
Using Macronutrient Distributions within Trees to Define a Branch Diameter Threshold for Biomass Harvest in Sugar Maple-Dominated Stands


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Stand Selection
2.2. Tree Sampling and Compartment Measurements
2.3. Nutrient Content Analyses
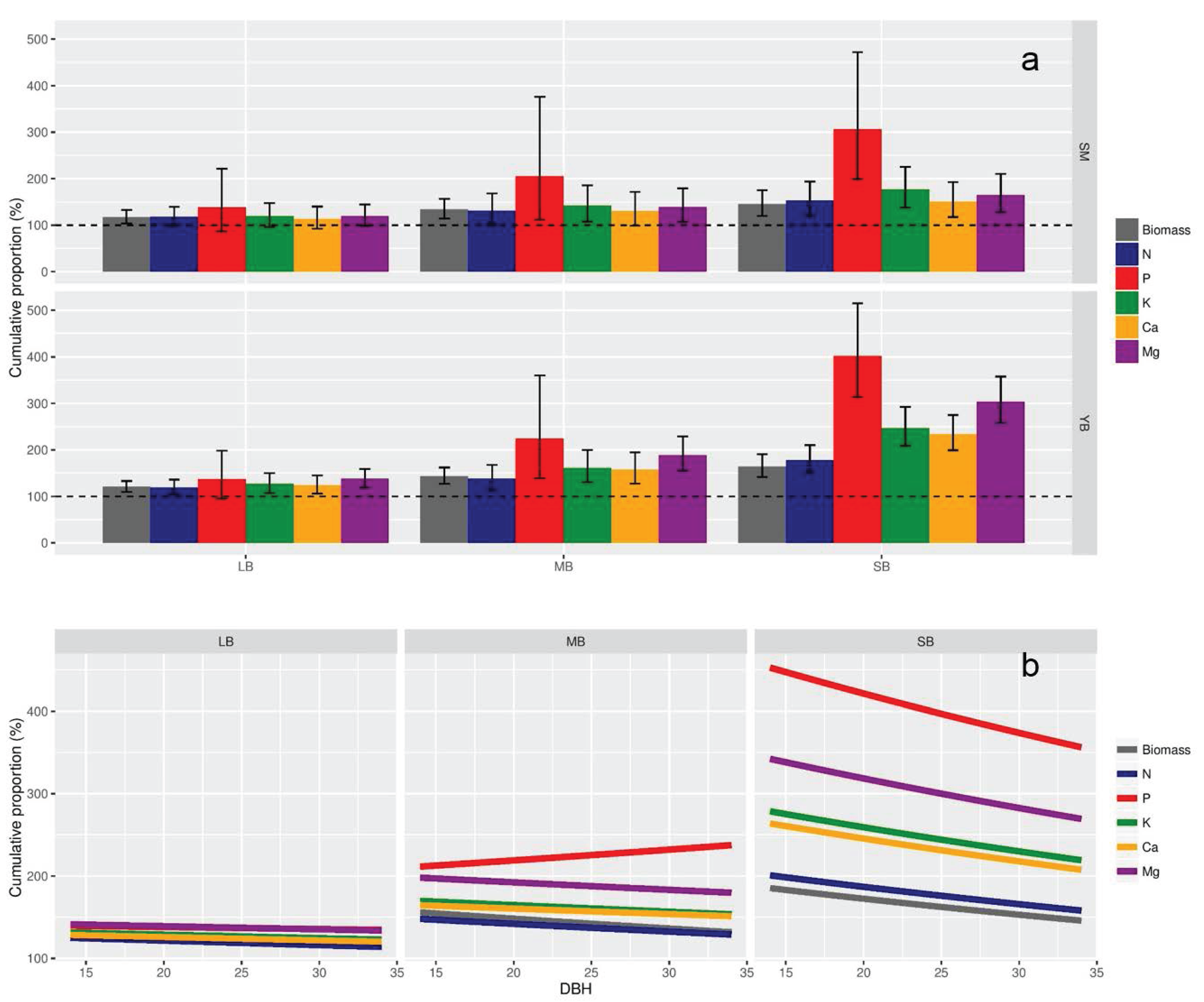
2.4. Cumulative Proportions of Nutrient and Biomass in Branch Compartments
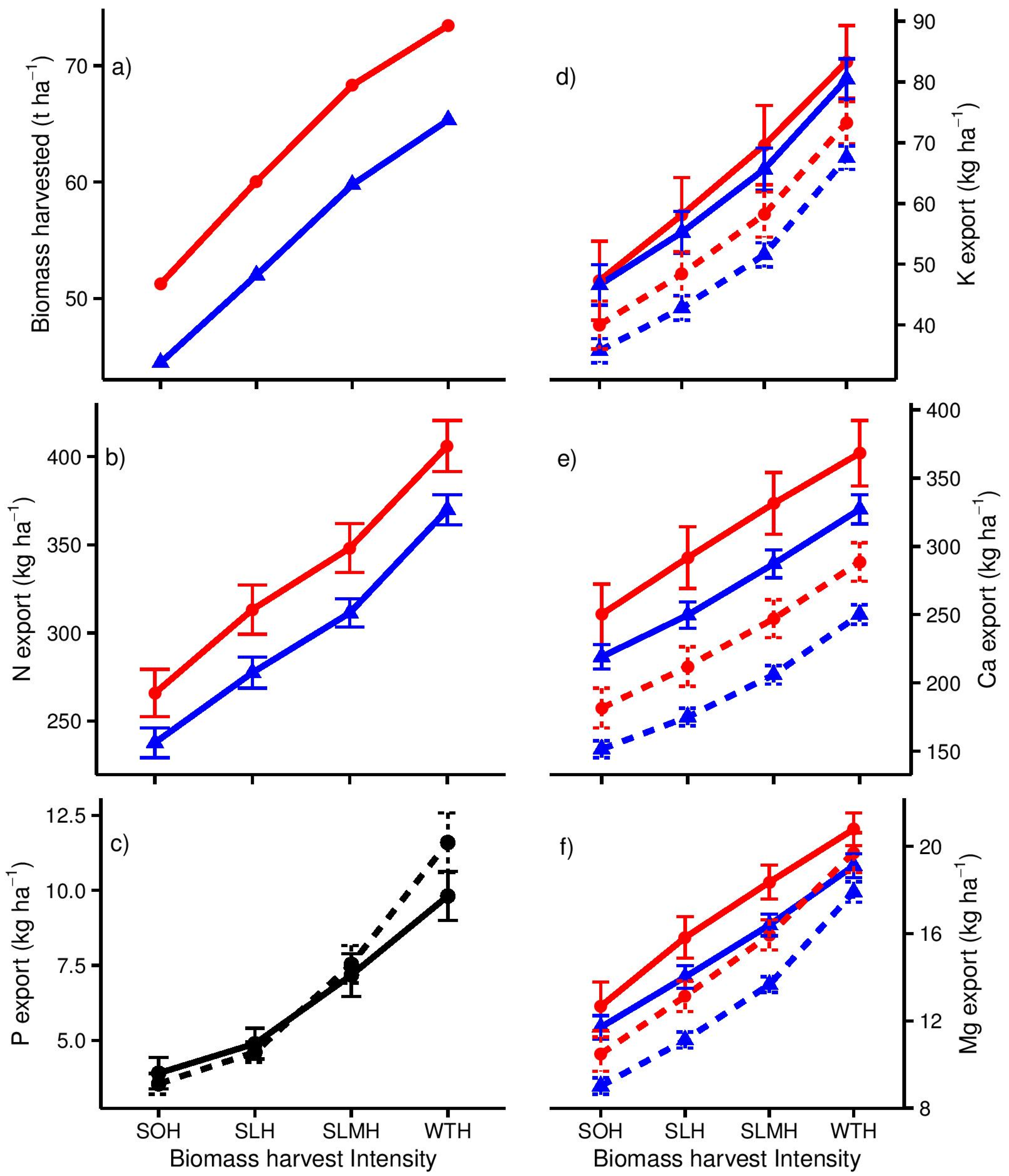
2.5. Modelling Nutrient Removal with Harvest
2.6. Statistical Analyses
3. Results
3.1. Cumulative Proportion of Nutrients and Biomass at the Tree Scale
3.2. Nutrient Export with Different Biomass Harvest Scenarios
4. Discussion
4.1. Defining a Branch Diameter Threshold Based on Nutrient Distribution at the Tree Scale
4.2. Estimating Nutrient Export with Harvest at the Stand Scale
4.3. Implications on Nutrient Pools and Future Forest Productivity
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paré, D.; Bernier, P.; Thiffault, E.; Titus, B.D. The potential of forest biomass as an energy supply for Canada. For. Chron. 2011, 87, 71–76. [Google Scholar] [CrossRef]
- Lattimore, B.; Smith, C.T.; Titus, B.D.; Stupak, I.; Egnell, G. Environmental factors in woodfuel production: Opportunities, risks, and criteria and indicators for sustainable practices. Biomass Bioenergy 2009, 33, 1321–1342. [Google Scholar] [CrossRef]
- Thiffault, E.; Hannam, K.D.; Pare, D.; Titus, B.D.; Hazlett, P.W.; Maynard, D.G.; Brais, S. Effects of forest biomass harvesting on soil productivity in boreal and temperate forests—A review. Environ. Rev. 2011, 19, 278–309. [Google Scholar] [CrossRef]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth—A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Achat, D.L.; Fortin, M.; Landmann, G.; Ringeval, B.; Augusto, L. Forest soil carbon is threatened by intensive biomass harvesting. Sci. Rep. 2015, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Fahey, T.J.; Hill, M.O.; Stevens, P.A.; Hornung, M.; Rowland, P. Nutrient accumulation in vegetation following conventional and whole-tree harvest of Sitka spruce plantations in North Wales. Forestry 1991, 64, 271–288. [Google Scholar] [CrossRef]
- Son, Y.; Gower, S.T. Nitrogen and phosphorus distribution for 5 plantation species in southwestern Wisconsin. For. Ecol. Manag. 1992, 53, 175–193. [Google Scholar] [CrossRef]
- Paré, D.; Bernier, P.; Lafleur, B.; Titus, B.D.; Thiffault, E.; Maynard, D.G.; Guo, X.J. Estimating stand-scale biomass, nutrient contents, and associated uncertainties for tree species of Canadian forests. Can. J. For. Res. 2013, 43, 599–608. [Google Scholar] [CrossRef]
- Martin, A.R.; Gezahegn, S.; Thomas, S.C. Variation in carbon and nitrogen concentration among major woody tissue types in temperate trees. Can. J. For. Res. 2015, 45, 744–757. [Google Scholar] [CrossRef]
- Andre, F.; Ponette, Q. Comparison of biomass and nutrient content between oak (Quercus petraea) and hornbeam (Carpinus betulus) trees in a coppice-with-standards stand in Chimay (Belgium). Ann. For. Sci. 2003, 60, 489–502. [Google Scholar] [CrossRef]
- Wernsdorfer, H.; Jonard, M.; Genet, A.; Legout, A.; Nys, C.; Saint-Andre, L.; Ponette, Q. Modelling of nutrient concentrations in roundwood based on diameter and tissue proportion: Evidence for an additional site-age effect in the case of Fagus sylvatica. For. Ecol. Manag. 2014, 330, 192–204. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin, Germany, 1995; p. 506. [Google Scholar]
- Aber, J.D.; Melillo, J.M. Terrestrial Ecosystems, 2nd ed.; Harcourt Academic Press: New York, NY, USA, 2001; p. 556. [Google Scholar]
- Yang, Y.; Yanai, R.D.; Fatemi, F.R.; Levine, C.R.; Lilly, P.J.; Briggs, R.D. Sources of variability in tissue chemistry in northern hardwood species. Can. J. For. Res. 2016, 46, 285–296. [Google Scholar] [CrossRef]
- Nyland, R.D. Selection system and its application to uneven-aged northern hardwoods. In Managing Northern Hardwoods: Proceedings of a Silvicultural Symposium, 23–25 June 1986, State University of New York, College of Environmental Science and Forestry, Syracuse, New York; Nyland, R.D., Ed.; SUNY College of Environmental Science and Forestry: New York, NY, USA, 1987; pp. 49–80. [Google Scholar]
- Nolet, P.; Doyon, F.; Messier, C. A new silvicultural approach to the management of uneven-aged northern hardwoods: Frequent low-intensity harvesting. Forestry 2014, 87, 39–48. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.-P. Paysages Régionaux du Québec Méridional; Publications du Québec: Sainte-Foy, QC, Canada, 1998; p. 213. [Google Scholar]
- Environment Canada. Canadian Climate Normals: 1981–2010 Climate Normals & Averages. Available online: http://climate.weather.gc.ca/climate_normals/index_e.html (accessed on 4 December 2016).
- Lajoie, P.G. Étude Pédologique des comtés de Gatineau et Pontiac, Québec; Service des recherches, Ministère de l’Agriculture du Canada, Ministère de l’Agriculture du Québec et Collège Macdonald, Université McGill: Montréal, QC, Canada, 1962. [Google Scholar]
- Lambert, M.C.; Ung, C.H.; Raulier, F. Canadian national tree aboveground biomass equations. Can. J. For. Res. 2005, 35, 1996–2018. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Zuur, A.F. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; p. 574. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Nlme: Linear and Nonlinear Mixed Effects Models, R Package Version 3.1-128; 2016. Available online: http://CRAN.R-project.org/package=nlme (accessed on 21 October 2016).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Yanai, R.D. The effect of whole-tree harvest on phosphorus cycling in a northern hardwood forest. For. Ecol. Manag. 1998, 104, 281–295. [Google Scholar] [CrossRef]
- Tremblay, S.; Ouimet, R.; Houle, D.; Duchesne, L. Base cation distribution and requirement of three common forest ecosystems in eastern Canada based on site-specific and general allometric equations. Can. J. For. Res. 2012, 42, 1796–1809. [Google Scholar] [CrossRef]
- Arthur, M.A.; Siccama, T.G.; Yanai, R.D. Calcium and magnesium in wood of northern hardwood forest species: Relations to site characteristics. Can. J. For. Res. 1999, 29, 339–346. [Google Scholar] [CrossRef]
- Park, B.B.; Yanai, R.D. Nutrient concentrations in roots, leaves and wood of seedling and mature sugar maple and American beech at two contrasting sites. For. Ecol. Manag. 2009, 258, 1153–1160. [Google Scholar] [CrossRef]
- Vadeboncoeur, M.A.; Hamburg, S.P.; Yanai, R.D.; Blum, J.D. Rates of sustainable forest harvest depend on rotation length and weathering of soil minerals. For. Ecol. Manag. 2014, 318, 194–205. [Google Scholar] [CrossRef]
- Watmough, S.A.; Dillon, P.J. Base cation and nitrogen budgets for seven forested catchments in central Ontario, 1983–1999. For. Ecol. Manag. 2003, 177, 155–177. [Google Scholar] [CrossRef]
- Federer, C.A.; Hornbeck, J.W.; Tritton, L.M.; Martin, C.W.; Pierce, R.S.; Smith, C.T. Long-term depletion of calcium and other nutrients in eastern United States forests. Environ. Manag. 1989, 13, 593–601. [Google Scholar] [CrossRef]
- Houle, D.; Paquin, R.; Camire, C.; Ouimet, R.; Duchesne, L. Response of the Lake Clair watershed (Duchesnay, Quebec) to changes in precipitation chemistry (1988–1994). Can. J. For. Res. 1997, 27, 1813–1821. [Google Scholar] [CrossRef]
- Watmough, S.A.; Aherne, J.; Alewell, C.; Arp, P.; Bailey, S.; Clair, T.; Dillon, P.; Duchesne, L.; Eimers, C.; Fernandez, I.; et al. Sulphate, nitrogen and base cation budgets at 21 forested catchments in Canada, the United States and Europe. Environ. Monit. Assess. 2005, 109, 1–36. [Google Scholar] [PubMed]
- Ouimet, R.; Duchesne, L. Base cation mineral weathering and total release rates from soils in three calibrated forest watersheds on the Canadian Boreal Shield. Can. J. Soil Sci. 2005, 85, 245–260. [Google Scholar] [CrossRef]
- Phillips, T.; Watmough, S.A. A nutrient budget for a selection harvest: Implications for long-term sustainability. Can. J. For. Res. 2012, 42, 2064–2077. [Google Scholar] [CrossRef]
- Gradowski, T.; Thomas, S.C. Phosphorus limitation of sugar maple growth in central Ontario. For. Ecol. Manag. 2006, 226, 104–109. [Google Scholar] [CrossRef]
- Vadeboncoeur, M.A. Meta-analysis of fertilization experiments indicates multiple limiting nutrients in northeastern deciduous forests. Can. J. For. Res. 2010, 40, 1766–1780. [Google Scholar] [CrossRef]
- Akselsson, C.; Westling, O.; Alveteg, M.; Thelin, G.; Fransson, A.M.; Hellsten, S. The influence of N load and harvest intensity on the risk of P limitation in Swedish forest soils. Sci. Total Environ. 2008, 404, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Vanguelova, E.; Pitman, R.; Luiro, J.; Helmisaari, H.S. Long term effects of whole tree harvesting on soil carbon and nutrient sustainability in the UK. Biogeochemistry 2010, 101, 43–59. [Google Scholar] [CrossRef]
- Zetterberg, T.; Olsson, B.A.; Löfgren, S.; Hyvönen, R.; Brandtberg, P.-O. Long-term soil calcium depletion after conventional and whole-tree harvest. For. Ecol. Manag. 2016, 369, 102–115. [Google Scholar] [CrossRef]
- Thiffault, E.; Bechard, A.; Pare, D.; Allen, D. Recovery rate of harvest residues for bioenergy in boreal and temperate forests: A review. Wiley Interdiscip. Rev. Energy Environ. 2015, 4, 429–451. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Tree Species | Compartment | Biomass (kg) | ||||
| 12–17 cm | 18–23 cm | 24–29 cm | 30–35 cm | |||
| SM | Stem bark | 11 (1.4) | 21.1 (3.6) | 43.8 (11.7) | 64.3 (15.9) | |
| Stem wood | 63.8 (8.2) | 127.4 (23) | 286.9 (79.4) | 442.9 (113.1) | ||
| Branches > 10 cm | 7.0 (0.6) | 11.0 (1.4) | 13.9 (1.6) | 14.1 (0.7) | ||
| Large branches | 22.8 (8.5) | 21.3 (8.9) | 41.3 (13.4) | 94.4 (41) | ||
| Medium branches | 14.6 (4.1) | 28.1 (8.4) | 50.4 (14.5) | 78.2 (14.6) | ||
| Small branches | 14.3 (3.9) | 18.9 (5.2) | 36.2 (11.6) | 44.7 (13) | ||
| YB | Stem bark | 6.8 (2.9) | 17.9 (4.2) | 37.7 (5.1) | 58.9 (10) | |
| Stem wood | 53.6 (22.4) | 137 (31.7) | 282.3 (38.9) | 432.5 (73.8) | ||
| Branches > 10 cm | 5.6 (1.6) | 9.5 (1.5) | 13.0 (1.7) | 14.5 (1.7) | ||
| Large branches | 20.3 (6.2) | 29.9 (14.4) | 56.3 (34.6) | 72.3 (16.1) | ||
| Medium branches | 18.3 (6.9) | 39.1 (9.3) | 61.3 (12.2) | 86.8 (28.3) | ||
| Small branches | 18.9 (5.1) | 39.4 (7.9) | 57.2 (10.3) | 65.9 (14) | ||
| Tree Species | Compartment | Nutrient Concentration (mg/kg) | ||||
| N | P | K | Ca | Mg | ||
| SM | Stem bark | 9833 (961) | 153 (64) | 3396 (857) | 27,500 (4054) | 855 (157) |
| Stem wood | 4849 (681) | 75 (57) | 742 (323) | 1425 (606) | 182 (54) | |
| Large branches | 5330 (633) | 146 (96) | 1114 (286) | 3775 (1382) | 280 (71) | |
| Medium branches | 4107 (1578) | 309 (235) | 1373 (210) | 4713 (1492) | 306 (65) | |
| Small branches | 9890 (3448) | 638 (405) | 3017 (369) | 8296 (1201) | 559 (117) | |
| YB | Stem bark | 10,301 (947) | 107 (65) | 1304 (359) | 10,784 (3968) | 355 (94) |
| Stem wood | 4747 (734) | 93 (62) | 478 (138) | 748 (239) | 117 (36) | |
| Large branches | 4923 (940) | 126 (88) | 718 (280) | 2243 (1475) | 255 (72) | |
| Medium branches | 4355 (1633) | 335 (268) | 823 (145) | 2704 (1054) | 308 (53) | |
| Small branches | 10,612 (4518) | 671 (392) | 2204 (329) | 6333 (1903) | 743 (158) | |
| Branch Compartment | Main Effects and Interactions | Pairwise Comparisons with Biomass Proportion | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Term | numDF | denDF | F-Value | p-Value | Nutrient | Sugar Maple | Yellow birch | |||
| t-Value | p-Value | t-Value | p-Value | |||||||
| Large branches (LB) | (Intercept) | 1 | 225 | 133,196.56 | 0.0000 | N | −1.473 | 0.1420 | −2.190 | 0.0296 |
| BP | 5 | 225 | 9.37 | 0.0000 | P | 0.500 | 0.6178 | 0.434 | 0.6647 | |
| DBH | 1 | 45 | 6.69 | 0.0130 | K | −1.221 | 0.2235 | −0.281 | 0.7792 | |
| sp | 1 | 45 | 2.04 | 0.1604 | Ca | −3.055 | 0.0025 | −1.081 | 0.2810 | |
| BP:DBH | 5 | 225 | 3.42 | 0.0054 | Mg | −2.427 | 0.0160 | 2.477 | 0.0140 | |
| BP:sp | 5 | 225 | 8.40 | 0.0000 | ||||||
| Medium branches (MB) | (Intercept) | 1 | 225 | 91,335.32 | 0.0000 | N | −1.577 | 0.1162 | −1.970 | 0.0501 |
| BP | 5 | 225 | 37.49 | 0.0000 | P | 0.488 | 0.6262 | 0.591 | 0.5550 | |
| DBH | 1 | 45 | 7.63 | 0.0083 | K | −0.540 | 0.5899 | 0.787 | 0.4323 | |
| sp | 1 | 45 | 8.64 | 0.0052 | Ca | −2.868 | 0.0045 | −0.081 | 0.9353 | |
| BP:DBH | 5 | 225 | 2.96 | 0.0131 | Mg | −1.316 | 0.1895 | 4.369 | 0.0000 | |
| BP:sp | 5 | 225 | 17.33 | 0.0000 | ||||||
| Small branches (SB) | (Intercept) | 1 | 230 | 64,708.90 | 0.0000 | N | 2.825 | 0.0051 | 3.081 | 0.0023 |
| BP | 5 | 230 | 97.83 | 0.0000 | P | 7.497 | 0.0000 | 8.676 | 0.0000 | |
| DBH | 1 | 45 | 20.48 | 0.0000 | K | 9.641 | 0.0000 | 13.449 | 0.0000 | |
| sp | 1 | 45 | 15.70 | 0.0003 | Ca | 2.726 | 0.0069 | 7.962 | 0.0000 | |
| BP:sp | 5 | 230 | 37.68 | 0.0000 | Mg | 7.781 | 0.0000 | 15.241 | 0.0000 | |
| Terms in the Model | Df | Biomass | N | P | |||
| Variance Explained | F-Value | Variance Explained | F-Value | Variance Explained | F-Value | ||
| Intensity | 3 | 80.39 | 1,007,521 | 85.24 | 111,459.57 | 91.98 | 96,438.34 |
| Composition | 1 | 0.02 | 855 | 0.14 | 554.64 | 0.45 | 1407.89 |
| Marking | 1 | 18.73 | 704,362 | 10.08 | 39,521.46 | 0.01 | 21.79 |
| Intensity: Composition | 3 | 0.15 | 1929 | 0.32 | 422.60 | 2.32 | 2437.55 |
| Intensity: Marking | 3 | 0.13 | 1686 | 0.11 | 140.74 | 0.12 | 126.88 |
| Composition: Marking | 1 | 0.13 | 4850 | 0.03 | 104.35 | 0.00 | 11.02 |
| Intensity: Composition: Marking | 3 | 0.01 | 80 | 0.01 | 10.23 | 0.04 | 38.76 |
| Residuals | 15,984 | 0.43 | 4.07 | 5.08 | |||
| Terms in the Model | Df | K | Ca | Mg | |||
| Variance Explained | F-Value | Variance Explained | F-Value | Variance Explained | F-Value | ||
| Intensity | 3 | 74.69 | 48,739.90 | 43.98 | 39,680.22 | 78.75 | 110,919.35 |
| Composition | 1 | 14.57 | 28,523.50 | 39.88 | 107,947.95 | 10.15 | 42,897.88 |
| Marking | 1 | 1.98 | 3869.75 | 9.88 | 26,734.00 | 6.18 | 26,133.32 |
| Intensity: Composition | 3 | 0.21 | 134.86 | 0.19 | 174.58 | 0.85 | 1190.45 |
| Intensity: Marking | 3 | 0.12 | 80.59 | 0.14 | 124.86 | 0.23 | 321.99 |
| Composition: Marking | 1 | 0.26 | 508.56 | 0.02 | 40.66 | 0.04 | 175.67 |
| Intensity: Composition: Marking | 3 | 0.00 | 2.36 | 0.00 | 3.30 | 0.01 | 14.84 |
| Residuals | 15,984 | 8.17 | 5.91 | 3.78 | |||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Royer-Tardif, S.; Delagrange, S.; Nolet, P.; Rivest, D. Using Macronutrient Distributions within Trees to Define a Branch Diameter Threshold for Biomass Harvest in Sugar Maple-Dominated Stands. Forests 2017, 8, 41. https://doi.org/10.3390/f8020041
Royer-Tardif S, Delagrange S, Nolet P, Rivest D. Using Macronutrient Distributions within Trees to Define a Branch Diameter Threshold for Biomass Harvest in Sugar Maple-Dominated Stands. Forests. 2017; 8(2):41. https://doi.org/10.3390/f8020041
Chicago/Turabian StyleRoyer-Tardif, Samuel, Sylvain Delagrange, Philippe Nolet, and David Rivest. 2017. "Using Macronutrient Distributions within Trees to Define a Branch Diameter Threshold for Biomass Harvest in Sugar Maple-Dominated Stands" Forests 8, no. 2: 41. https://doi.org/10.3390/f8020041