
Optimization of Phytase Production from Escherichia coli by Altering Solid-State Fermentation Conditions


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
2.1. E. coli Inoculum
2.2. Solid-State Fermentation

2.3. Effect of Nutrient Additives on Phytase Production
2.4. Effect of Substrate Moisture on Phytase Production
2.5. Effect of Inoculation Rate on Phytase Production
2.6. Effect of Incubation Period on Phytase Production
2.7. Effect of Temperature and pH on Phytase Activity
2.8. Phytase Activity Assay
2.9. Statistical Analyses
3. Results and Discussion
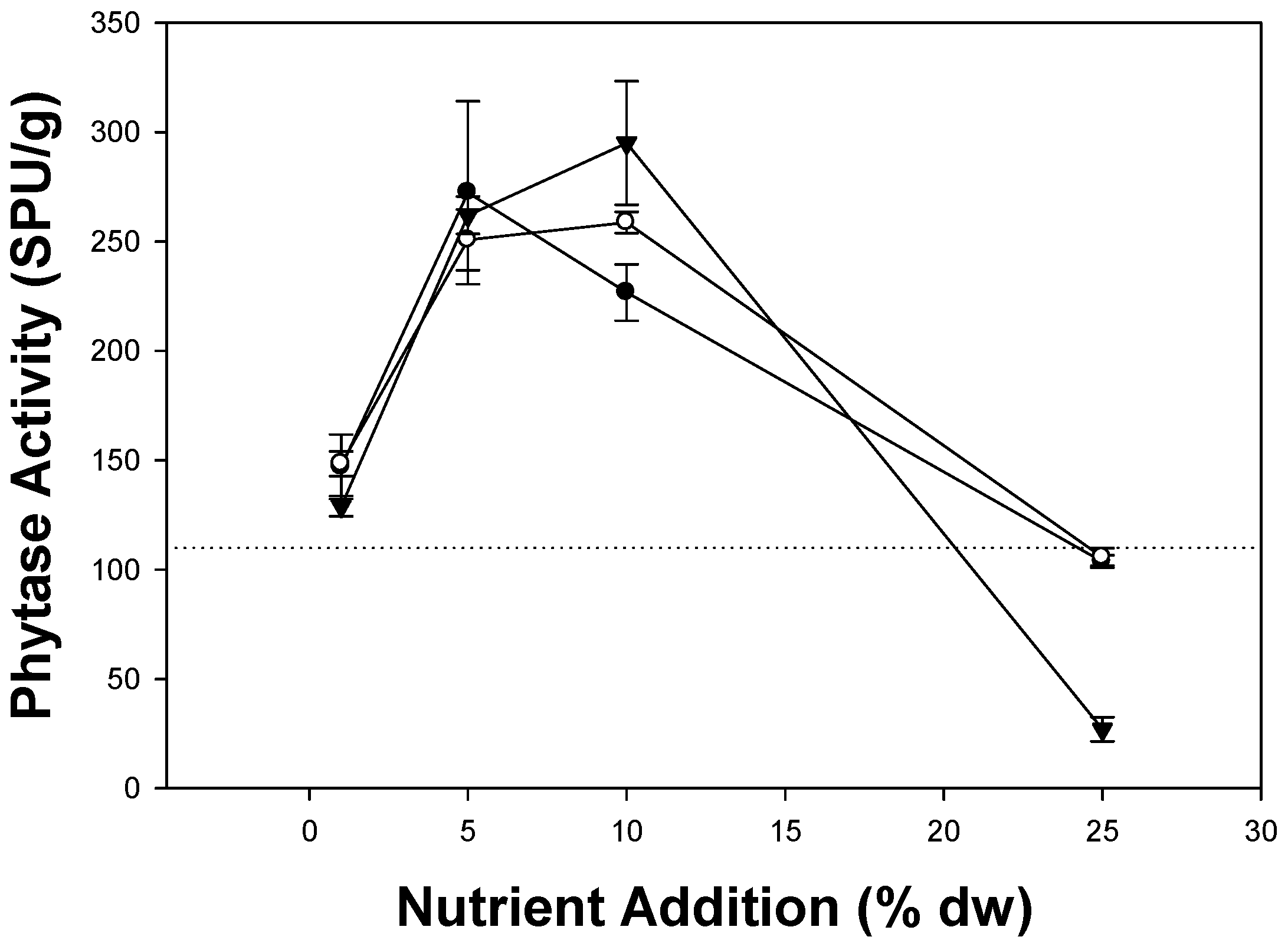
3.1. Effect of Additives on Phytase Production

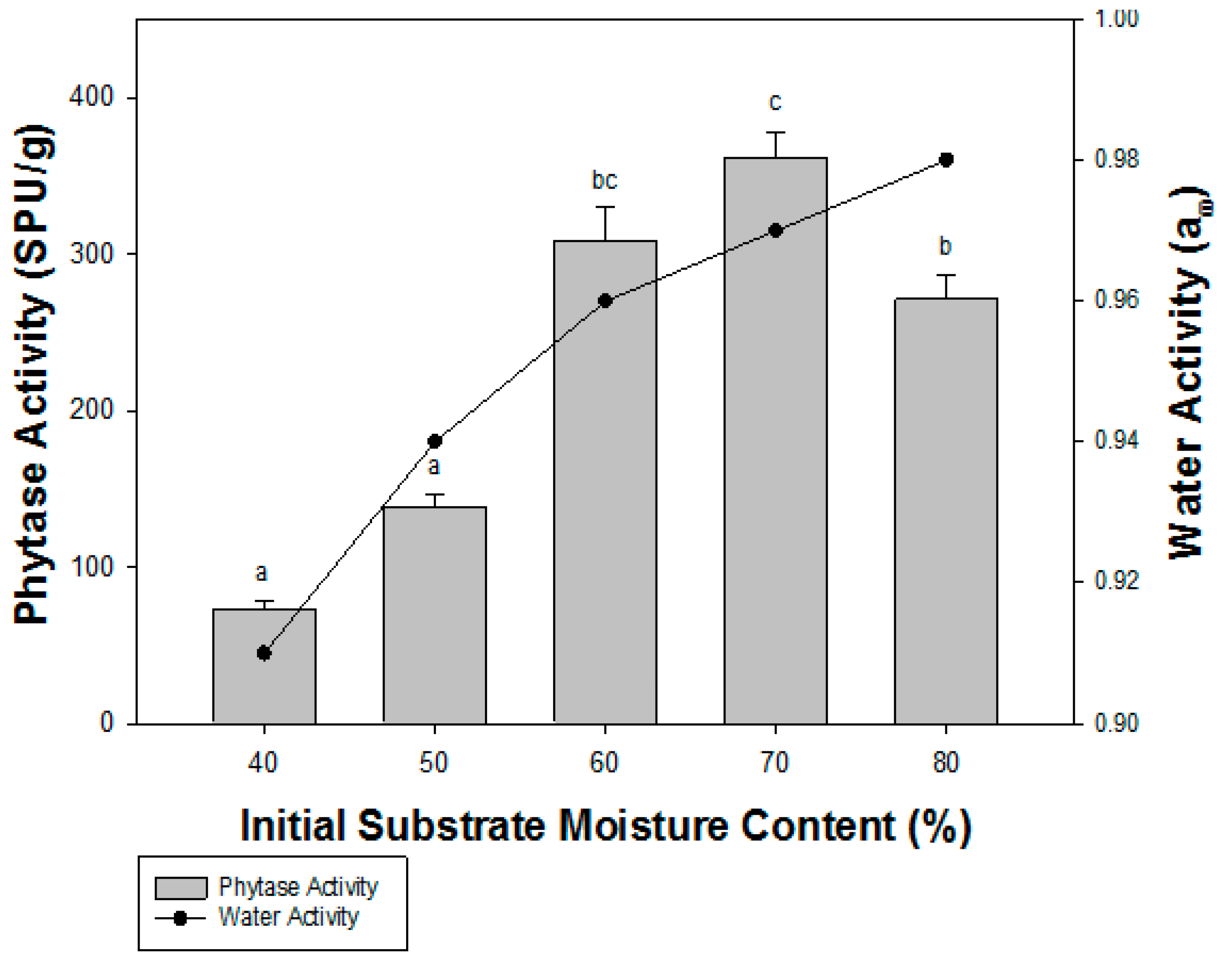
3.2. Effect of Moisture Level and Water Activity on Phytase Production

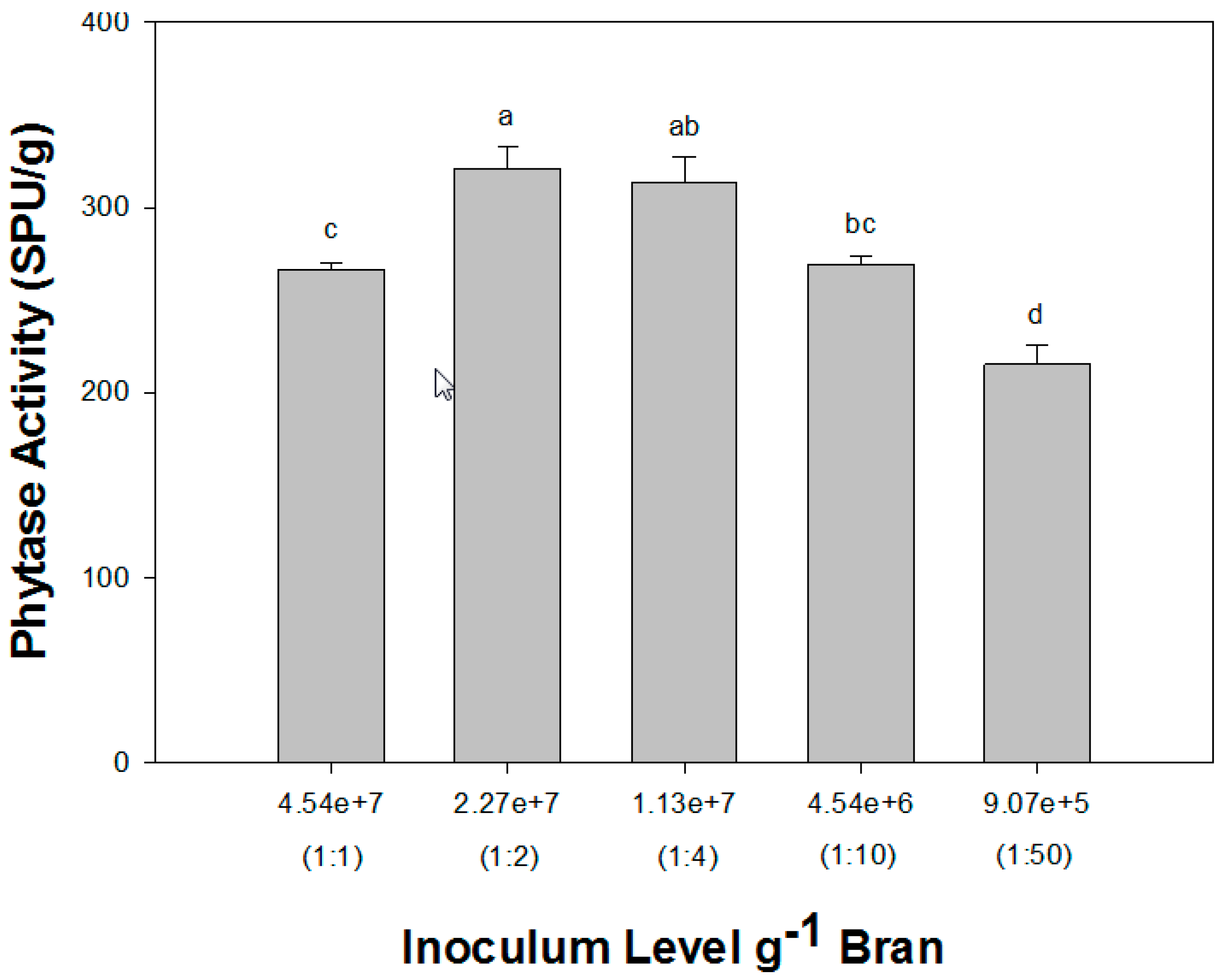
3.3. Effect of Inoculum Rate on Phytase Production

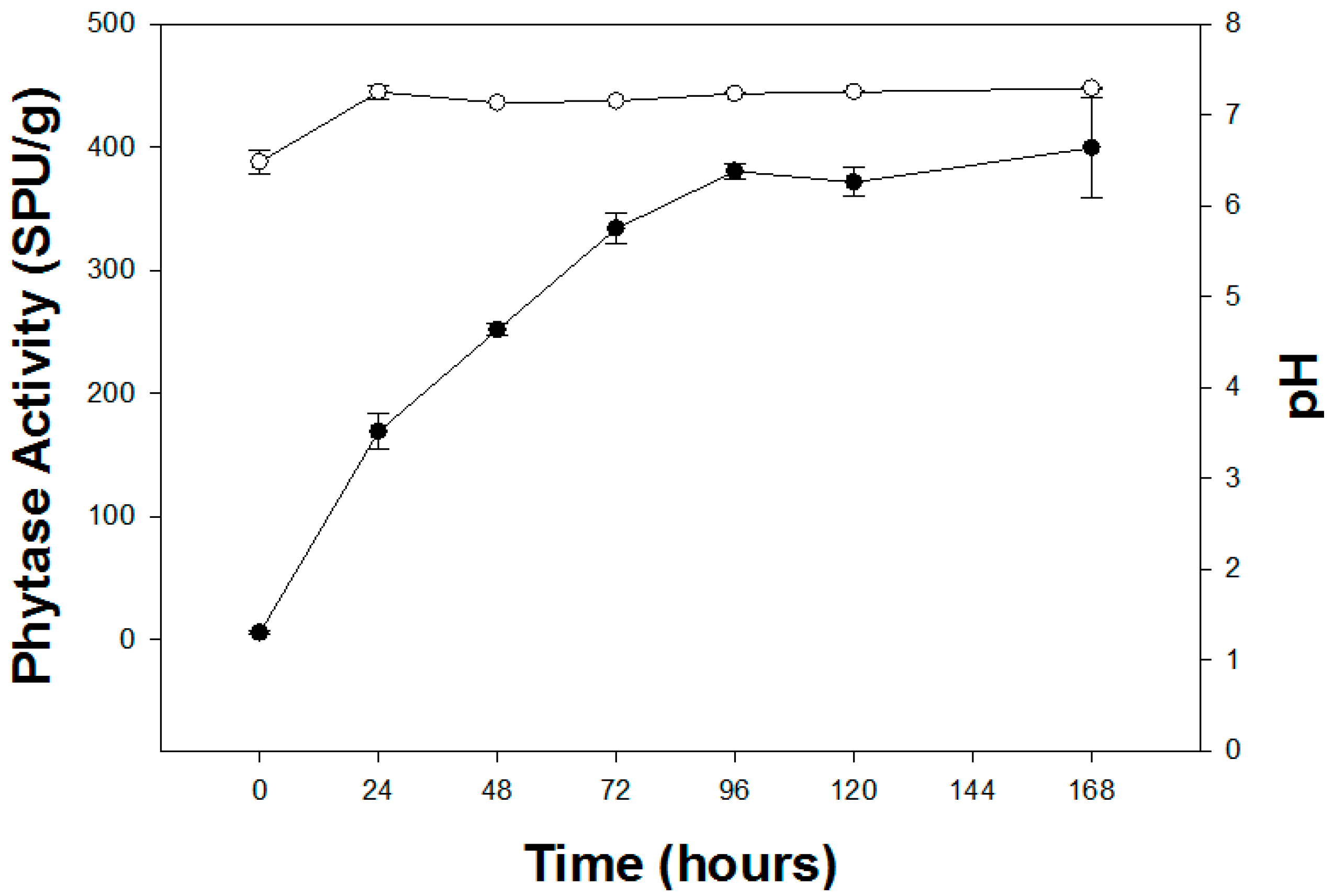
3.4. Effect of Incubation Period on Phytase Production

3.5. Comparison between SmF and SSF on Phytase Productivity
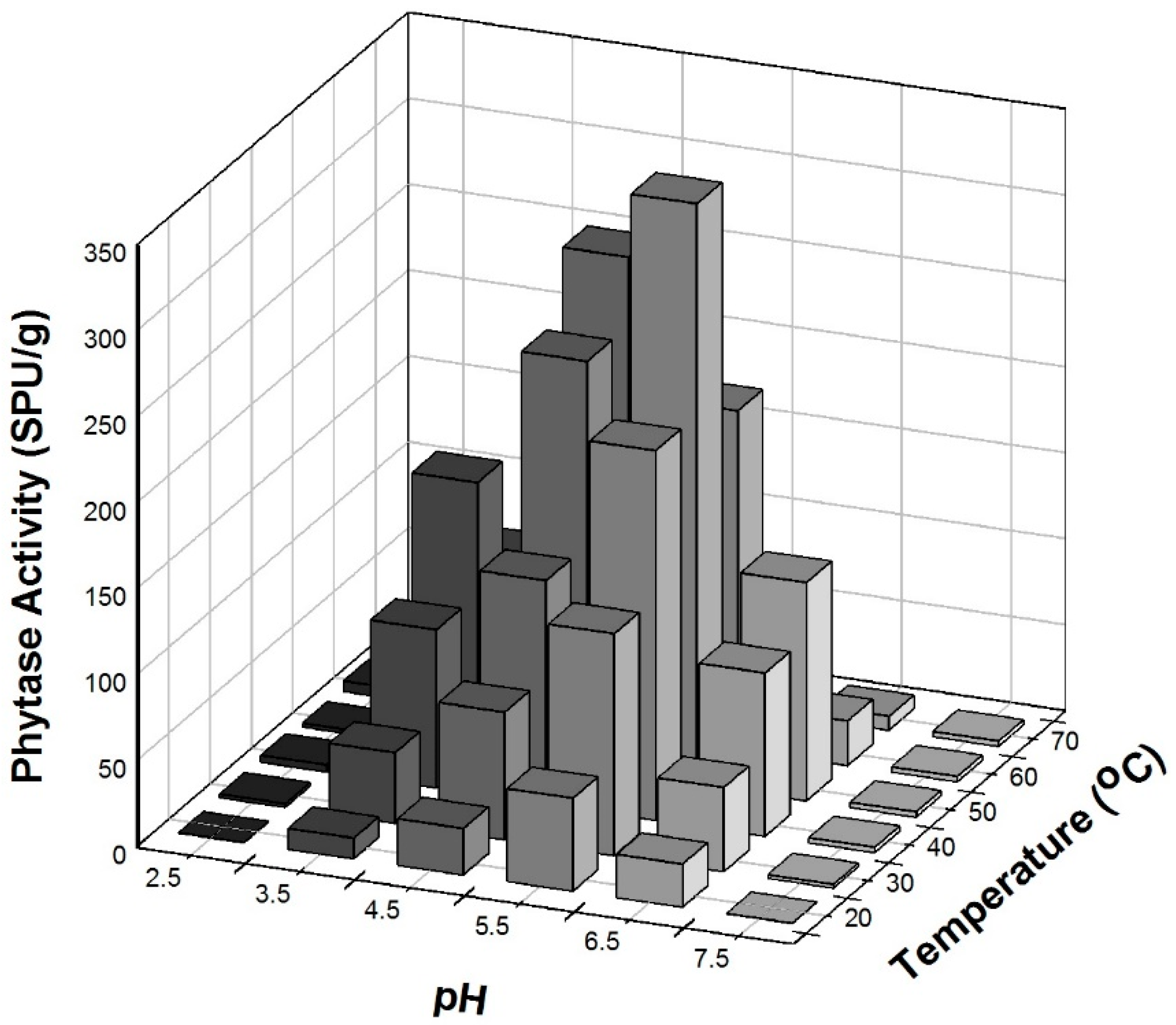
3.6. Effect of Temperature and pH on Phytase Activity

3.7. General Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lei, X.G.; Weaver, J.D.; Mullaney, E.; Ullah, A.H.; Azain, M.J. Phytase, a new life for an “Old” enzyme. Annu. Rev. Anim. Biosci. 2013, 1, 283–309. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.; Larroche, C.; Pandey, A. Current developments in solid-state fermentation. Biochem Eng. J. 2013, 81, 146–161. [Google Scholar] [CrossRef]
- Graminha, E.B.N.; Gonçalves, A.Z.L.; Pirota, R.D.P.B.; Balsalobre, M.A.A.; da Silva, R.; Gomes, E. Enzyme production by solid-state fermentation: Application to animal nutrition. Anim. Feed Sci. Tech. 2008, 144, 1–22. [Google Scholar] [CrossRef]
- El-Shishtawy, R.M.; Mohamed, S.A.; Asiri, A.M.; Gomaa, A.B.; Ibrahim, I.H.; Al-Talhi, H.A. Solid fermentation of wheat bran for hydrolytic enzymes production and saccharification content by a local isolate Bacillus megatherium. BMC Biotechnol. 2014, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Svihus, B. Effect of Digestive Tract Conditions, Feed Processing and Ingredients on Response to NSP Enzymes. In Enzymes in Farm Animal Nutrition; Bedford, M.R., Partridge, G.G., Eds.; CAB International: Bodmin, UK, 2011. [Google Scholar]
- Sreedevi, S.; Reddy, B.N. Isolation, screening, and optimization of phytase production from newly isolated Bacillus sp. C43. Int. J. Pharm. Biol. Sci. 2012, 2, 218–231. [Google Scholar]
- Igbansan, F.A.; Manner, K.; Miksch, G.; Borriss, R.; Farouk, A.; Simon, O. Comparative Studies on the In Vitro Properties of Phytases from Various Microbial Origins. Arch. Anim. Nutr. 2000, 53, 353–373. [Google Scholar]
- Vohra, A.; Satyanarayana, T. Phytases: Microbial sources, production, purification, and potential biotechnological applications. Crit. Rev. Biotechnol. 2003, 23, 29–60. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Soccol, C.R.; Larroche, C. Current Developments in Solid-State Fermentation; AsiaTech Publishers, Inc.: Patparganj, Delhi, India, 2008. [Google Scholar]
- Chen, J.; Zhu, Y. Solid State Fermentation for Foods and Beverages; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Bhargav, S.; Panda, B.P.; Ali, M.; Javed, S. Solid-State Fermenation: An Overview. Chem. Biochem. Eng. Q. 2008, 22, 49–70. [Google Scholar]
- Krishna, C. Solid-state fermentation systems—An overview. Crit. Rev. Biotechnol. 2005, 25, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Raimbault, M. General and microbiological aspects of solid substrate fermentation. J. Biotechnol. 1998, 1, 1–15. [Google Scholar] [CrossRef]
- Gervais, P.; Molin, P. The role of water in solid-state fermentation. Biochem Eng. J. 2003, 13, 85–101. [Google Scholar] [CrossRef]
- Montiel-González, A.M.; Viniegra-González, G.; Fernández, F.J.; Loera, O. Effect of water activity on invertase production in solid state fermentation by improved diploid strains of Aspergillus niger. Proc. Biochem. 2004, 39, 2085–2090. [Google Scholar] [CrossRef]
- Kavya, V.; Padmavathi, T. Optimization of growth conditions for xylanase production by Aspergillus niger in solid state fermentation. Pol. J .Microbiol. 2009, 58, 125–130. [Google Scholar] [PubMed]
- Naves, L.D.P.; Corrêa, A.; Bertechini, A.; Gomide, E.; Santos, C.D. Effect of ph and temperature on the activity of phytase products used in broiler nutrition. Rev. Bras. Cienc. Avic. 2012, 14, 181–185. [Google Scholar] [CrossRef]
- Shivanna, G.B.; Venkateswaran, G. Phytase production by Aspergillus niger CFR 335 and Aspergillus ficuum SGA 01 through submerged and solid-state fermentation. Sci. World J. 2014, 2014, 392615. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.M.; Suh, H.J.; Kim, J.M. Purification and properties of extracellular phytase from Bacillus sp. KHU-10. J. Protein. Chem. 2001, 20, 287–292. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McKinney, K.; Combs, J.; Becker, P.; Humphries, A.; Filer, K.; Vriesekoop, F. Optimization of Phytase Production from Escherichia coli by Altering Solid-State Fermentation Conditions. Fermentation 2015, 1, 13-23. https://doi.org/10.3390/fermentation1010013
McKinney K, Combs J, Becker P, Humphries A, Filer K, Vriesekoop F. Optimization of Phytase Production from Escherichia coli by Altering Solid-State Fermentation Conditions. Fermentation. 2015; 1(1):13-23. https://doi.org/10.3390/fermentation1010013
Chicago/Turabian StyleMcKinney, Kyle, Justin Combs, Patrick Becker, Andrea Humphries, Keith Filer, and Frank Vriesekoop. 2015. "Optimization of Phytase Production from Escherichia coli by Altering Solid-State Fermentation Conditions" Fermentation 1, no. 1: 13-23. https://doi.org/10.3390/fermentation1010013