
Changes in Species and Functional Diversity of the Herb Layer of Riparian Forest despite Six Decades of Strict Protection

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
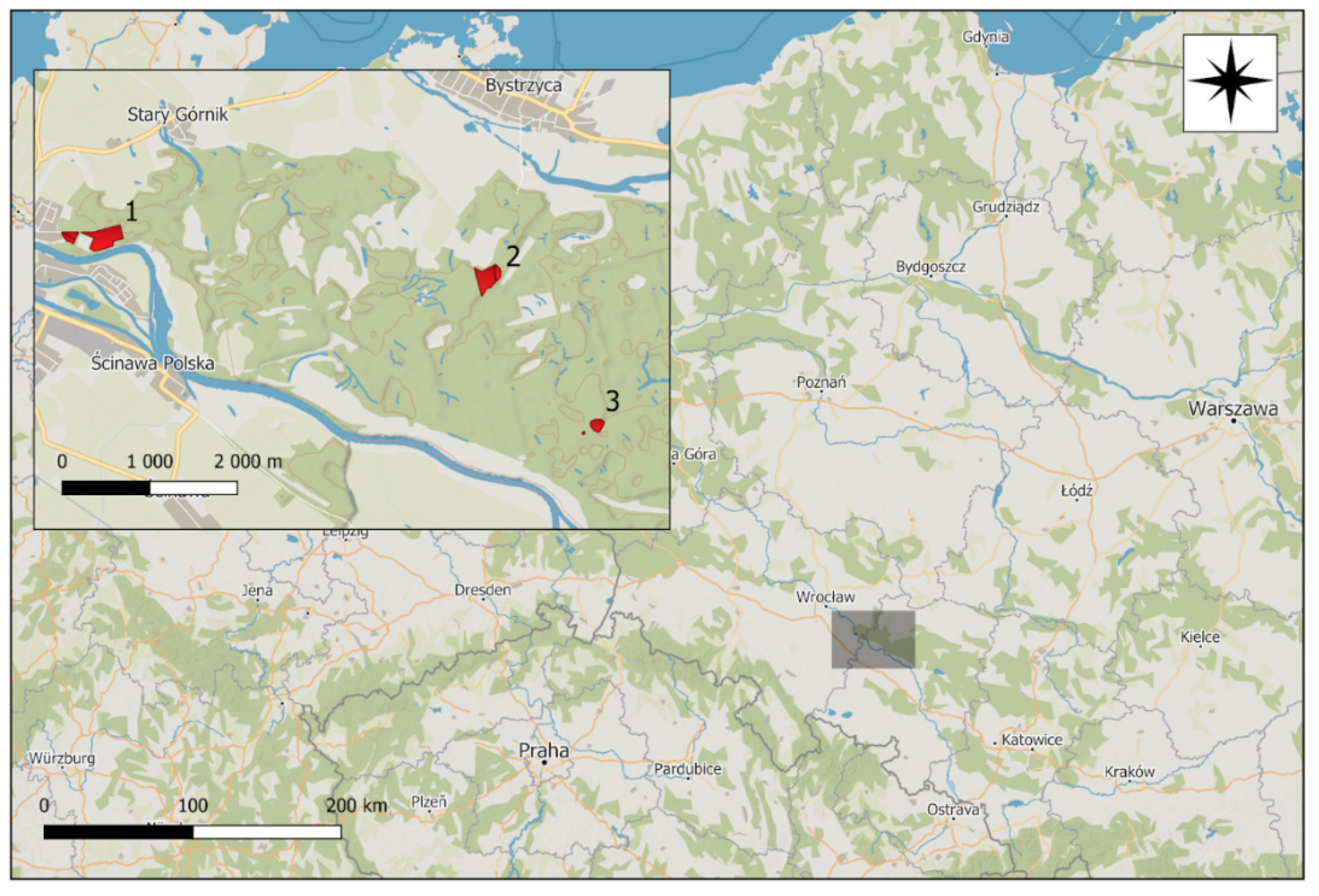
2.1. Study Area
2.2. Sampling
2.3. Data Analysis
2.3.1. The Overall Pattern in Vegetation
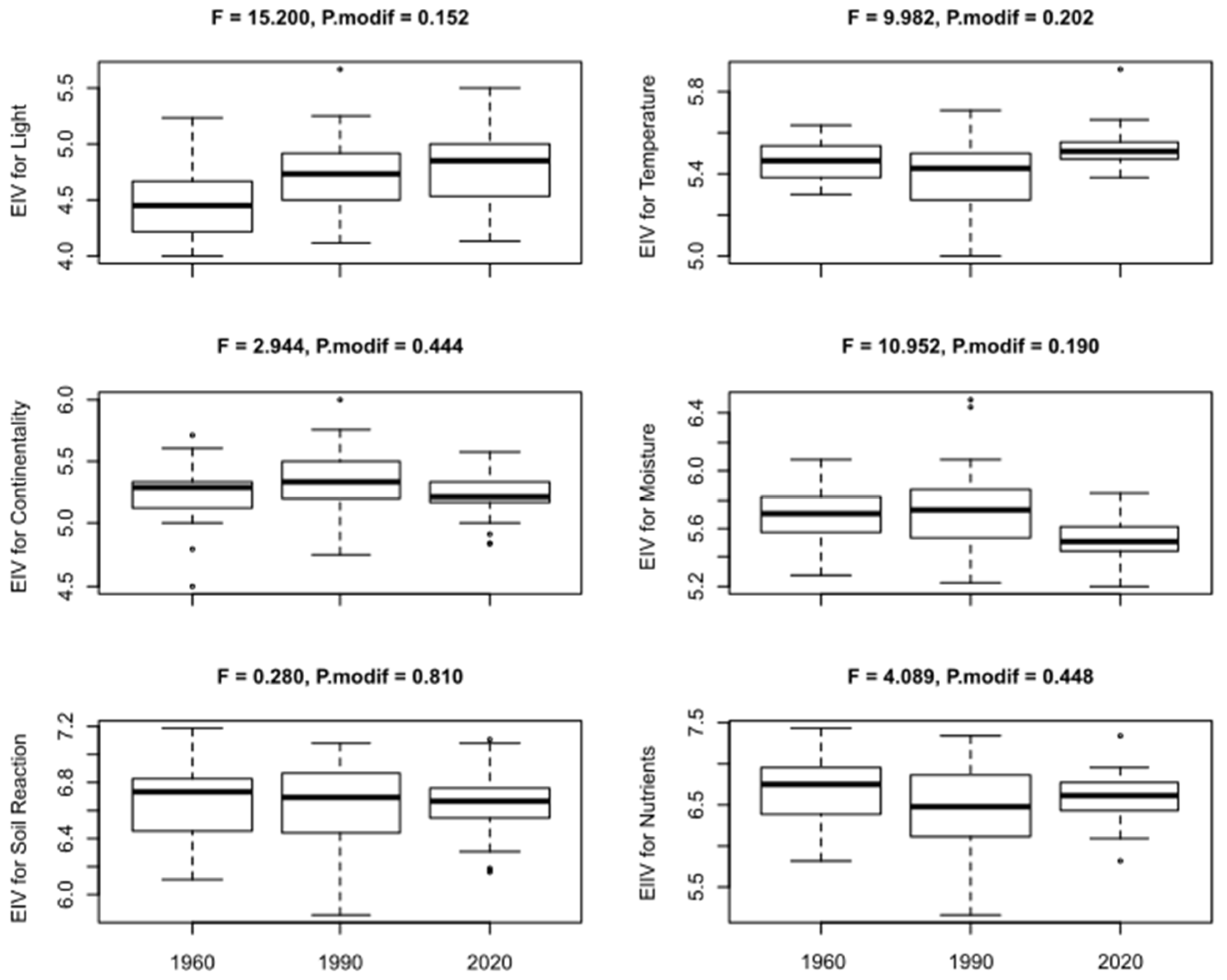
2.3.2. Habitat Conditions
2.3.3. Plant Functional Traits
3. Results
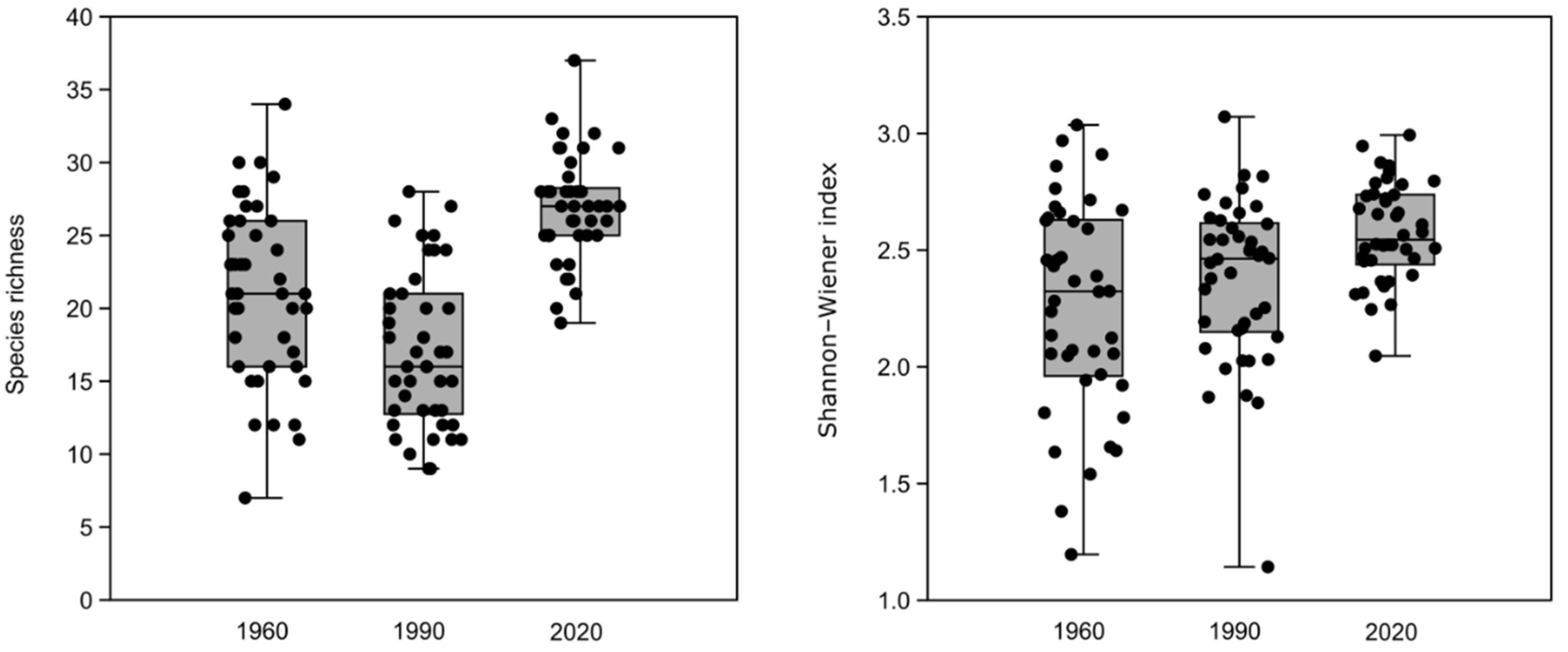
3.1. Changes in Species Diversity
3.2. Changes in Species Composition
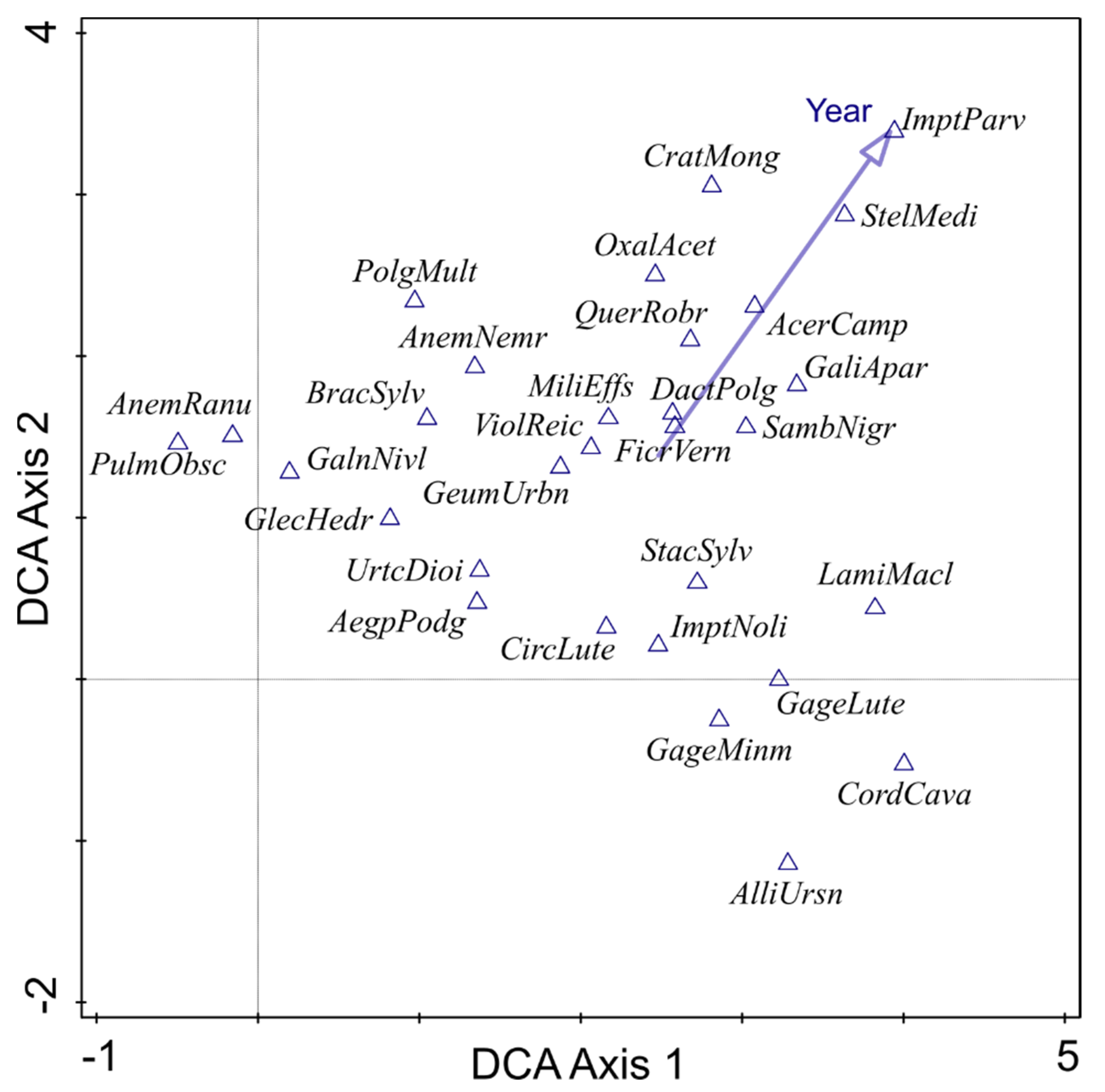
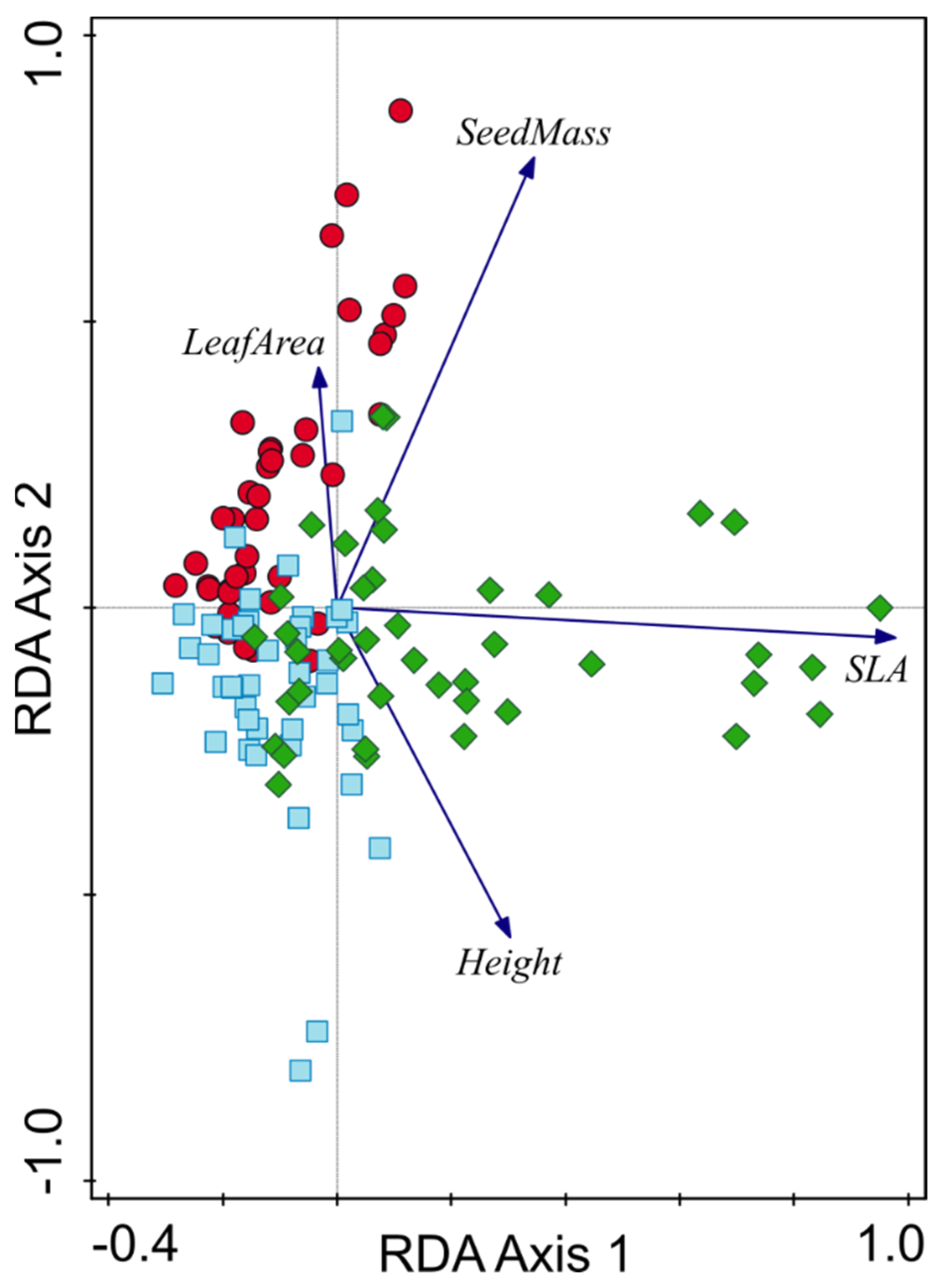
3.3. Temporal Changes in Herbaceous Vegetation Pattern
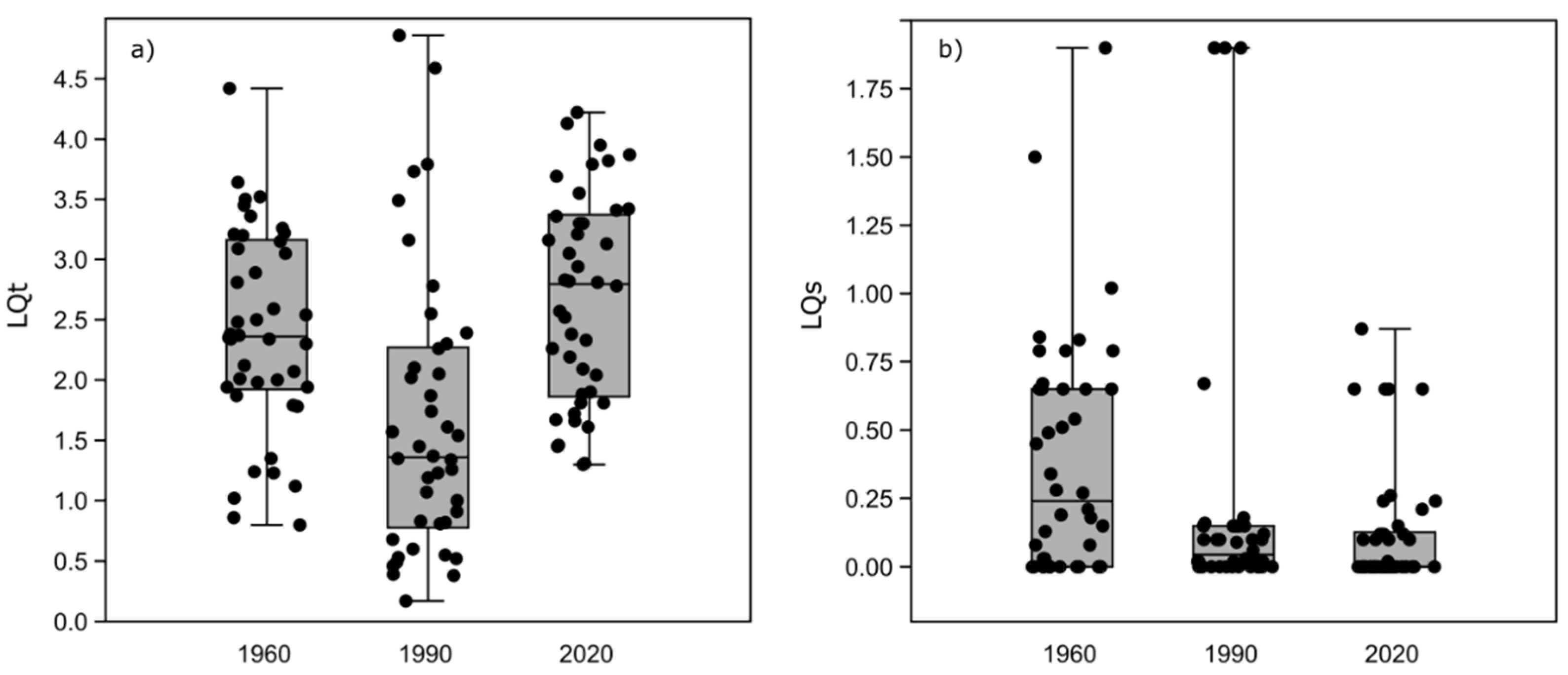
3.4. Changes in Litter Quality Index
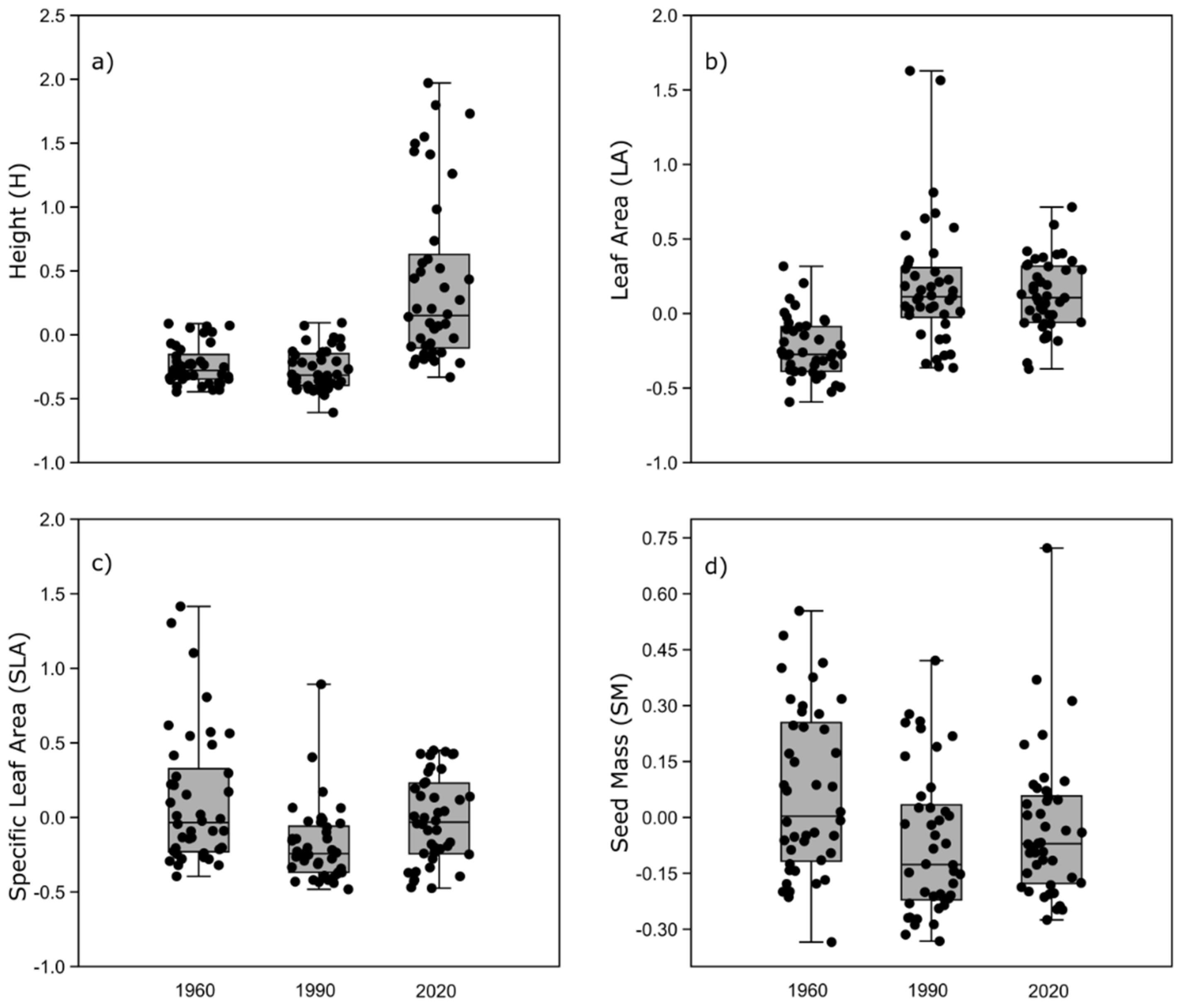
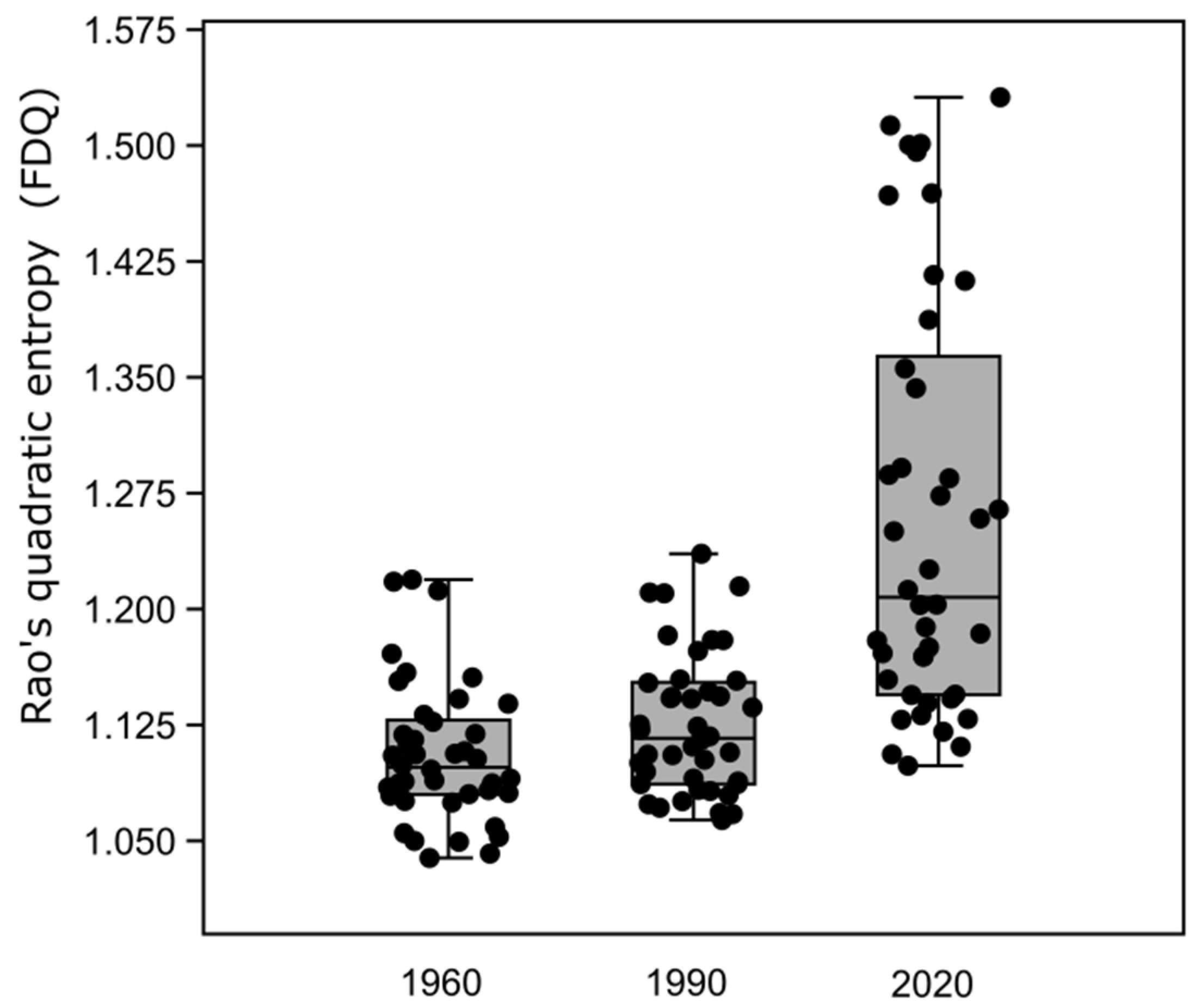
3.5. Changes in Functional Structure
4. Discussion
4.1. Species Composition and Diversity Change
4.2. Possible Reasons for the Change in Species Composition
4.3. Changes in Vegetation Type and Habitat Conditions
4.4. Functional Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Plant Functional Types and Ecosystem Function in Relation to Global Change. J. Veg. Sci. 1997, 8, 463–474. [Google Scholar] [CrossRef]
- Trumbore, S.; Brando, P.; Hartmann, H. Forest Health and Global Change. Science 2015, 349, 814–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanneste, T.; Valdés, A.; Verheyen, K.; Perring, M.P.; Bernhardt-Römermann, M.; Andrieu, E.; Brunet, J.; Cousins, S.A.O.; Deconchat, M.; De Smedt, P.; et al. Functional Trait Variation of Forest Understorey Plant Communities across Europe. Basic Appl. Ecol. 2019, 34, 1–14. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate Change Impacts, Adaptive Capacity, and Vulnerability of European Forest Ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Hansen, J.; Sato, M.; Ruedy, R.; Lo, K.; Lea, D.W.; Medina-Elizade, M. Global Temperature Change. Proc. Natl. Acad. Sci. USA 2006, 103, 14288–14293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Zohner, C.M.; Mo, L.; Renner, S.S.; Svenning, J.-C.; Vitasse, Y.; Benito, B.M.; Ordonez, A.; Baumgarten, F.; Bastin, J.-F.; Sebald, V.; et al. Late-Spring Frost Risk between 1959 and 2017 Decreased in North America but Increased in Europe and Asia. Proc. Natl. Acad. Sci. USA 2020, 117, 12192–12200. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Powers, J.; Cochard, H.; Choat, B. Hanging by a Thread? Forests and Drought. Science 2020, 368, 261–266. [Google Scholar] [CrossRef]
- Zellweger, F.; De Frenne, P.; Lenoir, J.; Vangansbeke, P.; Verheyen, K.; Bernhardt-Römermann, M.; Baeten, L.; Hédl, R.; Berki, I.; Brunet, J.; et al. Forest Microclimate Dynamics Drive Plant Responses to Warming. Science 2020, 368, 772–775. [Google Scholar] [CrossRef]
- Perring, M.P.; Diekmann, M.; Midolo, G.; Schellenberger Costa, D.; Bernhardt-Römermann, M.; Otto, J.C.J.; Gilliam, F.S.; Hedwall, P.-O.; Nordin, A.; Dirnböck, T.; et al. Understanding Context Dependency in the Response of Forest Understorey Plant Communities to Nitrogen Deposition. Environ. Pollut. 2018, 242, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Staude, I.R.; Pereira, H.M.; Daskalova, G.N.; Bernhardt-Römermann, M.; Diekmann, M.; Pauli, H.; Van Calster, H.; Vellend, M.; Bjorkman, A.D.; Brunet, J.; et al. Directional Turnover towards Larger-Ranged Plants over Time and across Habitats. Ecol. Lett. 2022, 25, 466–482. [Google Scholar] [CrossRef] [PubMed]
- Wagner, V.; Chytrý, M.; Jiménez-Alfaro, B.; Pergl, J.; Hennekens, S.; Biurrun, I.; Knollová, I.; Berg, C.; Vassilev, K.; Rodwell, J.S.; et al. Alien Plant Invasions in European Woodlands. Divers. Distrib. 2017, 23, 969–981. [Google Scholar] [CrossRef]
- Saout, S.L.; Hoffmann, M.; Shi, Y.; Hughes, A.; Bernard, C.; Brooks, T.M.; Bertzky, B.; Butchart, S.H.M.; Stuart, S.N.; Badman, T.; et al. Protected Areas and Effective Biodiversity Conservation. Science 2013, 342, 803–805. [Google Scholar] [CrossRef] [PubMed]
- Vačkář, D.; Harmáčková, Z.V.; Kaňková, H.; Stupková, K. Human Transformation of Ecosystems: Comparing Protected and Unprotected Areas with Natural Baselines. Ecol. Indic. 2016, 66, 321–328. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, Diversity, and Invasion: Implications for Conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A Meta-Analysis of Biotic Resistance to Exotic Plant Invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, B.; Aldridge, D.C.; González-Moreno, P.; Pergl, J.; Pizarro, M.; Pyšek, P.; Thuiller, W.; Yesson, C.; Vilà, M. Protected Areas Offer Refuge from Invasive Species Spreading under Climate Change. Glob. Chang. Biol. 2017, 23, 5331–5343. [Google Scholar] [CrossRef] [Green Version]
- Geldmann, J.; Barnes, M.; Coad, L.; Craigie, I.D.; Hockings, M.; Burgess, N.D. Effectiveness of Terrestrial Protected Areas in Reducing Habitat Loss and Population Declines. Biol. Conserv. 2013, 161, 230–238. [Google Scholar] [CrossRef]
- Rija, A.A.; Critchlow, R.; Thomas, C.D.; Beale, C.M. Global Extent and Drivers of Mammal Population Declines in Protected Areas under Illegal Hunting Pressure. PLoS ONE 2020, 15, e0227163. [Google Scholar] [CrossRef]
- Clark, N.E.; Boakes, E.H.; McGowan, P.J.K.; Mace, G.M.; Fuller, R.A. Protected Areas in South Asia Have Not Prevented Habitat Loss: A Study Using Historical Models of Land-Use Change. PLoS ONE 2013, 8, e65298. [Google Scholar] [CrossRef] [PubMed]
- Galford, G.L.; Soares-Filho, B.S.; Sonter, L.J.; Laporte, N. Will Passive Protection Save Congo Forests? PLoS ONE 2015, 10, e0128473. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, S.L.; Cazalis, V.; Dudley, N.; Hoffmann, M.; Rodrigues, A.S.L.; Stolton, S.; Visconti, P.; Woodley, S.; Kingston, N.; Lewis, E.; et al. Area-Based Conservation in the Twenty-First Century. Nature 2020, 586, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Lecina-Diaz, J.; Alvarez, A.; De Cáceres, M.; Herrando, S.; Vayreda, J.; Retana, J. Are Protected Areas Preserving Ecosystem Services and Biodiversity? Insights from Mediterranean Forests and Shrublands. Landsc. Ecol. 2019, 34, 2307–2321. [Google Scholar] [CrossRef]
- Foxcroft, L.C.; Pyšek, P.; Richardson, D.M.; Genovesi, P. Plant Invasions in Protected Areas: Patterns, Problems and Challenges; Springer Science & Business Media: Dordrecht, The Netherlands, 2013; ISBN 9789400777507. [Google Scholar]
- Chmura, D.; Sierka, E. The Occurrence of Invasive Alien Plant Species in Selected Forest Nature Reserves in Southern Poland as a Conservation Problem. Nat. Conserv. 2006, 62, 3–11. [Google Scholar]
- Chmura, D.; Gucwa-Przepióra, E. Interactions between Arbuscular Mycorrhiza and the Growth of the Invasive Alien Annual Impatiens Parviflora DC: A Study of Forest Type and Soil Properties in Nature Reserves (S Poland). Appl. Soil Ecol. 2012, 62, 71–80. [Google Scholar] [CrossRef]
- Riitters, K.; Potter, K.M.; Iannone, B.V.; Oswalt, C.; Guo, Q.; Fei, S. Exposure of Protected and Unprotected Forest to Plant Invasions in the Eastern United States. For. Trees Livelihoods 2018, 9, 723. [Google Scholar] [CrossRef] [Green Version]
- Lapin, K.; Oettel, J.; Steiner, H.; Langmaier, M.; Sustic, D.; Starlinger, F.; Kindermann, G.; Frank, G. Invasive Alien Plant Species in Unmanaged Forest Reserves, Austria. NeoBiota 2019, 48, 71–96. [Google Scholar] [CrossRef] [Green Version]
- Reczyńska, K.; Świerkosz, K. Does Protection Really Matter? A Case Study from Central European Oak Forests. Diversity 2019, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Hughes, R.F.; Vitousek, P.M.; Knapp, D.E.; Kennedy-Bowdoin, T.; Boardman, J.; Martin, R.E.; Eastwood, M.; Green, R.O. Invasive Plants Transform the Three-Dimensional Structure of Rain Forests. Proc. Natl. Acad. Sci. USA 2008, 105, 4519–4523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florens, F.B.V.; Baider, C.; Martin, G.M.N.; Seegoolam, N.B.; Zmanay, Z.; Strasberg, D. Invasive Alien Plants Progress to Dominate Protected and Best-Preserved Wet Forests of an Oceanic Island. J. Nat. Conserv. 2016, 34, 93–100. [Google Scholar] [CrossRef]
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; et al. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES: Bonn, Germany, 2019. [Google Scholar]
- Pyšek, P.; Jarošík, V.; Kučera, T. Inclusion of Native and Alien Species in Temperate Nature Reserves: An Historical Study from Central Europe. Conserv. Biol. 2003, 17, 1414–1424. [Google Scholar] [CrossRef]
- Matlack, G.R.; Monde, J. Consequences of Low Mobility in Spatially and Temporally Heterogeneous Ecosystems. J. Ecol. 2004, 92, 1025–1035. [Google Scholar] [CrossRef]
- EU Biodiversity Strategy for 2030: Bringing Nature Back into Our Lives; Publications Office of the European Union: Luxembourg, 2021.
- Directorate-General for Environment. Criteria and Guidance for Protected Areas Designations—Staff Working Document; European Comission: Brussels, Belgium, 2022. [Google Scholar]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where Are Europe’s Last Primary Forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef] [Green Version]
- Sabatini, F.M.; Bluhm, H.; Kun, Z.; Aksenov, D.; Atauri, J.A.; Buchwald, E.; Burrascano, S.; Cateau, E.; Diku, A.; Duarte, I.M.; et al. European Primary Forest Database v2.0. Sci. Data 2021, 8, 220. [Google Scholar] [CrossRef]
- Baran, J.; Pielech, R.; Bodziarczyk, J. No Difference in Plant Species Diversity between Protected and Managed Ravine Forests. For. Ecol. Manag. 2018, 430, 587–593. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional Richness, Functional Evenness and Functional Divergence: The Primary Components of Functional Diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- De Bello, F.; Lavorel, S.; Lavergne, S.; Albert, C.H.; Boulangeat, I.; Mazel, F.; Thuiller, W. Hierarchical Effects of Environmental Filters on the Functional Structure of Plant Communities: A Case Study in the French Alps. Ecography 2013, 36, 393–402. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the Concept of Trait Be Functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Lavorel, S.; McIntyre, S.; Landsberg, J.; Forbes, T.D. Plant Functional Classifications: From General Groups to Specific Groups Based on Response to Disturbance. Trends Ecol. Evol. 1997, 12, 474–478. [Google Scholar] [CrossRef]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bonisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—a Global Database of Plant Traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Díaz, S.; Purvis, A.; Cornelissen, J.H.C.; Mace, G.M.; Donoghue, M.J.; Ewers, R.M.; Jordano, P.; Pearse, W.D. Functional Traits, the Phylogeny of Function, and Ecosystem Service Vulnerability. Ecol. Evol. 2013, 3, 2958–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenderes, K.; Standovár, T. The Impact of Forest Management on Forest Floor Vegetation Evaluated by Species Traits. Community Ecol. 2003, 4, 51–62. [Google Scholar] [CrossRef]
- Dölle, M.; Petritan, A.M.; Biris, I.A.; Petritan, I.C. Relations between Tree Canopy Composition and Understorey Vegetation in a European Beech-Sessile Oak Old Growth Forest in Western Romania. Biologia 2017, 72, 1422–1430. [Google Scholar] [CrossRef]
- Ganivet, E.; Flores, O.; Balguerías, E.; de Nascimento, L.; Arévalo, J.R.; Fernández-Lugo, S.; Fernández-Palacios, J.M. Ecological Strategies of Tree Species in the Laurel Forest of Tenerife (Canary Islands): An Insight into Cloud Forest Natural Dynamics Using Long-Term Monitoring Data. Eur. J. For. Res. 2019, 138, 93–110. [Google Scholar] [CrossRef]
- Selwood, K.E.; Zimmer, H.C. Refuges for Biodiversity Conservation: A Review of the Evidence. Biol. Conserv. 2020, 245, 108502. [Google Scholar] [CrossRef]
- Pielech, R. Plant Species Richness in Riparian Forests: Comparison to Other Forest Ecosystems, Longitudinal Patterns, Role of Rare Species and Topographic Factors. For. Ecol. Manag. 2021, 496, 119400. [Google Scholar] [CrossRef]
- Krzemińska, A. Dynamika Zmian Warunków Wodnych W Lasach Na Terenach Polderowych—Analiza Interakcji; Uniwersytet Przyrodniczy we Wrocławiu: Wrocław, Poland, 2008. [Google Scholar]
- Jankowski, W.; Świerkosz, K. (Eds.) Korytarz Ekologiczny Doliny Odry—Stan-Funkcjonowanie-Zagrożenia; IUCN: Warszawa, Poland, 1995. [Google Scholar]
- Blachowski, J.; Markowicz-Judycka, E.; Zięba, D. Opracowanie Ekofizjograficzne dla Województwa Dolnośląskiego; Wojewódzkie Biuro Urbanistyczne: Wrocław, Poland, 2005; ISBN 9788391347256. [Google Scholar]
- Świerkosz, K.; Liberadzka, H.; Łysiak, M.; Zając, K. (Eds.) Obszary Natura 2000 Na Dolnym Śląsku; Regionalna Dyrekcja Ochrony Środowiska we Wrocławiu: Wrocław, Poland, 2012. [Google Scholar]
- Kuczyńska, I. Zbiorowiska Leśne Rezerwatów W Nadleśnictwie Oława. Prace Botaniczne 1965, 6, 1–29. [Google Scholar]
- Anioł-Kwiatkowska, J. Roślinność Rezerwatu “Grodzińska Ryczyńskie”. Prace Botaniczne 1995, 62, 39–52. [Google Scholar]
- Anioł-Kwiatkowska, J.; Weretelnik, E. Flora I Roślinność Rezerwatu “Zwierzyniec” Koło Oławy. Prace Botaniczne 1995, 62, 287–303. [Google Scholar]
- Anioł-Kwiatkowska, J.; Weretelnik, E. Flora I Roślinność Rezerwatu “Kanigóra”. Prace Botaniczne 1995, 62, 67–78. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Reprint; The Blackburn Press: Caldwell, NJ, USA, 2002; ISBN 9781930665736. [Google Scholar]
- Novák, P.; Willner, W.; Zukal, D.; Kollár, J.; Roleček, J.; Świerkosz, K.; Ewald, J.; Wohlgemuth, T.; Csiky, J.; Onyshchenko, V.; et al. Oak-Hornbeam Forests of Central Europe: A Formalized Classification and Syntaxonomic Revision. Preslia 2020, 92, 1–34. [Google Scholar] [CrossRef]
- Euro+Med PlantBase—the Information Resource for Euro-Mediterranean Plant Diversity. 2006. Available online: https://ww2.bgbm.org/EuroPlusMed/query.asp (accessed on 20 February 2022).
- Tichý, L. JUICE, Software for Vegetation Classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Roleček, J.; Tichý, L.; Zelený, D.; Chytrý, M. Modified TWINSPAN Classification in Which the Hierarchy Respects Cluster Heterogeneity. J. Veg. Sci. 2009, 20, 596–602. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Holt, J.; Botta-Dukát, Z. Determination of Diagnostic Species with Statistical Fidelity Measures. J. Veg. Sci. 2002, 13, 79–90. [Google Scholar] [CrossRef]
- Dengler, J.; Berg, C.; Jansen, F. New Ideas for Modern Phytosociological Monographs. Annali di Botanica 2005, 5, 195–213. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012; ISBN 9788362940349. [Google Scholar]
- Verheyen, K.; Baeten, L.; De Frenne, P.; Bernhardt-Römermann, M.; Brunet, J.; Cornelis, J.; Decocq, G.; Dierschke, H.; Eriksson, O.; Hédl, R.; et al. Driving Factors behind the Eutrophication Signal in Understorey Plant Communities of Deciduous Temperate Forests. J. Ecol. 2012, 100, 352–365. [Google Scholar] [CrossRef]
- Šipoš, J.; Chudomelová, M.; Vild, O.; Macek, M.; Kopecký, M.; Szabó, P.; Hédl, R. Plant Diversity in Deciduous Temperate Forests Reflects Interplay among Ancient and Recent Environmental Stress. J. Veg. Sci. 2020, 31, 53–62. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, W.; Werner, W.; Paulien, D. Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica, 18, 1–258. Scr. Geobot. 1991, 18, 1–258. [Google Scholar]
- Berg, C.; Welk, E.; Jäger, E.J. Revising Ellenberg’s Indicator Values for Continentality Based on Global Vascular Plant Species Distribution. Appl. Veg. Sci. 2017, 20, 482–493. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Zelený, D.; Schaffers, A.P. Too Good to Be True: Pitfalls of Using Mean Ellenberg Indicator Values in Vegetation Analyses. J. Veg. Sci. 2012, 23, 419–431. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Gleason, J. Comparative Power of the Anova, Randomization Anova, and Kruskal-Wallis Test. Ph.D. Thesis, Wayne State University, Detroit, MI, USA, 2013. [Google Scholar]
- Westoby, M. A Leaf-Height-Seed (LHS) Plant Ecology Strategy Scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting Changes in Community Composition and Ecosystem Functioning from Plant Traits: Revisiting the Holy Grail: Plant Response and Effect Groups. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Czortek, P.; Orczewska, A. Niche Differentiation, Competition or Habitat Filtering? Mechanisms Explaining Co-occurrence of Plant Species on Wet Meadows of High Conservation Value. J. Veg. Sci. 2021, 32, e12983. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New Handbook for Standardised Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global Patterns in Plant Height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Garnier, E.; Lavorel, S.; Ansquer, P.; Castro, H.; Cruz, P.; Doležal, J.; Eriksson, O.; Freitas, H.; Golodets, C.; et al. Impact of Abundance Weighting on the Response of Seed Traits to Climate and Land Use. J. Ecol. 2008, 96, 355–366. [Google Scholar] [CrossRef]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA Traitbase: A Database of Life-History Traits of the Northwest European Flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Paź-Dyderska, S.; Dyderski, M.K.; Szwaczka, P. Leaf Traits and Aboveground Biomass Variability of Forest Understory Herbaceous Plant Species. Ecosystems 2020, 23, 555–569. [Google Scholar] [CrossRef] [Green Version]
- Pakeman, R.J. Functional Trait Metrics Are Sensitive to the Completeness of the Species’ Trait Data? Methods Ecol. Evol. 2014, 5, 9–15. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing Functional Diversity in the Field—Methodology Matters! Funct. Ecol. 2007, 22, 134–147. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New Multidimensional Functional Diversity Indices for a Multifaceted Framework in Functional Ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Mason, N.W.H.; de Bello, F.; Mouillot, D.; Pavoine, S.; Dray, S. A Guide for Using Functional Diversity Indices to Reveal Changes in Assembly Processes along Ecological Gradients. J. Veg. Sci. 2013, 24, 794–806. [Google Scholar] [CrossRef]
- Garnier, E.; Lavorel, S.; Ansquer, P.; Castro, H.; Cruz, P.; Dolezal, J.; Eriksson, O.; Fortunel, C.; Freitas, H.; Golodets, C.; et al. Assessing the Effects of Land-Use Change on Plant Traits, Communities and Ecosystem Functioning in Grasslands: A Standardized Methodology and Lessons from an Application to 11 European Sites. Ann. Bot. 2007, 99, 967–985. [Google Scholar] [CrossRef] [Green Version]
- Botta-Dukát, Z. Rao’s Quadratic Entropy as a Measure of Functional Diversity Based on Multiple Traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Loster, S. Wskaźnikowe Gatunki Roślin Starych Lasów I Ich Znaczenie Dla Ochrony Przyrody I Kartografii Roślinności. Prace Geograficzne 2001, 178, 119–132. [Google Scholar]
- Richardson, D.M.; Holmes, P.M.; Esler, K.J.; Galatowitsch, S.M.; Stromberg, J.C.; Kirkman, S.P.; Pyšek, P.; Hobbs, R.J. Riparian Vegetation: Degradation, Alien Plant Invasions, and Restoration Prospects. Divers. Distrib. 2007, 13, 126–139. [Google Scholar] [CrossRef]
- Douda, J.; Boublík, K.; Slezák, M.; Biurrun, I.; Nociar, J.; Havrdová, A.; Doudová, J.; Aćić, S.; Brisse, H.; Brunet, J.; et al. Vegetation Classification and Biogeography of European Floodplain Forests and Alder Carrs. Appl. Veg. Sci. 2016, 19, 147–163. [Google Scholar] [CrossRef]
- Staude, I.R.; Waller, D.M.; Bernhardt-Römermann, M.; Bjorkman, A.D.; Brunet, J.; De Frenne, P.; Hédl, R.; Jandt, U.; Lenoir, J.; Máliš, F.; et al. Replacements of Small- by Large-Ranged Species Scale up to Diversity Loss in Europe’s Temperate Forest Biome. Nat. Ecol. Evol. 2020, 4, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catford, J.A.; Daehler, C.C.; Murphy, H.T.; Sheppard, A.W.; Hardesty, B.D.; Westcott, D.A.; Rejmánek, M.; Bellingham, P.J.; Pergl, J.; Horvitz, C.C.; et al. The Intermediate Disturbance Hypothesis and Plant Invasions: Implications for Species Richness and Management. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 231–241. [Google Scholar] [CrossRef]
- Boch, S.; Prati, D.; Müller, J.; Socher, S.; Baumbach, H.; Buscot, F.; Gockel, S.; Hemp, A.; Hessenmöller, D.; Kalko, E.K.V.; et al. High Plant Species Richness Indicates Management-Related Disturbances rather than the Conservation Status of Forests. Basic Appl. Ecol. 2013, 14, 496–505. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Baeten, L.; Craven, D.; De Frenne, P.; Hédl, R.; Lenoir, J.; Bert, D.; Brunet, J.; Chudomelová, M.; Decocq, G.; et al. Drivers of Temporal Changes in Temperate Forest Plant Diversity Vary across Spatial Scales. Glob. Chang. Biol. 2015, 21, 3726–3737. [Google Scholar] [CrossRef]
- Naaf, T.; Kolk, J. Initial Site Conditions and Interactions between Multiple Drivers Determine Herb-Layer Changes over Five Decades in Temperate Forests. For. Ecol. Manag. 2016, 366, 153–165. [Google Scholar] [CrossRef]
- Verstraeten, G.; Baeten, L.; Van den Broeck, T.; De Frenne, P.; Demey, A.; Tack, W.; Muys, B.; Verheyen, K. Temporal Changes in Forest Plant Communities at Different Site Types. Appl. Veg. Sci. 2013, 16, 237–247. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Kučera, T. Patterns of Invasion in Temperate Nature Reserves. Biol. Conserv. 2002, 104, 13–24. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Römermann, C.; Pillar, V.D.; Kudernatsch, T.; Fischer, A. High Functional Diversity Is Related to High Nitrogen Availability in a Deciduous Forest—Evidence from a Functional Trait Approach. Folia Geobotanica 2010, 45, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Härdtle, W.; Bergmeier, E.; Fichtner, A.; Heinken, T.; Hölzel, N.; Remy, D.; Schneider, S.; Schwabe, A.; Tischew, S.; Dierschke, H. Pflanzengesellschaft Des Jahres 2021: Hartholz-Auenwald (Ficario-Ulmetum). Tuexenia 2020, 40, 373–399. [Google Scholar] [CrossRef]
- Reczyńska, K.; Świerkosz, K. Compositional Changes in Thermophilous Oak Forests in Poland over Time: Do They Correspond to European Trends? Appl. Veg. Sci. 2017, 20, 293–303. [Google Scholar] [CrossRef]
- Gioria, M.; Osborne, B.A. Resource Competition in Plant Invasions: Emerging Patterns and Research Needs. Front. Plant Sci. 2014, 5, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grime, J.P.; Hodgson, J.G.; Hunt, R. Comparative Plant Ecology. A Functional Approach to Common British Species; Springer-Science+Business Media, B.V.: Dordrecht, The Netherlands, 1988; ISBN 9780412741708. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date of Investigation | 1960 | 1990 | 2020 | |
|---|---|---|---|---|
| No. of Relevés | 42 | 42 | 42 | |
| Ratio | ||||
| Aegopodium podagraria | 1.7 | 35 86 | __ 50 | __ 50 |
| Symphytum tuberosum | 2.8 | 29 26 | __ 2 | __ 10 |
| Paris quadrifolia | 2.2 | 29 45 | __ 21 | __ 14 |
| Ajuga reptans | 2.1 | 27 29 | __ 12 | __ 5 |
| Elymus caninus | 100 | __ . | 48 31 | __ . |
| Deschampsia cespitosa | 5.4 | __ . | 37 26 | __ 5 |
| Impatiens parviflora | 4 | __ . | __ 23 | 78 93 |
| Chelidonium majus | 16.4 | __ . | __ 2 | 51 38 |
| Galeopsis tetrahit | 3.8 | __ 12 | __ 2 | 45 45 |
| Stellaria media | 2.5 | __ 10 | __ 21 | 39 52 |
| Chaerophyllum aromaticum | 4.4 | __ . | __ 7 | 39 31 |
| Date of Investigation | 1960 | 1990 | 2020 |
|---|---|---|---|
| No. of Relevés | 42 | 42 | 42 |
| Impatiens parviflora | . | 24 (2) | 93 (14) |
| Ballota nigra | . | 2 (1) | 5 (1) |
| Solidago gigantea | . | . | 10 (1) |
| Juglans regia (juv.) | . | . | 5 (2) |
| Sonchus oleraceus | . | . | 2 (1) |
| Quercus rubra (juv.) | . | . | 2 (1) |
| Aesculus hippocastanum (juv.) | . | . | 2 (2) |
| Axis 1 | Axis 2 | Axis 3 | Axis 4 | |||||
|---|---|---|---|---|---|---|---|---|
| Rho | P. Modif | Rho | P. Modif | Rho | P. Modif | Rho | P. Modif | |
| Light | 0.345 | 0.100 | −0.150 | 0.388 | 0.277 | 0.088 | 0.115 | 0.472 |
| Temperature | 0.123 | 0.616 | 0.006 | 0.992 | 0.118 | 0.476 | −0.265 | 0.088 |
| Continentality | −0.221 | 0.432 | 0.082 | 0.720 | 0.162 | 0.296 | −0.015 | 0.932 |
| Moisture | −0.221 | 0.360 | −0.500 | 0.012 | 0.162 | 0.316 | −0.015 | 0.960 |
| Soil reaction | −0.268 | 0.244 | −0.341 | 0.044 | −0.286 | 0.056 | 0.222 | 0.236 |
| Nutrients | 0.069 | 0.732 | −0.133 | 0.480 | 0.149 | 0.356 | −0.129 | 0.368 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reczyńska, K.; Orczewska, A.; Yurchenko, V.; Wójcicka-Rosińska, A.; Świerkosz, K. Changes in Species and Functional Diversity of the Herb Layer of Riparian Forest despite Six Decades of Strict Protection. Forests 2022, 13, 747. https://doi.org/10.3390/f13050747
Reczyńska K, Orczewska A, Yurchenko V, Wójcicka-Rosińska A, Świerkosz K. Changes in Species and Functional Diversity of the Herb Layer of Riparian Forest despite Six Decades of Strict Protection. Forests. 2022; 13(5):747. https://doi.org/10.3390/f13050747
Chicago/Turabian StyleReczyńska, Kamila, Anna Orczewska, Valeriia Yurchenko, Anna Wójcicka-Rosińska, and Krzysztof Świerkosz. 2022. "Changes in Species and Functional Diversity of the Herb Layer of Riparian Forest despite Six Decades of Strict Protection" Forests 13, no. 5: 747. https://doi.org/10.3390/f13050747