How Do Mediterranean Pine Trees Respond to Drought and Precipitation Events along an Elevation Gradient?

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
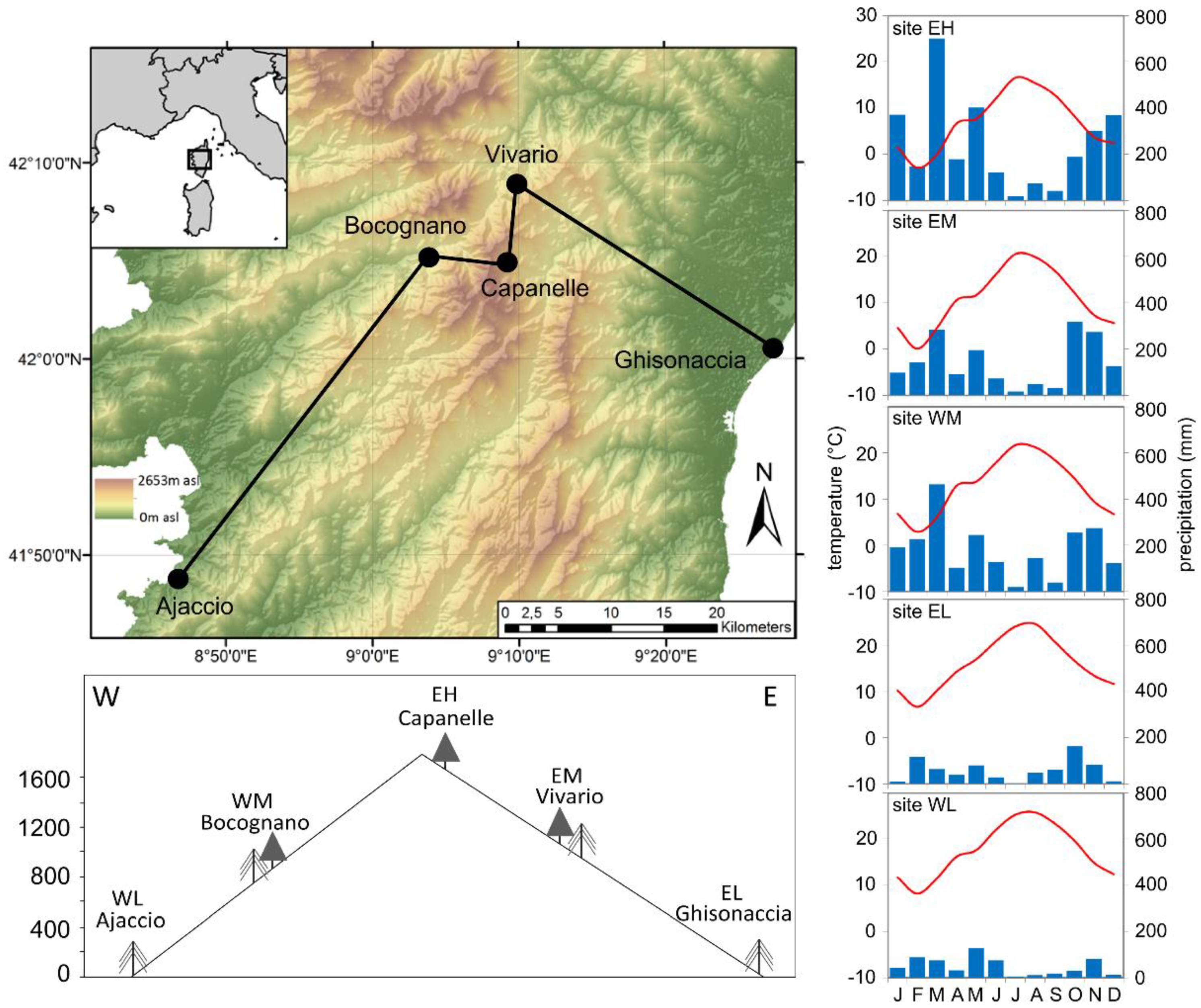
2.1. Study Sites
2.2. Meteorological Data
2.3. Dendrometer Data
3. Results and Discussion
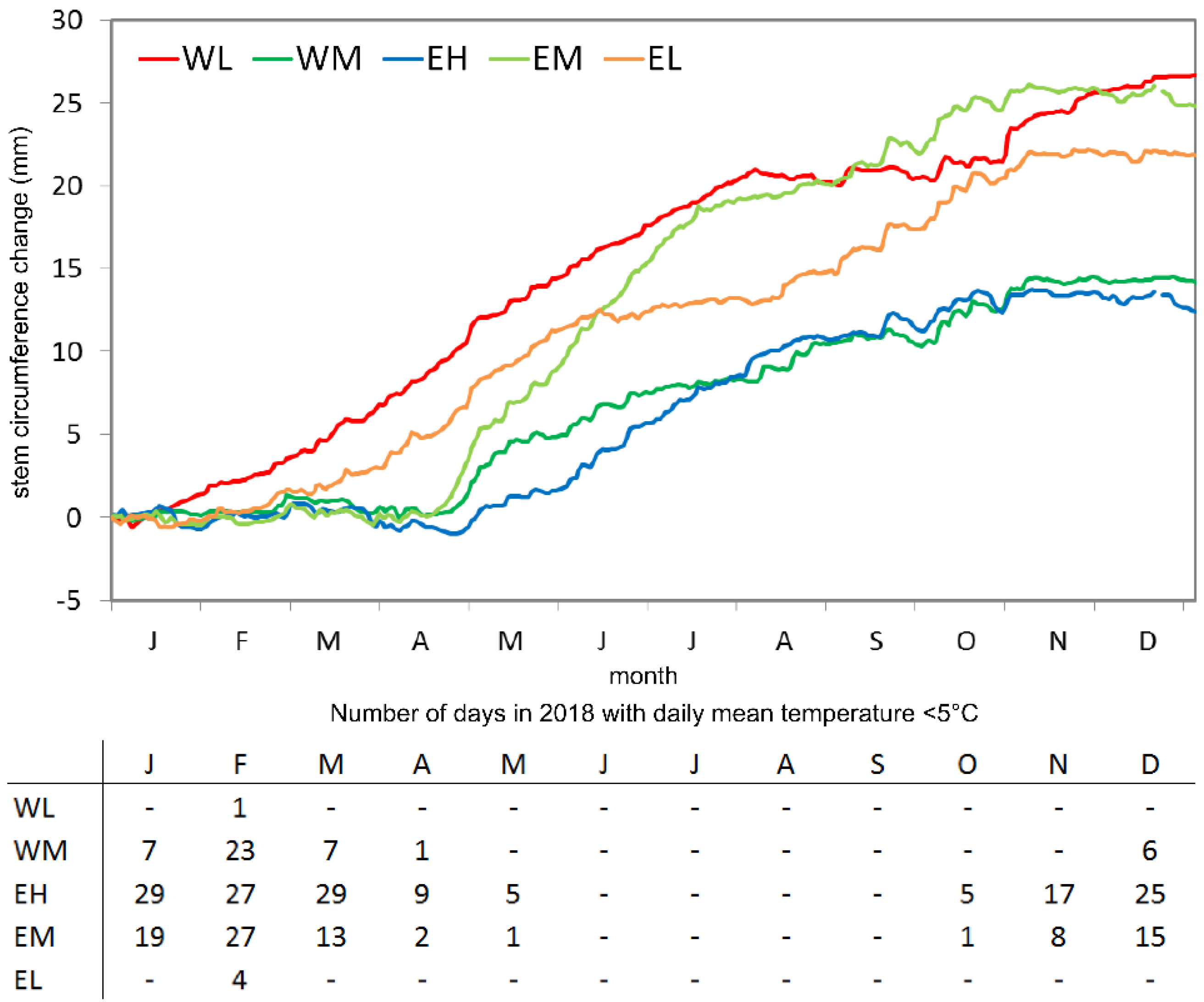
3.1. Site-Specific Stem Circumference Changes and Growing Periods
3.2. Precipitation Events and Dry Periods
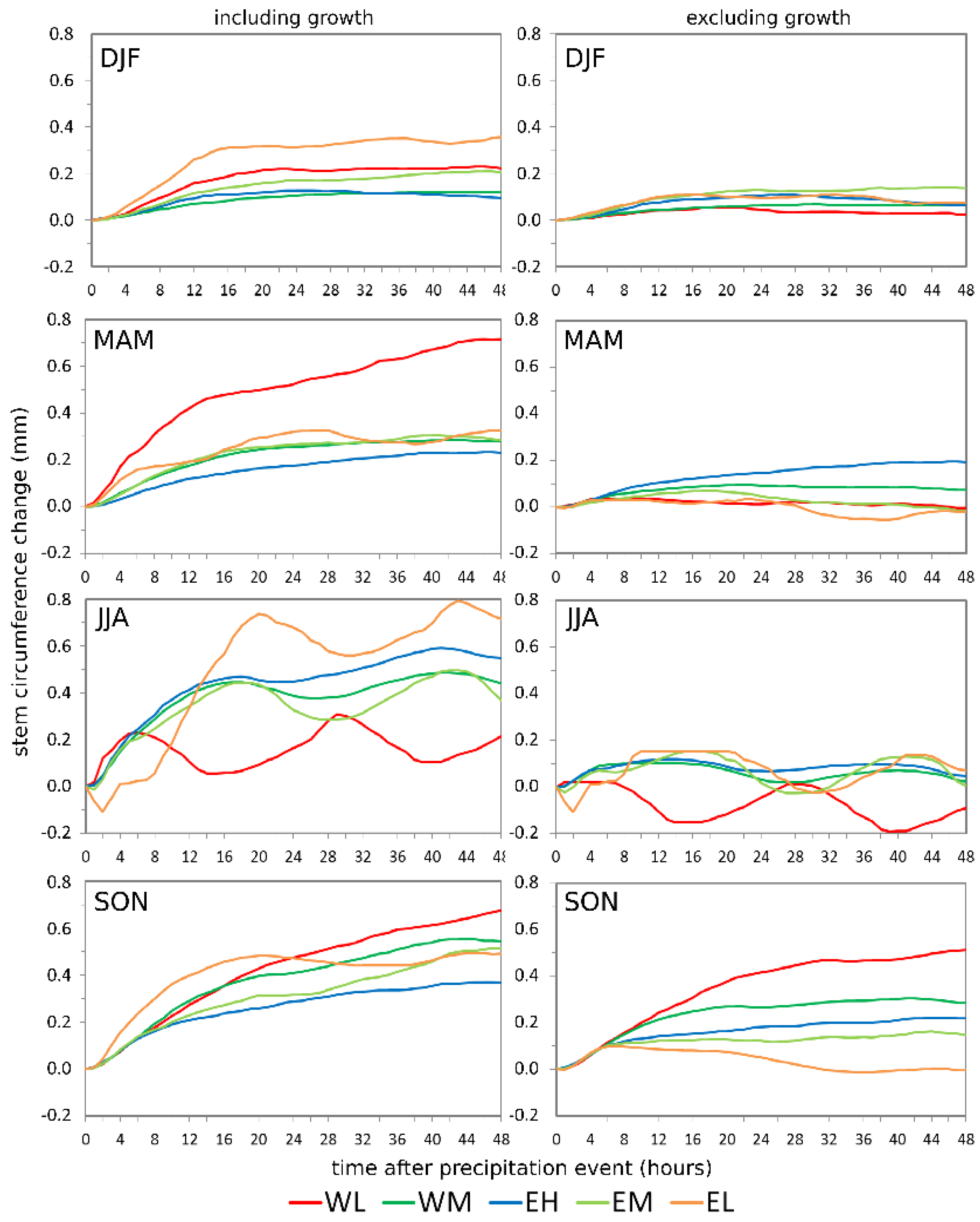
3.3. Tree Response to Precipitation Events
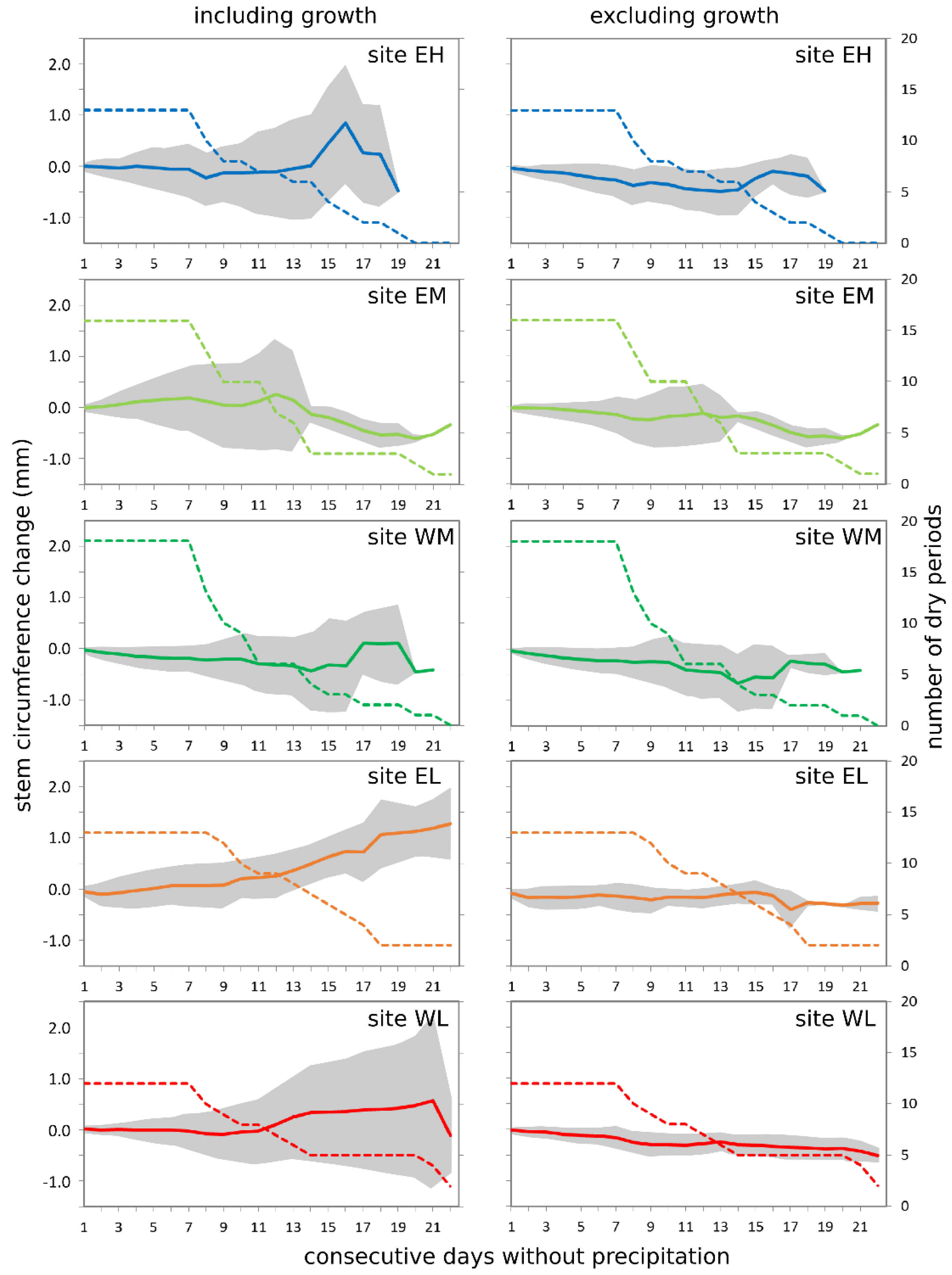
3.4. Tree Response to Dry Periods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meinzer, F.C.; Brooks, J.R.; Gartner, B.L.; Warren, J.M.; Wodruff, D.R.; Bible, K.; Shaw, D.C. Dynamics of water transport and storage in conifers studied with deuterium and heat tracing techniques. Plant Cell Environ. 2006, 29, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battipaglia, G.; DeMicco, V.; Brand, W.A.; Saurer, M.; Aronne, G.; Linke, P.; Cherubini, P. Drought impact on water use efficiency and intra-annual density fluctuations in Erica arborea on Elba (Italy). Plant Cell Environ. 2013, 37, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Flint, A.; Huang, C.-Y.; Flint, L.; Berry, J.A.; Davis, F.W.; Sperry, J.S.; Field, C.B. Tree mortality predicted from drought-induced vascular damage. Nat. Geosci. 2015. [Google Scholar] [CrossRef] [Green Version]
- Hember, R.A.; Kurz, W.A.; Coops, N.C. Relationships between the individual-tree mortality and water-balance variables indicate positive trends in water stress-induced tree mortality across North America. Glob. Chang. Biol. 2017, 23, 1691–1710. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Schäfer, C.; Rötzer, T.; Thurm, E.A.; Biber, P.; Kallenbach, C.; Pretzsch, H. Growth and tree water deficit of mixed Norway spruce and European beech at different heights in a tree and under heavy drought. Forests 2019, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Steudle, E. Water uptake by roots: Effects of water deficit. J. Exp. Bot. 2000, 51, 1531–1542. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.; Moore, L.; Hoffmann, W.; Pockman, W.; Linder, C. Ecosystem rooting depth determined with caves and DNA. Proc. Natl. Acad. Sci. USA 1999, 96, 11387–11392. [Google Scholar] [CrossRef] [Green Version]
- Ogle, K.; Reynolds, J.F. Plant responses to precipitation in desert ecosystems: Integrating functional types, pulses, thresholds, and delays. Oecologia 2004, 141, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Eggemeyer, K.D.; Awada, T.; Harvey, F.E.; Wedin, D.A.; Zhou, X.; Zanner, C.W. Seasonal changes in depth of water uptake for encroaching trees Juniperus virginiana and Pinus ponderosa and two dominant C4 grasses in a semiarid grassland. Tree Physiol. 2009, 29, 157–169. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Virginia, R.A.; Kemp, P.R.; de Soyza, A.G.; Tremmel, D.C. Impact of drought on desert shrubs: Effects of seasonality and degree of resource island development. Ecol. Monogr. 1999, 69, 69–106. [Google Scholar] [CrossRef]
- Gebauer, R.L.W.; Ehleringer, J.R. Water and nitrogen uptake patterns following moisture pulses in a cold desert community. Ecology 2000, 81, 1415–1424. [Google Scholar] [CrossRef]
- Zweifel, R.; Zimmermann, L.; Newbery, D.M. Modeling tree water deficit from microclimate: An approach to quantifying drought stress. Tree Physiol. 2005, 25, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mencuccini, M.; Hölttä, T.; Sevanto, S.; Nikinmaa, E. Concurrent measurements of change in the bark and xylem diameters of trees reveal a phloem-generated turgor signal. New Phytol. 2013, 198, 1143–1154. [Google Scholar] [CrossRef]
- Deslauriers, A.; Rossi, S.; Anfodillo, T. Dendrometer and intra-annual tree growth: What kind of information can be inferred? Dendrochronologia 2007, 25, 113–124. [Google Scholar] [CrossRef] [Green Version]
- King, G.; Fonti, P.; Nievergelt, D.; Büntgen, U.; Frank, D. Climatic drivers of hourly to yearly tree radius variations along a 6 °C natural warming gradient. Agric. Meteorol. 2013, 168, 36–46. [Google Scholar] [CrossRef]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Sesaonal and daily cycles of stem radial variation of Pinus pinaster in a drought-prone environment. Agric. Meteorol. 2013, 180, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Siegmund, J.F.; Sanders, T.G.M.; Heinrich, I.; van der Maaten, E.; Simard, S.; Helle, G.; Donner, R.V. Meteorological drivers of extremes in daily stem radius variations of beech, oak, and pine in northeastern Germany: An event coincidence analysis. Front. Plant Sci. 2016, 7, 733. [Google Scholar] [CrossRef] [Green Version]
- Raffelsbauer, V.; Spannl, S.; Pena, K.; Pucha-Cofrep, D.; Steppe, K.; Bräuning, A. Tree circumference changes and species-specific growth recovery after extreme dry events in a montane rainforest in southern Ecuador. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Zweifel, R. Radial stem variations—A source of tree physiological information not fully exploited yet. Plant Cell Environ. 2016, 39, 231–232. [Google Scholar] [CrossRef]
- Oberhuber, W.; Gruber, A.; Kofler, W.; Swirdak, I. Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur. J. Res. 2014, 133, 467–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güney, A.; Gülsoy, S.; Sentürk, Ö.; Niessner, A.; Küppers, M. Environmental control of daily stem radius increment in the montane conifer Cedrus libani. J. Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Chan, T.; Hölttä, T.; Berninger, F.; Mäkinen, H.; Nöjd, P.; Mencuccini, M.; Nikinmaa, E. Separating water-potential induced swelling and shrinking from measured radial stem variations reveals a cambial growth and osmotic concentration signal. Plant Cell Environ. 2016, 39, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Mencuccini, M.; Salmon, Y.; Mitchell, P.; Hölttä, T.; Choat, B.; Meir, P.; O’Grady, A.; Tissue, D.; Zweifel, R.; Sevanto, S.; et al. An empirical method that separates irreversible stem radial growth from bark water content changes in trees: Theory and case studies. Plant Cell Environ. 2017, 40, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Zweifel, R.; Häsler, R. Dynamics of water storage in mature subalpine Picea abies: Temporal and spatial patterns of change in stem radius. Tree Physiol. 2001, 21, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Barnard, R.L.; Salmon, Y.; Kodama, N.; Sörgel, K.; Holst, J.; Rennenberg, H.; Gessler, A.; Buchmann, N. Evaporative enrichment and time lags between δ18O of leaf water and organic pools in a pine stand. Plant Cell Environ. 2007, 30, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Brandes, E.; Wenninger, J.; Koeninger, P.; Schindler, D.; Rennenberg, H.; Leibundgut, C.; Mayer, H.; Gessler, A. Assessing environmental and physiological controls over water relations in a Scots pine (Pinus sylvestris L.) stand through analyses of stable isotope composition of water and organic matter. Plant Cell Environ. 2007, 30, 113–127. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.L.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.H.; Zhao, X.; Liang, S.L.; Zhou, T.; Huang, K.C.; Tang, B.J.; Zhao, W.Q. Time-lag effects of global vegetation responses to climate change. Glob. Chang. Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef]
- Bruno, C.; Dupre, G.; Giorgetti, G.; Borne, D. Chì Tempu Face? Météorologie, Climat et Microclimats de la Corse; CNDP-CRDP de Corse/MétéoFrance: Ajaccio, France, 2001. [Google Scholar]
- Häusser, M.; Szymczak, S.; Garel, E.; Santoni, S.; Huneau, F.; Bräuning, A. Growth variability of two native pine species on Corsica as a function of altitude. Dendrochronologia 2019, 54, 49–55. [Google Scholar] [CrossRef]
- Shuttle Radar Topography Mission of the United States Geological Survey. Available online: https://lta.cr.usgs.gov/SRTM/ (accessed on 15 January 2018).
- MétéoFrance. Climate Data from Stations Ajaccio, Bocognano, Evisa, Sampolo and Solenzara. Available online: https://www.meteofrance.fr (accessed on 20 May 2020).
- Cheng, X.L.; An, S.Q.; Li, B.; Chen, J.Q.; Lin, G.H.; Liu, Y.H.; Luo, Y.; Liu, S. Summer rain pulse size and rainwater uptake by three dominant desert plants in a desertified grassland ecosystem in northwestern China. Plant Ecol. 2006, 184, 1–12. [Google Scholar] [CrossRef]
- Aryal, S.; Häusser, M.; Grießinger, J.; Fan, Z.-X.; Bräuning, A. DendRoAnalyst: A Complete Tool for Processing and Analyzing Dendrometer Data. R Package Version 0.1.0. 2020. Available online: https://CRAN.R-project.org/package=dendRoAnalyst (accessed on 7 May 2020).
- Thabeet, A.; Vennetier, M.; Gadbin-Henry, C.; Denelle, N.; Roux, M.; Caraglio, Y.; Vila, B. Response of Pinus sylvestris L. to recent climatic events in the French Mediterranean region. Trees 2009, 23, 843–853. [Google Scholar] [CrossRef] [Green Version]
- Hänninen, H.; Tannino, K. Tree seasonality in a warming climate. Trends Plant. Sci. 2011, 16, 412–416. [Google Scholar] [CrossRef]
- Rossi, S.; Morin, H.; Deslauriers, A.; Plourde, P.Y. Predicting xylem phenology in black spruce under climate warming. Glob. Chang. Biol. 2011, 17, 614–625. [Google Scholar] [CrossRef]
- Swidrak, I.; Gruber, A.; Kofler, W.; Oberhuber, W. Effects of environmental conditions on onset of xylem growth in Pinus sylvestris under drought. Tree Physiol. 2011, 31, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Gricar, J.; Seo, J.-W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Güney, A.; Kerr, D.; Sökücü, A.; Zimmermann, R.; Küppers, M. Cambial activity and xylogenesis in stems of Cedrus libani A. Rich at different altitudes. Bot. Stud. 2015. [Google Scholar] [CrossRef]
- Liu, X.; Nie, Y.; Wen, F. Seasonal dynamics of stem radial increment of Pinus taiwanensis Hayata and its response to environmental factors in the Lushan mountains, southeastern China. Forests 2018, 9, 387. [Google Scholar] [CrossRef] [Green Version]
- De Luis, M.; Gričar, J.; Čufar, K.; Raventós, J. Sesaonal dynamics of wood formation in Pinus halepensis from dry and semi-arid ecosystems in Spain. IAWA J. 2007, 28, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Cocozza, C.; Palombo, C.; Tognetti, R.; La Porta, N.; Anichini, M.; Giovannelli, A.; Emiliani, G. Monitoring intra-annual dynamics of wood formation with microcores and dendrometers in Picea abies at two different altitudes. Tree Physiol. 2016, 36, 832–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deslauriers, A.; Morin, H.; Begin, Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Can. J. For. Res. 2003, 33, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, A.; Morin, H.; Krause, C.; Deslauriers, A.; Thibeault-Martel, M. The timing of spring rehydration and its relation with the onset of wood formation in black spruce. Agric. Meteorol. 2009, 149, 1403–1409. [Google Scholar] [CrossRef]
- Klippel, L.; Hartl-Meier, C.; Lindén, J.; Kochbeck, M.; Emde, K.; Esper, J. Hourly resolved climate response of Picea abies beyond its natural distribution range. Balt. For. 2017, 23, 556–563. [Google Scholar]
- Čermak, J.; Kučerá, J.; Bauerle, W.L.; Phillips, N.; Hinckley, T.M. Tree water storage and its diurnal dynamics related to sap flow and changes in stem volume in old-growth Douglas-fir trees. Tree Physiol. 2007, 27, 181–198. [Google Scholar] [CrossRef]
- Holbrook, N.M. Stem water storage. In Plant Stems: Physiology and Functional Morphology; Gartner, B.L., Ed.; Academic Press: San Diego, CA, USA, 1995; pp. 151–175. [Google Scholar]
- Grip, H.; Hällgren, J.E. Water cycling in coniferous forest ecosystems. In Ecosystems of the World. Volume 6. Coniferous Forests; Andersson, F., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 385–426. [Google Scholar]
- Szymczak, S.; Barth, J.; Bendix, J.; Huneau, F.; Garel, E.; Häusser, M.; Juhlke, T.; Knerr, I.; Santoni, S.; Trachte, K.; et al. Tracking the oxygen isotope composition from source to sink in pine trees along an elevation gradient in a Mediterranean ecosystem. Chem. Geol. 2020. under review. [Google Scholar] [CrossRef]
- Schütt, P.; Weisgerber, H.; Schuck, H.J.; Lang, U.; Stimm, B.; Roloff, A. Lexikon der Nadelbäume; Nikol: Hamburg, Germany, 2004; p. 639. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Trees | Tree Species | Mean Age (yr) ± 1 STD | Mean Stem Circumference (cm) ± 1 STD | |
|---|---|---|---|---|
| WL | 6 | PIPI | 32 ± 10 | 131 ± 24 |
| WM | 12 | PIPI, PINI | 42 ± 7 | 116 ± 24 |
| EH | 6 | PINI | 59 ± 10 | 109 ± 15 |
| EM | 12 | PIPI, PINI | 53 ± 10 | 120 ± 30 |
| EL | 6 | PIPI | 47 ± 20 | 139 ± 19 |
| Precipitation Events | ||||||
|---|---|---|---|---|---|---|
| WL | WM | EH | EM | EL | ||
| mean precipitation sum per event (mm) | 11.5 | 20.8 | 33.0 | 18.6 | 14.3 | |
| mean duration of precipitation events (h) | 6 | 9 | 14 | 10 | 6 | |
| mean intensity per event (mm/h) | 2.6 | 2.7 | 2.9 | 2.3 | 3.3 | |
| number of events per season | DJF | 6 | 42 | 41 | 31 | 7 |
| MAM | 8 | 43 | 48 | 29 | 5 | |
| JJA | 1 | 19 | 13 | 12 | 1 | |
| SON | 10 | 34 | 24 | 26 | 24 | |
| number of events per amount class | 5–9.9 mm | 14 | 47 | 38 | 38 | 14 |
| 10–19.9 mm | 7 | 45 | 26 | 37 | 14 | |
| 20–29.9 mm | 4 | 17 | 24 | 9 | 7 | |
| 30–39.9 mm | - | 13 | 10 | 5 | 1 | |
| 40–49.9 mm | - | 5 | 3 | 1 | 1 | |
| 50–99.9 mm | - | 10 | 16 | 7 | - | |
| > 100 mm | - | - | 9 | 1 | - | |
| total number of precipitation events | 25 | 138 | 126 | 98 | 37 | |
| Dry periods with at least 7 days without precipitation | ||||||
| WL | WM | EH | EM | EL | ||
| maximum duration (days) | 62 | 21 | 19 | 33 | 27 | |
| mean duration (days) | 18 | 10 | 11 | 13 | 14 | |
| number of dry periods per season | DJF | 3 | 2 | 3 | 3 | 1 |
| MAM | 3 | 5 | 3 | 4 | 2 | |
| JJA | 3 | 7 | 6 | 7 | 8 | |
| SON | 7 | 6 | 4 | 5 | 3 | |
| total number of dry periods | 16 | 20 | 16 | 19 | 14 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szymczak, S.; Häusser, M.; Garel, E.; Santoni, S.; Huneau, F.; Knerr, I.; Trachte, K.; Bendix, J.; Bräuning, A. How Do Mediterranean Pine Trees Respond to Drought and Precipitation Events along an Elevation Gradient? Forests 2020, 11, 758. https://doi.org/10.3390/f11070758
Szymczak S, Häusser M, Garel E, Santoni S, Huneau F, Knerr I, Trachte K, Bendix J, Bräuning A. How Do Mediterranean Pine Trees Respond to Drought and Precipitation Events along an Elevation Gradient? Forests. 2020; 11(7):758. https://doi.org/10.3390/f11070758
Chicago/Turabian StyleSzymczak, Sonja, Martin Häusser, Emilie Garel, Sébastien Santoni, Frédéric Huneau, Isabel Knerr, Katja Trachte, Jörg Bendix, and Achim Bräuning. 2020. "How Do Mediterranean Pine Trees Respond to Drought and Precipitation Events along an Elevation Gradient?" Forests 11, no. 7: 758. https://doi.org/10.3390/f11070758