Enhanced Light-Induced Biosynthesis of Fatty Acids Suitable for Biodiesel Production by the Yellow-Green Alga Eustigmatos magnus



Abstract
:1. Introduction
2. Results
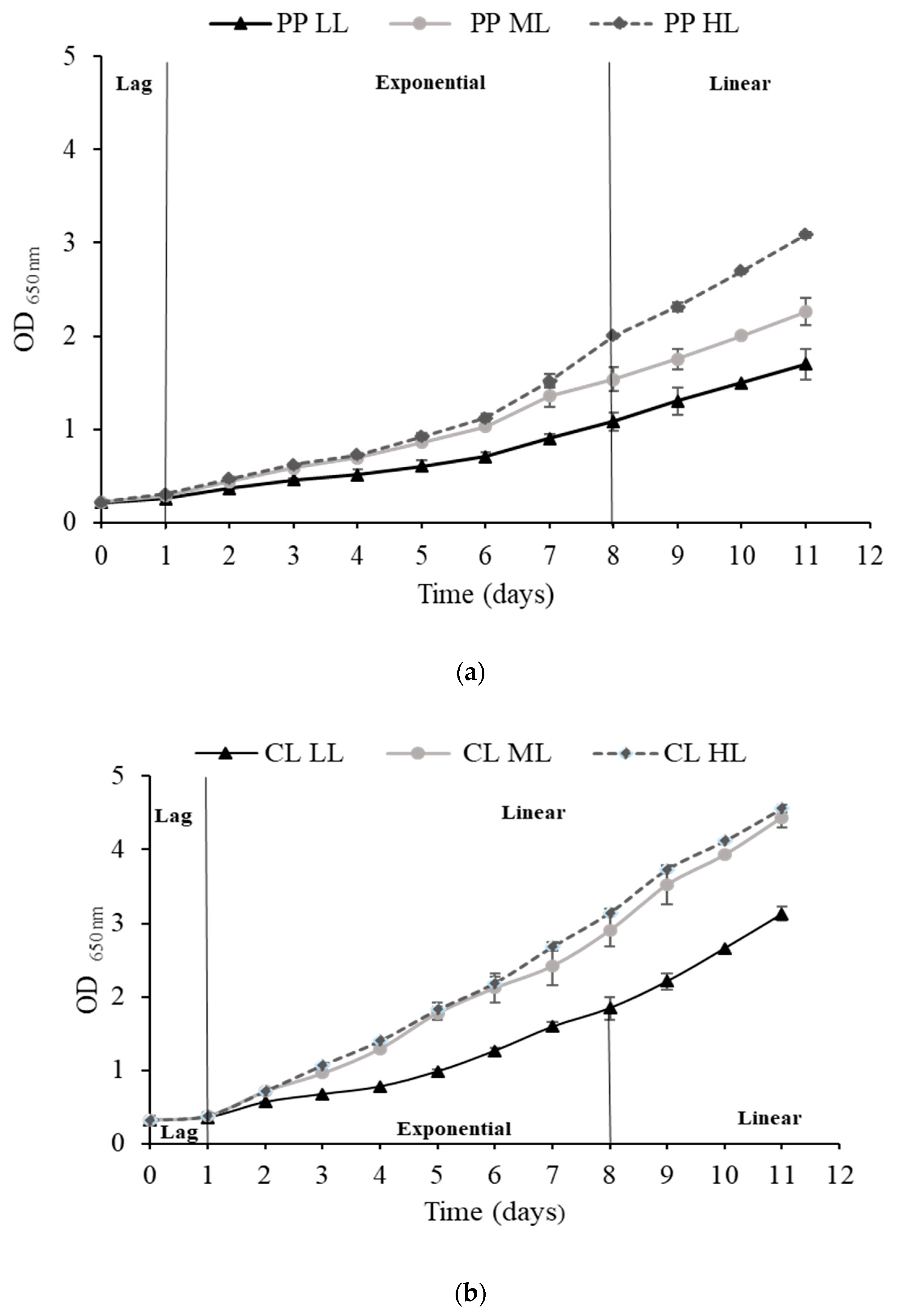
2.1. Analysis of Eustigmatos magnus Growth
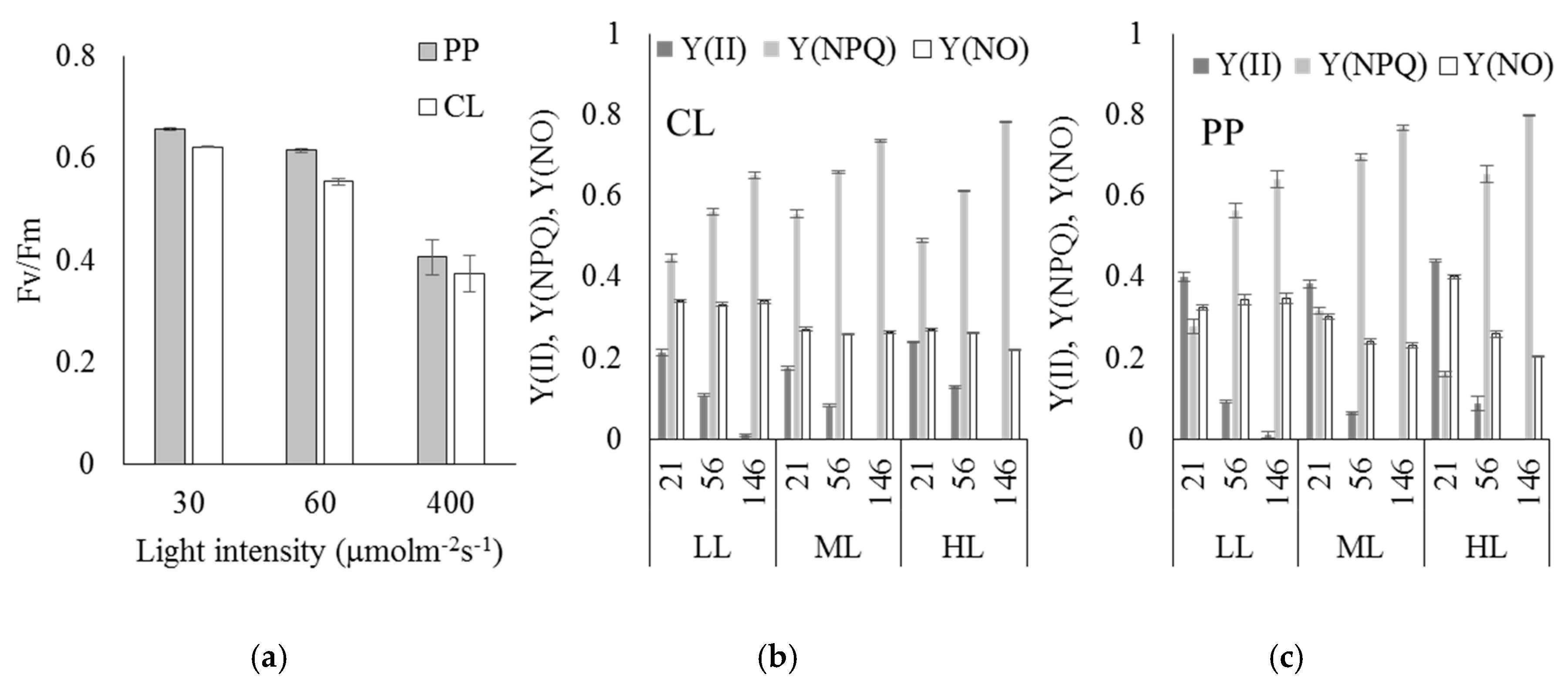
2.2. Content of Pigments and Analysis of Chlorophyll Fluorescence
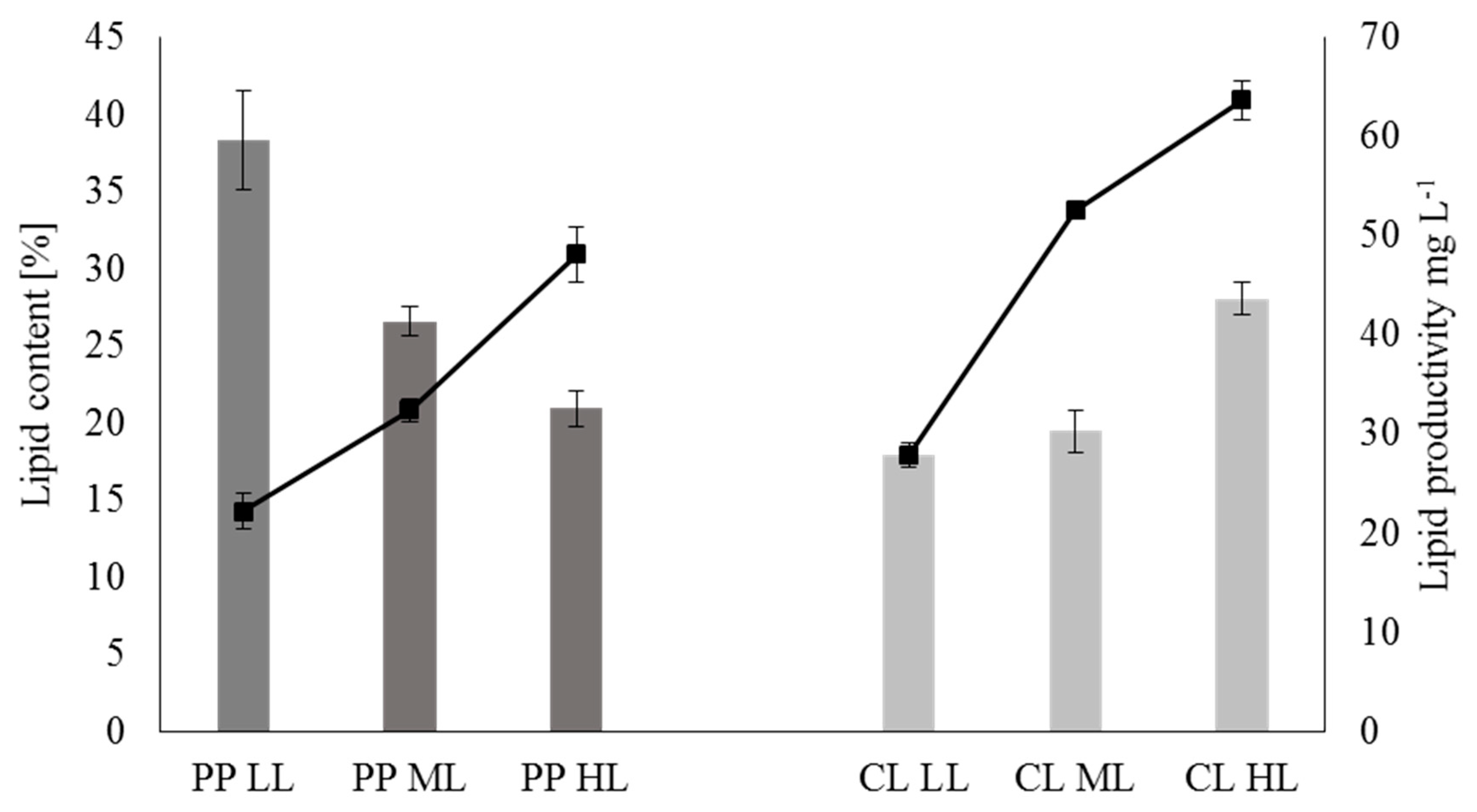
2.3. Analysis of Lipid Metabolism
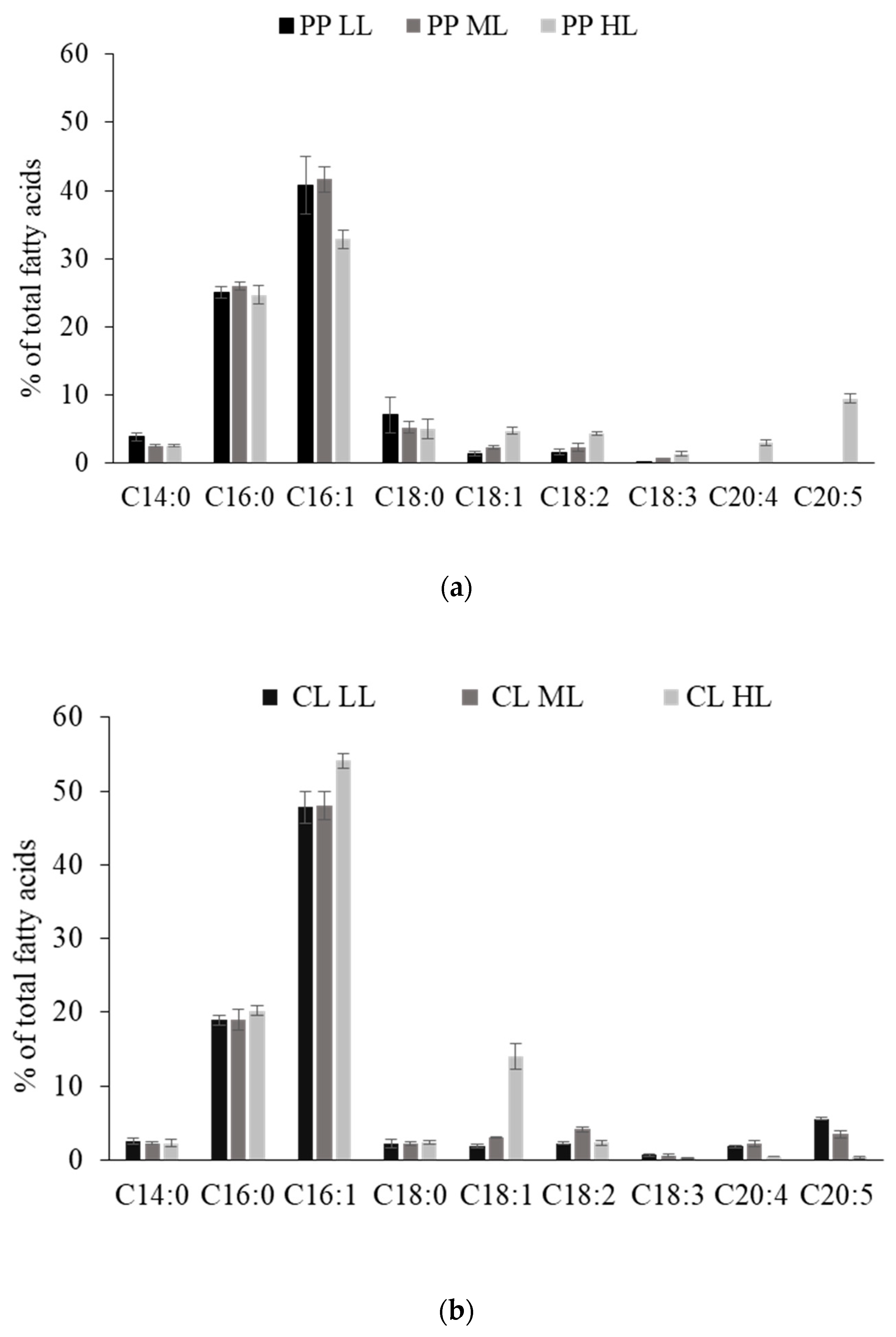
2.4. Analysis of the Fatty Acid Profile
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Microalgal Strain and Culture Conditions
5.2. Growth Density Measurement
5.3. Biomass Determination
5.4. Pigment Analysis
5.5. Lipid Extraction
5.6. Determination of the Fatty Acid Composition
5.7. Determination of the Effective Quantum Yield of PSII Photochemistry
Author Contributions
Funding
Conflicts of Interest
References
- Magierek, E.; Krzemińska, I. Effect of stress conditions on improvement of lipid and carbohydrate accumulation under photoautotrophic cultivation of Chlorophyta. Phycologia 2018, 57, 601–618. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Khozin-Goldberg, I.; Solovchenko, A.; Pal, D.; Cohen, Z.; Boussiba, S. Omega-3 and omega-6 LC-PUFA from photosynthetic microalgae: Studies on Parietochlorisincisa and Nannochloropsis sp. In Polyunsaturated Fatty Acids: Sources, Antioxidant Properties and Health Benefits; Catala, A., Ed.; Nova Science Publishers: New York, NY, USA, 2006. [Google Scholar]
- Tamburic, B.; Guruprasad, S.; Radford, D.T.; Szabó, M.; McC Lilley, R.; Larkum, A.W.D.; Franklin, J.B.; Kramer, D.M.; Blackburn, S.I.; Raven, J.A.; et al. The effect of diel temperature and light cycles on the growth of Nannochloropsis oculata in a photobioreactor matrix. PLoS ONE 2014, 9, e86047. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Ma, X.; Li, A.; Zhang, C. A novel potential source of β-carotene: Eustigmatos cf. polyphem (Eustigmatophyceae) and pilot β-carotene production in bubble column and flat panel photobioreactors. Bioresour. Technol. 2012, 117, 257–263. [Google Scholar] [CrossRef]
- Gruber-Brunhumer, M.R.; Schöberl, A.; Zohar, E.; Koenigsberger, S.; Bochmann, G.; Uher, B.; Lang, I.; Schagerl, M.; Fuchs, W.; Drosg, B. Cultivation of the microalga Eustigmatos magnus in different photobioreactor geometries and subsequent anaerobic digestion of pre-treated biomass. Biomass Bioenergy 2019, 128, 105303. [Google Scholar] [CrossRef]
- Krzemińska, I.; Pawlik-Skowrońska, B.; Trzcińska, M.; Tys, J. Influence of photoperiods on the growth rate and biomass productivity of green microalgae. Bioprocess Biosyst. Eng. 2014, 37, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Sirisuk, P.; Ra, C.-H.; Jeong, G.-T.; Kim, S.-K. Effects of wavelength mixing ratio and photoperiod on microalgal biomass and lipid production in a two-phase culture system using LED illumination. Bioresour. Technol. 2018, 253, 1715–1781. [Google Scholar] [CrossRef]
- Meseck, S.L.; Alix, J.H.; Wikfors, G.H. Photoperiod and light intensity effects on growth and utilization of nutrients by the aquaculture feed microalga, Tetraselmischui (PLY429). Aquaculture 2005, 246, 393–404. [Google Scholar] [CrossRef]
- Mitra, M.; Patidar, S.K.; George, B.; Shah, F.; Mishra, S. A euryhaline Nannochloropsis gaditana with potential for nutraceutical (EPA) and biodiesel production. Algal Res. 2015, 8, 161–167. [Google Scholar] [CrossRef]
- Matos, Â.P.; Cavanholi, M.G.; Moecke, E.H.S.; Sant’Anna, E.S. Effects of different photoperiod and trophic conditions on biomass, protein and lipid production by the marine alga Nannochloropsis gaditana at optimal concentration of desalination concentrate. Bioresour. Technol. 2017, 224, 490–497. [Google Scholar] [CrossRef]
- Li, Z.; Sun, M.; Li, Q.; Li, A.; Zhang, C. Profiling of carotenoids in six microalgae (Eustigmatophyceae) and assessment of their β-carotene productions in bubble column photobioreactor. Biotechnol. Lett. 2012, 34, 2049–2053. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Huang, L.; Gao, B.; Zhang, C. Optimum production conditions, purification, identification, and antioxidant activity of violaxanthin from microalga Eustigmatos cf. polyphem (Eustigmatophyceae). Mar. Drugs 2018, 16, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedzwiedzki, D.M.; Wolf, B.M.; Blankenship, R.E. Excitation energy transfer in the far-red absorbing violaxanthin/vaucheriaxanthin chlorophyll a complex from the eustigmatophyte alga FP5. Photosynth. Res. 2019, 140, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Keşan, G.; Litvín, R.; Bína, D.; Durchan, M.; Šlouf, V.; Polívka, T. Efficient light-harvesting using non-carbonyl carotenoids: Energy transfer dynamics in the VCP complex from Nannochloropsis oceanica. Biochim. Et Biophys. Acta Bioenerg. 2016, 1857, 370–379. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.; Stoykova, P.; Uzunov, B.; Dincheva, I.; Atanassov, I.; Draganova, P.; Borisova, C.; Gärtner, G. Carotenoids in five aeroterrestrial strains from Vischeria/Eustigmatos group: Updating the pigment pattern of Eustigmatophyceae. Biotechnol. Biotechnol. Equip. 2019, 33, 250–267. [Google Scholar] [CrossRef] [Green Version]
- Lubián, L.M.; Montero, O. Excess light-induced violaxanthin cycle activity in Nannochloropsis gaditana (Eustigmatophyceae): Effects of exposure time and temperature. Phycologia 1998, 37, 16–23. [Google Scholar] [CrossRef]
- Larkum, A.W.D.; Grossmann, A.R.; Raven, J.A. Photosynthesis in Algae: Biochemical and Physiological Mechanisms. In Advances in Photosynthesis and Respiration Including Bioenergy and Related Processes; Larkum, A.W.D., Grossmann, A.R., Raven, J.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Wolf, B.M.; Niedzwiedzki, D.M.; Magdaong, N.C.M.; Roth, R.; Goodenough, U.; Blankenship, R.E. Characterization of a newly isolated freshwater Eustigmatophyte alga capable of utilizing far-red light as its sole light source. Photosynth. Res. 2018, 135, 177–189. [Google Scholar] [CrossRef]
- Litvín, R.; Bína, D.; Herbstová, M.; Gardian, Z. Architecture of the light-harvesting apparatus of the eustigmatophyte alga Nannochloropsis oceanica. Photosynth. Res. 2016, 130, 137–150. [Google Scholar] [CrossRef]
- Lubián, L.M.; Montero, O.; Moreno-Garrido, I.; Huertas, I.E.; Sobrino, C.; González-del Valle, M.; Parés, G. Nannochloropsis (Eustigmatophyceae) as source of commercially valuable pigments. J. Appl. Phycol. 2000, 12, 249–255. [Google Scholar] [CrossRef]
- Simionato, D.; Sforza, E.; Corteggiani Carpinelli, E.; Bertucco, A.; Giacometti, G.M.; Morosinotto, T. Acclimation of Nannochloropsis gaditana to different illumination regimes: Effects on lipids accumulation. Bioresour. Technol. 2011, 102, 6026–6032. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Harvey, P.J. The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina (chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 2016, 106, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Richmond, A. Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Blackwell Publishing Company: Oxford, UK, 2004; pp. 125–177. [Google Scholar]
- Huang, W.; Zhang, S.-B.; Cao, K.-F. Evidence for leaf fold to remedy the deficiency of physiological photoprotection for photosystem II. Photosynth. Res. 2012, 110, 185–191. [Google Scholar] [CrossRef]
- Perkins, R.; Williamson, C.; Lavaud, J.; Mouget, J.-L.; Campbell, D.A. Time-dependent upregulation of electron transport with concomitant induction of regulated excitation dissipation in Haslea diatoms. Photosynth. Res. 2018, 137, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Zhang, X.; Xu, D.; Fan, X.; Mou, S.; Wang, Y.; Ye, N.; Wang, W. A transthylakoid proton gradient and inhibitors induce a non-photochemical fluorescence quenching in unicellular algae Nannochloropsis sp. Febs Lett. 2013, 587, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Krzemińska, I.; Piasecka, A.; Nosalewicz, A.; Simionato, D.; Wawrzykowski, J. Alterations of the lipid content and fatty acid profile of Chlorella protothecoides under different light intensities. Bioresour. Technol. 2015, 196, 72–77. [Google Scholar] [CrossRef]
- Goold, H.D.; Cuiné, S.; Legeret, B.; Liang, Y.; Brugière, S.; Auroy, P.; Javot, H.; Tardif, M.; Jones, B.; Beisson, F.; et al. Saturating light induces sustained accumulation of oil in plastidal lipid droplets in Chlamydomonas reinhardtii. Plant Physiol. 2016, 171, 2406–2417. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.J.; Harrison, S.T.L. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Appl. Phycol. 2009, 21, 493–507. [Google Scholar] [CrossRef]
- Meng, Y.; Jiang, J.; Wang, H.; Cao, X.; Xue, S.; Yang, Q.; Wang, W. The characteristics of TAG and EPA accumulation in Nannochloropsis oceanica IMET1 under different nitrogen supply regimes. Bioresour. Technol. 2015, 179, 483–489. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef]
- Pinzi, S.; Garcia, I.L.; Lopez-Gimenez, F.J.; Luque de Castro, M.D.; Dorado, G.; Dorado, M.P. The ideal vegetable oil-based biodiesel composition: A review of social, economical and technical implications. Energy Fuels 2009, 23, 2325–2341. [Google Scholar] [CrossRef]
- Sharma, K.K.; Schuhmann, H.; Schenk, P.M. High lipid induction in microalgae for biodiesel production. Energies 2012, 5, 1532–1553. [Google Scholar] [CrossRef] [Green Version]
- Go, S.; Lee, S.-J.; Jeong, G.-T.; Kim, S.-K. Factors affecting the growth and the oil accumulation of marine microalgae, Tetraselmis suecica. Bioprocess Biosyst. Eng. 2011, 35, 145–150. [Google Scholar] [CrossRef]
- Nascimento, I.A.; Marques, S.S.I.; Cabanelas, I.T.D.; Pereira, S.A.; Druzian, J.I.; de Souza, C.O.; Vich, D.V.; de Carvalho, G.C.; Nascimento, M.A. Screening microalgae strains for biodiesel production: Lipid productivity and estimation of fuel quality based on fatty acids profiles as selective criteria. Bioenerg. Res. 2012, 6, 1–13. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Illumination Intensity (µmol m−2 s−1) | Specific Growth Rate 0–7 Days µ(d−1) | Doubling Time 0–7 Days (hr) | Biomass Productivity (g L−1 day−1) |
|---|---|---|---|
| 16/8-h light and dark cycle (PP) | |||
| 30 LL | 0.207 ± 0.01 | 80.26 ± 2.67 | 0.083 ± 0.007 |
| 60 ML | 0.256 ± 0.01 | 64.64 ± 1.37 | 0.143 ± 0.01 |
| 400 HL | 0.275 ± 0.01 | 60.32 ± 1.81 | 0.251 ± 0.012 |
| Continuous 24-h light (CL) | |||
| 30 LL | 0.251 ± 0.006 | 66.10 ± 1.50 | 0.161 ± 0.007 |
| 60 ML | 0.310 ± 0.02 | 54.32 ± 2.74 | 0.283 ± 0.016 |
| 400 HL | 0.344 ± 0.007 | 48.32 ± 1.11 | 0.343 ± 0.028 |
| Individual Carotenoids and Chlorophyll Content [%] of the Total Pigment | ||||||
|---|---|---|---|---|---|---|
| Pigment | Continuous 24-h Light CL | 16/8-h Light and Dark Cycle PP | ||||
| LL | ML | HL | LL | ML | HL | |
| Violaxanthin | 33.00 ± 3.00 | 21.50 ± 1.02 | 35.91 ± 2.38 | 20.68 ± 3.15 | 17.29 ± 0.69 | 22.60 ± 1.14 |
| Xantophyll-like voucheriaxanthin structure | 20.01 ± 2.01 | 13.78 ± 0.38 | 17.82 ± 0.60 | 16.39 ± 0.87 | 11.75 ± 0.61 | 11.90 ± 0.79 |
| Zeaxanthin | 0.82 ± 0.33 | 0.69 ± 0.14 | 3.58 ± 0.21 | 0.21 ± 0.06 | 0.23 ± 0.02 | 2.55 ± 0.10 |
| Chlorophyll a | 40.68 ± 1.65 | 59.25 ± 3.31 | 27.61 ± 0.85 | 57.97 ± 0.06 | 69.47 ± 2.70 | 53.52 ± 1.44 |
| β-carotene | 5.71 ± 0.88 | 4.13 ± 0.72 | 15.72 ± 1.14 | 4.96 ± 0.33 | 2.29 ± 0.04 | 11.12 ± 1.39 |
| Fatty Acid Profile | |||
|---|---|---|---|
| 16/8-h Light and Dark Cycle (PP) | |||
| illumination intensity (µmol m−2 s−1) | 30 LL | 60 ML | 400 HL |
| saturated fatty acids | 35.953 ± 2.907 | 34.468 ± 1.405 | 30.202 ± 3.083 |
| monounsaturated fatty acids | 43.320 ± 4.681 | 44.68 ± 2.036 | 38.238 ± 1.495 |
| polyunsaturated fatty acids | 2.395 ± 0.429 | 3.625 ± 0.455 | 14.973 ± 2.066 |
| total C16–18 (%,w/w) | 75.957 ± 1.110 | 78.993 ± 2.094 | 73.070 ± 1.570 |
| Continuous 24-h light (CL) | |||
| illumination intensity (µmol m−2 s−1) | 30 LL | 60 ML | 400 HL |
| saturated fatty acids | 23.703 ± 0.427 | 22.470 ± 1.483 | 24.685 ± 1.011 |
| monounsaturated fatty acids | 49.816 ± 2.398 | 51.597 ± 2.517 | 68.305 ± 2.537 |
| polyunsaturated fatty acids | 9.849 ± 2.777 | 7.750 ± 1.592 | 4.512 ± 0.238 |
| total C16–18 (%,w/w) | 78.355 ± 3.487 | 78.948 ± 5.424 | 93.960 ± 1.599 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzemińska, I.; Nosalewicz, A.; Reszczyńska, E.; Pawlik-Skowrońska, B. Enhanced Light-Induced Biosynthesis of Fatty Acids Suitable for Biodiesel Production by the Yellow-Green Alga Eustigmatos magnus. Energies 2020, 13, 6098. https://doi.org/10.3390/en13226098
Krzemińska I, Nosalewicz A, Reszczyńska E, Pawlik-Skowrońska B. Enhanced Light-Induced Biosynthesis of Fatty Acids Suitable for Biodiesel Production by the Yellow-Green Alga Eustigmatos magnus. Energies. 2020; 13(22):6098. https://doi.org/10.3390/en13226098
Chicago/Turabian StyleKrzemińska, Izabela, Artur Nosalewicz, Emilia Reszczyńska, and Barbara Pawlik-Skowrońska. 2020. "Enhanced Light-Induced Biosynthesis of Fatty Acids Suitable for Biodiesel Production by the Yellow-Green Alga Eustigmatos magnus" Energies 13, no. 22: 6098. https://doi.org/10.3390/en13226098