
The Dry and the Wet Case: Tree Growth Response in Climatologically Contrasting Years on the Island of Corsica

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
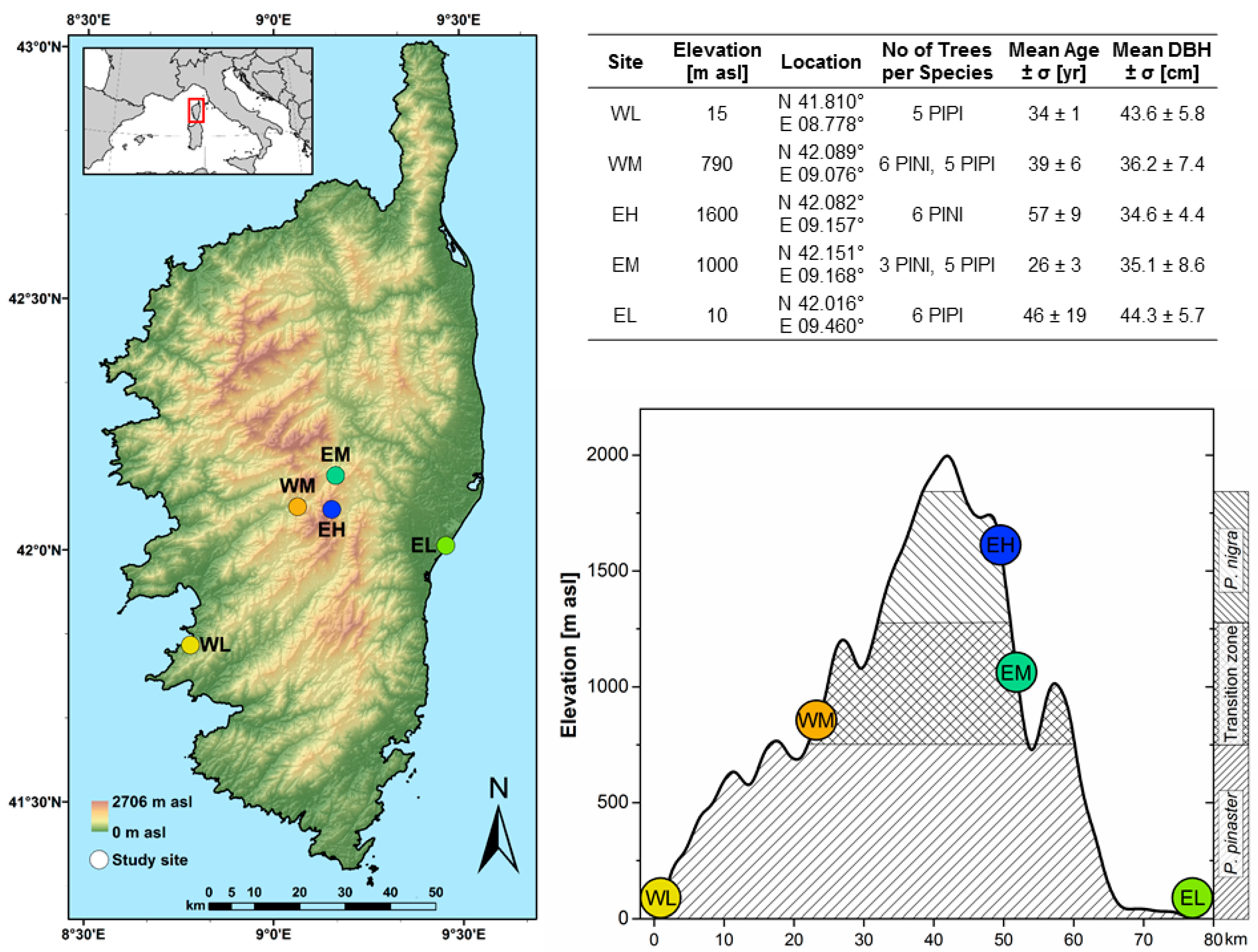
2.1. Study Area and Species
2.2. Dendroecological Methods
2.3. Meteorological Methods
3. Results and Discussion
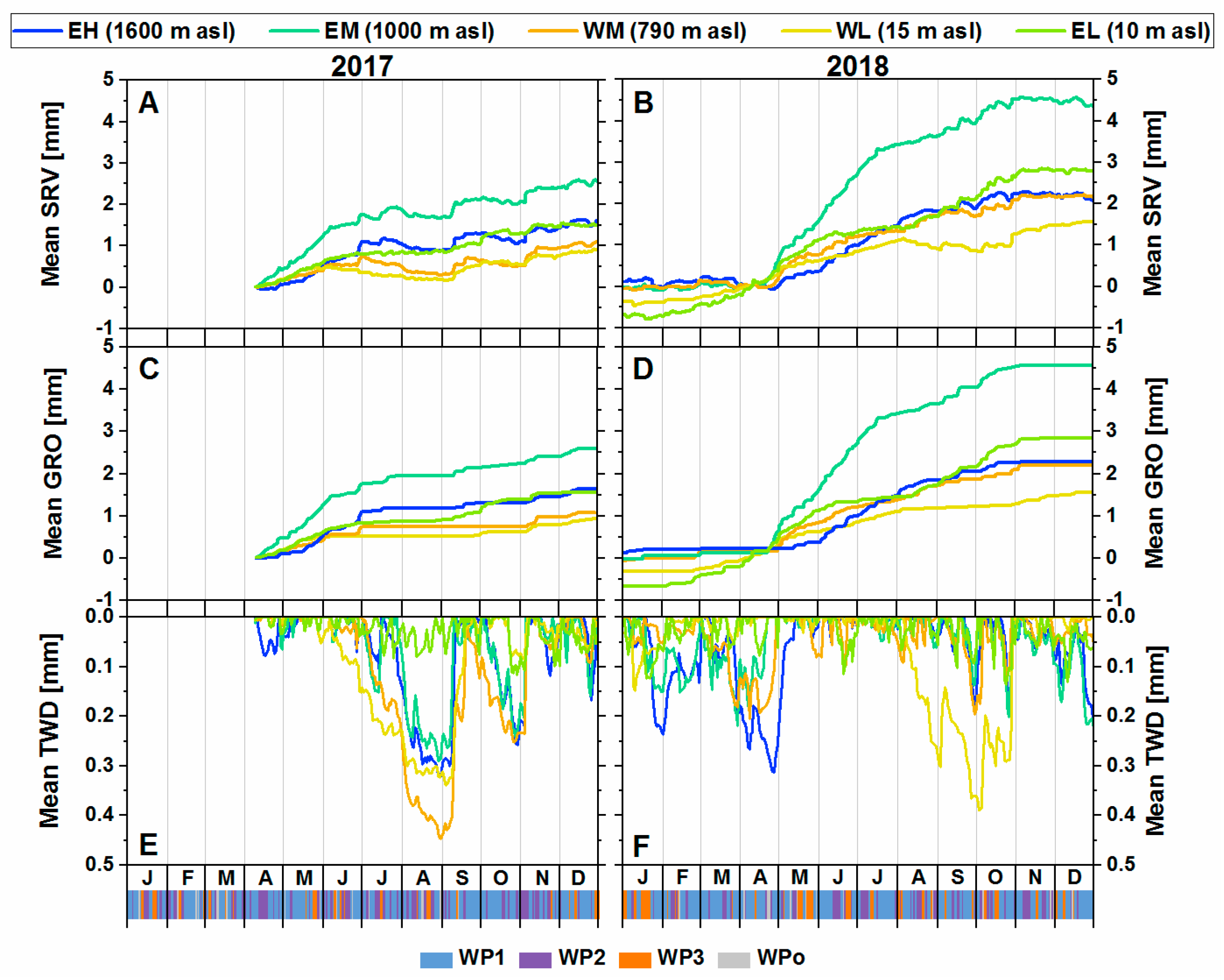
3.1. Stem Radial Variations
3.1.1. Growth Response in 2017 and 2018
3.1.2. Irreversible Growth and Tree Water Deficit
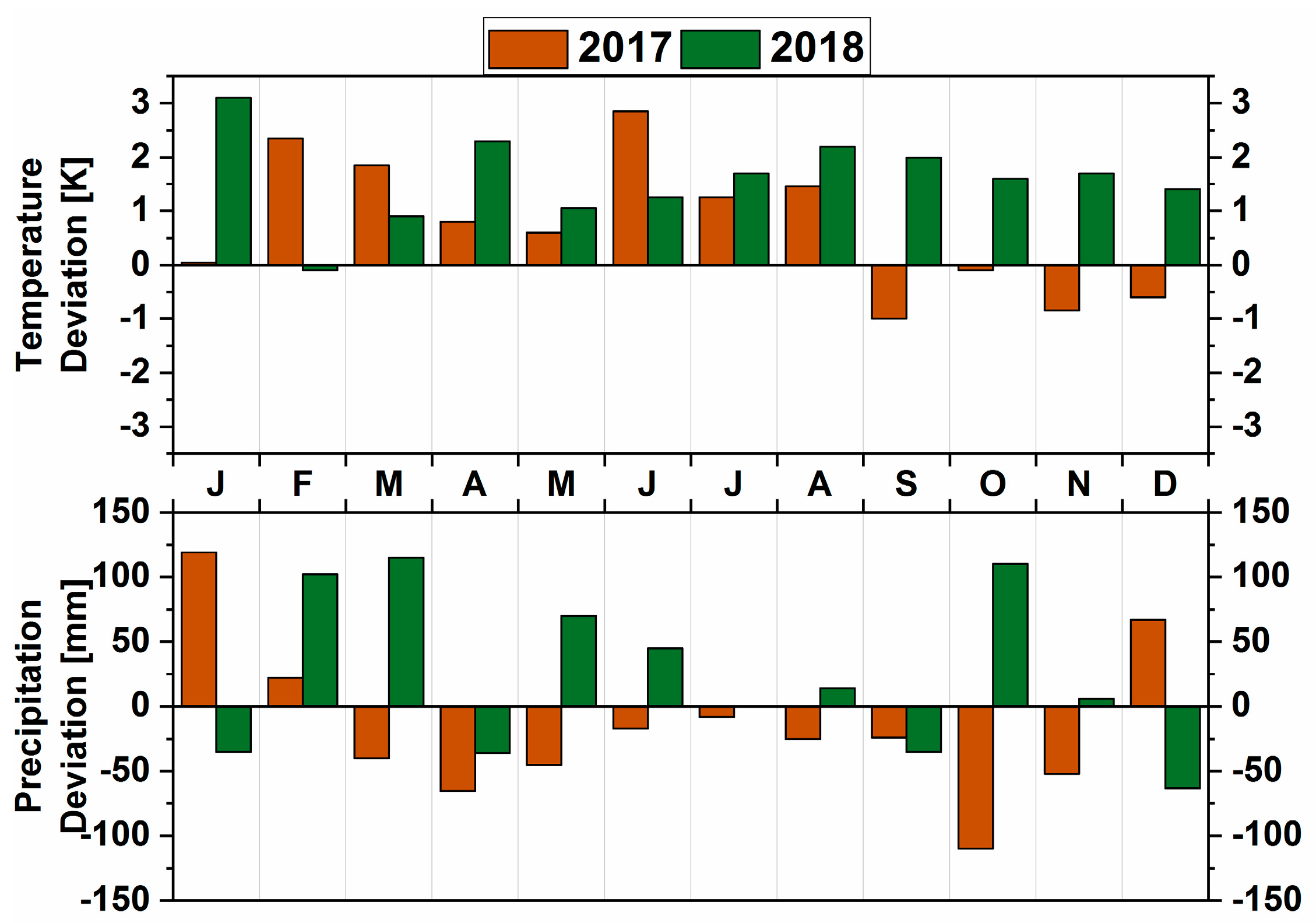
3.2. Meteorological Parameters
3.2.1. Comparison of the Contrasting Years 2017 and 2018
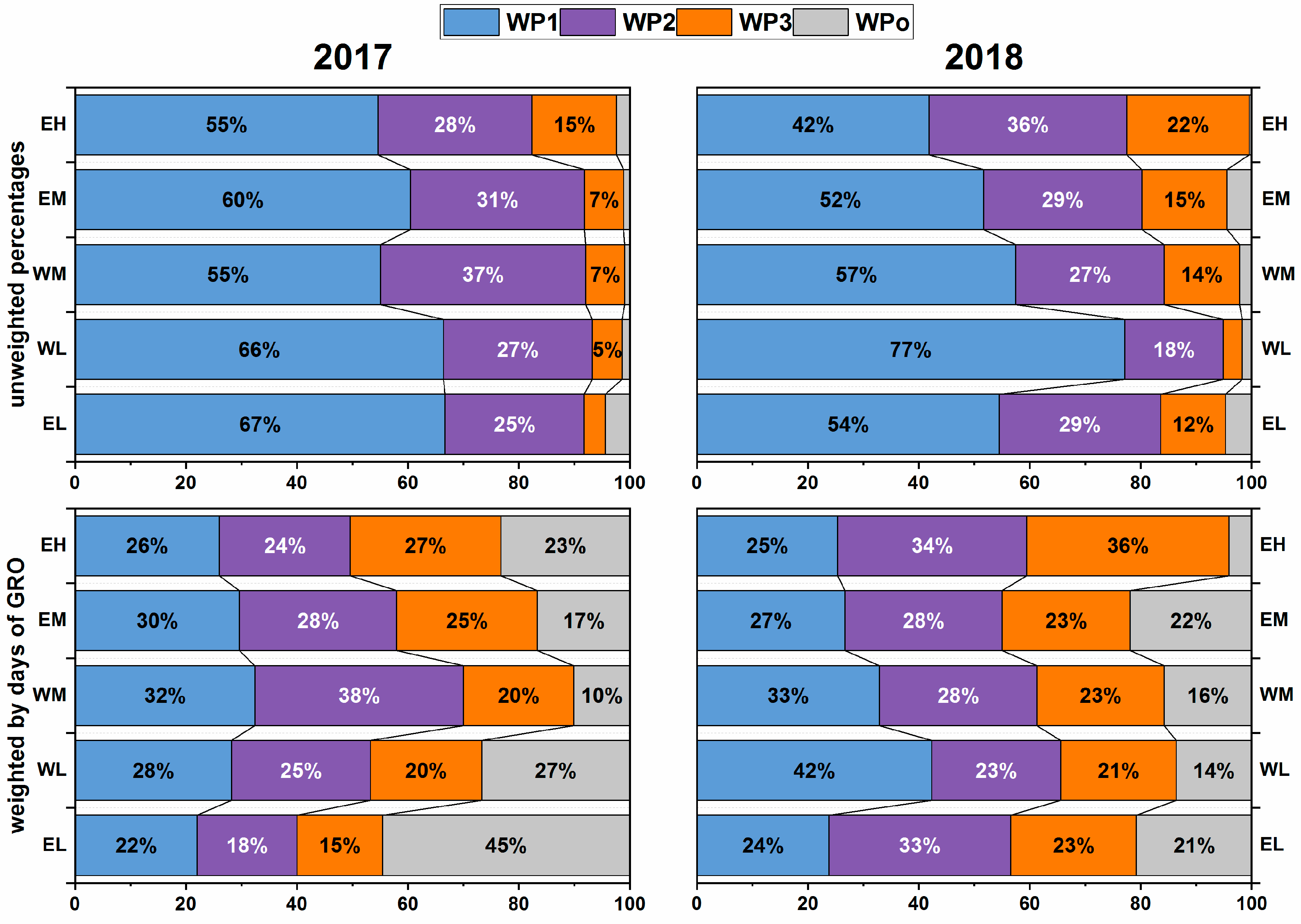
3.2.2. Frequency of the Prevalent Weather Patterns and their Contribution to Precipitation
3.3. Integration of Meteorology and Tree Growth Response
3.3.1. Site-Specific Precipitation Regimes and Respective Impact on Tree Growth
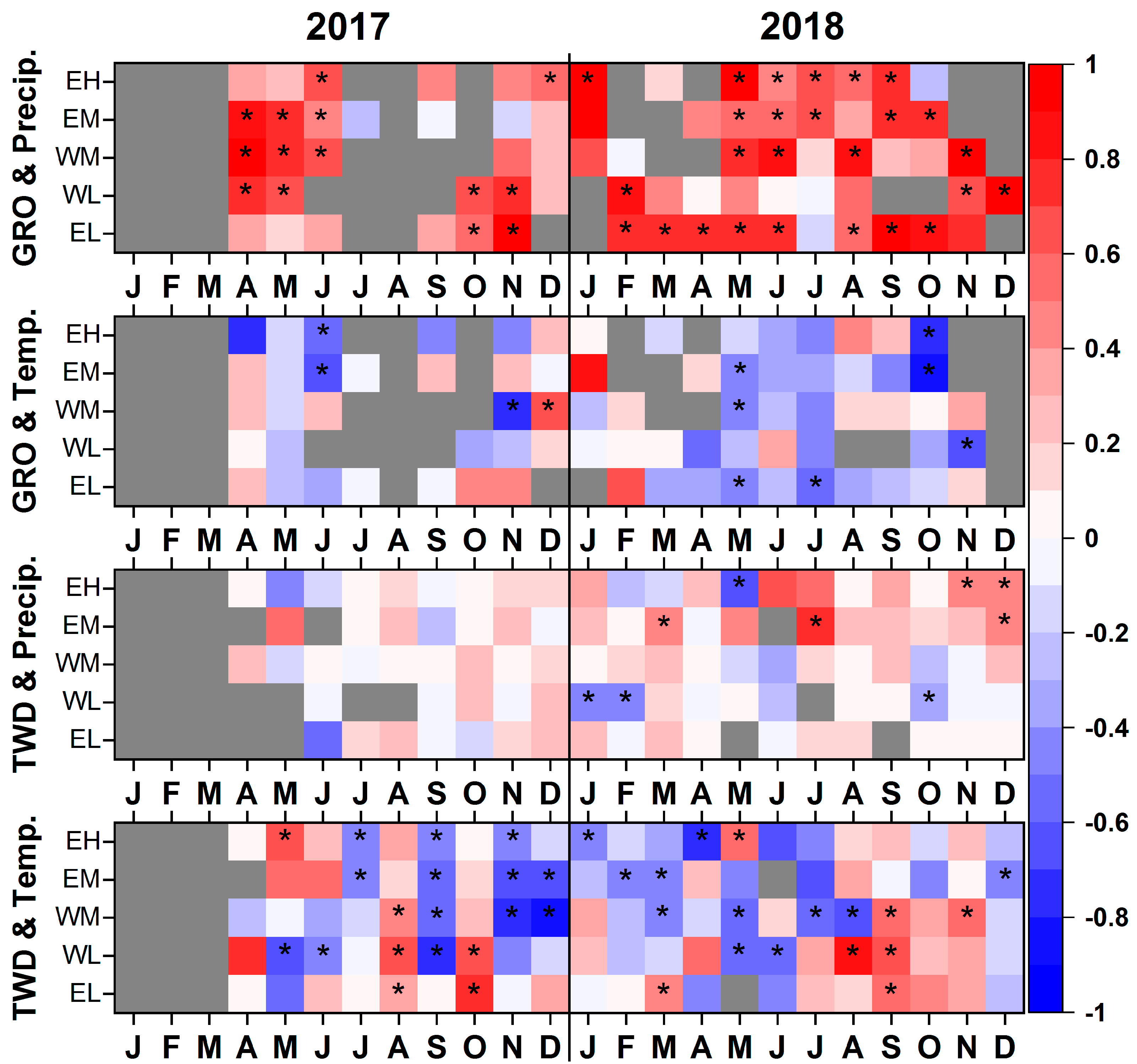
3.3.2. Correlations between Meteorological and Growth Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovern-Mental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Nardini, A.; Lo Gullo, M.A.; Trifilò, P.; Salleo, S. The challenge of the Mediterranean climate to plant hydraulics: Responses and adaptations. Environ. Exp. Bot. 2014, 103, 68–79. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2015. [Google Scholar]
- Beniston, M.; Diaz, H.F. The 2003 heat wave as an example of summers in a greenhouse climate? Observations and climate model simulations for Basel, Switzerland. Glob. Planet. Chang. 2004, 44, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Serrano, S.M.; Lopez-Moreno, J.-I.; Beguería, S.; Lorenzo-Lacruz, J.; Sanchez-Lorenzo, A.; García-Ruiz, J.M.; Azorin-Molina, C.; Morán-Tejeda, E.; Revuelto, J.; Trigo, R.; et al. Evidence of increasing drought severity caused by temperature rise in southern Europe. Environ. Res. Lett. 2014, 9, 44001. [Google Scholar] [CrossRef]
- Zittis, G.; Hadjinicolaou, P.; Fnais, M.; Lelieveld, J. Projected changes in heat wave characteristics in the eastern Mediterranean and the Middle East. Reg. Environ. Chang. 2016, 16, 1863–1876. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, art129. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Bigler, C.; Bräker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an Inciting Mortality Factor in Scots Pine Stands of the Valais, Switzerland. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Akkemik, Ü. Tree Rings of Cedrus libani at the Northern Boundary of its Natural Distribution. IAWA J. 2003, 24, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Touchan, R.; Xoplaki, E.; Funkhouser, G.; Luterbacher, J.; Hughes, M.K.; Erkan, N.; Akkemik, Ü.; Stephan, J. Reconstructions of spring/summer precipitation for the Eastern Mediterranean from tree-ring widths and its connection to large-scale atmospheric circulation. Clim. Dyn. 2005, 25, 75–98. [Google Scholar] [CrossRef]
- Schuster, R.; Oberhuber, W. Drought sensitivity of three co-occurring conifers within a dry inner Alpine environment. Trees 2013, 27, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Häusser, M.; Szymczak, S.; Garel, E.; Santoni, S.; Huneau, F.; Bräuning, A. Growth variability of two native pine species on Corsica as a function of elevation. Dendrochronologia 2019, 54, 49–55. [Google Scholar] [CrossRef]
- Galván, J.; Büntgen, U.; Ginzler, C.; Grudd, H.; Gutiérrez, E.; Labuhn, I.; Julio Camarero, J. Drought-induced weakening of growth–temperature associations in high-elevation Iberian pines. Glob. Planet. Chang. 2015, 124, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Bouriaud, O.; Leban, J.-M.; Bert, D.; Deleuze, C. Intra-annual variations in climate influence growth and wood density of Norway spruce. Tree Physiol. 2005, 25, 651–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathgeber, C.B.K.; Rossi, S.; Bontemps, J.-D. Cambial activity related to tree size in a mature silver-fir plantation. Ann. Bot. 2011, 108, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Deslauriers, A.; Morin, H.; Begin, Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Can. J. For. Res. 2003, 33, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Linares, J.C.; Camarero, J.J.; Carreira, J.A. Plastic responses of Abies pinsapo xylogenesis to drought and competition. Tree Physiol. 2009, 29, 1525–1536. [Google Scholar] [CrossRef]
- Michelot, A.; Simard, S.; Rathgeber, C.; Dufrêne, E.; Damesin, C. Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol. 2012, 32, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood Formation in Trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef]
- Zhai, L.; Bergeron, Y.; Huang, J.-G.; Berninger, F. Variation in intra-annual wood formation, and foliage and shoot development of three major Canadian boreal tree species. Am. J. Bot. 2012, 99, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Xylogenesis of Pinus pinaster under a Mediterranean climate. Ann. For. Sci. 2014, 71, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Nie, Y.; Wen, F. Seasonal Dynamics of Stem Radial Increment of Pinus taiwanensis Hayata and Its Response to Environmental Factors in the Lushan Mountains, Southeastern China. Forests 2018, 9, 387. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Huang, J.-G.; Rossi, S.; Ma, Q.; Yu, B.; Zhai, L.; Luo, D.; Guo, X.; Fu, S.; Zhang, W. Intra-annual dynamics of xylem growth in Pinus massoniana submitted to an experimental nitrogen addition in Central China. Tree Physiol. 2017, 37, 1546–1553. [Google Scholar] [CrossRef] [Green Version]
- Gruber, A.; Strobl, S.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol. 2010, 30, 490–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deslauriers, A.; Rossi, S.; Anfodillo, T. Dendrometer and intra-annual tree growth: What kind of information can be inferred? Dendrochronologia 2007, 25, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Herzog, K.M.; Häsler, R.; Thum, R. Diurnal changes in the radius of a subalpine Norway spruce stem: Their relation to the sap flow and their use to estimate transpiration. Trees 1995, 10, 94–101. [Google Scholar] [CrossRef]
- Zweifel, R.; Item, H.; Häsler, R. Stem radius changes and their relation to stored water in stems of young Norway spruce trees. Trees 2000, 15, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Güney, A.; Gülsoy, S.; Şentürk, Ö.; Niessner, A.; Küppers, M. Environmental control of daily stem radius increment in the montane conifer Cedrus libani. J. For. Res. 2020, 31, 1159–1171. [Google Scholar] [CrossRef] [Green Version]
- Oberhuber, W.; Gruber, A.; Kofler, W.; Swidrak, I. Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur. J. For. Res. 2014, 133, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Raffelsbauer, V.; Spannl, S.; Peña, K.; Pucha-Cofrep, D.; Steppe, K.; Bräuning, A. Tree Circumference Changes and Species-Specific Growth Recovery After Extreme Dry Events in a Montane Rainforest in Southern Ecuador. Front. Plant Sci. 2019, 10, 342. [Google Scholar] [CrossRef]
- Daudet, F.-A.; Améglio, T.; Cochard, H.; Archilla, O.; Lacointe, A. Experimental analysis of the role of water and carbon in tree stem diameter variations. J. Exp. Bot. 2005, 56, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Häsler, R. Frost-induced reversible shrinkage of bark of mature subalpine conifers. Agric. For. Meteorol. 2000, 102, 213–222. [Google Scholar] [CrossRef]
- Zweifel, R.; Häsler, R. Dynamics of water storage in mature subalpine Picea abies: Temporal and spatial patterns of change in stem radius. Tree Physiol. 2001, 21, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Seasonal and daily cycles of stem radial variation of Pinus pinaster in a drought-prone environment. Agric. For. Meteorol. 2013, 180, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Guerrero-Campo, J.; Gutierrez, E. Tree-Ring Growth and Structure of Pinus uncinata and Pinus sylvestris in the Central Spanish Pyrenees. Arct. Alp. Res. 1998, 30, 1. [Google Scholar] [CrossRef]
- Martinez del Castillo, E.; Longares, L.A.; Gričar, J.; Prislan, P.; Gil-Pelegrín, E.; Čufar, K.; de Luis, M. Living on the Edge: Contrasted Wood-Formation Dynamics in Fagus sylvatica and Pinus sylvestris under Mediterranean Conditions. Front. Plant Sci. 2016, 7, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Luis, M.; Novak, K.; Raventós, J.; Gričar, J.; Prislan, P.; Čufar, K. Cambial activity, wood formation and sapling survival of Pinus halepensis exposed to different irrigation regimes. For. Ecol. Manag. 2011, 262, 1630–1638. [Google Scholar] [CrossRef]
- Güney, A.; Küppers, M.; Rathgeber, C.; Şahin, M.; Zimmermann, R. Intra-annual stem growth dynamics of Lebanon cedar along climatic gradients. Trees 2017, 31, 587–606. [Google Scholar] [CrossRef] [Green Version]
- Knerr, I.; Trachte, K.; Garel, E.; Huneau, F.; Santoni, S.; Bendix, J. Partitioning of Large-Scale and Local-Scale Precipitation Events by Means of Spatio-Temporal Precipitation Regimes on Corsica. Atmosphere 2020, 11, 417. [Google Scholar] [CrossRef] [Green Version]
- MétéoFrance. Bulletins Climatiques de France Métropolitaine et Outremer. 2020. Available online: https://donneespubliques.meteofrance.fr/?fond=produit&id_produit=129&id_rubrique=29 (accessed on 6 July 2020).
- ONF. Contribution à la Conduite des Peuplements de Pin Laricio et Habitats Associés: Patrimoine et Richesses; Office National des Fôrets: Sarreguemines, France, 2006. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Aryal, S.; Häusser, M.; Grießinger, J.; Fan, Z.; Bräuning, A. “dendRoAnalyst”: A tool for processing and analysing dendrometer data. Dendrochronologia 2020, 2020, 125772. [Google Scholar] [CrossRef]
- Zweifel, R.; Haeni, M.; Buchmann, N.; Eugster, W. Are trees able to grow in periods of stem shrinkage? New Phytol. 2016, 211, 839–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MétéoFrance. Climate Data from Stations Ajaccio, Bocognano, Evisa, Sampolo and Solenzara. 2020. Available online: https://www.meteofrance.fr (accessed on 20 May 2020).
- Schultz, J.A.; Beck, C.; Menz, G.; Neuwirth, B.; Ohlwein, C.; Philipp, A. Sensitivity of proxies on non-linear interactions in the climate system. Sci. Rep. 2015, 5, 18560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hersbach, H.; Bell, B.; Berrisford, P.; Hirahara, S.; Horányi, A.; Muñoz-Sabater, J.; Nicolas, J.; Peubey, C.; Radu, R.; Schepers, D.; et al. The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- Oberhuber, W.; Kofler, W.; Schuster, R.; Wieser, G. Environmental effects on stem water deficit in co-occurring conifers exposed to soil dryness. Int. J. Biometeorol. 2015, 59, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, C.; Rötzer, T.; Thurm, E.A.; Biber, P.; Kallenbach, C.; Pretzsch, H. Growth and Tree Water Deficit of Mixed Norway Spruce and European Beech at Different Heights in a Tree and under Heavy Drought. Forests 2019, 10, 557. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Zimmermann, L.; Zeugin, F.; Newbery, D.M. Intra-annual radial growth and water relations of trees: Implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loris, K.; Havranek, W.M.; Wieser, G. Ecological significance of thickness changes in stem, branches and twigs of Pinus cembra L. during winter. PHYTON-HORN- 1999, 39, 117–122. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
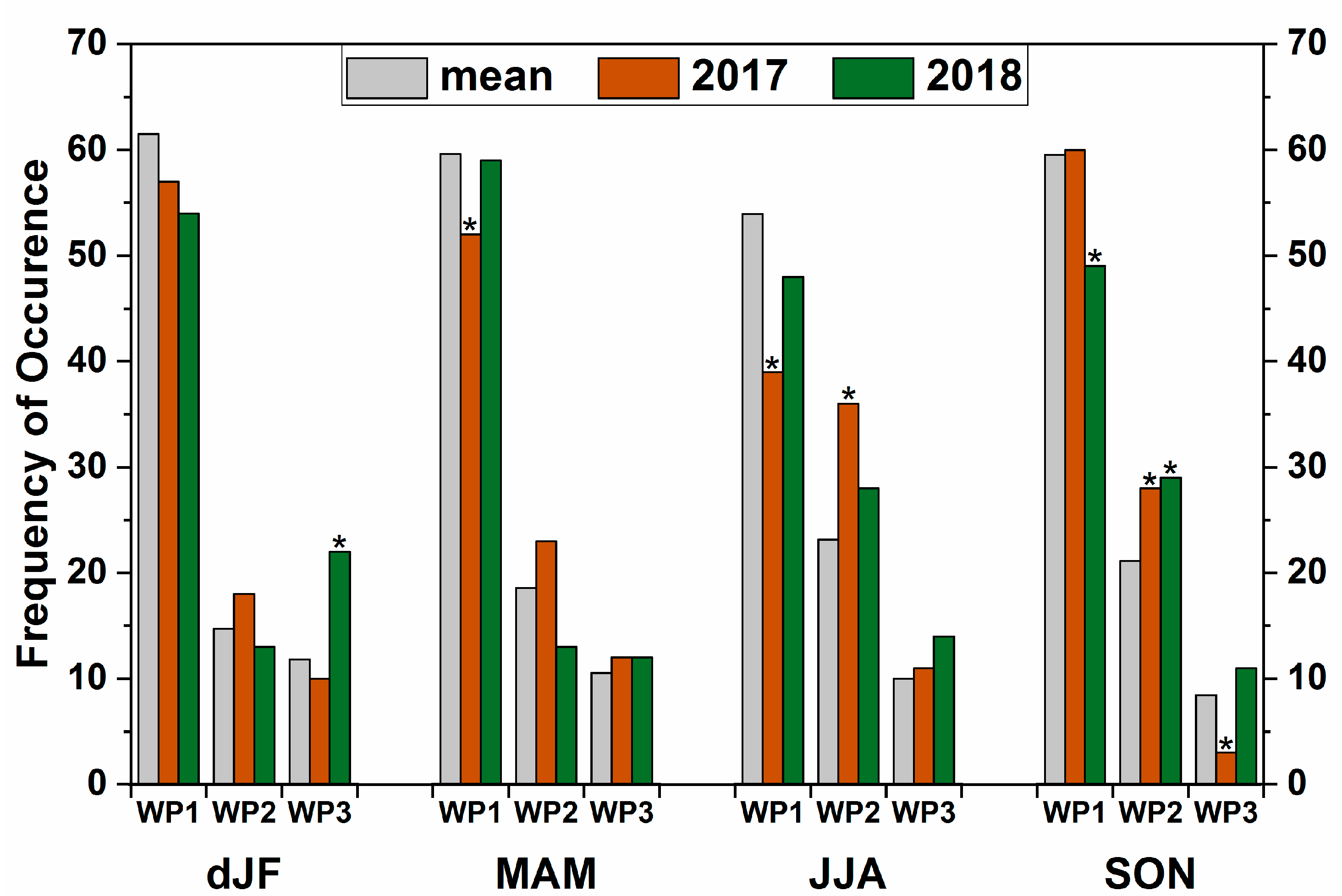{kind=link}
{kind=link}
{kind=link}
| Site | Cumulative GRO [mm/Year] | Maximum TWD [mm/Day] | ||||
|---|---|---|---|---|---|---|
| 2017 | 2018 | Δ | 2017 | 2018 | Δ | |
| WL (15 m asl) | 0.92 | 1.56 | +70% | 0.34 at DOY 247 | 0.39 at DOY 277 | +14% |
| WM (790 m asl) | 1.08 | 2.21 | +104% | 0.45 at DOY 243 | 0.21 at DOY 098 | −53% |
| EH (1600 m asl) | 1.64 | 2.28 | +39% | 0.32 at DOY 243 | 0.31 at DOY 117 | −3% |
| EM (1000 m asl) | 2.59 | 4.57 | +76% | 0.29 at DOY 241 | 0.22 at DOY 364 | −24% |
| EL (10 m asl) | 1.54 | 2.85 | +85% | 0.12 at DOY 302 | 0.14 at DOY 297 | +16% |
| Site | Mean Temperature [°C] | Rainy Days [n] | Precipitation Sum [mm] | Mean Soil Water Content [m3/m3] | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | ∆ | 2017 | 2018 | ∆ | 2017 | 2018 | ∆ | 2017 | 2018 | ∆ | |
| WL (15 m asl) | 17.0 | 17.4 | +0.4 | 77 | 133 | +73% | 319.4 | 609.3 | +91% | 0.03 | 0.04 | +33% |
| WM (790 m asl) | 13.3 | 12.7 | −0.6 | 112 | 217 | +94% | 1193.4 | 2208.4 | +85% | 0.08 | 0.19 | +138% |
| EH (1600 m asl) | 7.7 | 7.1 | −0.6 | 135 | 251 | +86% | 1594.3 | 2918.4 | +83% | NA | NA | NA |
| EM (1000 m asl) | 11.8 | 10.9 | −0.9 | 112 | 182 | +63% | 990.8 | 1695.4 | +71% | 0.04 | 0.12 | +200% |
| EL (10 m asl) | 16.0 | 15.9 | −0.1 | 93 | 140 | +51% | 721.5 | 709.5 | −2% | 0.07 | 0.18 | +157% |
| Site | A: Percentage of Rainy Days by Weather Pattern [%] | B: Percentage of Precipitation Amount by Weather Pattern [%] | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | |||||||||
| WP1 | WP2 | WP3 | WP1 | WP2 | WP3 | WP1 | WP2 | WP3 | WP1 | WP2 | WP3 | |
| WL (15 m asl) | 55 | 25 | 18 | 61 | 24 | 14 | 46 | 36 | 17 | 70 | 19 | 11 |
| WM (790 m asl) | 52 | 29 | 14 | 59 | 22 | 17 | 47 | 28 | 25 | 65 | 17 | 17 |
| EH (1600 m asl) | 53 | 31 | 12 | 55 | 25 | 17 | 59 | 21 | 19 | 58 | 20 | 21 |
| EM (1000 m asl) | 52 | 29 | 14 | 56 | 23 | 18 | 42 | 24 | 32 | 62 | 17 | 20 |
| EL (10 m asl) | 52 | 37 | 10 | 66 | 20 | 12 | 22 | 39 | 32 | 55 | 31 | 13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Häusser, M.; Szymczak, S.; Knerr, I.; Bendix, J.; Garel, E.; Huneau, F.; Trachte, K.; Santoni, S.; Bräuning, A. The Dry and the Wet Case: Tree Growth Response in Climatologically Contrasting Years on the Island of Corsica. Forests 2021, 12, 1175. https://doi.org/10.3390/f12091175
Häusser M, Szymczak S, Knerr I, Bendix J, Garel E, Huneau F, Trachte K, Santoni S, Bräuning A. The Dry and the Wet Case: Tree Growth Response in Climatologically Contrasting Years on the Island of Corsica. Forests. 2021; 12(9):1175. https://doi.org/10.3390/f12091175
Chicago/Turabian StyleHäusser, Martin, Sonja Szymczak, Isabel Knerr, Jörg Bendix, Emilie Garel, Frédéric Huneau, Katja Trachte, Sébastien Santoni, and Achim Bräuning. 2021. "The Dry and the Wet Case: Tree Growth Response in Climatologically Contrasting Years on the Island of Corsica" Forests 12, no. 9: 1175. https://doi.org/10.3390/f12091175