
Cyanobacterial Blooms: Current Knowledge and New Perspectives


1
CIIMAR/CIMAR, Interdisciplinary Centre of Marine and Environmental Research, University of Porto, Terminal de Cruzeiros do Porto de Leixões, Av. General Norton de Matos s/n, 4050-208 Porto, Portugal
2
Department of Biology, Faculty of Sciences, University of Porto, Rua do Campo Alegre, 4169-007 Porto, Portugal
*
Author to whom correspondence should be addressed.
Earth 2022, 3(1), 127-135; https://doi.org/10.3390/earth3010010
Submission received: 11 December 2021
/
Revised: 28 January 2022
/
Accepted: 29 January 2022
/
Published: 3 February 2022
Abstract
:Cyanobacteria are ancient prokaryotes responsible for bloom formation in many freshwater resources worldwide. These dense agglomerations are a result of the rise of nutrient input (N and P) or temperature. The toxin content and illness associated with contact impair human health with repercussions in water quality. Produced by a wide variety of cyanobacteria species, CyanoBlooms are in need of a literature review to achieve a global scenario of its current impacts on freshwater resources aiming at changing behaviors towards CyanoBlooms globally and by making communities more resilient to this recurrent problem. With a global distribution, recent data highlight the impacts of climate change on CyanoBlooms occurrence, namely through the rise of temperature and nutrient input from storms and heavy rainfall. With current worldwide regulations based on the enumeration of the nutrient input of freshwater ecosystems, the increase in field monitoring regarding CyanoBlooms occurrence is demanded since evaluation of this parameter may conceal these massive agglomerations resulting in human health episodes and cyanotoxin outbreaks.
1. Introduction
Cyanobacteria are ancient prokaryotes responsible for oxygen production in the Earth’s atmosphere. Their emergence dates back from 2.8 billion years ago [1]. Inhabitants of a wide range of ecosystems, from the Polar regions to other habitats such as cold and hot deserts, hot springs, and flood plain soils [2,3,4,5], cyanobacteria are microorganisms that possess a global occurrence. As primary producers of aquatic and terrestrial food chains, cyanobacteria currently have increasing biotechnological applications and as a source of new drugs for medical treatment [6]. In nature, the toxic and non-toxic forms of cyanobacteria can co-exist, but it is under a CyanoBloom that the amount of toxic forms increases, producing and releasing to the water column potent toxins designated as cyanotoxins. A CyanoBloom can be defined and characterized as an increase in cyanobacterial biomass in a relatively short period of time (few days to 2 weeks) and with the dominance (above 80%) of one or a few species of the phytoplankton community [7]. Factors such as eutrophication, anthropogenic pressure, or increase of temperature are associated with the proliferation of cyanobacteria and consequently of bloom formation. For these reasons, Cyanobacterial Blooms are in need of further insights due to their possible linkage to climate change. In addition, as toxin producers, CyanoBlooms, particularly in freshwater resources, are in need of constant vigilance and study since the changes in composition of blooms which may reflect toxin profile alterations have been reported, causing severe cyanotoxin outbreaks and associated human illness and animal deaths [8].
Freshwater systems have drinking, irrigation or recreational impacts and possess a diverse range of exposure routes to intoxications for humans and animals. Though climate change effects are more studied in the marine areas, the impacts of this phenomenon on freshwater ecosystems where CyanoBlooms may appear demand the need to intensify surveillance programs through analysis of bloom composition, occurrence, and toxicity. With statistical models, it is possible to predict future CyanoBlooms episodes by correlating the data (cyanobacterial biomass) with other environmental data [9]. In a CyanoBloom, toxins can be released to the water column where microcystins, cylindrospermopsins, anatoxins, and saxitoxins are of main concern [10]. Therefore, due to the underlined toxicity, nuisance, and linkage to climate change, a review is demanded of the current literature and knowledge to improve behaviors towards CyanoBlooms and make communities more resilient to this phenomenon fostering water quality and public health to tackle CyanoBlooms globally.
2. CyanoBlooms: Current Regulations
As ubiquitous microorganisms of freshwater resources, cyanobacteria tend to form blooms in the ecosystems, particularly after high levels of nutrients are found in the water or through the rise of temperature. Some strains find their ideal conditions for growth, compete, and proliferate intensively on the water surface causing visible masses of cyanobacteria that can be toxic and cause severe diseases (Figure 1). In both hemispheres, there have been reports on the occurrence of CyanoBlooms [11]. Climate alterations and increasing urbanization close to freshwater resources can exacerbate the occurrence of CyanoBlooms, leading to intoxication episodes due to unintentional contact of humans and domestic animals. In Portugal, recently, microcystins were found in the kidneys of death farmed cows after blooms were observed in a near water resource used for animal drinking [12]. Lack of knowledge and dissemination of the toxic properties of CyanoBlooms has led to intoxication episodes globally, constituting a problem in need of constant vigilance. Due to intensifications of CyanoBlooms, regulations on other cyanotoxins, apart from the common microcystins, were implemented in several countries. In fact, microcystins have currently an established guideline value of 1 μg/L in drinking water adopted by the World Health Organization (WHO) that implemented this provisional guideline [13]. In cylindrospermopsins, a guideline value for drinking water similar to microcystins has been proposed by Humpage and Falconer [14], despite the lack of recommendation by the WHO. However, on a global scale, three countries adopted the enumeration of cylindrospermopsins into their national legislation which include Australia (1 μg/L), New Zealand (1 μg/L), and Brazil (15 μg/L) [15]. Neurotoxins guideline values have also been adopted to the national legislation where New Zealand regulates both anatoxins (6 μg/L) and saxitoxins (3 μg/L), and Australia and Brazil regulate only saxitoxins (3 μg/L) [15]. These altogether reinforce the surveillance of freshwater systems through campaigns where screening of cyanotoxins can occur prior to bloom onset.
3. CyanoBlooms: Toxicity
Cyanotoxins are a chemically and biologically diverse group of secondary metabolites that are not produced by the primary metabolism of cyanobacteria. They are classified according to their mode of action in hepatotoxins (microcystins and nodularins), cytotoxins (cylindrospermopsin), neurotoxins (anatoxins and saxitoxins), and dermal toxins (aplysiatoxin and lyngbyatoxin) [16]. Some of these have proven effects on DNA damage (genotoxic) and promotion of cancer (carcinogenic) such as is the case for microcystins and cylindrospermopsins [17,18]. Human fatalities and animal deaths have been attributed to cyanotoxins worldwide. In Brazil, water contaminated with microcystins and cylindrospermopsins was responsible for the death of 60 patients in a dyalisis center that died after acute liver failure associated with the presence in the treatment water of these two cyanotoxins [19,20]. Another incident associated with cyanotoxins intoxication was reported in Australia where 148 people, mostly children, were hospitalized with symptoms of gastroenteritis [20]. Later, this episode was attributed to water contaminated with the cytotoxin cylindrospermopsins. Incidents with neurotoxins have been reported in France where dogs died after drinking water contaminated with anatoxins [21]. Cyanotoxins are synthetized non-ribosomically and are organized in gene clusters [22,23]. Their main characteristic is that they are non-strain specific meaning that more than one genus of cyanobacteria can synthetize the same cyanotoxin and the same genus can produce more than one type of cyanotoxin. Among all cyanotoxins, microcystins and cylindrospermopsins are the two most studied worldwide while the neurotoxins are the most dangerous [11].
3.1. Microcystins
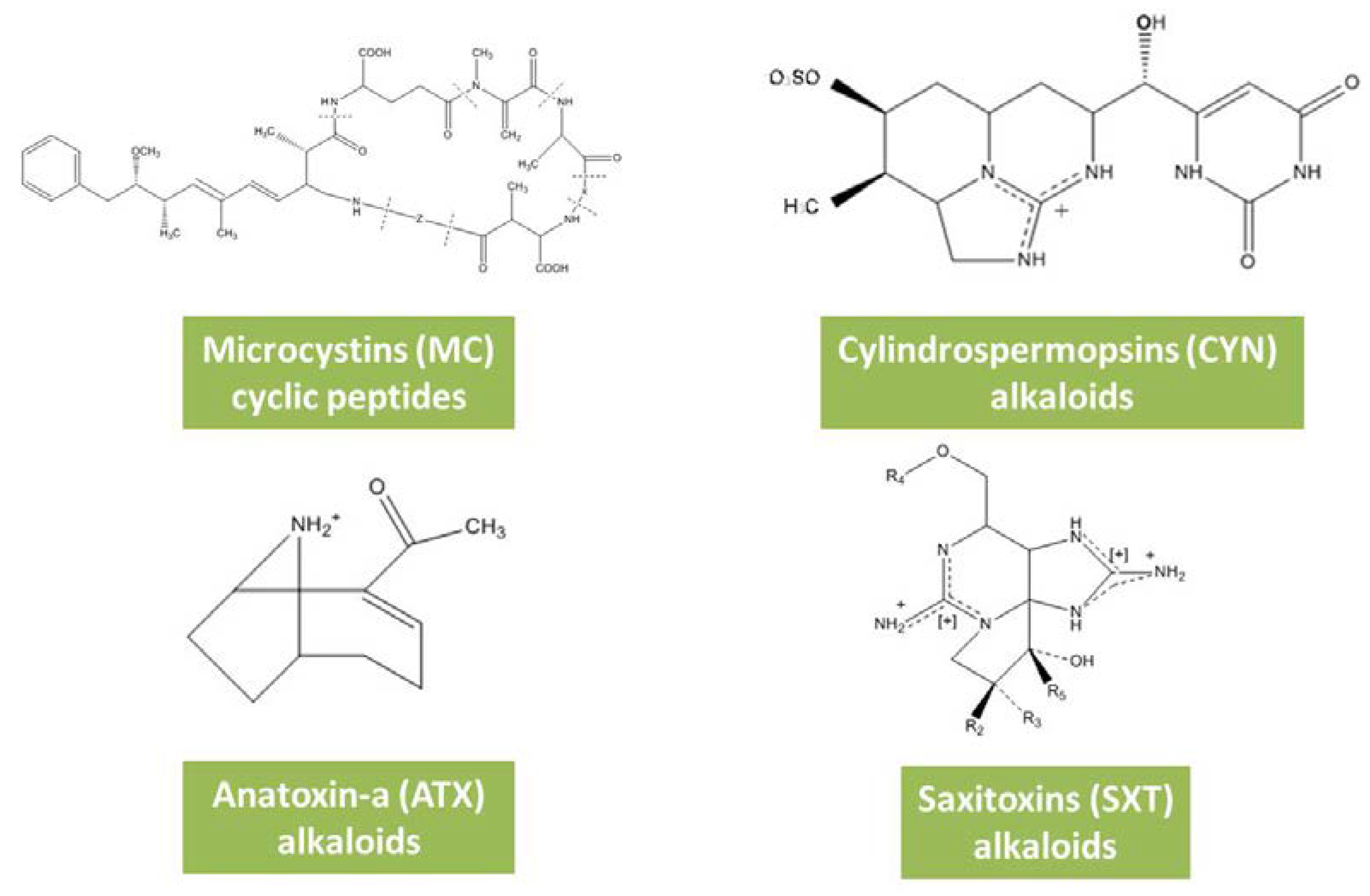
Microcystins (MCs) (Table 1) are hepatotoxins characterized by a group of cyclic heptapeptides whose chemical structure is characterized as a cyclo(D-Ala-L-X-D-erythro-β-methylAsp-L-Z-Adda-D-Glu-N-methyldehydro-Ala), where Adda is translated as the β-amino acid (2S,3S,8S,9S)-3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4,6-dienoic acid, which is found solely in the Cyanobacteria group [24]. X and Z positions in MC chemical structure correspond to varied amino acid residues where the most common are leucine and arginine whose outcome is MCLR (Figure 2). With these variations, more than 200 structural isoforms of MCs can be produced by cyanobacteria [24]. MCs act as specific inhibitors of protein phosphatases type 1 and 2A resulting in increased phosphorylation of proteins in liver cells which may affect metabolic pathways or cell division [25,26]. MCs producing genera include Chrysosporum, Dolichospermum, Limnothrix, Microcystis, Nostoc, Phormidium, and Planktothrix [24]. CyanoBlooms of MCs due to its varied species production are still the most commonly reported, globally demonstrating that, despite global changes, this is the cyanotoxin of most concern in water quality and human health [27,28].
3.2. Cylindrospermopsins
Cylindrospermopsins (CYN) (Table 1) are cytotoxins chemically characterized as alkaloids that can also affect the liver and nervous systems [29,30]. CYN chemical structure encompasses a guanidine group and a hydroxide group associated with a tricyclic-carbon skeleton (Figure 2) [31]. Despite CYN high stability, the molecule can suffer minor changes in its chemical structure, being currently found to possess four variants: 7-epi-CYN, 7-deoxy-CYN, 7-deoxy-desulfo-CYN and 7-deoxy-desulfo-12-acetyl-CYN [32]. CYN acts as a potent inhibitor of protein synthesis [33]. Initially described as a liver toxin, CYN is currently a cytotoxin also with genotoxic effects and a carcinogenic potential [17]. Produced by a variety of species including Cylindrospermopsis raciborskii, Aphanizomenon ovalisporum, Aphanizomenon flos-aquae, Aphanizomenon gracile, Aphanizomenon klebahnii, Umezakia natans, Raphidiopsis curvata, Anabaena bergii, Anabaena planctonica, Anabaena lapponica and Lyngbya wollei [34]. Blooms of CYN are at the moment poorly reported, despite CYN being considered the second most studied cyanotoxin worldwide. The Palm Island incident, after a bloom of C. raciborskii in the Soloman dam, is until now the only CYN CyanoBloom described, despite its production by diverse strains.
3.3. Anatoxins
Anatoxins (ATX) (Table 1) are a group of cyanotoxins with neurotoxic effects chemically characterized as an alkaloid composed of a bicyclic secondary amine (Figure 2) [35]. Anatoxin-a is highly unstable in nature, being easily converted into non-toxic metabolites, namely dihydroanatoxin-a and epoxyanatoxin-a [35]. The most studied metabolite is anatoxin-a with proven toxicity in the genera Anabaena, Aphanizomenon, Cylindrospermum, Oscillatoria, Microcystis, Raphidiopsis, Planktothrix, Artrospira, Nostoc and Phormidium [35,36]. ATX outbreaks relate to an episode associated with blooms of Phormidium spp. that had neurotoxin production and resulted in the death of dogs after making contact with these cyanobacterium mats in both France and New Zealand [20,37]. In fact, with regard to ATX, no human fatalities have been reported until now, despite the several deaths reported in wild and domestic animals [38].
3.4. Saxitoxins
Saxitoxins (SXT) (Table 1) are a neurotoxic alkaloid where the chemical structure is represented by tetrahydropurines with varied amino acids in the R positions (Figure 2) [39,40]. Its tricyclic structure brings to saxitoxins a high resistance to extreme environmental conditions [40]. Currently presenting 58 variants, saxitoxins can be produced by varied genera of marine dinoflagellates including Alexandrium, Gymnodinium, and Pyrodinium. In cyanobacteria, Anabaena, Aphanizomenon, Cylindrospermopsis, Lyngbya, and Planktothrix genera are the most frequently found [29,39,40]. Despite its reported production in several genera, SXT CyanoBlooms are currently only reported in dinoflagellates and intoxications normally associated with seafood consumption [29].
4. CyanoBlooms: Evaluation Methods
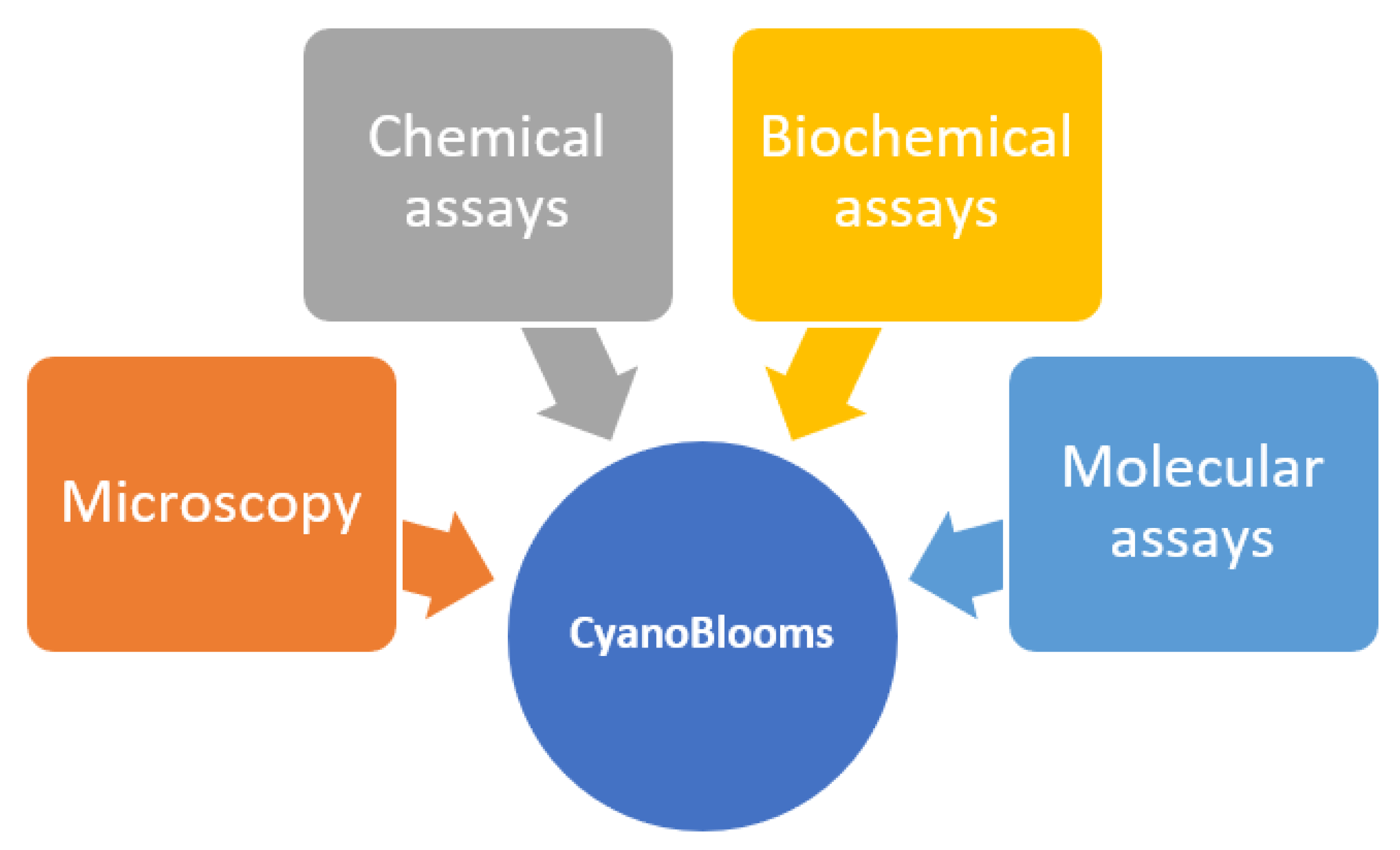
4.1. Microscopy
Assessing cyanobacteria species or genera can be performed by applying microscopy methods (Figure 3). The most traditional is light microscopy and in this the inspection for cyanobacteria composing CyanoBlooms can be achieved. Although microscopy is the most common method of all the main limitations, it requires extensive expertise and is unable to identify toxic taxa. Microscopy methods though greatly applied are becoming surpassed by other methods that permit the enumeration of cyanotoxins, most of them required by legislation. Despite their disadvantages, they have been used in the inspection of CyanoBloom samples [8].
4.2. Chemical Assays
Detection and enumeration of cyanotoxins from a complex matrix such as CyanoBlooms can be achieved by applying chemical methods (Figure 3). These englobe mainly High-Performance Liquid Chromatography (HPLC) and Liquid Chromatography Mass Spectrometry (LCMS) coupled with MS that permit in a few days from sampling the retrieval of the cyanotoxins molecule present in a CyanoBloom or in a water sample. As high specific methods, limitations encompass the lack of detection and enumeration of all main cyanotoxins in a given sample. The most recent chemical method developed englobes the screening through the LCMS technique of nine cyanotoxins in pure and natural waters including CYN, ATX, and six variants of MC including the common MCLR [41]. Chemical methods have also been applied in cyanotoxin screening in samples such as food supplements [42] and also in CyanoBloom samples [43]. These methods were the first to be applied in cyanotoxins investigations, being associated with the report of the first cyanotoxin ever enumerated, the anatoxin-a, from a cattle poisonous bloom of Anabaena flos-aquae [44].
4.3. Biochemical Assays
Biochemical assays englobe the most commonly known as Enzyme-Linked Immunosorbent Assay (ELISA) that possess a higher sensitivity and lower specificity to cyanotoxins than the chemical assays (Figure 3). Despite these specificities, these methods are commonly used in water samples to screen for cyanotoxin presence in the study of ecosystems for cyanotoxin occurrence [8]. These methods also allow, similarly to the chemical methods, the detection and enumeration of cyanotoxins if the proper commercial cyanotoxin kit is used, meaning that, if multiple toxins are present, a kit for each cyanotoxin needs to be purchased. Abraxis currently commercializes ELISA kits for all main cyanotoxins with a time response of a few hours.
4.4. Molecular Assays
Discovered since the mid 2000s, molecular methods constitute an alternative method for cyanotoxin detection (Figure 3). The potential of cyanotoxicity of these methods are a limitation since the presence of toxicity genes does not represent actual toxicity due to the reported presence of a gene inactivation [45]. Despite toxicity genes being described in the environment, the producing capability of the strains needs to be clarified by other methods, namely the chemical ones. As the gene clusters of each cyanotoxin are annotated and sequenced in varied strains, the development of new tools (primers) that in a general or specific manner contribute to the environmental detection of toxigenic strains is increasing. Polymerase chain reaction (PCR) methods are the most commonly applied and developed, being normally associated with primer development [8,46]. These methods further allow other inferences through phylogenetic analysis and possible biogeographic investigations [47,48].
4.5. Artificial Intelligence Methods
In CyanoBlooms and in cyanotoxins research, there is no known application of artificial intelligence in their study. However, the outcome of interactive rooms portraying ecosystems with CyanoBlooms may permit surpassing limitations such as poor education and foster a change in behaviors to tackle CyanoBlooms health impairment. Promising machines with artificial intelligence on the identification and toxicity of CyanoBlooms are lacking at the moment.
5. Future Perspectives
Evaluating nutrient input is mandatory but not satisfactory since evaluation leads to classification of freshwater resources as sensible to eutrophication, but several factors can contribute to CyanoBlooms. These include rise of temperature, nutrient input by agricultural runoff and sewage discharges and finally climate change storms where heat events and rainfall can also lead to the rise of nutrients in freshwater resources. For this, the monitoring of freshwater resources for CyanoBlooms occurrence is relevant to infer on the effects of climate change and on the effects on human health contributing to altering behaviors towards Cyanobacteria Blooms globally and simultaneously making communities more resilient to this frequent problem, namely by the implementation of artificial intelligence methods. Therefore, continued research is required to assess possible relations between nutrient input and the occurrence of CyanoBlooms with particular emphasis on current climate conditions.
6. Conclusions
As toxin producers and a health hazard, CyanoBlooms are required to be continuously studied and reported globally. Under the scenario of global changes, the risk of CyanoBloom occurrence may increase, requiring a literature review to surpass limitations in its study and foster new challenges and behaviors. As an environmental assessment, its monitoring frequency should be increased since the presence of nutrients may hamper CyanoBloom reports that can occur after climate change phenomena such as heat waves or storms. In view of this relationship, much needs to be investigated with artificial intelligence methods assisting in the alert and education of communities. Therefore, with this review, the essential information on CyanoBlooms is summarized, globally reinforcing their environmental procurement and the monitorization for their occurrence, toxicity, and composition.
Author Contributions
Conceptualization, C.M.; investigation, C.M.; resources, C.M.; data curation, C.M.; writing—original draft preparation, C.M.; writing—review and editing, V.V. and A.A. visualization, C.M.; supervision, V.V. and A.A; project administration, A.A.; funding acquisition, V.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by national funds through FCT-Foundation for Science and Technology within the scope of UIDB/04423/2020 and UIDP/04423/2020 and to the Postdoctoral fellowship attributed to Cristiana Moreira (SFRH/BPD/122909/2016) by the FCT.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are thankful for all the constructive comments and suggestions provided by the four anonymous reviewers, which greatly improved the quality of this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Summons, R.E.; Jahnke, L.L.; Hope, J.M.; Logan, G.A. 2-Methylhopanoids as biomarkers for cyanobacterial oxygenic photosynthesis. Nature 1999, 400, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.D.; Hawes, I.; Mountfort, D.; Hitzfeld, B.; Dietrich, D.R.; Burns, B.P.; Neilan, B.A. Diversity within cyanobacterial mat communities in variable salinity meltwater ponds of McMurdo Ice Shelf, Antarctica. Environ. Microbiol. 2005, 7, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papke, R.T.; Ramsing, N.B.; Bateson, M.M.; Ward, D.M. Geographic isolation in hot spring cyanobacteria. Environ. Microbiol. 2003, 5, 650–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinklebe, J.; Langer, U. Microbial diversity in three floodplain soils at the Elbe River (Germany). Soil Biol. Biochem. 2006, 38, 2144–2151. [Google Scholar] [CrossRef]
- Garcia-Navarro, F.J.; Perona, E.; Cubero, S.; Bravo, S.; Jiménez-Ballesta, R. Primary producers and anthropic signs related to the flood plain soils of the Tablas de Daimiel Wetland. Geosciences 2018, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. (Tokyo) 2011, 64, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Humbert, J.-F.; Fastner, J. Ecology of Cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2017; pp. 11–18. [Google Scholar]
- Moreira, C.; Gomes, C.; Vasconcelos, V.; Antunes, A. Cyanotoxins Occurrence in Portugal: A New Report on Their Recent Multiplication. Toxins 2020, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Christensen, V.G.; Stelzer, E.A.; Eikenberry, B.C.; Olds, H.T.; LeDuc, J.F.; Maki, R.P.; Saley, A.M.; Norland, J.; Khan, E. Cyanotoxin mixture models: Relating environmental variables and toxin co-occurrence to human exposure risk. J. Haz. Mat. 2021, 415, 125560. [Google Scholar] [CrossRef]
- van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.A.; Bakker, G.J.I. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef]
- Falconer, I.R.; Humpage, A.R. Health risk assessment of cyanobacterial (blue-green algal) toxins in drinking water. Int. J. Environ. Res. Public Health 2005, 2, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Menezes, C.; Nova, R.; Vale, M.; Azevedo, J.; Vasconcelos, V.; Pinto, C. First description of an outbreak of cattle intoxication by cyanobacteria (blue-green algae) in the South of Portugal. Bov. Pract. 2019, 53, 66–70. [Google Scholar]
- WHO. Guidelines for Drinking-water Quality. In Addendum to Vol. 2: Health Criteria and Other Supporting Information, 2nd ed.; World Health Organization: Geneva, Switzerland, 1998; 283p. [Google Scholar]
- Humpage, A.R.; Falconer, I.R. Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice: Determination of no observed adverse effect level for deriving a drinking water guideline value. Environ. Toxicol. 2003, 18, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Burch, M.D. Effective doses, guidelines & regulations. Adv. Exp. Med. Biol. 2008, 619, 831–853. [Google Scholar] [PubMed]
- Carmichael, W.W.; Li, R. Cyanobacteria toxins in the Salton Sea. Saline Syst. 2006, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Moreira, C.; Azevedo, J.; Antunes, A.; Vasconcelos, V. Cylindrospermopsin: Occurrence, methods of detection and toxicology. J. Appl. Microbiol. 2012, 114, 605–620. [Google Scholar] [CrossRef]
- Zanchett, G.; Oliveira-Filho, E.C. Cyanobacteria and Cyanotoxins: From Impacts on Aquatic Ecosystems and Human Health to Anticarcinogenic Effects. Toxins 2013, 5, 1896–1917. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Azevedo, S.M.F.O.; An, J.S.; Molica, R.J.R.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.S.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.; Antunes, M.B.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.C.; Humbert, J.F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef]
- Carmichael, W. Cyanobacteria secondary metabolites—the cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef]
- Pearson, L.A.; Moffitt, M.C.; Ginn, H.P.; Neilan, B.A. The molecular genetics and regulation of cyanobacterial peptide hepatotoxin biosynthesis. Crit. Rev. Toxicol. 2008, 38, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Catherine, A.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2017; pp. 109–126. [Google Scholar]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. The phosphoprotein phosphatase family of Ser/Thr phosphatases as principal targets of naturally occurring toxins. Crit. Rev. Toxicol. 2011, 41, 83–110. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. Computational study of the covalent bonding of microcystins to cysteine residues—A reaction involved in the inhibition of the PPP family of protein phosphatases. FEBS J. 2013, 280, 674–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.; Pereira, M.J. Microcystin-producing blooms—A serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Dhanji-Rapkova, M.; O’Neill, A.; Coates, L.; Lewis, A.; Lewis, K. Analysis of Microcystins in Cyanobacterial Blooms from Freshwater Bodies in England. Toxins 2018, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B.A. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [Green Version]
- Neilan, B.A.; Pearson, L.A.; Muenchoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Muenchoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [Green Version]
- WHO. Cyanobacterial toxins: Cylindrospermopsins. In Background Document for Development of WHO Guidelines for Drinking-Water Quality and Guidelines for Safe Recreational Water Environments; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Kokocinski, M.; Camean, A.M.; Carmeli, S.; Guzmán-Guillén, R.; Jos, Á.; Mankiewicz-Boczek, J.; Metcalf, J.S.; Moreno, I.M.; Prieto, A.I.; Sukenik, A. Cylindrospermopsin and Congeners. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2017; pp. 127–137. [Google Scholar]
- Moreira, C.; Mendes, R.; Azevedo, J.; Vasconcelos, V.; Antunes, A. First occurrence of cylindrospermopsin in Portugal: A contribution to its continuous global dispersal. Toxicon 2017, 130, 87–90. [Google Scholar] [CrossRef]
- Osswald, J.; Rellán, S.; Gago, A.; Vasconcelos, V. Toxicology and detection methods of the alkaloid neurotoxin produced by cyanobacteria, anatoxin-a. Environ. Int. 2007, 33, 1070–1089. [Google Scholar] [CrossRef]
- Ballot, A.; Fastner, J.; Lentz, M.; Wiedner, C. First report of anatoxin-a producing cyanobacterium Aphanizomenon issatschenkoi in northeastern Germany. Toxicon 2010, 56, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Selwood, A.I.; Rueckert, A.; Holland, P.T.; Milne, J.R.; Smith, K.F.; Smits, B.; Watts, L.F.; Cary, C.S. First report of homoanatoxin-a and associated dog neurotoxicosis in New Zealand. Toxicon 2007, 50, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Colas, S.; Marie, B.; Lance, E.; Quiblier, C.; Tricoire-Leignel, H.; Mattei, C. Anatoxin-a: Overview on a harmful cyanobacterial neurotoxin from the environmental scale to the molecular target. Environ. Res. 2021, 193, 110590. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Cyanobacteria toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; WHO: Geneva, Switzerland; E&FN Spon: London, UK, 1999. [Google Scholar]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptores. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef]
- Yen, H.K.; Lin, T.F.; Liao, P.C. Simultaneous detection of nine cyanotoxins in drinking water using dual solid-phase extraction and liquid chromatography-mass spectrometry. Toxicon 2011, 58, 209–218. [Google Scholar] [CrossRef]
- Roy-Lachapelle, A.; Solliec, M.; Bouchard, M.F.; Sauvé, S. Detection of Cyanotoxins in Algae Dietary Supplements. Toxins 2017, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Teta, R.; Della Sala, G.; Glukhov, E.; Gerwick, L.; Gerwick, W.H.; Mangoni, A.; Costantino, V. Combined LC-MS/MS and Molecular Networking Approach Reveals New Cyanotoxins from the 2014 Cyanobacterial Bloom in Green Lake, Seattle. Environ. Sci. Technol. 2015, 49, 14301–14310. [Google Scholar] [CrossRef]
- Devlin, J.P.; Edwards, O.E.; Gorham, P.R.; Hunter, N.R.; Pike, R.K.; Stavric, B. Anatoxin-a, a toxic alkaloid from Anabaena flos-aquae NRC-44h. Can. J. Chem. 1977, 55, 1367–1371. [Google Scholar] [CrossRef]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef]
- Moreira, C.; Pimentel, A.; Vasconcelos, V.; Antunes, A. Preliminary evidence on the presence of cyanobacteria and cyanotoxins from culture enrichments followed by PCR analysis: New perspectives from Africa (Mali) and South Pacific (Fiji) countries. Environ. Sci. Pollut. Res. 2021, 28, 31731–31745. [Google Scholar] [CrossRef]
- Ribeiro, K.F.; Duarte, L.; Crossetti, L.O. Everything is not everywhere: A tale on the biogeography of cyanobacteria. Hydrobiologia 2018, 820, 23–48. [Google Scholar] [CrossRef]
- Moreira, C.; Vasconcelos, V.; Antunes, A. Genetic characterization of Microcystis aeruginosa isolates from Portuguese freshwater systems. World J. Microbiol. Biotechnol. 2016, 32, 118. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Photograph of a CyanoBloom (Microcystis sp.) in a river in the North of Portugal.

Figure 2.
Chemical structure of cyanotoxins associated with CyanoBlooms.

Figure 3.
Current methods available in CyanoBloom investigations.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the cyanobacterial toxicity associated with CyanoBlooms.
| Cyanotoxins | Chemical Structure | Effect | Strains | Maximum Permissible Concentration | References |
|---|---|---|---|---|---|
| Microcystins | Cyclic peptides | Hepatotoxins | Chrysosporum, Dolichospermum, Limnothrix, Microcystis, Nostoc, Phormidium and Planktothrix | 1 μg/L | [12,24,25] |
| Cylindrospermopsins | Alkaloids | Cytotoxins | Cylindrospermopsis raciborskii, Aphanizomenon ovalisporum, Aphanizomenon flos-aquae, Aphanizomenon gracile, Aphanizomenon klebahnii, Umezakia natans, Raphidiopsis curvata, Anabaena bergii, Anabaena planctonica, Anabaena lapponica and Lyngbya wollei | 1 μg/L | [14,29,34] |
| Anatoxins | Alkaloids | Neurotoxins | Anabaena, Aphanizomenon, Cylindrospermum, Oscillatoria, Microcystis, Raphidiopsis, Planktothrix, Artrospira, Nostoc and Phormidium | 6 μg/L | [14,35,36] |
| Saxitoxins | Alkaloids | Neurotoxins | Anabaena, Aphanizomenon, Cylindrospermopsis, Lyngbya and Planktothrix | 3 μg/L | [14,29,39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moreira, C.; Vasconcelos, V.; Antunes, A. Cyanobacterial Blooms: Current Knowledge and New Perspectives. Earth 2022, 3, 127-135. https://doi.org/10.3390/earth3010010
AMA Style
Moreira C, Vasconcelos V, Antunes A. Cyanobacterial Blooms: Current Knowledge and New Perspectives. Earth. 2022; 3(1):127-135. https://doi.org/10.3390/earth3010010
Chicago/Turabian StyleMoreira, Cristiana, Vitor Vasconcelos, and Agostinho Antunes. 2022. "Cyanobacterial Blooms: Current Knowledge and New Perspectives" Earth 3, no. 1: 127-135. https://doi.org/10.3390/earth3010010