
The Change in Growth, Osmolyte Production and Antioxidant Enzymes Activity Explains the Cadmium Tolerance in Four Tree Species at the Saplings Stage

 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Cadmium Application
2.3. Growth and Dry Biomass Production
2.4. Physiological Parameters
2.5. Measurement of Cd Concentration in Different Plant Organs (Leaves, Stem and Root)
2.6. Measurement of the Translocation Factor
2.7. Measurement of Hydrogen Peroxide (H2O2), Superoxide Radical (O2–) and Osmolytes Accumulation
2.8. Measurement of Antioxidant Enzymes
2.9. Statistical Analysis
3. Results
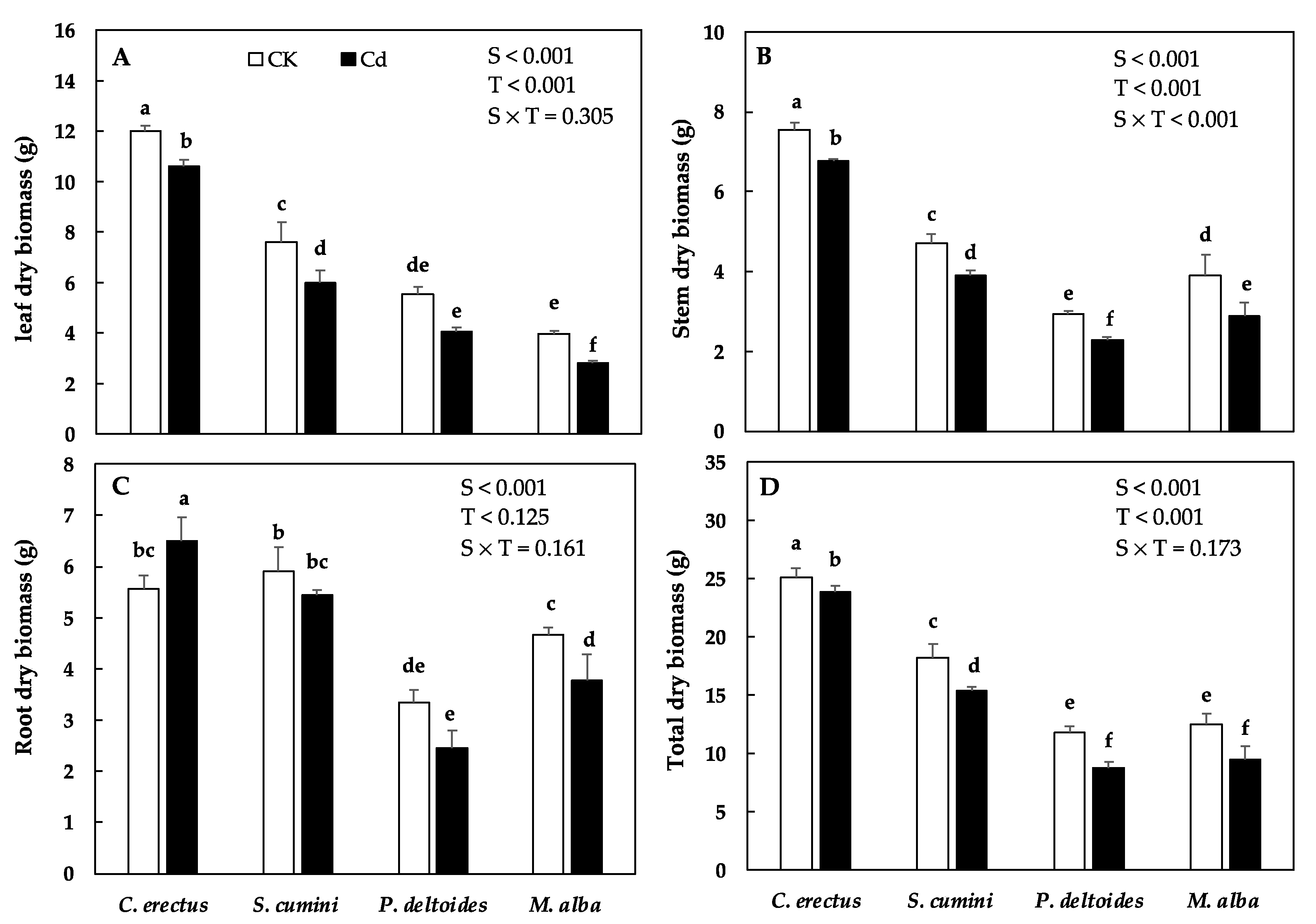
3.1. Cd Treatment and Growth and Biomass Production
3.2. Cd Treatment and Chl a, b and Carotenoid Contents
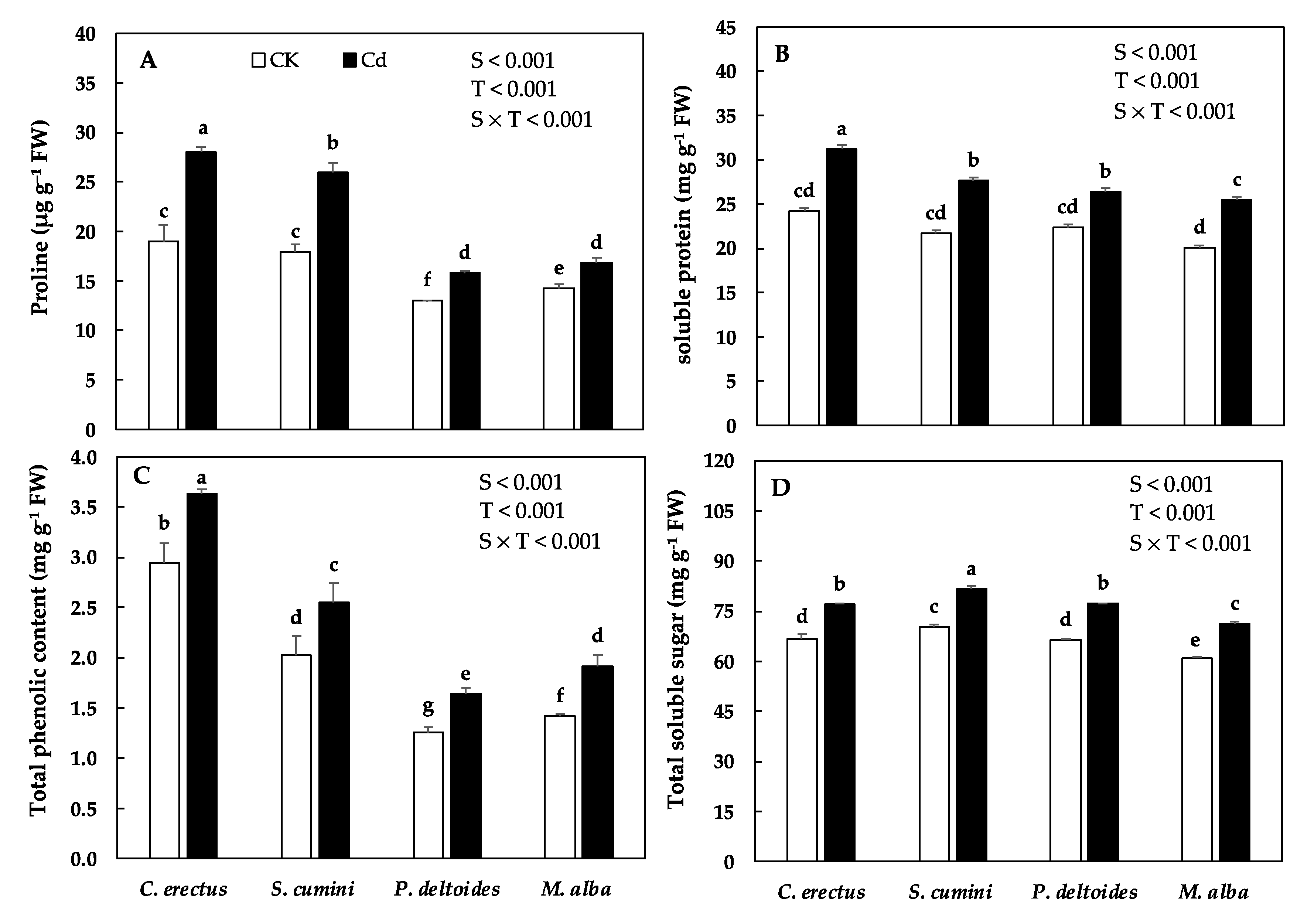
3.3. Cd Treatment and Proline and Osmolytes Accumulation
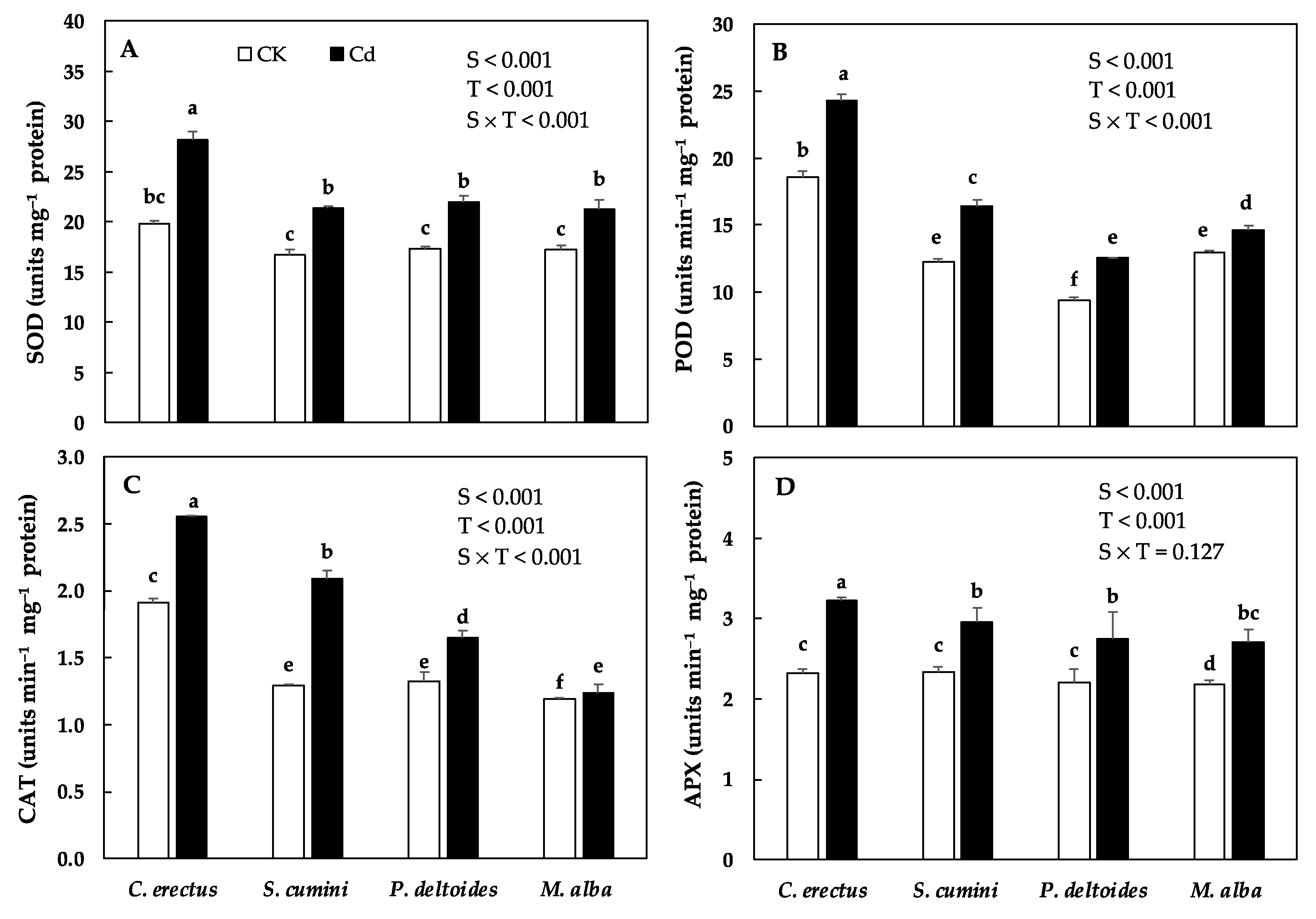
3.4. Cd Treatment and Oxidants and Antioxidants Enzyme Activity
3.5. Content of Cd in Various Plant Organs and the Translocation Factor
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, J.; Liu, C.; Hsu, P.C.; Zhao, J.; Wu, T.; Tang, J.; Liu, K.; Cui, Y. Remediation of heavy metal contaminated soil by asymmetrical alternating current electrochemistry. Nat. Commun. 2019, 10, 2440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Exp. Suppl. 2012, 101, 133–164. [Google Scholar] [PubMed] [Green Version]
- Xu, Z.; Guo, Z.; Xiao, X.; Zeng, P.; Xue, Q. Effect of inorganic potassium compounds on the hydrothermal carbonization of Cd-contaminated rice straw for experimental scale hydrochar. Biomass Bioenergy 2019, 130, 105357. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Khan, S.; Brusseau, M.L.; Shah, M.T. Lead and cadmium contamination and exposure risk assessment via consumption of vegetables grown in agricultural soils of five-selected regions of Pakistan. Chemosphere 2017, 168, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhou, K.; Qin, W.; Tian, C.; Qi, M.; Yan, X.; Han, W. A review on heavy metals contamination in soil: Effects, sources, and remediation techniques. Soil Sediment Contam. Int. J. 2019, 28, 380–394. [Google Scholar] [CrossRef]
- Kramer, U. Metal hyperaccumulation in plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry (ATSDR). Available online: https://wwwn.cdc.gov/TSP/substances/ToxSubstance.aspx?toxid=15 (accessed on 15 May 2022).
- Rahim, H.U.; Akbar, W.A.; Alatalo, J.M. A Comprehensive Literature Review on Cadmium (Cd) Status in the Soil Environment and Its Immobilization by Biochar-Based Materials. Agronomy 2022, 12, 877. [Google Scholar] [CrossRef]
- Mohsin, M.; Salam, M.M.A.; Nawrot, N.; Kaipiainen, E.; Lane, D.J.; Wojciechowska, E.; Kinnunen, N.; Heimonen, M.; Tervahauta, A.; Peräniemi, S.; et al. Phytoextraction and recovery of rare earth elements using willow (Salix spp.). Sci. Total Environ. 2022, 809, 152209. [Google Scholar] [CrossRef]
- Kaur, B.; Singh, B.; Kaur, N.; Singh, D. Phytoremediation of cadmium-contaminated soil through multipurpose tree species. Agrofor. Syst. 2018, 92, 473–483. [Google Scholar] [CrossRef]
- Peuke, A.D.; Heinz, R. Phytoremediation Molecular Biology, Requirements for Application, Environmental Protection, Public Attention and Feasibility. EMBO Rep. 2005, 6, 497–501. [Google Scholar] [CrossRef]
- Mohsin, M.; Kaipiainen, E.; Salam, M.M.A.; Evstishenkov, N.; Nawrot, N.; Villa, A.; Wojciechowska, E.; Kuittinen, S.; Pappinen, A. Biomass Production and Removal of Nitrogen and Phosphorus from Processed Municipal Wastewater by Salix schwerinii: A Field Trial. Water 2021, 13, 2298. [Google Scholar] [CrossRef]
- Ahmad, Z.; Khan, S.M.; Ali, M.I.; Fatima, N.; Ali, S. Pollution in cadmium and marble waste polluted ecosystem; role of selected indicator plants in phytoremediation and determination of pollution zones. J. Clean. Prod. 2019, 236, 117709. [Google Scholar] [CrossRef]
- Luo, J.; Yin, D.; Cheng, H.; Davison, W.; Zhang, H. Plant induced changes to rhizosphere characteristics affecting supply of Cd to Noccaea caerulescens and Ni to Thlaspi goes in gense. Environ. Sci. Technol. 2018, 52, 5085–5093. [Google Scholar] [CrossRef] [PubMed]
- El Rasafi, T.; Oukarroum, A.; Haddioui, A.; Song, H.; Kwon, E.E.; Bolan, N.; Tack, F.M.; Sebastian, A.; Prasad, M.N.V.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Environ. Sci. Technol. 2021, 52, 1–52. [Google Scholar] [CrossRef]
- Fine, P.; Rathod, P.H.; Beriozkin, A.; Mingelgrin, U. Uptake of cadmium by hydroponically grown, mature Eucalyptus camaldulensis saplings and the effect of organic ligands. Int. J. Phytoremed. 2013, 15, 585–601. [Google Scholar] [CrossRef]
- Vassilev, A.; Perez-Sanz, A.; Semane, B.; Carleer, R.; Vangronsveld, J. Cadmium accumulation and tolerance of two Salix genotypes hydroponically grown in presence of Cadmium. J. Plant. Nutr. 2005, 28, 2159–2177. [Google Scholar] [CrossRef]
- Faroon, O.; Ashizawa, A.; Wright, S.; Tucker, P.; Jenkins, K.; Ingerman, L.; Rudisill, C. Toxicological Profile of Cadmium; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2012; pp. 273–274. [Google Scholar]
- Hussain, Z.; Rasheed, F.; Tanvir, M.A.; Zafar, Z.; Rafay, M.; Mohsin, M.; Pulkkinen, P.; Ruffner, C. Increased antioxidative enzyme activity mediates the phytoaccumulation potential of Pb in four agroforestry tree species: A case study under municipal and industrial wastewater irrigation. Int. J. Phytoremed. 2020, 23, 704–714. [Google Scholar] [CrossRef]
- Bashir, A.; Rizwan, M.; Ali, S.; Adress, M.; Qayyum, M.F. Effect of composted organic amendments and zinc oxide nanoparticles on growth and cadmium accumulation by wheat; a life cycle study. Environ. Sci. Pollut. Res. 2020, 27, 23926–23936. [Google Scholar] [CrossRef]
- Iori, V.; Pietrini, F.; Zacchini, M. Assessment of ibuprofen tolerance and removal capability in Populus nigra L. by in vitro culture. J. Hazard Mater. 2012, 229, 217–223. [Google Scholar] [CrossRef]
- Zhao, Z.; Xi, M.; Jiang, G.; Liu, X.; Bai, Z.; Huang, Y. Effects of IDSA, EDDS and EDTA on heavy metals accumulation in hydroponically grown maize (Zea mays L.). J. Hazard Mater. 2010, 181, 455–459. [Google Scholar] [CrossRef]
- He, F.; Zhao, Q.; Huang, J.L.; Niu, M.X.; Feng, H.C.; Shi, Y.J.; Zhao, K.J.; Cui, X.L.; Wu, X.L.; Mi, J.X.; et al. External application of nitrogen alleviates toxicity of cadmium on poplars via starch and sucrose metabolism. Tree Physiol. 2021, 41, 2126–2141. [Google Scholar] [CrossRef]
- Si, L.; Zhang, J.; Hussain, A.; Qiao, Y.; Zhou, J.; Wang, X. Accumulation, and translocation of food chain in soil-mulberry (Morus alba L.)-silkworm (Bombyx mori) under single and combined stress of lead and cadmium. Ecotoxicol. Environ. Saf. 2021, 208, 111582. [Google Scholar] [CrossRef]
- Rasheed, F.; Zafar, Z.; Waseem, Z.A.; Rafay, M.; Abdullah, M.; Salam, M.M.A.; Mohsin, M.; Khan, R.W. Phytoaccumulation of Zn, Pb, and Cd in Conocarpus lancifolius irrigated with wastewater: Does physiological response influence heavy metal uptake? Int. J. Phytoremed. 2020, 22, 287–294. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zafar, Z.; Rasheed, F.; Delagrange, S.; Abdullah, M.; Ruffner, C. Acclimatization of Terminalia arjuna saplings to salt stress: Characterization of growth, biomass, and photosynthetic parameters. J. Sustain. For. 2020, 39, 76–91. [Google Scholar] [CrossRef]
- Guo, B.; Dai, S.; Wang, R.; Guo, J.; Ding, Y.; Xu, Y. Combined effects of elevated CO2 and Cd-contaminated soils on the growth, gas exchange, antioxidants defense and Cd accumulation of poplars and willows. Environ. Exp. Bot. 2015, 115, 1–10. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bai, T.; Li, C.; Ma, F.; Feng, F.; Shu, H. Responses of growth and antioxidant system to root-zone hypoxia stress in two Malus species. Plant Soil 2010, 327, 95–105. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Bayer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Method. Enzymol. 1955, 2, 764–775. [Google Scholar]
- Knörzer, O.C.; Burner, J.; Boger, P. Alterations in the antioxidative system of suspension-cultured soybean cells (Glycine max) induced by oxidative stress. Physiol. Plant. 1996, 97, 388–396. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Liu, L.; Li, W.; Song, W.; Guo, M. Remediation techniques for heavy metal-contaminated soils: Principles and applicability. Sci. Total Environ. 2018, 633, 206–219. [Google Scholar] [CrossRef]
- Bae, J.; Benoit, D.L.; Watson, A.K. Effect of heavy metals on seed germination and seedling growth of common ragweed and roadside ground cover legumes. Environ. Pollut. 2016, 213, 112–118. [Google Scholar] [CrossRef]
- Wu, F.; Yang, W.; Zhang, J.; Zhou, L. Cadmium accumulation and growth responses of a poplar (Populus deltoides × Populus nigra) in cadmium contaminated purple soil and alluvial soil. J. Hazard Mater. 2010, 177, 268–273. [Google Scholar] [CrossRef]
- Zacchini, M.; Pietrini, F.; Scarascia Mugnozza, G.; Iori, V.; Pietrosanti, L.; Massacci, A. Metal tolerance accumulation and translocation in poplar and willow clones treated with cadmium in hydroponics. Water Air Soil Pollut. 2009, 197, 23–34. [Google Scholar] [CrossRef]
- Guerra, F.; Gainza, F.; Perez, R.; Zamudio, F. Phytoremediation of heavy metals using poplars (Populus spp.): A glimpse of the plant responses to copper, cadmium and zinc stress. In Handbook of Phytoremediation; Golubev, I.A., Ed.; Nova Science: New York, NY, USA, 2011; pp. 387–413. [Google Scholar]
- Zacchini, M.; Iori, V.; Scarascia-Mugnozza, G.; Pietrini, F.; Massacci, A. Cadmium accumulation and tolerance in Populus nigra and Salix alba. Biol. Plant. 2011, 55, 383–386. [Google Scholar] [CrossRef]
- Pietrini, F.; Iori, V.; Bianconi, D.; Mughini, G.; Massacci, A.; Zacchini, M. Assessment of physiological and biochemical responses, metal tolerance and accumulation in two eucalyptus hybrid clones for phytoremediation of cadmium-contaminated waters. J. Environ. Manag. 2015, 162, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, P.; Planchon, S.; Oufir, M.; Ziebel, J.; Dommes, J.; Hoffmann, L.; Hausman, J.F.; Renaut, J. Combining proteomics and metabolite analyses to unravel cadmium stress-response in poplar leaves. J. Proteome Res. 2009, 8, 400–417. [Google Scholar] [CrossRef] [PubMed]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Lannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Shabir, R.; Abbas, G.; Saqib, M.; Shahid, M.; Shah, G.M.; Akram, M.; Niazi, N.K.; Naeem, M.A.; Hussain, M.; Ashraf, F. Cadmium tolerance and phytoremediation potential of acacia (Acacia nilotica L.) under salinity stress. Int. J. Phytoremed. 2017, 20, 739–746. [Google Scholar] [CrossRef]
- Tőzsér, D.; Magura, T.; Simon, E. Heavy metal uptake by plant parts of willow species: A meta-analysis. J. Hazard. Mater. 2017, 336, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Pietrini, F.; Zacchini, M.; Iori, V.; Pietrosanti, L.; Bianconi, D.; Massacci, A. Screening of poplar clones for cadmium phytoremediation using photosynthesis, biomass and cadmium content analyses. Int. J. Phytoremed. 2010, 12, 105–120. [Google Scholar] [CrossRef]
- Liu, H.; Wang, H.; Ma, Y.; Wang, H.; Shi, Y. Role of transpiration and metabolism in translocation and accumulation of cadmium in tobacco plants (Nicotiana tabacum L.). Chemosphere 2016, 144, 1960–1965. [Google Scholar] [CrossRef]
- Ashraf, F.; Abbas, G.; Murtaza, B.; Amjad, M.; Imran, M.; Naeem, M.A.; Saqib, M.; Niazi, N.K.; Zakir, A.; Hussain, M.; et al. Comparative tolerance and Phytostabilization Potential of Conocarpus erectus and Eucalyptus camaldulensis grown in cadmium contaminated soil. Pak. J. Agric. Sci. 2018, 55, 521–529. [Google Scholar]
- He, J.; Qin, J.; Long, L.; Ma, Y.; Li, H.; Li, K.; Jiang, X.; Liu, T.; Polle, A.; Liang, Z.; et al. Net cadmium flux and accumulation reveal tissue-specific oxidative stress and detoxification in Populus × canescens. Physiol. Plant. 2011, 143, 50–63. [Google Scholar] [CrossRef]
- Schützendübel, A.; Polle, A. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot. 2002, 53, 135. [Google Scholar] [CrossRef]
- Rodriguez-Serrano, M.; Romero-Puertas, M.C.; Sparkes, I.; Hawes, C.; del Rio, L.A.; Sandalio, L.M. Peroxisome dynamics in Arabidopsis plants under oxidative stress induced by cadmium. Free Radic. Biol. Med. 2009, 47, 1632–1639. [Google Scholar] [CrossRef]
- Heyno, E.; Klose, C.; Krieger-Liszkay, A. Origin of cadmium induced reactive oxygen species production: Mitochondrial electron transfer versus plasma membrane NADPH oxidase. New Phytol. 2008, 179, 687–699. [Google Scholar] [CrossRef]
- Sidhu, G.P.S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Tolerance and hyperaccumulation of cadmium by a wild, unpalatable herb Coronopus didymus (L.) Sm. (Brassicaceae). Ecotoxicol. Environ. Saf. 2017, 135, 209–215. [Google Scholar] [CrossRef]
- Zhang, B.L.; Shang, S.H.; Zhang, H.T.; Jabeen, Z.; Zhang, G.P. Sodium chloride enhances cadmium tolerance through reducing cadmium accumulation and increasing anti-oxidative enzyme activity in tobacco. Environ. Toxicol. Chem. 2013, 32, 1420–1425. [Google Scholar] [CrossRef]
- Zhu, J.J.; Zhang, J.L.; Liu, H.C.; Cao, K.F. Photosynthesis, non-photochemical pathways, and activities of antioxidant enzymes in a resilient evergreen oak under different climatic conditions from a valley-savanna in Southwest China. Physiol. Plant. 2009, 135, 62–72. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants, and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Ghorbani, A.; Razavi, S.M.; Omran, V.O.G.; Pirdashti, H. Piriformospora indica alleviates salinity by boosting redox poise and antioxidative potential of tomato. Russ. J. Plant Physiol. 2018, 65, 898–907. [Google Scholar]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [Green Version]
- Manousaki, E.; Kalogerakis, N. Phytoextraction of Pb and Cd by the Mediterranean saltbush (Atriplex halimus L.): Metal uptake in relation to salinity. Environ. Sci. Pollut. Res. 2009, 16, 844–854. [Google Scholar] [CrossRef]
- Zhou, J.; Cheng, K.; Zheng, J.; Liu, Z.; Shen, W.; Fan, H.; Jin, Z. Physiological and Biochemical Characteristics of Cinnamomum camphora in Response to Cu- and Cd contaminated Soil. Water Air Soil Pollut. 2019, 230, 15. [Google Scholar] [CrossRef]
- Nocito, F.F.; Lancilli, C.; Dendena, B.; Lucchini, G.; Sacchi, G.A. Cadmium retention in rice roots is influenced by cadmium availability, chelation and translocation. Plant Cell Environ. 2011, 43, 994–1008. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plant Height (cm) | Stem Diameter (mm) | No. of Leaves | Root–Shoot Ratio | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Carotenoid (mg g−1 FW) | ||
|---|---|---|---|---|---|---|---|---|
| C. erectus | CK | 65.5 ± 1.57 | 5.63 ± 0.17 | 50.7 ± 2.12 | 0.26 ± 0.01 | 1.44 ± 0.07 | 1.65 ± 0.09 | 0.90 ± 0.01 |
| Cd | 56.6 ± 1.47 | 4.96 ± 0.18 | 47.3 ± 2.09 | 0.38 ± 0.02 | 1.34 ± 0.06 | 1.45 ± 0.02 | 0.76 ± 0.01 | |
| S. cumini | CK | 50.8 ± 0.73 | 5.76 ± 0.14 | 21.5 ± 0.86 | 0.47 ± 0.01 | 1.73 ± 0.05 | 1.44 ± 0.02 | 0.92 ± 0.04 |
| Cd | 40.6 ± 1.02 | 3.89 ± 0.18 | 16.7 ± 0.51 | 0.54 ± 0.03 | 1.61 ± 0.13 | 1.31 ± 0.01 | 0.64 ± 0.01 | |
| P. deltoides | CK | 53.8 ± 1.37 | 4.17 ± 0.15 | 17.0 ± 0.37 | 0.31 ± 0.02 | 1.38 ± 0.01 | 1.36 ± 0.01 | 0.80 ± 0.01 |
| Cd | 43.6 ± 0.78 | 3.29 ± 0.32 | 12.7 ± 1.12 | 0.47 ± 0.03 | 1.18 ± 0.04 | 1.06 ± 0.09 | 0.46 ± 0.05 | |
| M. alba | CK | 46.9 ± 2.15 | 3.77 ± 0.11 | 22.5 ± 0.51 | 0.51 ± 0.05 | 1.18 ± 0.01 | 1.12 ± 0.01 | 0.71 ± 0.01 |
| Cd | 37.5 ± 0.78 | 3.14 ± 0.06 | 14.2 ± 0.71 | 0.63 ± 0.05 | 0.96 ± 0.12 | 0.95 ± 0.05 | 0.46 ± 0.05 | |
| S effect | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | |
| T effect | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | |
| S × T Effect | p = 0.988 | p = 0.030 | p = 0.529 | p = 0.173 | p = 0.218 | p < 0.001 | p = 0.096 |
| H2O2 (μmol g–1 FW) | O2– (μmol g–1 FW) | Cd Content Leaves (mg Kg−1) | Cd Content Stem (mg Kg−1) | Cd Content Root (mg Kg−1) | Translocation Factor | ||
|---|---|---|---|---|---|---|---|
| C. erectus | CK | 5.62 ± 0.80 | 1.12 ± 0.01 | 0.35 ± 0.01 | 0.79 ± 0.01 | 0.67 ± 0.03 | 1.04 ± 0.11 |
| Cd | 7.11 ± 0.62 | 1.18 ± 0.01 | 57.1 ± 2.65 | 37.0 ± 2.41 | 75.9 ± 0.05 | 1.24 ± 0.34 | |
| S. cumini | CK | 7.33 ± 0.15 | 1.16 ± 0.06 | 0.45 ± 0.02 | 0.93 ± 0.01 | 0.44 ± 0.02 | 0.97 ± 0.16 |
| Cd | 9.94 ± 0.34 | 1.28 ± 0.02 | 42.6 ± 0.67 | 20.3 ± 0.49 | 60.1 ± 0.47 | 1.03 ± 0.21 | |
| P. deltoides | CK | 8.95 ± 0.16 | 1.22 ± 0.02 | 0.55 ± 0.01 | 0.93 ± 0.01 | 0.88 ± 0.01 | 1.53 ± 0.09 |
| Cd | 11.1 ± 0.61 | 1.33 ± 0.01 | 44.5 ± 0.25 | 24.3 ± 1.57 | 67.1 ± 1.03 | 1.02 ± 0.13 | |
| M. alba | CK | 8.95 ± 0.04 | 1.27 ± 0.01 | 0.55 ± 0.01 | 0.88 ± 0.01 | 0.98 ± 0.01 | 0.73 ± 0.06 |
| Cd | 11.0 ± 0.09 | 1.59 ± 0.04 | 39.1 ± 1.93 | 22.2 ± 1.78 | 62.2 ± 2.49 | 0.98 ± 0.11 | |
| S effect | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | |
| T effect | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.977 | |
| S × T Effect | p = 0.134 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar, Z.; Rasheed, F.; Khan, W.R.; Mohsin, M.; Rashid, M.Z.; Magiman, M.M.; Raza, Z.; Rosli, Z.; Afzal, S.; Bakar, F.A. The Change in Growth, Osmolyte Production and Antioxidant Enzymes Activity Explains the Cadmium Tolerance in Four Tree Species at the Saplings Stage. Forests 2022, 13, 1343. https://doi.org/10.3390/f13091343
Zafar Z, Rasheed F, Khan WR, Mohsin M, Rashid MZ, Magiman MM, Raza Z, Rosli Z, Afzal S, Bakar FA. The Change in Growth, Osmolyte Production and Antioxidant Enzymes Activity Explains the Cadmium Tolerance in Four Tree Species at the Saplings Stage. Forests. 2022; 13(9):1343. https://doi.org/10.3390/f13091343
Chicago/Turabian StyleZafar, Zikria, Fahad Rasheed, Waseem Razzaq Khan, Muhammad Mohsin, Muhammad Zahid Rashid, Mohamad Maulana Magiman, Zohaib Raza, Zamri Rosli, Shazia Afzal, and Fauziah Abu Bakar. 2022. "The Change in Growth, Osmolyte Production and Antioxidant Enzymes Activity Explains the Cadmium Tolerance in Four Tree Species at the Saplings Stage" Forests 13, no. 9: 1343. https://doi.org/10.3390/f13091343