
Time-Lag Effect of Climate Conditions on Vegetation Productivity in a Temperate Forest–Grassland Ecotone

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
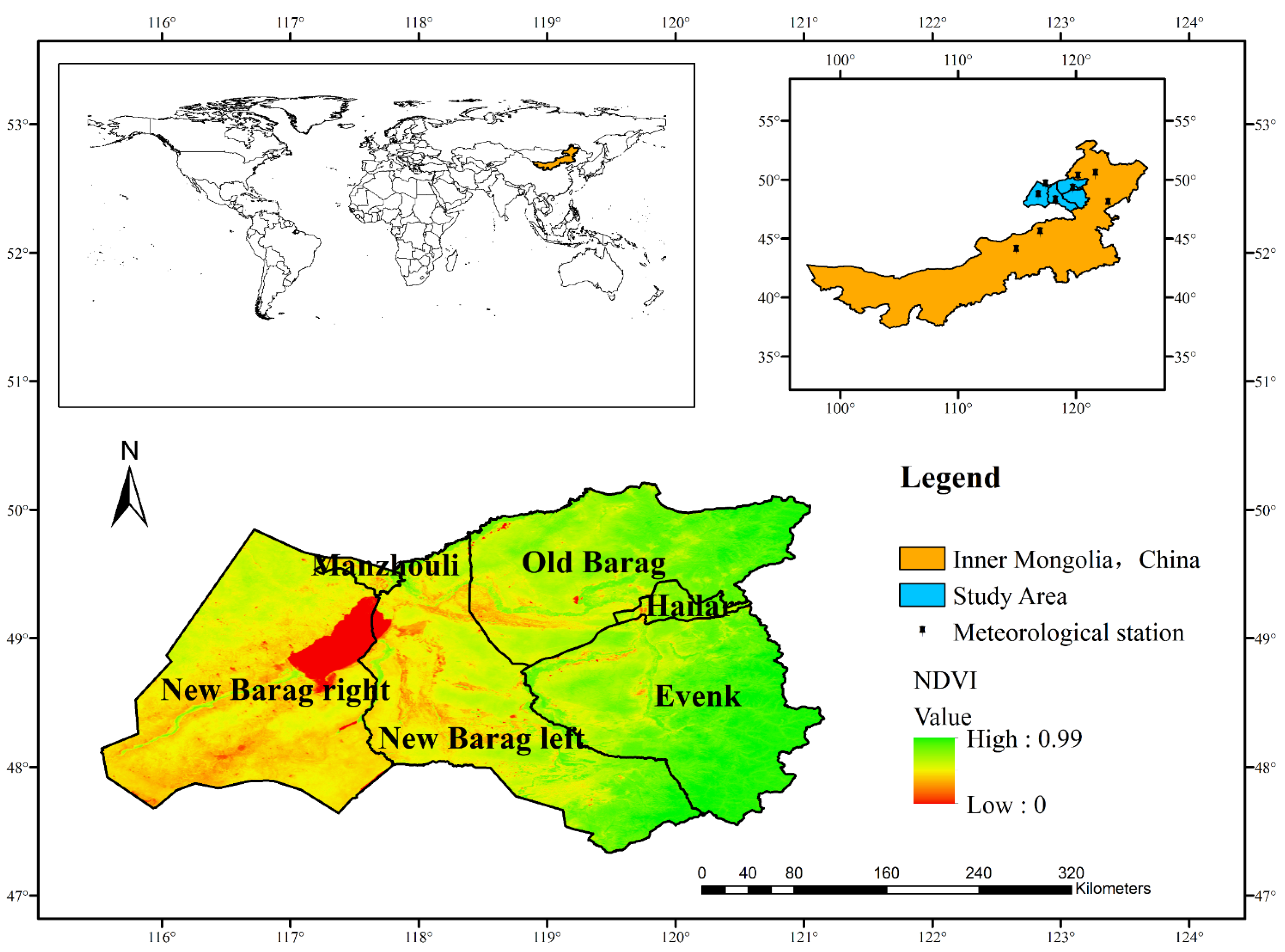
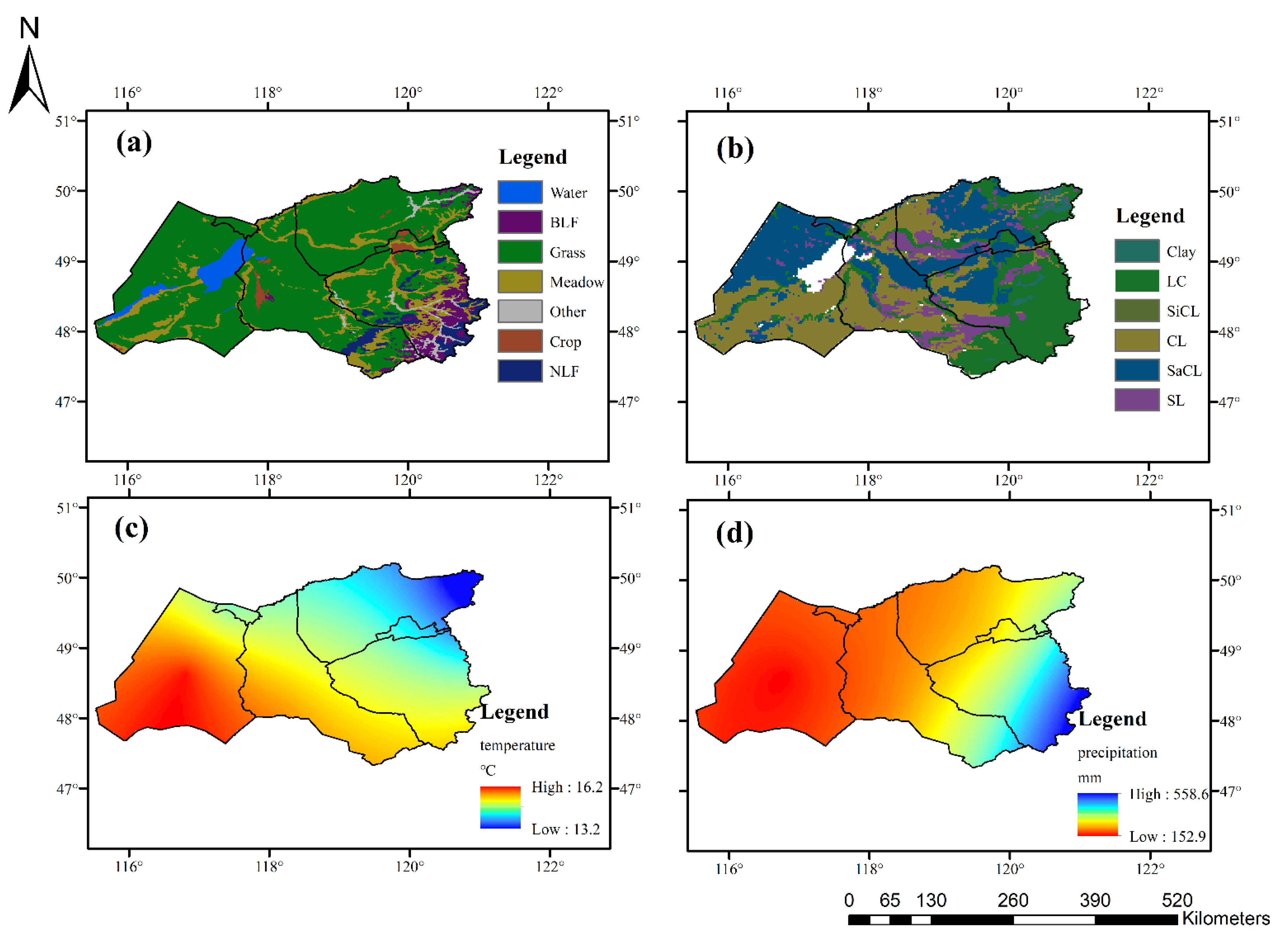
2.1. Study Area
2.2. Data Acquisition
2.3. Data Analysis
2.3.1. NPP Estimation
2.3.2. Time Lag Estimation
- Partial correlation coefficient (PCC) method
- 2.
- Multiple linear regression model method
2.3.3. The Geodetector Model
3. Results
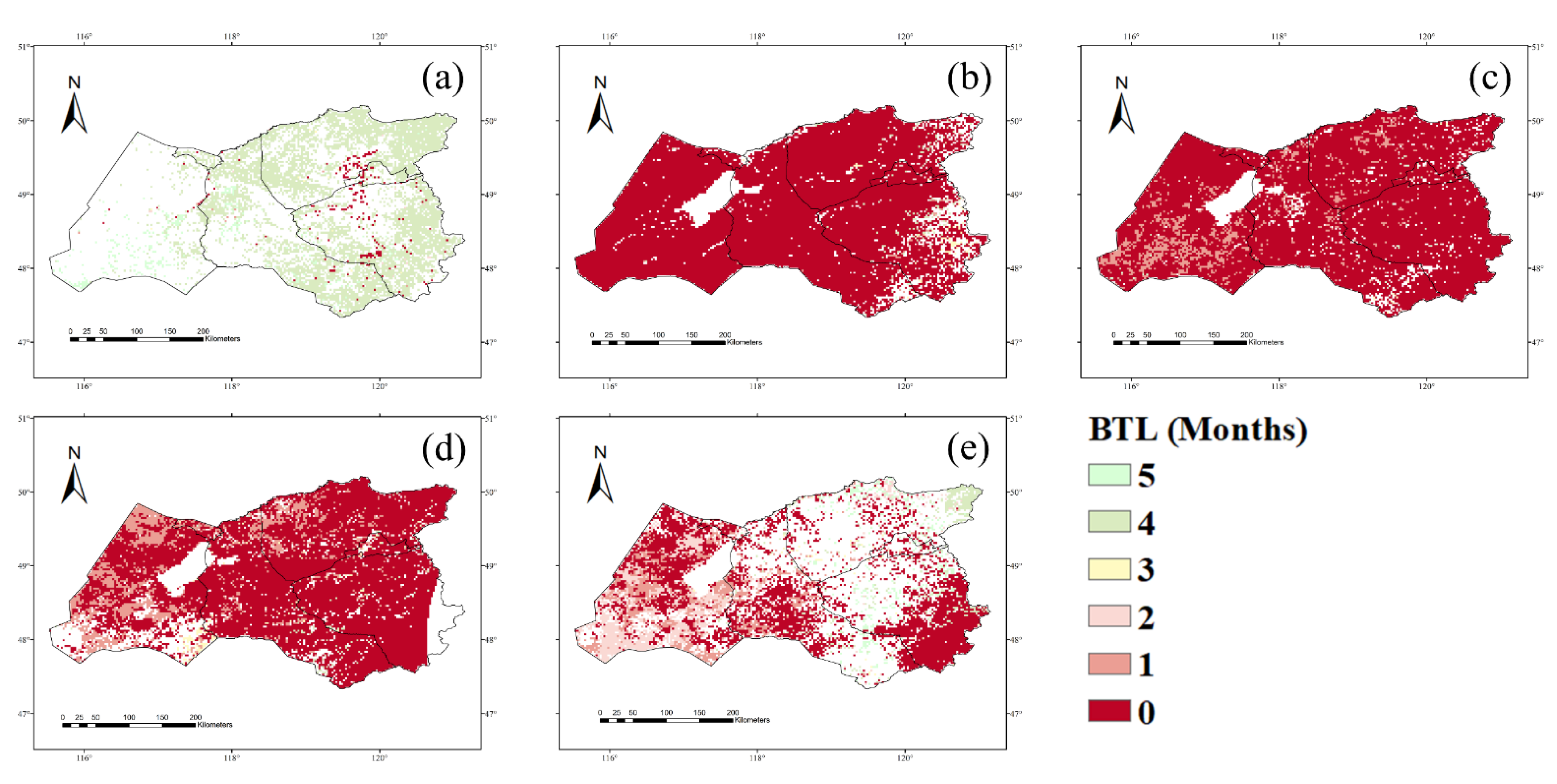
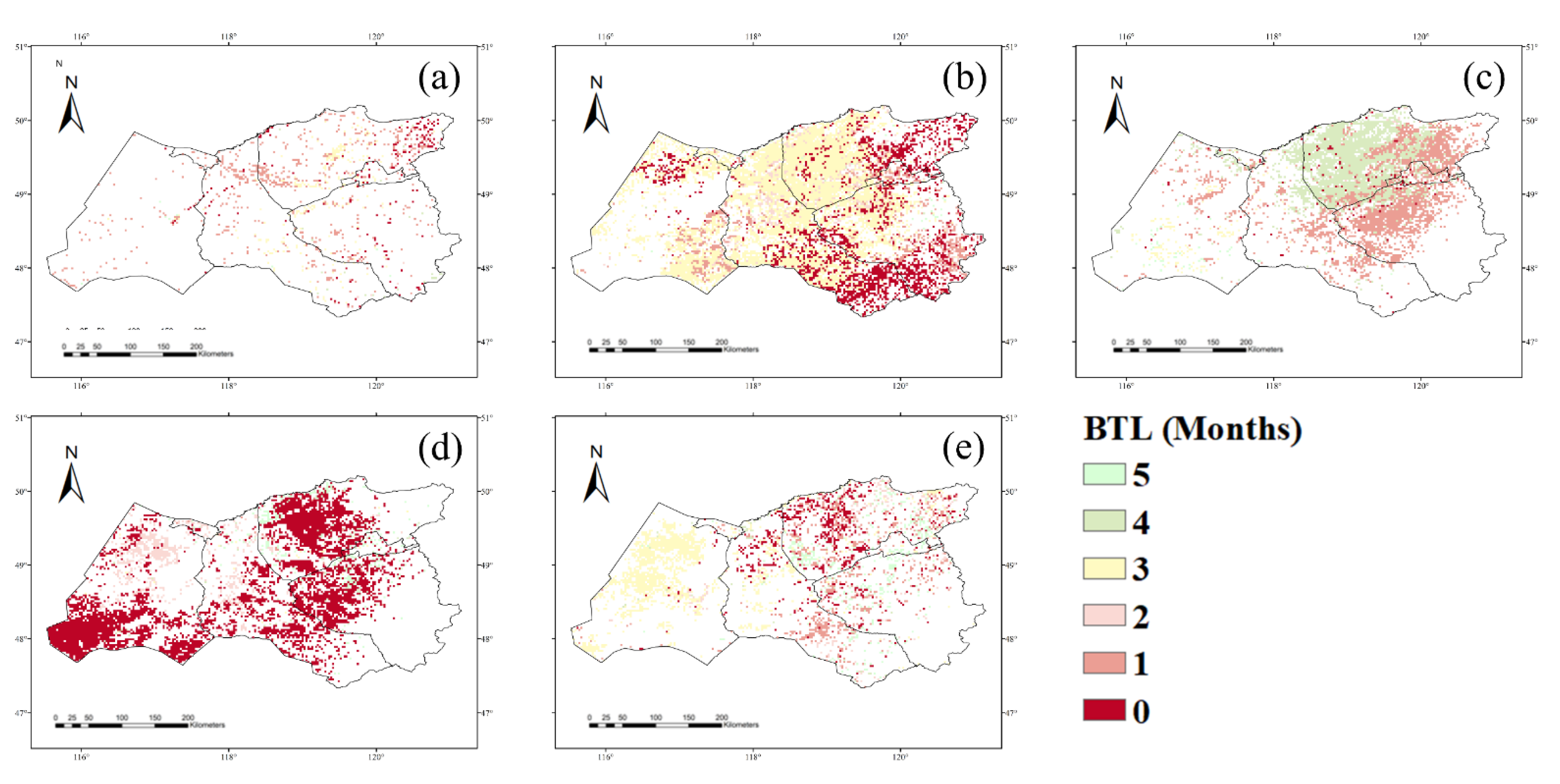
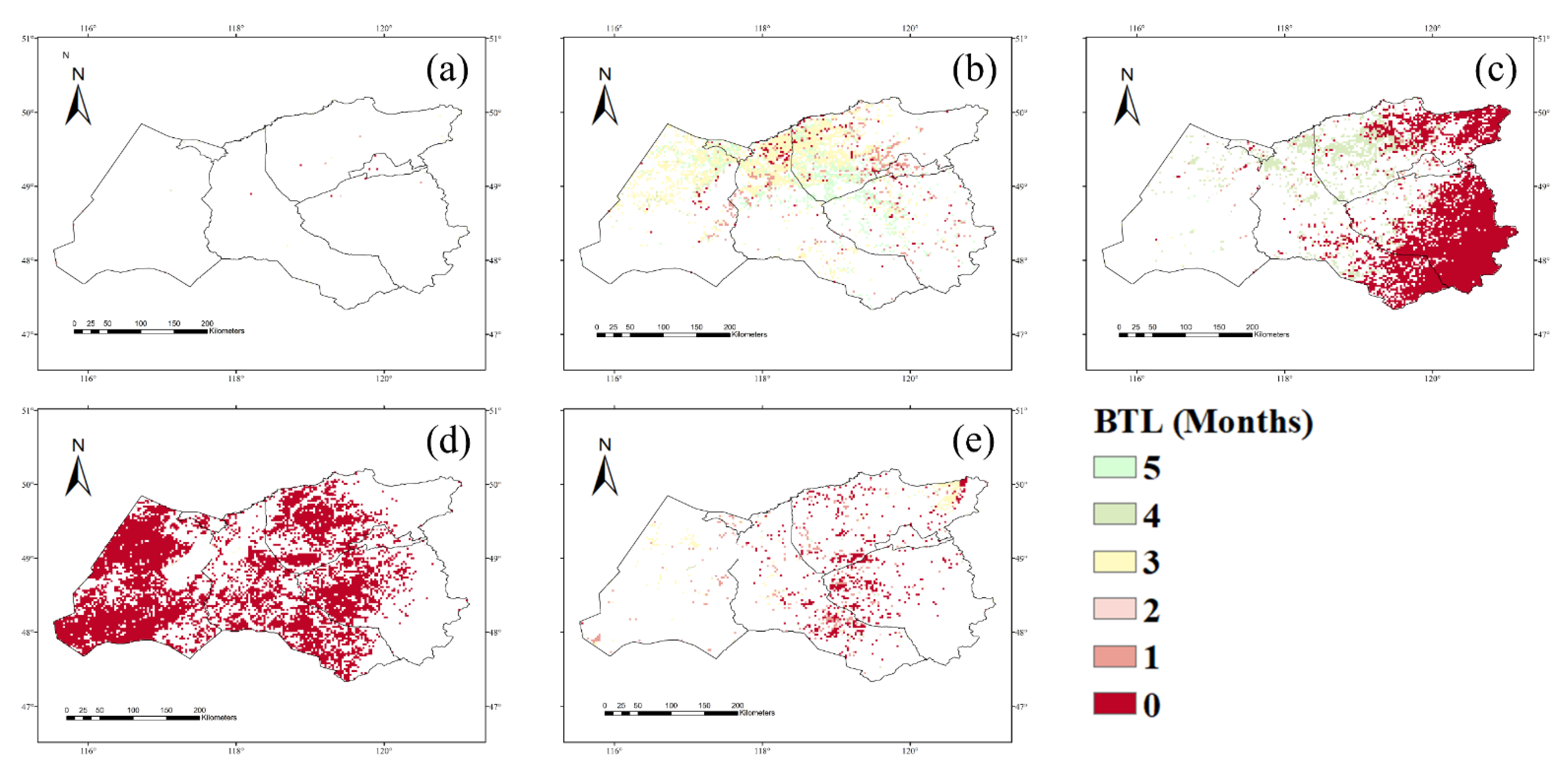
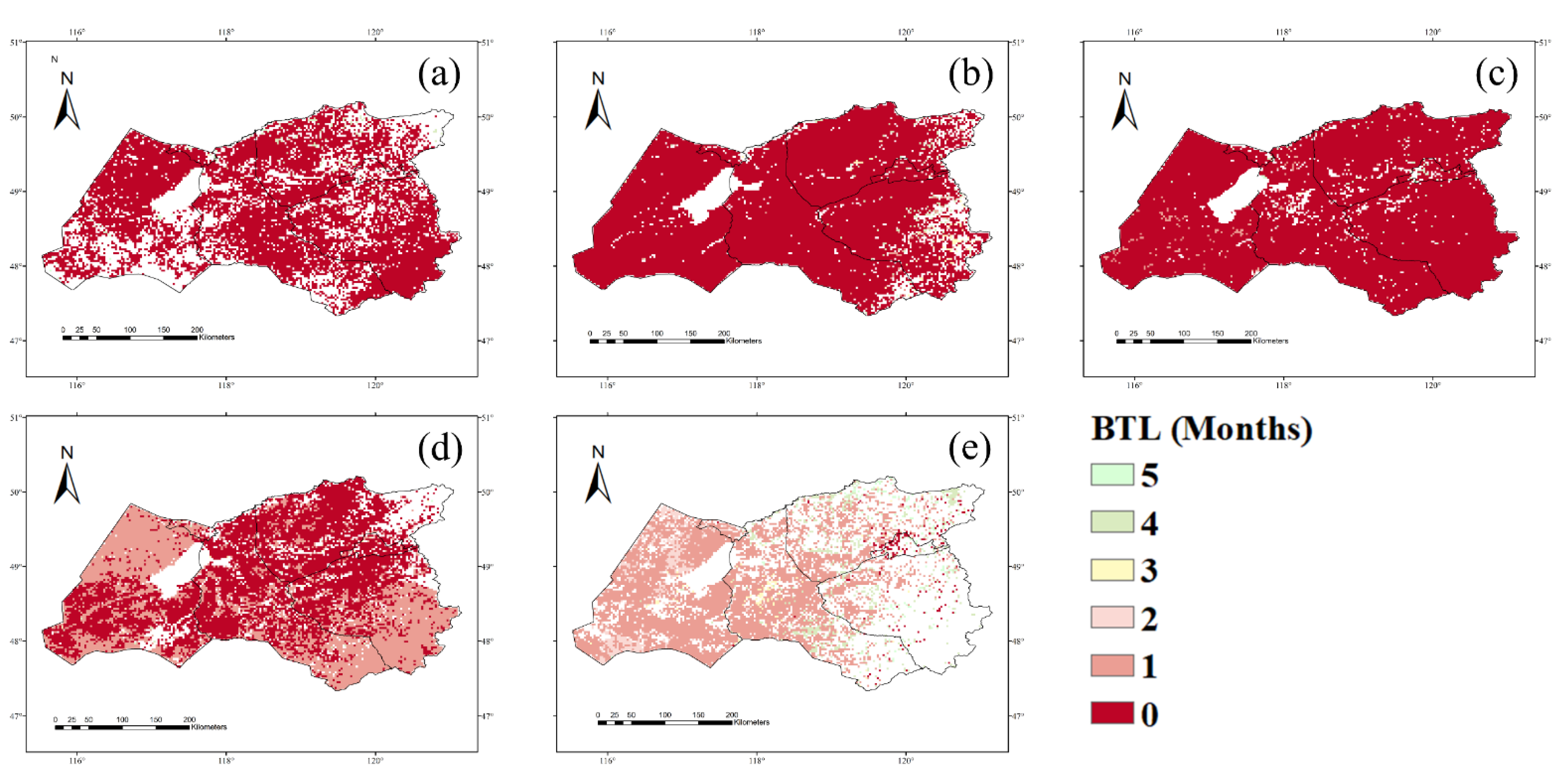
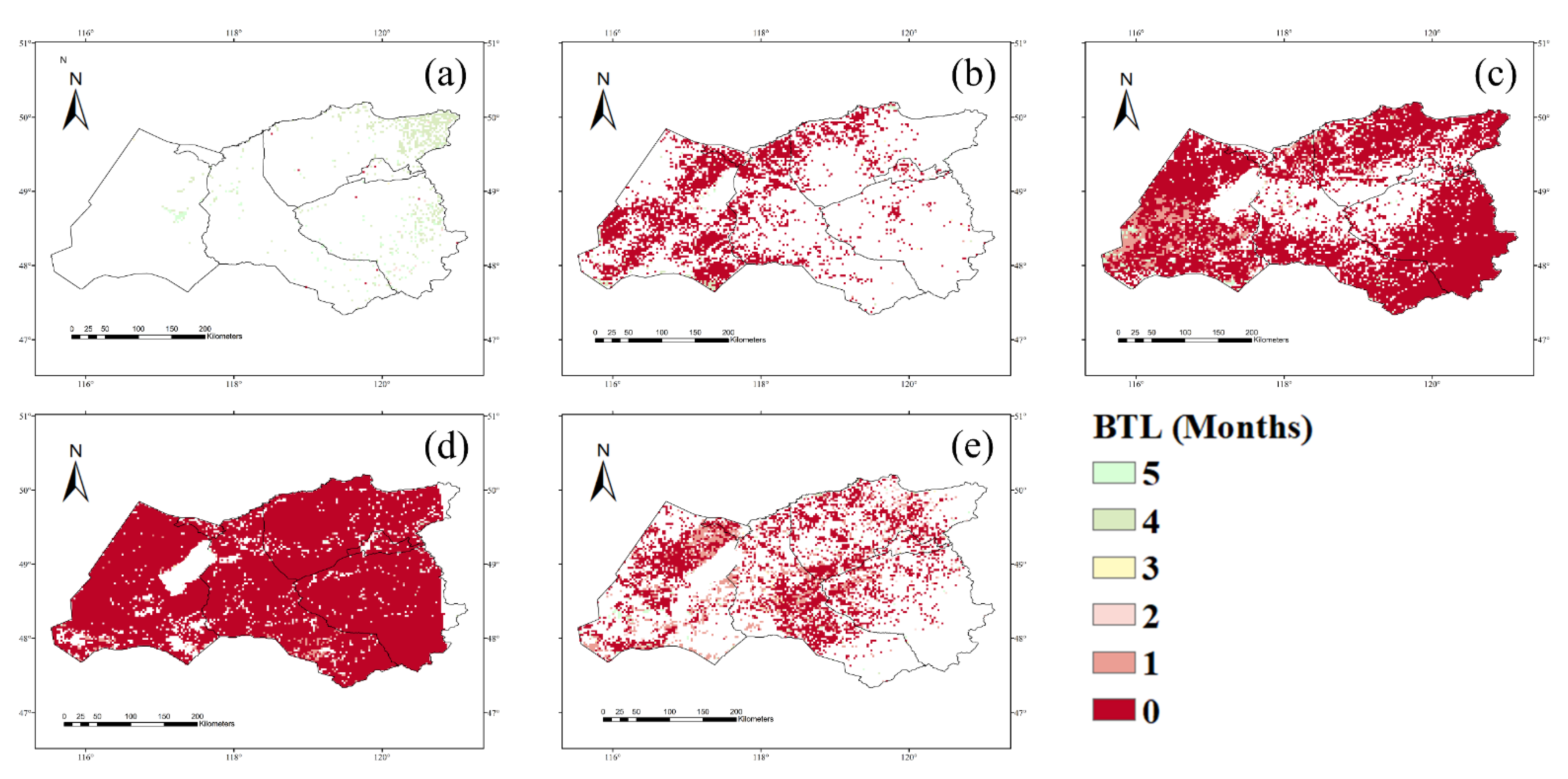
3.1. NPP Response to Climate Conditions with Varied Time Lags
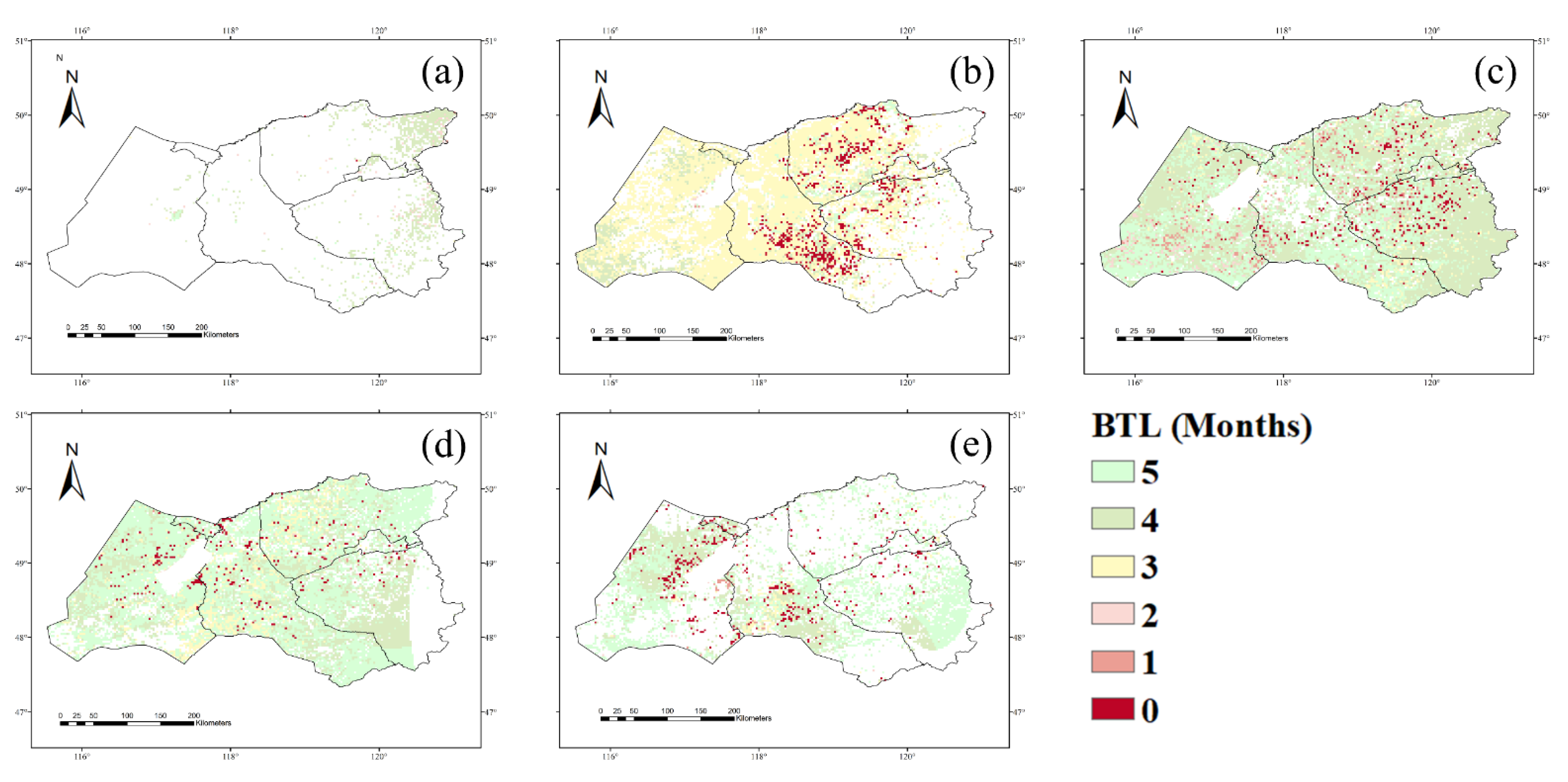
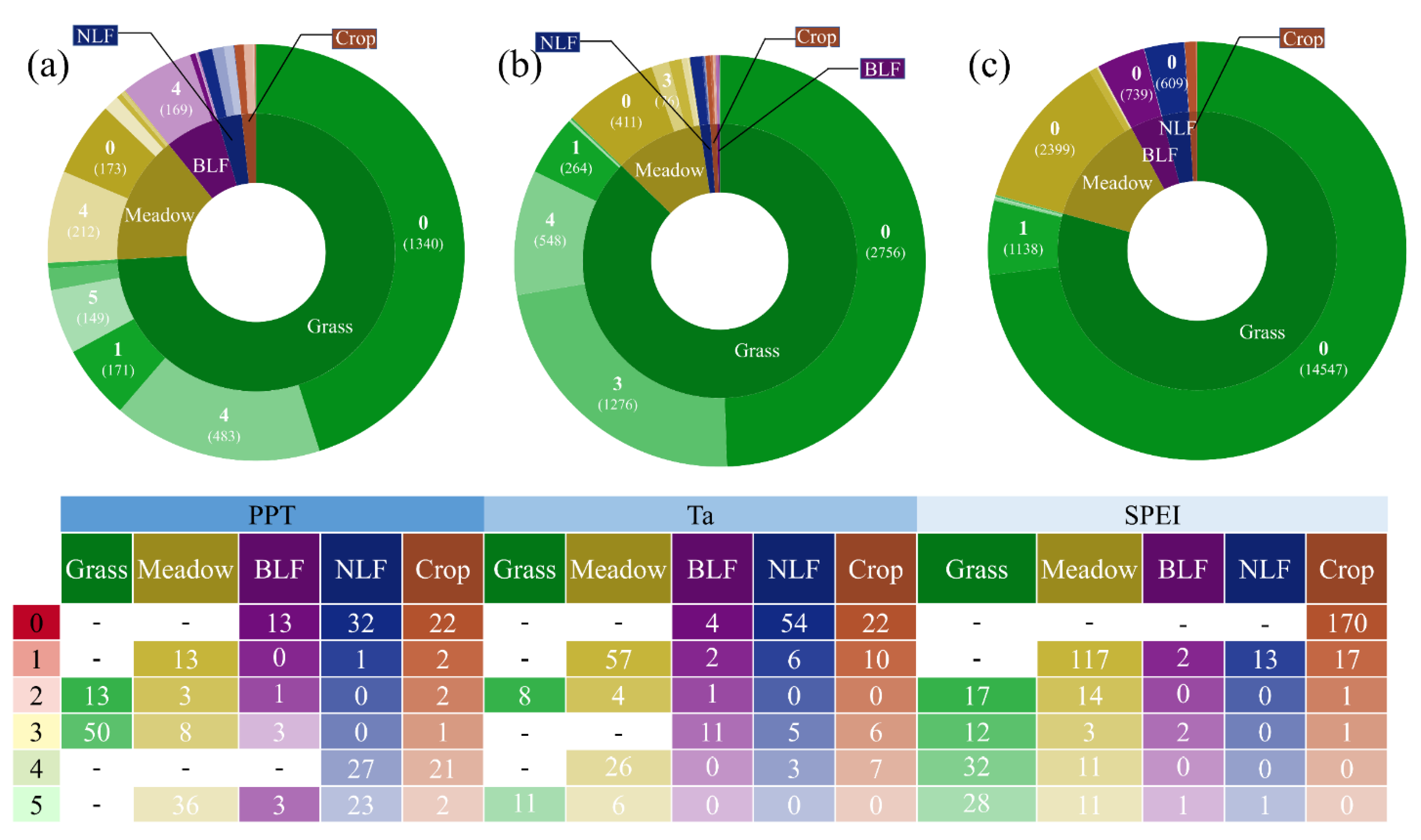
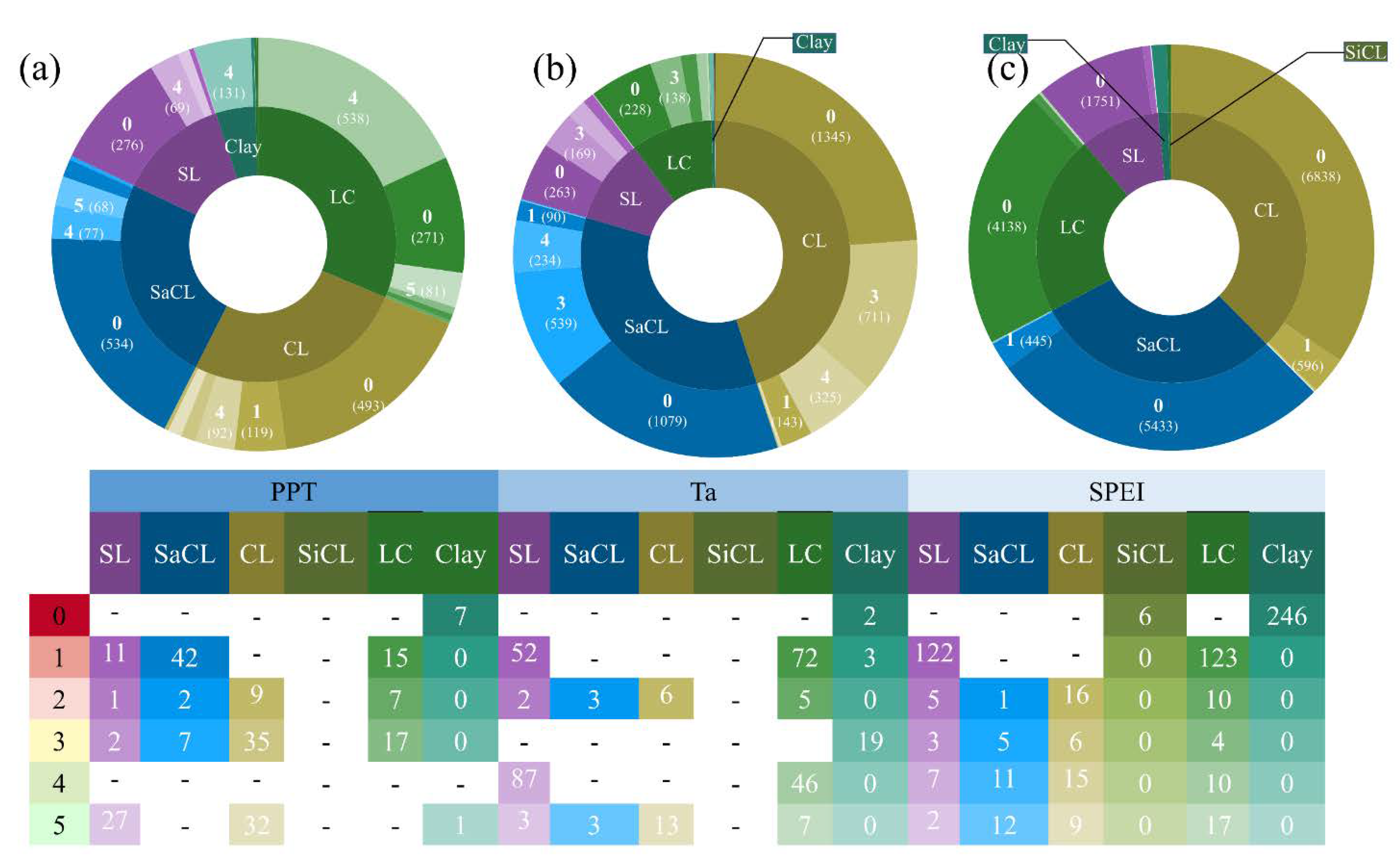
3.2. Time-Lag Effects of Climate Conditions on NPP with Vegetation Type and Soil Texture
4. Discussion
4.1. Time Lags of the NPP Response to Precipitation, Temperature, and Drought
4.2. Time-Lag Effects of the NPP Response to Precipitation, Temperature, and Drought with Different Vegetation Types and Soil Textures
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Particle Size Composition/% | |||
|---|---|---|---|---|
| Clay | Silt | Sand | ||
| sandy soil and sandy loam | SSSL | 0–15 | 0–15 | 85–100 |
| sandy loam | SL | 0–15 | 0–45 | 55–85 |
| loam | Loam | 0–15 | 30–45 | 40–55 |
| silty loam | SiL | 0–15 | 45–100 | 0–55 |
| sandy clay loam | SaCL | 15–25 | 0–30 | 55–85 |
| clay loam | CL | 15–25 | 20–45 | 30–55 |
| silty clay loam | SiCL | 15–25 | 45–85 | 0–40 |
| sandy clay | SaC | 25–45 | 0–20 | 55–75 |
| loamy clay | LC | 25–45 | 0–45 | 10–55 |
| silty clay | SiC | 25–45 | 45–75 | 0–30 |
| clay | Clay | 45–65 | 0–35 | 0–55 |
| heavy clay | HC | 65–100 | 0–35 | 0–35 |
Appendix B
Appendix C
Appendix D
References
- Jia, X.; Zha, T.; Gong, J.; Wang, B.; Zhang, Y.; Wu, B.; Qin, S.; Peltola, H. Carbon and water exchange over a temperate semi-arid shrubland during three years of contrasting precipitation and soil moisture patterns. Agric. For. Meteorol. 2016, 228–229, 120–129. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Seneviratne, S.I.; Zhang, X.; Adnan, M.; Badi, W.; Dereczynski, C.; Di Luca, A.; Ghosh, S.; Iskandar, I.; Kossin, J.; Lewis, S.; et al. Weather and Climate Extreme Events in a Changing Climate. In Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; in press. [Google Scholar]
- Compagnoni, A.; Levin, S.; Childs, D.Z.; Harpole, S.; Paniw, M.; Römer, G.; Burns, J.H.; Che-Castaldo, J.; Rüger, N.; Kunstler, G.; et al. Herbaceous perennial plants with short generation time have stronger responses to climate anomalies than those with longer generation time. Nat. Commun. 2021, 12, 1824. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Wang, Y.; Xu, J.; Yin, J.; Chen, S. Effects of Hydrothermal Conditions on the Net Primary Productivity in the Source Region of Yangtze River, China. Sci. Rep. 2021, 11, 1376. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Y.; Yang, Y.; Zhou, W.; Gang, C.; Zhang, Y.; Li, J.; An, R.; Wang, K.; Odeh, I.; et al. Quantitative assess the driving forces on the grassland degradation in the Qinghai–Tibet Plateau, in China. Ecol. Inform. 2016, 33, 32–44. [Google Scholar] [CrossRef]
- Liu, C.; Dong, X.; Liu, Y. Changes of NPP and their relationship to climate factors based on the transformation of different scales in Gansu, China. Catena 2015, 125, 190–199. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.; Ji, L.; Lei, L.; Wang, C.; Yan, D.; Li, B.; Li, J. Vegetation greenness trend (2000 to 2009) and the climate controls in the Qinghai-Tibetan Plateau. J. Appl. Remote Sens. 2013, 7, 073572. [Google Scholar] [CrossRef]
- Li, B.; Huang, F.; Qin, L.; Qi, H.; Sun, N. Spatio-Temporal Variations of Carbon Use Efficiency in Natural Terrestrial Ecosystems and the Relationship with Climatic Factors in the Songnen Plain, China. Remote Sens. 2019, 11, 2513. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E.; Gherardi, L.A.; Reichmann, L.; Jobbágy, E.; Peters, D. Legacies of precipitation fluctuations on primary production: Theory and data synthesis. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3135–3144. [Google Scholar] [CrossRef] [Green Version]
- Arnone, J.A., III; Verburg, P.S.J.; Johnson, D.W.; Larsen, J.D.; Jasoni, R.L.; Lucchesi, A.J.; Batts, C.M.; Von Nagy, C.; Coulombe, W.G.; Schorran, D.E.; et al. Prolonged suppression of ecosystem carbon dioxide uptake after an anomalously warm year. Nature 2008, 455, 383–386. [Google Scholar] [CrossRef]
- Wu, D.H.; Zhao, X.; Liang, S.L.; Zhou, T.; Huang, K.C.; Tang, B.J.; Zhao, W.Q. Time-lag effects of global vegetation responses to climate change. Glob. Change Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef]
- Li, X.; Liu, M.; Hajek, O.L.; Yin, G. Different Temporal Stability and Responses to Droughts between Needleleaf Forests and Broadleaf Forests in North China during 2001–2018. Forests 2021, 12, 1331. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Li, C.; Cai, Y.; Yang, Z.; Yi, Y. NDVI dynamics under changing meteorological factors in a shallow lake in future metropolitan, semiarid area in North China. Sci. Rep. 2018, 8, 15971. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Kong, D.; Shi, P.; Singh, V.P.; Sun, P. Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Marumbwa, F.M.; Cho, M.A.; Chirwa, P.W. An assessment of remote sensing-based drought index over different land cover types in southern Africa. Int. J. Remote Sens. 2020, 41, 7368–7382. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, X.; Zhou, T.; Wu, D.; Tang, B.; Wei, H. Climatic factors driving vegetation declines in the 2005 and 2010 Amazon droughts. PLoS ONE 2017, 12, e0175379. [Google Scholar] [CrossRef]
- Huang, N.; Wang, L.; Song, X.-P.; Black, T.A.; Jassal, R.S.; Myneni, R.B.; Wu, C.; Song, W.; Ji, D.; Yu, S.; et al. Spatial and temporal variations in global soil respiration and their relationships with climate and land cover. Sci. Adv. 2020, 6, eabb8508. [Google Scholar] [CrossRef]
- Hu, Y.; Yao, Y.; Kou, Z. Exploring on the climate regionalization of Qinling-Daba mountains based on Geodetector-SVM model. PLoS ONE 2020, 15, e0241047. [Google Scholar]
- Manns, H.R.; Berg, A.A.; Colliander, A. Soil organic carbon as a factor in passive microwave retrievals of soil water content over agricultural croplands. J. Hydrol. 2015, 528, 643–651. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Pan, Y.; Ji, C. Aboveground biomass in Tibetan grasslands. J. Arid Environ. 2009, 73, 91–95. [Google Scholar] [CrossRef]
- Ohanty, B.P.; Famiglietti, J.S.; Skaggs, T.H. Evolution of soil moisture spatial structure in a mixed vegetation pixel during the Southern Great Plains 1997 (SGP97) Hydrology Experiment. Water Resour. Res. 2000, 36, 3675–3686. [Google Scholar] [CrossRef] [Green Version]
- Evans, P.; Brown, C.D. The boreal–temperate forest ecotone response to climate change. Environ. Rev. 2017, 25, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Breshears, D.D. Drought-induced shift of a forest–woodland ecotone: Rapid landscape response to climate variation. Proc. Natl. Acad. Sci. USA 1998, 95, 14839–14842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Wu, J.; Xing, Q.; Pan, Q.; Huang, J.; Yang, D.; Han, X. Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau. Ecology 2008, 89, 2140–2153. [Google Scholar] [CrossRef]
- Dai, E.; Huang, Y.; Wu, Z.; Zhao, D. Analysis of spatio-temporal features of a carbon source/sink and its relationship to climatic factors in the Inner Mongolia grassland ecosystem. J. Geogr. Sci. 2016, 26, 297–312. [Google Scholar] [CrossRef] [Green Version]
- Loydi, A.; Lohse, K.; Otte, A.; Donath, T.W.; Eckstein, R.L. Distribution and effects of tree leaf litter on vegetation composition and biomass in a forest–grassland ecotone. J. Plant Ecol. 2013, 7, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Loehle, C. Forest ecotone response to climate change: Sensitivity to temperature response functional forms. Can. J. For. Res. 2000, 30, 1632–1645. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J. Grassland Productivity Response to Climate Change in the Hulunbuir Steppes of China. Sustainability 2019, 11, 6760. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Tian, H.; Dangal, S.R.S.; Ouyang, Z.; Lu, C.; Yang, J.; Tao, B.; Ren, W.; Banger, K.; Yang, Q.; et al. Impacts of climate variability and extremes on global net primary production in the first decade of the 21st century. J. Geogr. Sci. 2015, 25, 1027–1044. [Google Scholar] [CrossRef]
- Chuai, X.W.; Huang, X.J.; Wang, W.J.; Bao, G. NDVI, temperature and precipitation changes and their relationships with different vegetation types during 1998-2007 in Inner Mongolia, China. Int. J. Clim. 2012, 33, 1696–1706. [Google Scholar] [CrossRef]
- Na, R.; Du, H.; Na, L.; Shan, Y.; He, H.S.; Wu, Z.; Zong, S.; Yang, Y.; Huang, L. Spatiotemporal changes in the Aeolian desertification of Hulunbuir Grassland and its driving factors in China during 1980–2015. Catena 2019, 182, 104123. [Google Scholar] [CrossRef]
- Peng, F.; Fan, W.; Xu, X.; Liu, X. Analysis on temporal-spatial change of vegetation coverage in Hulunbuir Steppe (2000–2014). In Proceedings of the 2016 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Beijing, China, 10–15 July 2016. [Google Scholar] [CrossRef]
- Yang, X.; Liang, P.; Zhang, D.; Li, H.; Rioual, P.; Wang, X.; Xu, B.; Ma, Z.; Liu, Q.; Ren, X.; et al. Holocene aeolian stratigraphic sequences in the eastern portion of the desert belt (sand seas and sandy lands) in northern China and their palaeoenvironmental implications. Sci. China Earth Sci. 2019, 62, 1302–1315. [Google Scholar] [CrossRef]
- Liu, M.; Liu, G.; Gong, L.; Wang, D.; Sun, J. Relationships of Biomass with Environmental Factors in the Grassland Area of Hulunbuir, China. PLoS ONE 2014, 9, e102344. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Shi, Z.; Xu, L.; Yang, X.; Jia, Z.; Lü, S.; Feng, C.; Shang, J. Precipitation variability in Hulunbuir, northeastern China since 1829 AD reconstructed from tree-rings and its linkage with remote oceans. J. Arid Environ. 2013, 95, 14–21. [Google Scholar] [CrossRef]
- Zhang, G.; Xu, X.; Zhou, C.; Zhang, H.; Ouyang, H. Responses of grassland vegetation to climatic variations on different temporal scales in Hulun Buir Grassland in the past 30 years. J. Geogr. Sci. 2011, 21, 634–650. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Water Requirements, FAO Irrigation and Drainage Paper 56; Food agriculture of the United Nations: Remo, Italy, 1998; ISBN 92-5-104219-5. [Google Scholar]
- Piao, S.L.; Fang, J.Y.; Guo, Q.H. Application of CASA model to the estimation of Chinese terrestrial net primary productivity. Acta Phytoecol. Sin. 2001, 25, 603–608. [Google Scholar]
- Zhang, S.; Zhang, R.; Liu, T.; Song, X.; Adams, M.A. Empirical and model-based estimates of spatial and temporal variations in net primary productivity in semi-arid grasslands of Northern China. PLoS ONE 2017, 12, e0187678. [Google Scholar] [CrossRef] [Green Version]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Scurlock, J.M.O.; Johnson, K.; Olson, R.J. Estimating net primary productivity from grassland biomass dynamics measurements. Glob. Change Biol. 2002, 8, 736–753. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xue, Y. Expansion of the Sahara Desert and shrinking of frozen land of the Arctic. Sci. Rep. 2020, 10, 4109. [Google Scholar] [CrossRef]
- Potter, C.S.; Randerson, J.T.; Field, C.B.; Matson, P.A.; Vitousek, P.M.; Mooney, H.A.; Klooster, S.A. Terrestrial ecosystem production: A process model based on global satellite and surface data. Glob. Biogeochem. Cycles 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhao, J.; Zhu, Z.; Zhang, H.; Zhang, Z.; Guo, X.; Bi, Y.; Sun, L. Remotely Sensed Estimation of Net Primary Productivity (NPP) and Its Spatial and Temporal Variations in the Greater Khingan Mountain Region, China. Sustainability 2017, 9, 1213. [Google Scholar] [CrossRef] [Green Version]
- Field, C.B.; Randerson, J.T.; Malmström, C.M. Global net primary production: Combining ecology and remote sensing. Remote Sens. Environ. 1995, 51, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Liu, X.; Du, G. Nonuniform Time-Lag Effects of Asymmetric Warming on Net Primary Productivity across Global Terrestrial Biomes. Earth Interact. 2018, 22, 1–26. [Google Scholar] [CrossRef]
- Zhou, G.S.; Zhou, X.S. A natural vegetation NPP model. Acta Phytoecol. Sin. 1995, 19, 193–200. [Google Scholar]
- Zhu, W.Q.; Pan, Y.Z.; Zhang, J.S. Estimation of net primary productivity of Chinese terrestrial vegetation based on remote sensing. Chin. J. Plant Ecol. 2007, 31, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Liu, H.; Yin, Y.; He, S. Drivers of greening trend across vertically distributed biomes in temperate arid Asia. Geophys. Res. Lett. 2007, 34, L07707. [Google Scholar] [CrossRef]
- Xu, G.; Zhang, H.; Chen, B.; Zhang, H.; Innes, J.L.; Wang, G.; Yan, J.; Zheng, Y.; Zhu, Z.; Myneni, R.B. Changes in Vegetation Growth Dynamics and Relations with Climate over China’s Landmass from 1982 to 2011. Remote Sens. 2014, 6, 3263–3283. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, A.; Liu, C.; Zhao, Y.; Zhao, A.; Wang, D. Analysis of the time-lag effects of climate factors on grassland productivity in Inner Mongolia. Glob. Ecol. Conserv. 2021, 30, e01751. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Chen, X. Quantifying Influences of Natural and Anthropogenic Factors on Vegetation Changes Based on Geodetector: A Case Study in the Poyang Lake Basin, China. Remote Sens. 2021, 13, 5081. [Google Scholar] [CrossRef]
- Song, Y.; Wang, J.; Ge, Y.; Xu, C. An optimal parameters-based geographical detector model enhances geographic characteristics of explanatory variables for spatial heterogeneity analysis: Cases with different types of spatial data. GISci. Remote Sens. 2020, 57, 593–610. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Guo, Q.; Henderson, M.; Ji, W.; Li, Y.; Tao, S. Interannual variations of monthly and seasonal normalized difference vegetation index (NDVI) in China from 1982 to 1999. J. Geophys. Res. Earth Surf. 2003, 108, 1–13. [Google Scholar] [CrossRef]
- Zhao, A.; Yu, Q.; Feng, L.; Zhang, A.; Pei, T. Evaluating the cumulative and time-lag effects of drought on grassland vegetation: A case study in the Chinese Loess Plateau. J. Environ. Manag. 2020, 261, 110214. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, Z.; Peng, S. Global analysis of time-lag and -accumulation effects of climate on vegetation growth. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102179. [Google Scholar] [CrossRef]
- Kong, D.; Miao, C.; Wu, J.; Zheng, H.; Wu, S. Time lag of vegetation growth on the Loess Plateau in response to climate factors: Estimation, distribution, and influence. Sci. Total Environ. 2020, 744, 140726. [Google Scholar] [CrossRef]
- Li, Z.; Guo, X. Detecting Climate Effects on Vegetation in Northern Mixed Prairie Using NOAA AVHRR 1-km Time-Series NDVI Data. Remote Sens. 2012, 4, 120–134. [Google Scholar] [CrossRef] [Green Version]
- Sherry, R.A.; Weng, E.; Iii, J.A.A.; Johnson, D.W.; Schimel, D.S.; Verburg, P.S.; Wallace, L.L.; Luo, Y. Lagged effects of experimental warming and doubled precipitation on annual and seasonal aboveground biomass production in a tallgrass prairie. Glob. Change Biol. 2008, 14, 2923–2936. [Google Scholar] [CrossRef]
- Robertson, T.R.; Bell, C.W.; Zak, J.C.; Tissue, D.T. Precipitation timing and magnitude differentially affect aboveground annual net primary productivity in three perennial species in a Chihuahuan Desert grassland. New Phytol. 2008, 181, 230–242. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Fu, J. Vegetation response to precipitation anomalies under different climatic and biogeographical conditions in China. Sci. Rep. 2020, 10, 830. [Google Scholar] [CrossRef] [Green Version]
- Doi, R.D. Vegetational response of rainfall in Rajasthan using AVHRR imagery. J. Indian Soc. Remote Sens. 2001, 29, 213–224. [Google Scholar] [CrossRef]
- Au, T.F.; Maxwell, J.T. Drought Sensitivity and Resilience of Oak–Hickory Stands in the Eastern United States. Forests 2022, 13, 389. [Google Scholar] [CrossRef]
- Camberlin, P.; Martiny, N.; Philippon, N.; Richard, Y. Determinants of the interannual relationships between remote sensed photosynthetic activity and rainfall in tropical Africa. Remote Sens. Environ. 2007, 106, 199–216. [Google Scholar] [CrossRef] [Green Version]
- Ni, J. Estimating net primary productivity of grasslands from field biomass measurements in temperate northern China. Plant Ecol. 2004, 174, 217–234. [Google Scholar] [CrossRef]
- Jobbagy, E.G.; Sala, O.E. Controls of Grass and Shrub Aboveground Production in the Patagonian Steppe. Ecol. Appl. 2000, 10, 541–549. [Google Scholar] [CrossRef]
- Villegas, J.C.; Breshears, D.D.; Zou, C.; Royer, P.D. Seasonally Pulsed Heterogeneity in Microclimate: Phenology and Cover Effects along Deciduous Grassland–Forest Continuum. Vadose Zone J. 2010, 9, 537–547. [Google Scholar] [CrossRef]
- Sui, X.; Zhou, G.; Zhuang, Q. Sensitivity of carbon budget to historical climate variability and atmospheric CO2 concentration in temperate grassland ecosystems in China. Clim. Change 2012, 117, 259–272. [Google Scholar] [CrossRef]
- Peaucelle, M.; Janssens, I.A.; Stocker, B.D.; Ferrando, A.D.; Fu, Y.H.; Molowny-Horas, R.; Ciais, P.; Peñuelas, J. Spatial variance of spring phenology in temperate deciduous forests is constrained by background climatic conditions. Nat. Commun. 2019, 10, 5388. [Google Scholar] [CrossRef] [Green Version]










| Interaction Relationship | Interaction Types |
|---|---|
| q(X1∩X2) < Min [q(X1), q(X2)] | Nonlinear weakened |
| Min [q(X1), q(X2)] < q(X1∩X2) < Max [q(X1), q(X2)] | Univariable weakened |
| q(X1∩X2) = q(X1) + q(X2) | Independent |
| Max(q(X1), q(X2)) < q(X1∩X2) < q(X1) + q(X2) | Bivariable enhanced |
| q(X1∩X2) > q(X1) + q(X2) | Nonlinear enhanced |
| Vegetation Types | Soil Textures | Vegetation Types ∩ Soil Textures | |
|---|---|---|---|
| PPT | 0.15 ** | 0.31 ** | 0.37 |
| Ta | 0.10 ** | 0.04 ** | 0.13 |
| SPEI | 0.00 | 0.00 | 0.00 |
| Climate Conditions | Time Lag | Timescale | Study Area | Time Span | Reference |
|---|---|---|---|---|---|
| precipitation | 4-month lag | monthly scale | Hulunbuir | 2000–2018 | in this study |
| 0.55 ± 0.95-month lag | monthly scale | Global | 1982–2015 | [57] | |
| 7.9~17.7-day lag | daily | The Chinese Loess Plateau | 1982–2015 | [58] | |
| 40-day lag | daily | Grasslands National Park, southern Saskatchewan, Canada | 1985–2007 | [59] | |
| temperature | 0~4-month lag | monthly scale | Hulunbuir | 2000–2018 | in this study |
| 0.56 ± 1.04-month lag | monthly scale | Global | 1982–2015 | [57] | |
| 3-month lag | seasonal scale | China | 1982–1999 | [55] | |
| 6.2~25.3-day lag | daily | The Chinese Loess Plateau | 1982–2015 | [58] | |
| 10-day lag | daily | Grasslands National Park, southern Saskatchewan, Canada | 1985–2007 | [59] | |
| drought | 0~2-month lag | monthly scale | Hulunbuir | 2000–2018 | in this study |
| 2~3-month lag | seasonal scale | The Chinese Loess Plateau | 2000–2010 | [56] | |
| 2-month lag | seasonal scale | Southern Africa | 2015–2016 | [16] | |
| 8-month lag | seasonal scale | Southern Africa | 2015–2016 | [16] | |
| solar radiation | 0.50 ± 0.94-month lag | monthly scale | Global | 1982–2015 | [57] |
| soil water availability | 2–9-month lag | Kessler Farm Field Laboratory, Central Redbed Plains of Oklahoma | From 20 February 2003 to 20 February 2004 | [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Tian, Y.; Liu, S.; Jiang, L.; Mao, J.; Jia, X.; Zha, T.; Zhang, K.; Wu, Y.; Zhou, J. Time-Lag Effect of Climate Conditions on Vegetation Productivity in a Temperate Forest–Grassland Ecotone. Forests 2022, 13, 1024. https://doi.org/10.3390/f13071024
Liu X, Tian Y, Liu S, Jiang L, Mao J, Jia X, Zha T, Zhang K, Wu Y, Zhou J. Time-Lag Effect of Climate Conditions on Vegetation Productivity in a Temperate Forest–Grassland Ecotone. Forests. 2022; 13(7):1024. https://doi.org/10.3390/f13071024
Chicago/Turabian StyleLiu, Xinyue, Yun Tian, Shuqin Liu, Lixia Jiang, Jun Mao, Xin Jia, Tianshan Zha, Kebin Zhang, Yuqing Wu, and Jianqin Zhou. 2022. "Time-Lag Effect of Climate Conditions on Vegetation Productivity in a Temperate Forest–Grassland Ecotone" Forests 13, no. 7: 1024. https://doi.org/10.3390/f13071024