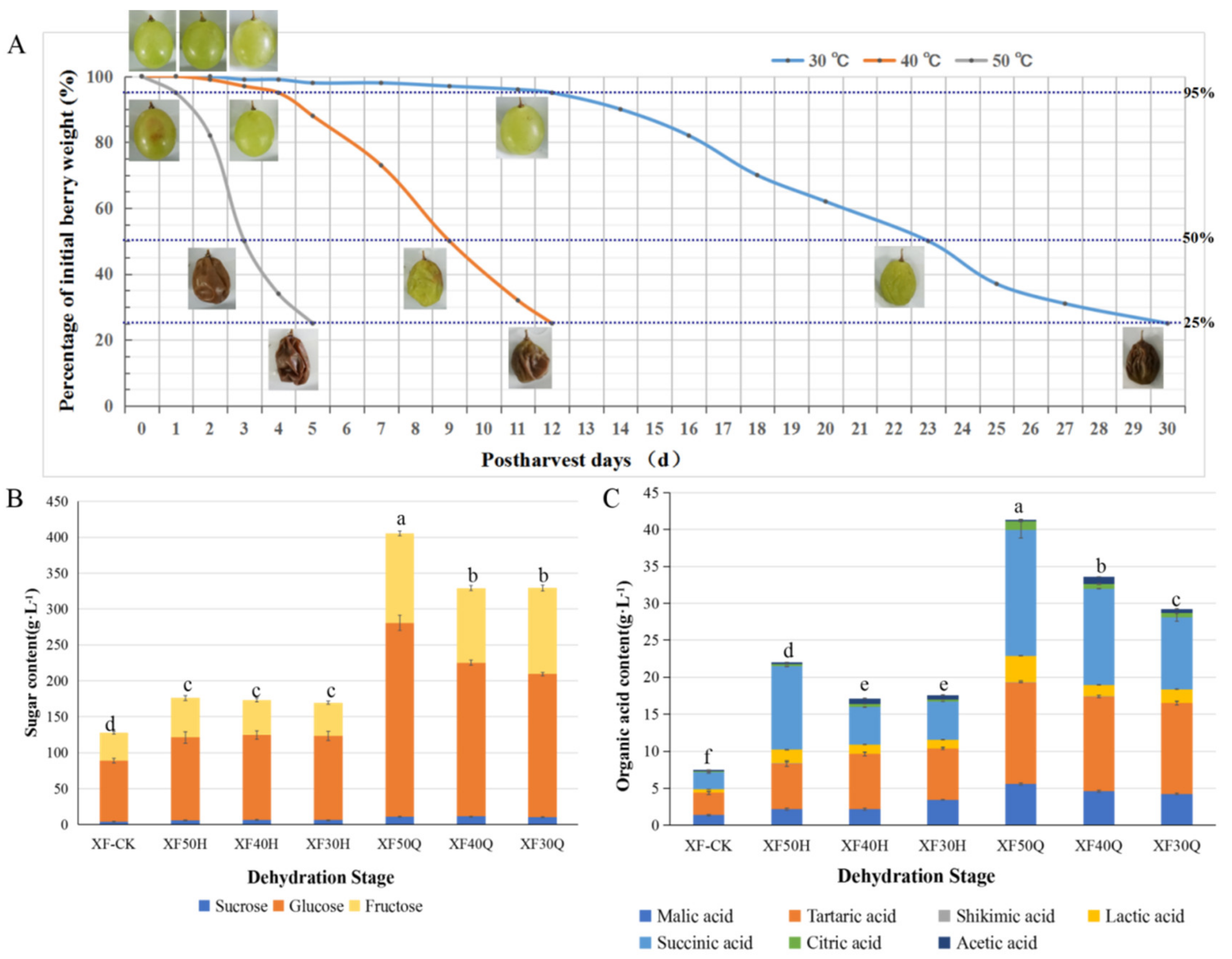
3.1. Changes in Water Content, Soluble Sugar Components, and Organic Acid Components in Grape Berries Subjected to HT Dehydration Mode
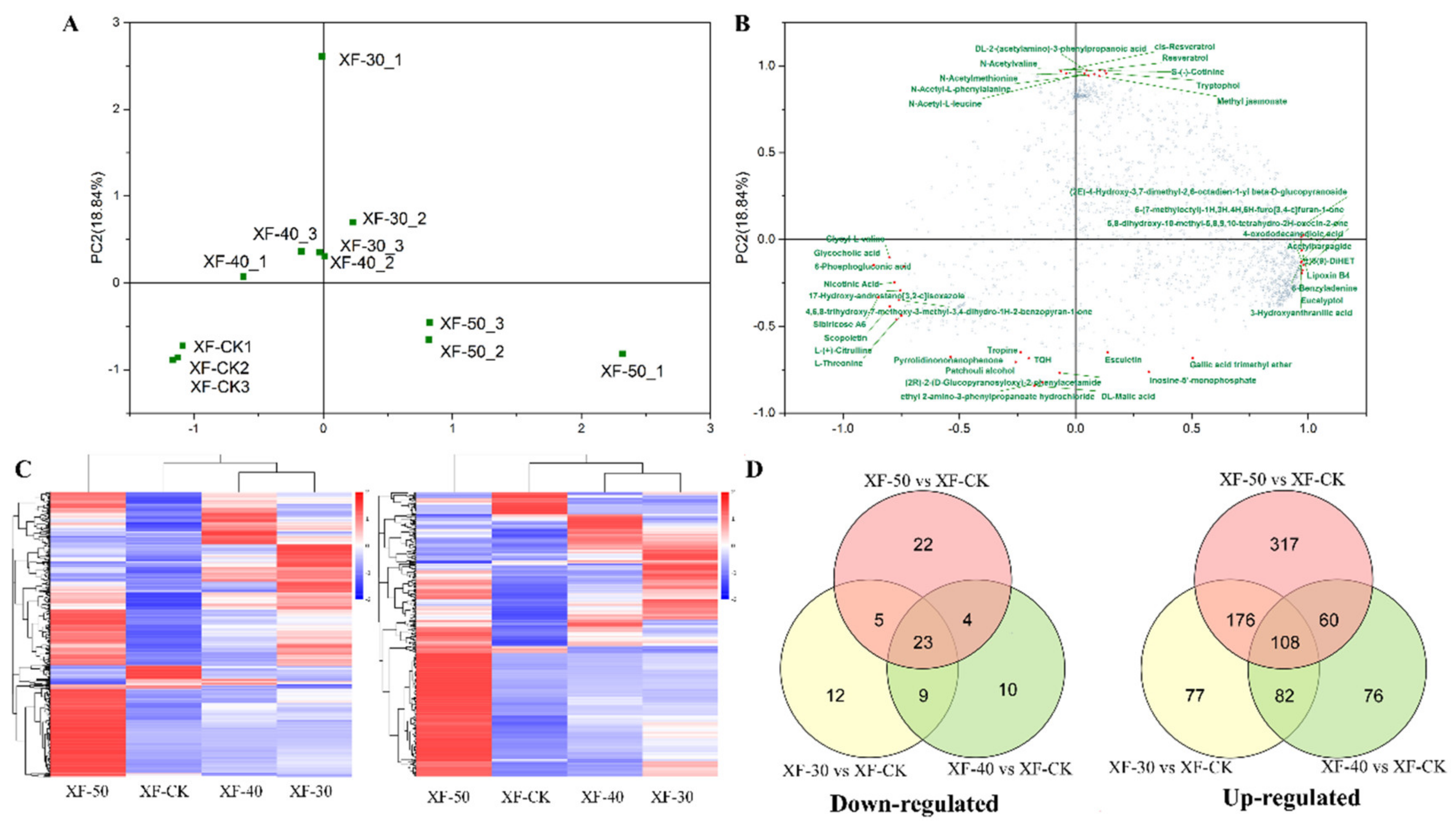
3.2. Differences in the Metabolic Profiles of Raisins Produced under Different HT Dehydration Modes
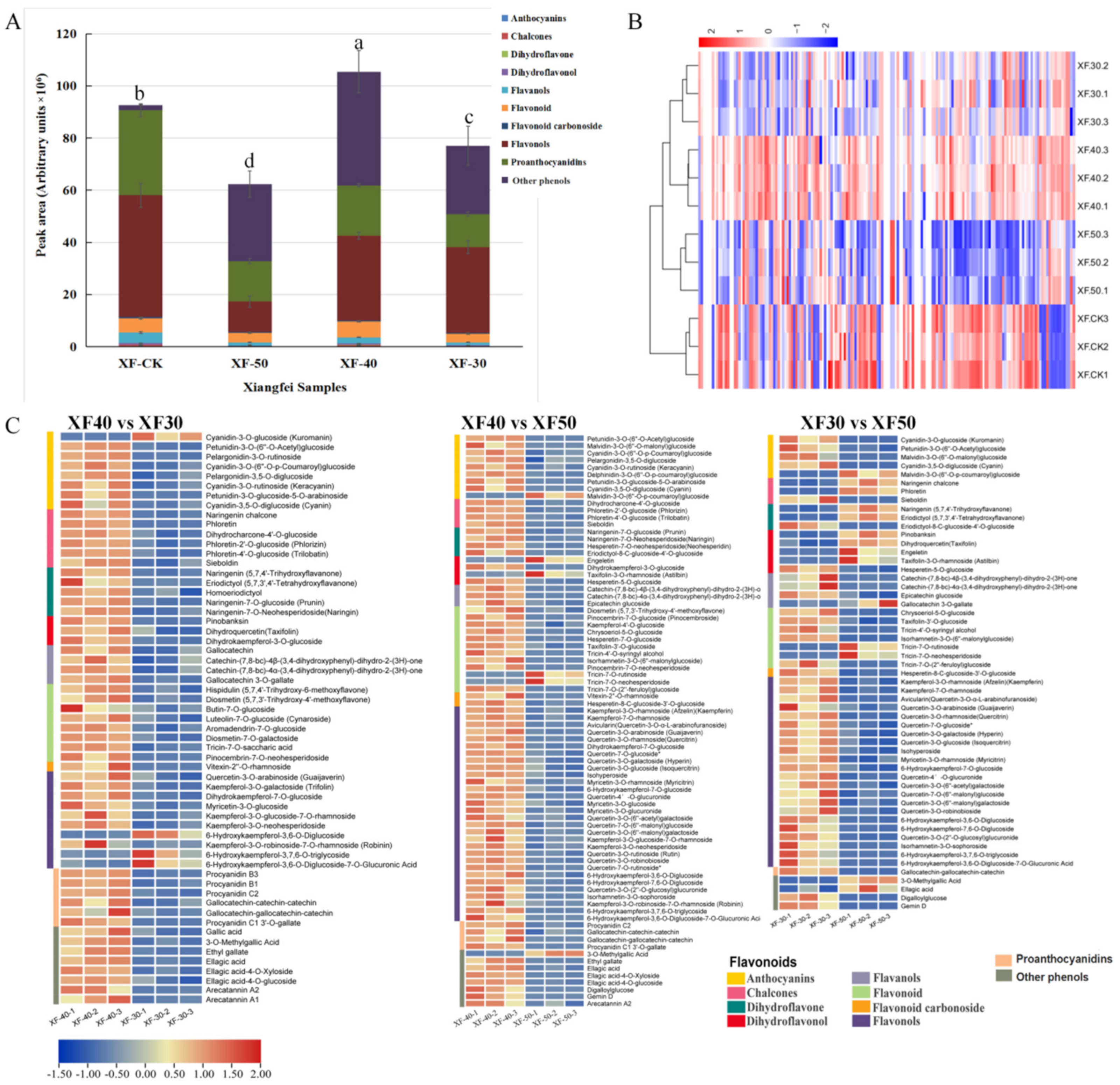
3.3. Difference in Flavonoids, Proanthocyanidins and Other Phenols in Raisins Produced under Different HT Dehydration Modes
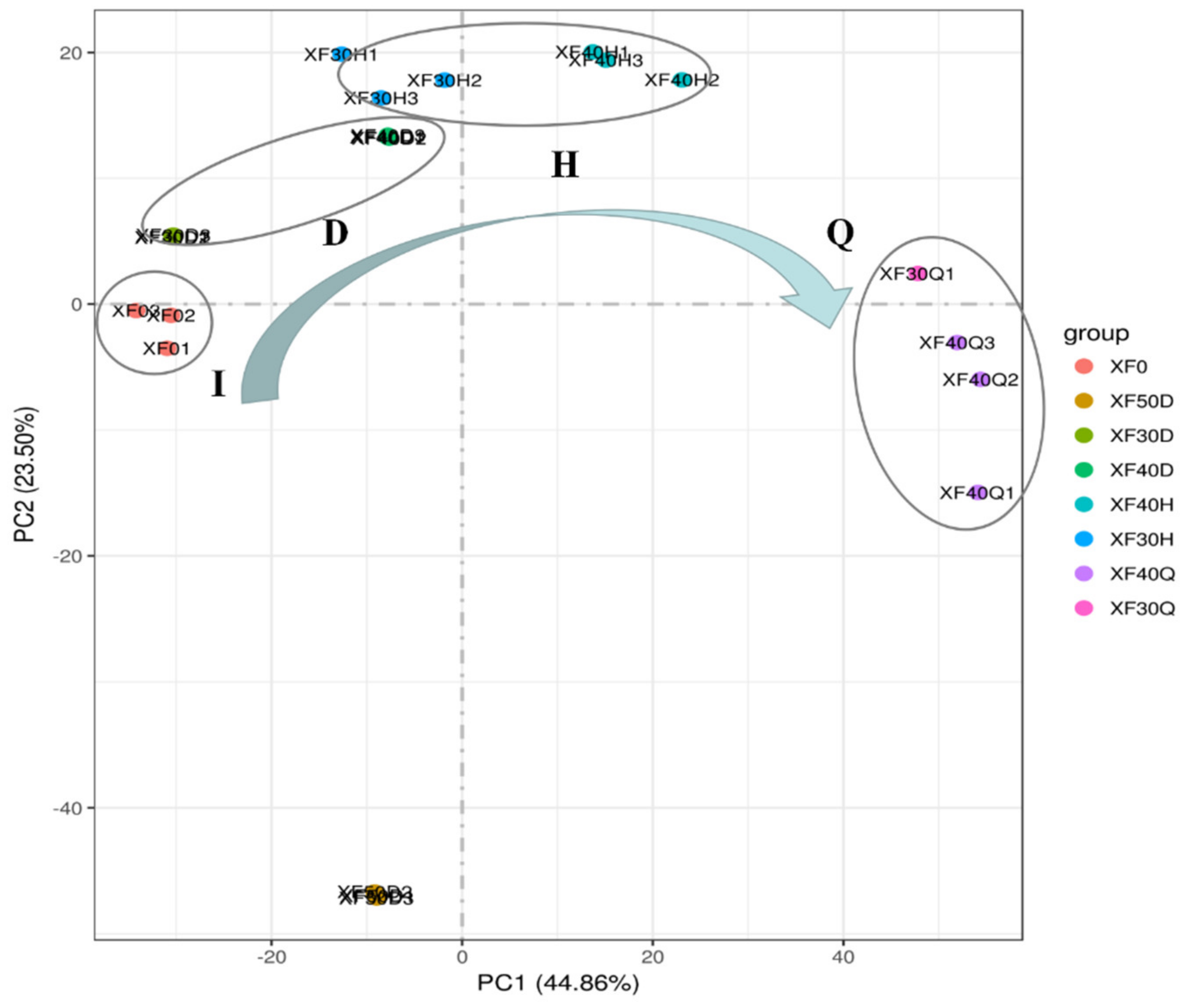
3.4. Transcriptome Analysis in Grape Berries under Different HT Dehydration Modes
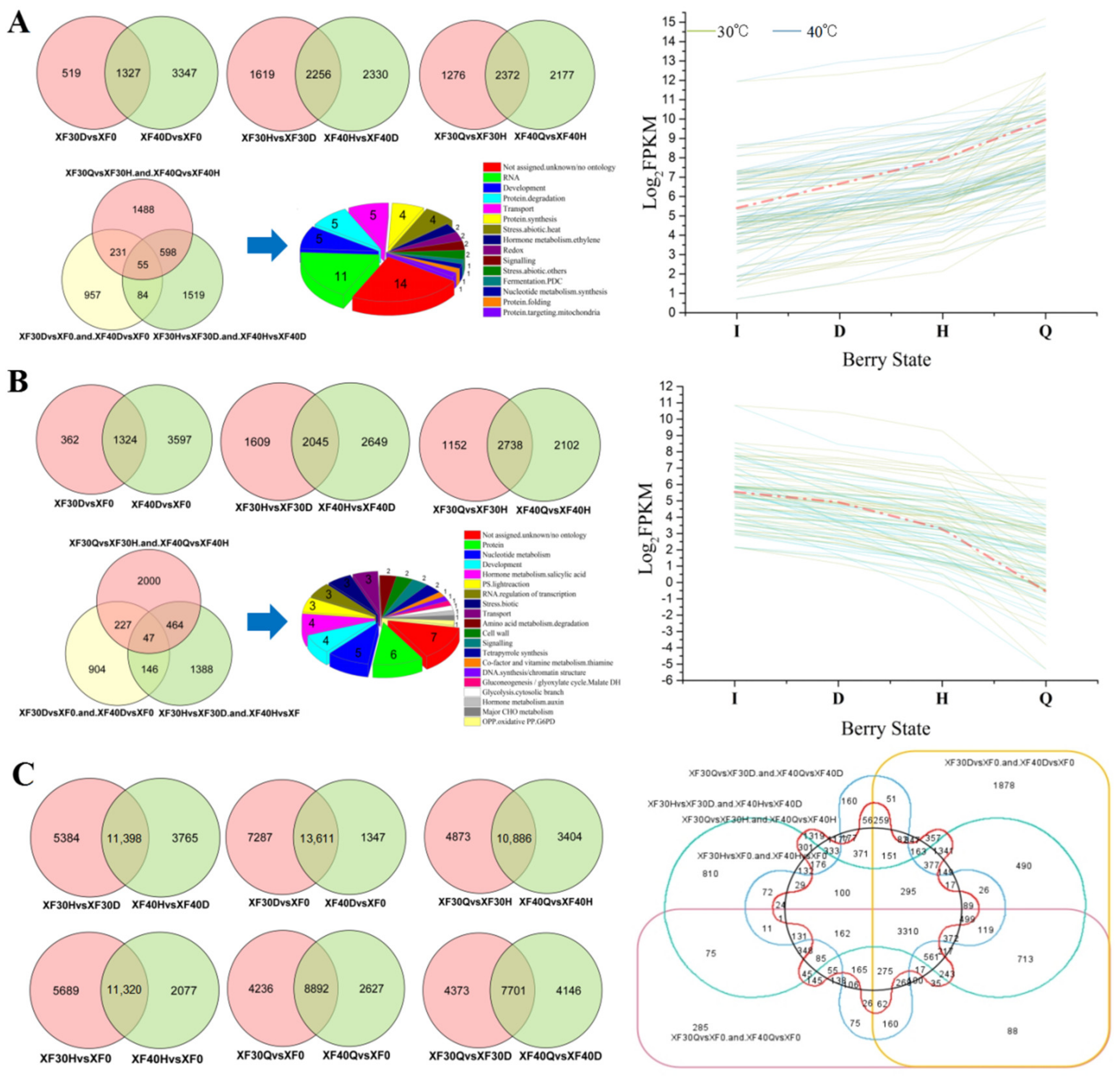
3.5. Changes in the Transcriptome Programme in Grape Berries under Different Dehydration States
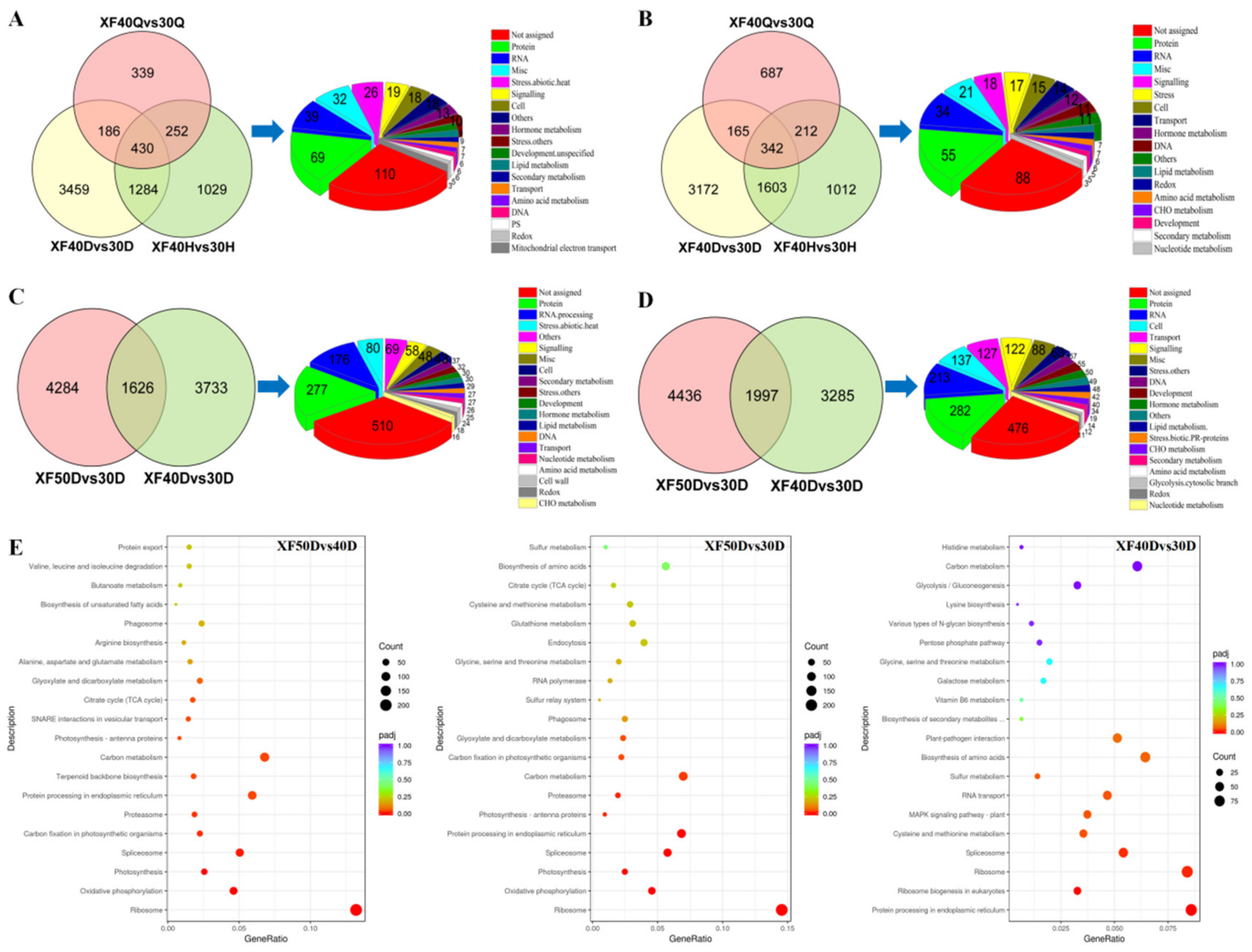
3.6. Analysis of Differentially Expressed Genes under Different Dehydration Temperatures
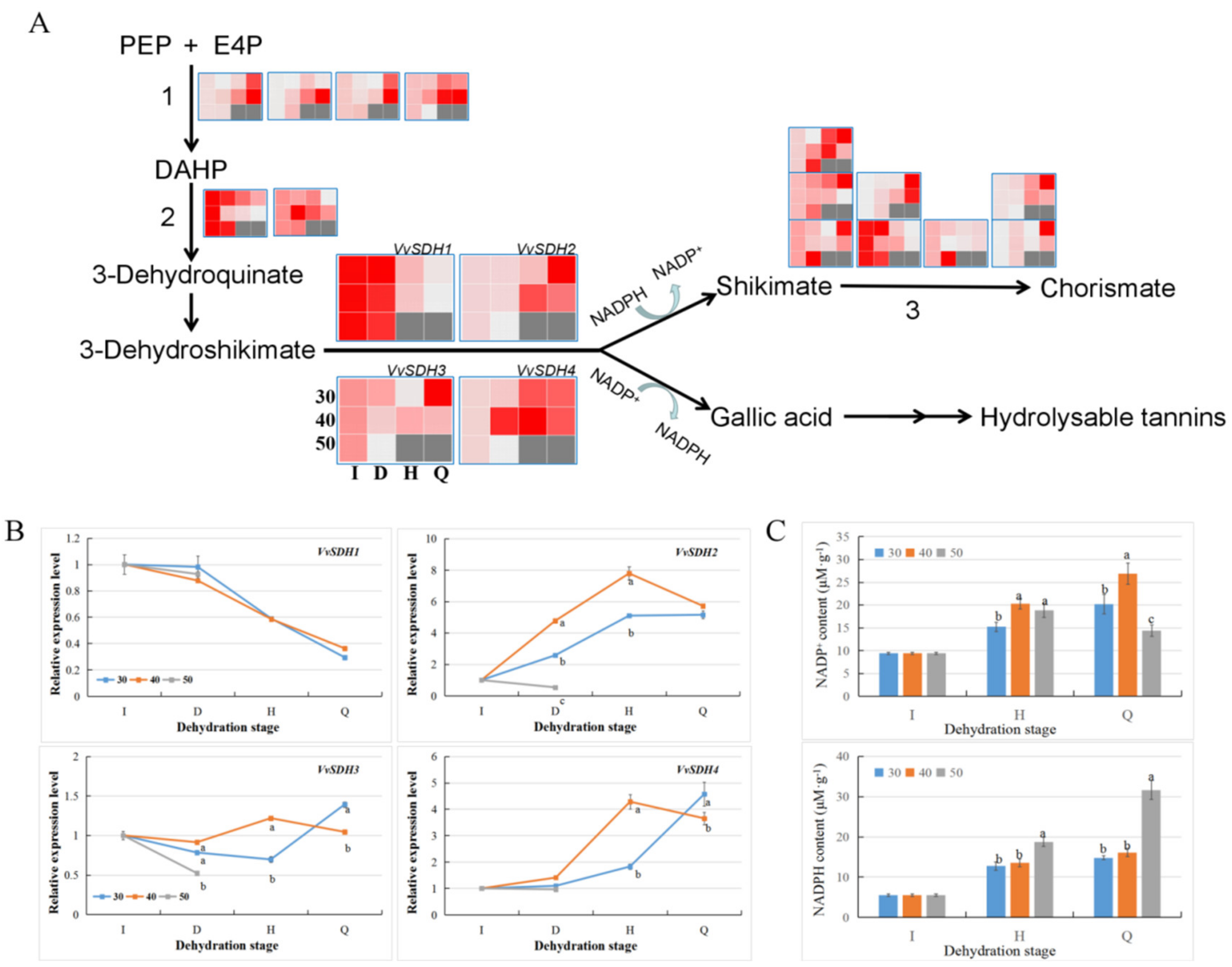
3.7. Differences in the Gallic acid Biosynthesis Pathway in Grape Berries under Different HT Modes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kelebek, H.; Jourdes, M.; Selli, S.; Teissedre, P.L. Comparative evaluation of the phenolic content and antioxidant capacity of sun-dried raisins. J. Sci. Food Agric. 2013, 93, 2963–2972. [Google Scholar] [CrossRef]
- Olmo-Cunillera, A.; Escobar-Avello, D.; Pérez, A.J.; Marhuenda-Muñoz, M.; Lamuela-Raventós, R.M.; Vallverdú-Queralt, A. Is Eating Raisins Healthy? Nutrients 2019, 12, 54. [Google Scholar] [CrossRef]
- Williamson, G.; Carughi, A. Polyphenol content and health benefits of raisins. Nutr. Res. 2010, 30, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Khiari, R.; Zemni, H.; Mihoubi, D. Raisin processing: Physicochemical, nutritional and microbiological quality characteristics as affected by drying process. Food Rev. Int. 2019, 35, 246–298. [Google Scholar] [CrossRef]
- Javed, H.U.; Wang, D.; Shi, Y.; Wu, G.-F.; Xie, H.; Pan, Y.-Q.; Duan, C.-Q. Changes of free-form volatile compounds in pre-treated raisins with different packaging materials during storage. Food Res. Int. 2018, 107, 649–659. [Google Scholar] [CrossRef]
- Zenoni, S.; Fasoli, M.; Guzzo, F.; Santo, S.D.; Amato, A.; Anesi, A.; Commisso, M.; Herderich, M.; Ceoldo, S.; Avesani, L.; et al. Disclosing the Molecular Basis of the Postharvest Life of Berry in Different Grapevine Genotypes. Plant Physiol. 2016, 172, 1821–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Bejarano, M.J.; Duran-Guerrero, E.; Castro, R.; Barroso, C.G.; Rodriguez-Dodero, M.C. Use of Sensory Analysis to Investigate the Influence of Climate Chambers and Other Process Variables in the Production of Sweet Wines. Foods 2020, 9, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonghi, C.; Rizzini, F.M.; Gambuti, A.; Moio, L.; Chkaiban, L.; Tonutti, P. Phenol compound metabolism and gene expression in the skin of wine grape (Vitis vinifera L.) berries subjected to partial postharvest dehydration. Postharvest Biol. Technol. 2012, 67, 102–109. [Google Scholar] [CrossRef]
- Mencarelli, F.; Bellincontro, A.; Nicoletti, I.; Cirilli, M.; Muleo, R.; Corradini, D. Chemical and biochemical change of healthy phenolic fractions in winegrape by means of postharvest dehydration. J. Agric. Food Chem. 2010, 58, 7557–7564. [Google Scholar] [CrossRef] [PubMed]
- Zoccatelli, G.; Zenoni, S.; Savoi, S.; Santo, S.D.; Tononi, P.; Zandonà, V.; Cin, A.D.; Guantieri, V.; Pezzotti, M.; Tornielli, G. Skin pectin metabolism during the postharvest dehydration of berries from three distinct grapevine cultivars. Aust. J. Grape Wine Res. 2013, 19, 171–179. [Google Scholar] [CrossRef]
- Rizzini, F.M.; Bonghi, C.; Tonutti, P. Postharvest water loss induces marked changes in transcript profiling in skins of wine grape berries. Postharvest Biol. Technol. 2009, 52, 247–253. [Google Scholar] [CrossRef]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Gouot, J.C.; Smith, J.P.; Holzapfel, B.P.; Walker, A.R.; Barril, C. Grape berry flavonoids: A review of their biochemical responses to high and extreme high temperatures. J. Exp. Bot. 2019, 70, 397–423. [Google Scholar] [CrossRef] [PubMed]
- Lecourieux, F.; Kappel, C.; Pieri, P.; Charon, J.; Pillet, J.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Lecourieux, D. Dissecting the Biochemical and Transcriptomic Effects of a Locally Applied Heat Treatment on Developing Cabernet Sauvignon Grape Berries. Front. Plant Sci. 2017, 8, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K.; Sugaya, S.; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic. 2005, 105, 319–330. [Google Scholar] [CrossRef]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot. 2007, 58, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Jeong, S.T.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Vitic. 2006, 57, 54–59. [Google Scholar]
- Azuma, A.; Yakushiji, H.; Sato, A. Postharvest light irradiation and appropriate temperature treatment increase anthocyanin accumulation in grape berry skin. Postharvest Biol. Technol. 2019, 147, 89–99. [Google Scholar] [CrossRef]
- Conde, A.; Soares, F.; Breia, R.; Gerós, H. Postharvest dehydration induces variable changes in the primary metabolism of grape berries. Food Res. Int. 2018, 105, 261–270. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhao, P.; Zhang, P.; Su, L.; Jia, H.; Wei, X.; Fang, J.; Jia, H. Integrative transcriptomics and metabolomics data exploring the effect of chitosan on postharvest grape resistance to Botrytis cinerea. Postharvest Biol. Technol. 2020, 167, 111248. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 7, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Liu, Z.; Guan, L.; Zheng, T.; Jiu, S.; Zhu, X.; Jia, H.; Fang, J. Changes of Anthocyanin Component Biosynthesis in ‘Summer Black’ Grape Berries after the Red Flesh Mutation Occurred. J. Agric. Food Chem. 2018, 35, 9209–9218. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, B.; Fan, P.; Xu, H.; Li, S. Inheritance of sugars and acids in berries of grape (Vitis vinifera L.). Euphytica 2007, 1, 99–107. [Google Scholar] [CrossRef]
- Zenoni, S.; Amato, A.; D’Inca, E.; Guzzo, F.; Tornielli, G.B. Rapid dehydration of grape berries dampens the post-ripening transcriptomic program and the metabolite profile evolution. Hortic. Res. 2020, 7, 141. [Google Scholar] [CrossRef]
- Bontpart, T.; Marlin, T.; Vialet, S.; Guiraud, J.L.; Pinasseau, L.; Meudec, E.; Sommerer, N.; Cheynier, V.; Terrier, N. Two shikimate dehydrogenases, VvSDH3 and VvSDH4, are involved in gallic acid biosynthesis in grapevine. J. Exp. Bot. 2016, 67, 3537–3550. [Google Scholar] [CrossRef] [Green Version]
- Zamboni, A.; Minoia, L.; Ferrarini, A.; Tornielli, G.B.; Zago, E.; Delledonne, M.; Pezzotti, M. Molecular analysis of post-harvest withering in grape by AFLP transcriptional profiling. J. Exp. Bot. 2008, 59, 4145–4159. [Google Scholar] [CrossRef] [Green Version]
- Olivati, C.; Nishiyama, Y.P.D.O.; de Souza, R.T.; Janzantti, N.S.; Mauro, M.A.; Gomes, E.; Hermosín-Gutiérrez, I.; da Silva, R.; Lago-Vanzela, E.S. Effect of the pre-treatment and the drying process on the phenolic composition of raisins produced with a seedless Brazilian grape cultivar. Food Res. Int. 2019, 116, 190–199. [Google Scholar] [CrossRef] [Green Version]
- Barbanti, D.; Mora, B.; Ferrarini, R.; Tornielli, G.B.; Cipriani, M. Effect of various thermo-hygrometric conditions on the withering kinetics of grapes used for the production of “Amarone” and “Recioto” wines. J. Food Eng. 2008, 85, 350–358. [Google Scholar] [CrossRef]
- An, X.; Xu, Y.; Jiang, L.; Huan, C.; Yu, Z. Effects of postharvest temperature on apoptosis-related enzyme activity and gene expression in peach fruits (Prunus persica L. cv. Xiahui 8). Sci. Hortic. 2019, 245, 178–184. [Google Scholar] [CrossRef]
- Wu, Z.; Yuan, X.; Li, H.; Liu, F.; Wang, Y.; Li, J.; Cai, H.; Wang, Y. Heat acclimation reduces postharvest loss of table grapes during cold storage—Analysis of possible mechanisms involved through a proteomic approach. Postharvest Biol. Technol. 2015, 105, 26–33. [Google Scholar] [CrossRef]
- Nasef, I.N. Short hot water as safe treatment induces chilling tolerance and antioxidant enzymes, prevents decay and maintains quality of cold-stored cucumbers. Postharvest Biol. Technol. 2018, 138, 1–10. [Google Scholar] [CrossRef]
- Ma, Q.; Suo, J.; Huber, D.J.; Dong, X.; Han, Y.; Zhang, Z.; Rao, J. Effect of hot water treatments on chilling injury and expression of a new C-repeat binding factor (CBF) in ‘Hongyang’ kiwifruit during low temperature storage. Postharvest Biol. Technol. 2014, 97, 102–110. [Google Scholar] [CrossRef]
- Pu, H.; Shan, S.; Wang, Z.; Duan, W.; Tian, J.; Zhang, L.; Li, J.; Song, H.; Xu, X. Dynamic Changes of DNA Methylation Induced by Heat Treatment Were Involved in Ethylene Signal Transmission and Delayed the Postharvest Ripening of Tomato Fruit. J. Agric. Food Chem. 2020, 68, 8976–8986. [Google Scholar] [CrossRef]
- Xie, Y.; Xu, D.; Cui, W.; Shen, W. Mutation of Arabidopsis HY1 causes UV-C hypersensitivity by impairing carotenoid and flavonoid biosynthesis and the down-regulation of antioxidant defence. J. Exp. Bot. 2012, 63, 3869–3883. [Google Scholar] [CrossRef] [Green Version]
- Gouot, J.C.; Smith, J.P.; Holzapfel, B.P.; Barril, C. Impact of short temperature exposure of Vitis vinifera L. cv. Shiraz grapevine bunches on berry development, primary metabolism and tannin accumulation. Environ. Exp. Bot. 2019, 168, 103866. [Google Scholar] [CrossRef]
- Shinomiya, R.; Fujishima, H.; Muramoto, K.; Shiraishi, M. Impact of temperature and sunlight on the skin coloration of the ‘Kyoho’ table grape. Sci. Hortic. 2015, 193, 77–83. [Google Scholar] [CrossRef]
- Nievierowski, T.H.; Veras, F.F.; Silveira, R.D.; Dachery, B.; Hernandes, K.C.; Lopes, F.C.; Scortegagna, E.; Zini, C.A.; Welke, J.E. Role of partial dehydration in a naturally ventilated room on the mycobiota, ochratoxins, volatile profile and phenolic composition of Merlot grapes intended for wine production. Food Res. Int. 2021, 141, 110145. [Google Scholar] [CrossRef]
- Ossipov, V.; Salminen, J.; Ossipova, S.; Haukioja, E.; Pihlaja, K. Gallic acid and hydrolysable tannins are formed in birch leaves from an intermediate compound of the shikimate pathway. Biochem. Syst. Ecol. 2003, 31, 3–16. [Google Scholar] [CrossRef]
- Habashi, R.; Hacham, Y.; Dhakarey, R.; Matityahu, I.; Holland, D.; Tian, L.; Amir, R. Elucidating the role of shikimate dehydrogenase in controlling the production of anthocyanins and hydrolysable tannins in the outer peels of pomegranate. BMC Plant Biol. 2019, 19, 476. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.-D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Liu, G.T.; Ma, L.; Duan, W.; Wang, B.-C.; Li, J.-H.; Xu, H.-G.; Yan, X.-Q.; Yan, B.-F.; Li, S.-H.; Wang, L.-J. Differential proteomic analysis of grapevine leaves by iTRAQ reveals responses to heat stress and subsequent recovery. BMC Plant Biol. 2014, 14, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basha, E.; O’Neill, H.; Vierling, E. Small heat shock proteins and a-crystallins: Dynamic proteins with flexible functions. Trends Biochem. Sci. 2012, 37, 106–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, S.M.; Genest, O.; Wickner, S. Protein rescue from aggregates by powerful molecular chaperone machines. Nat. Rev. Mol. Cell Biol. 2013, 14, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- McLoughlin, F.; Basha, E.; Fowler, M.E.; Kim, M.; Bordowitz, J.; Katiyar-Agarwal, S.; Vierling, E. Class I and II Small Heat Shock Proteins Together with HSP101 Protect Protein Translation Factors during Heat Stress. Plant Physiol. 2016, 172, 1221–1236. [Google Scholar]
- Liu, G.T.; Jiang, J.-F.; Liu, X.-N.; Jiang, J.-Z.; Sun, L.; Duan, W.; Li, R.-M.; Wang, Y.; Lecourieux, D.; Liu, C.-H.; et al. New insights into the heat responses of grape leaves via combined phosphoproteomic and acetylproteomic analyses. Hortic. Res. 2019, 6, 100. [Google Scholar] [CrossRef] [Green Version]
- Bokszczanin, K.L.; Fragkostefanakis, S. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 2013, 4, 315. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, X.; Liu, C.; Liu, G.; Li, S.; Wang, L. Integrating Omics and Alternative Splicing Reveals Insights into Grape Response to High Temperature. Plant Physiol. 2017, 2, 1502–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Wu, Q.; Duan, X.; Yun, Z.; Jiang, Y. Proteomic and transcriptomic analysis to unravel the influence of high temperature on banana fruit during postharvest storage. Funct. Integr. Genom. 2019, 19, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.A.; Sepulveda, G.; Kahlaoui, B. Phytohormone Interaction Modulating Fruit Responses to Photooxidative and Heat Stress on Apple (Malus domestica Borkh.). Front. Plant Sci. 2017, 8, 2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Sun, J.; Li, Z.; Zhang, J.; Li, Z.; Chen, L.; Li, W.; Fang, Y.; Zhang, K. Postharvest Dehydration Temperature Modulates the Transcriptomic Programme and Flavonoid Profile of Grape Berries. Foods 2021, 10, 687. https://doi.org/10.3390/foods10030687
Chen K, Sun J, Li Z, Zhang J, Li Z, Chen L, Li W, Fang Y, Zhang K. Postharvest Dehydration Temperature Modulates the Transcriptomic Programme and Flavonoid Profile of Grape Berries. Foods. 2021; 10(3):687. https://doi.org/10.3390/foods10030687
Chicago/Turabian StyleChen, Keqin, Jiahua Sun, Zhihao Li, Junxia Zhang, Ziyu Li, Li Chen, Wanping Li, Yulin Fang, and Kekun Zhang. 2021. "Postharvest Dehydration Temperature Modulates the Transcriptomic Programme and Flavonoid Profile of Grape Berries" Foods 10, no. 3: 687. https://doi.org/10.3390/foods10030687