Complexity Changes in Brain Activity in Healthy Ageing: A Permutation Lempel-Ziv Complexity Study of Magnetoencephalograms

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
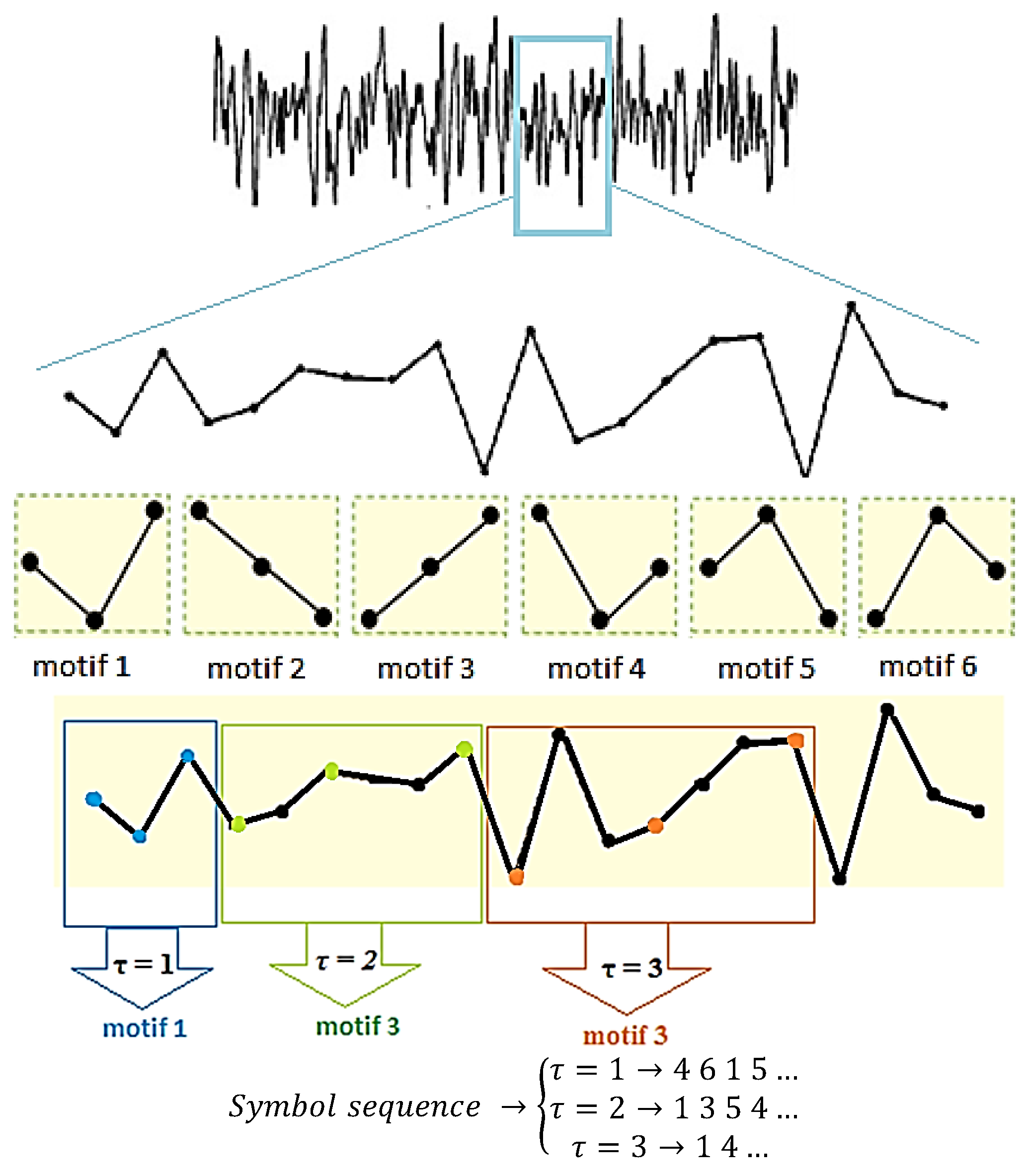
2.1. Permutation Lempel-Ziv Complexity
2.2. Statistical Analysis
3. Results
3.1. Selection of Motif Length
3.2. Analysis of Short Term Variability of Complexity Values
- In group 1, there were no significant differences (p > 0.0125) between the complexity values of minutes 3 to 5, however minutes 1 and 2 contained complexity values that were significantly different to the other minutes of recording (p < 0.0125),
- In group 2, only minute 1 had significantly different complexity values (p < 0.0125) when compared to the complexity values of the other 4 min of recording,
- In group 3, minute 5 had significantly different PLZC values (p < 0.0125) when compared to other minutes, however, there were no significant differences between complexity values between minutes 1 and 4.
- Group 1, only minutes 1 and 2 had similar complexity values (p > 0.0125), while minutes 3 to 5 had significantly different values (p < 0.0125),
- Group 2, although minutes 1, 4 and 5 did not have significantly different values (p > 0.0125), minutes 2, 3 and 4 also did not have any significantly different values (p > 0.0125), thus, no clear and obvious trends were found, as all minutes were observed to have varying PLZC values,
- Group 3, similar to the observation made for females, minute 5 had complexity values that were significantly different to all other minutes of recording.
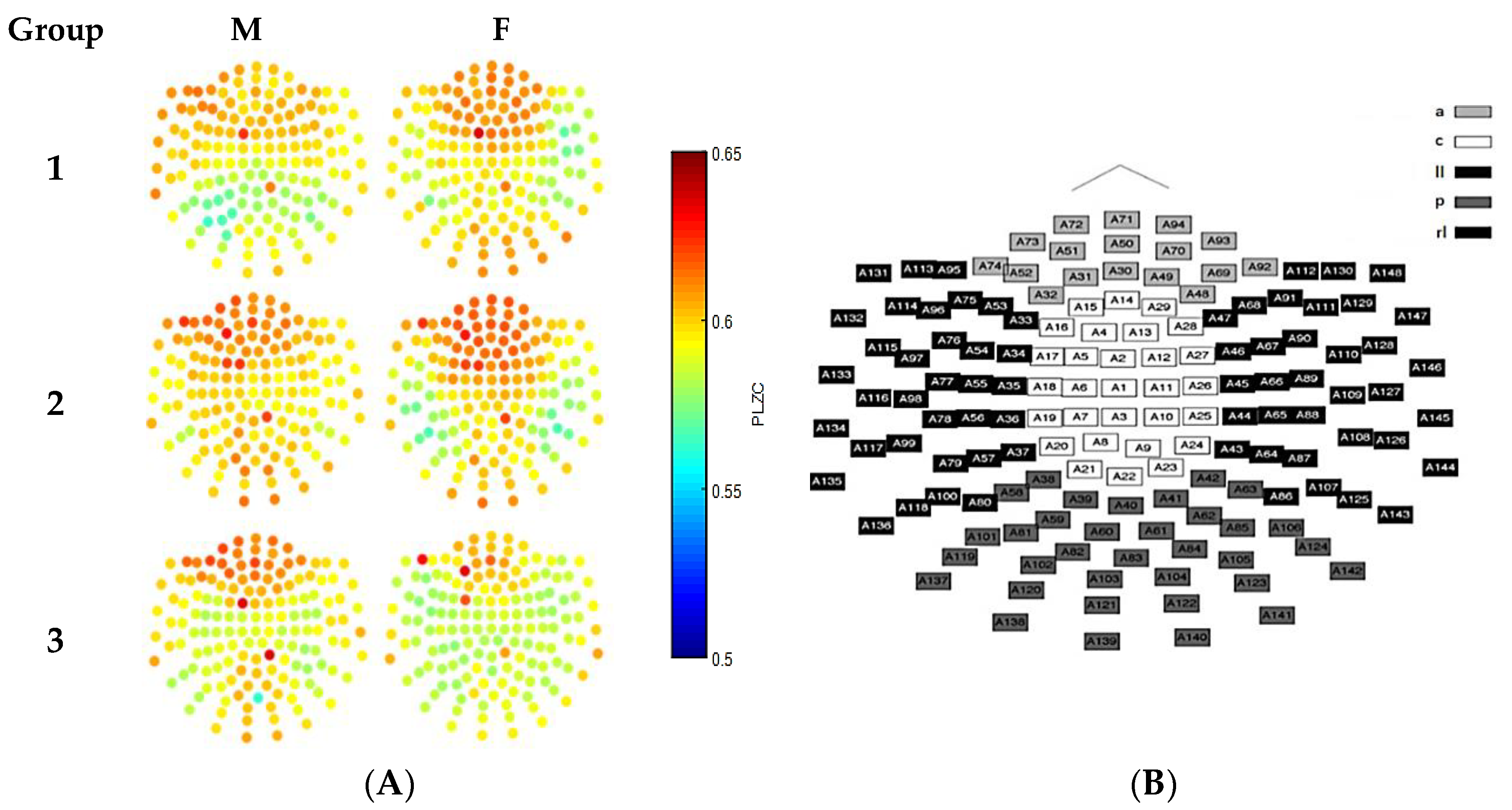
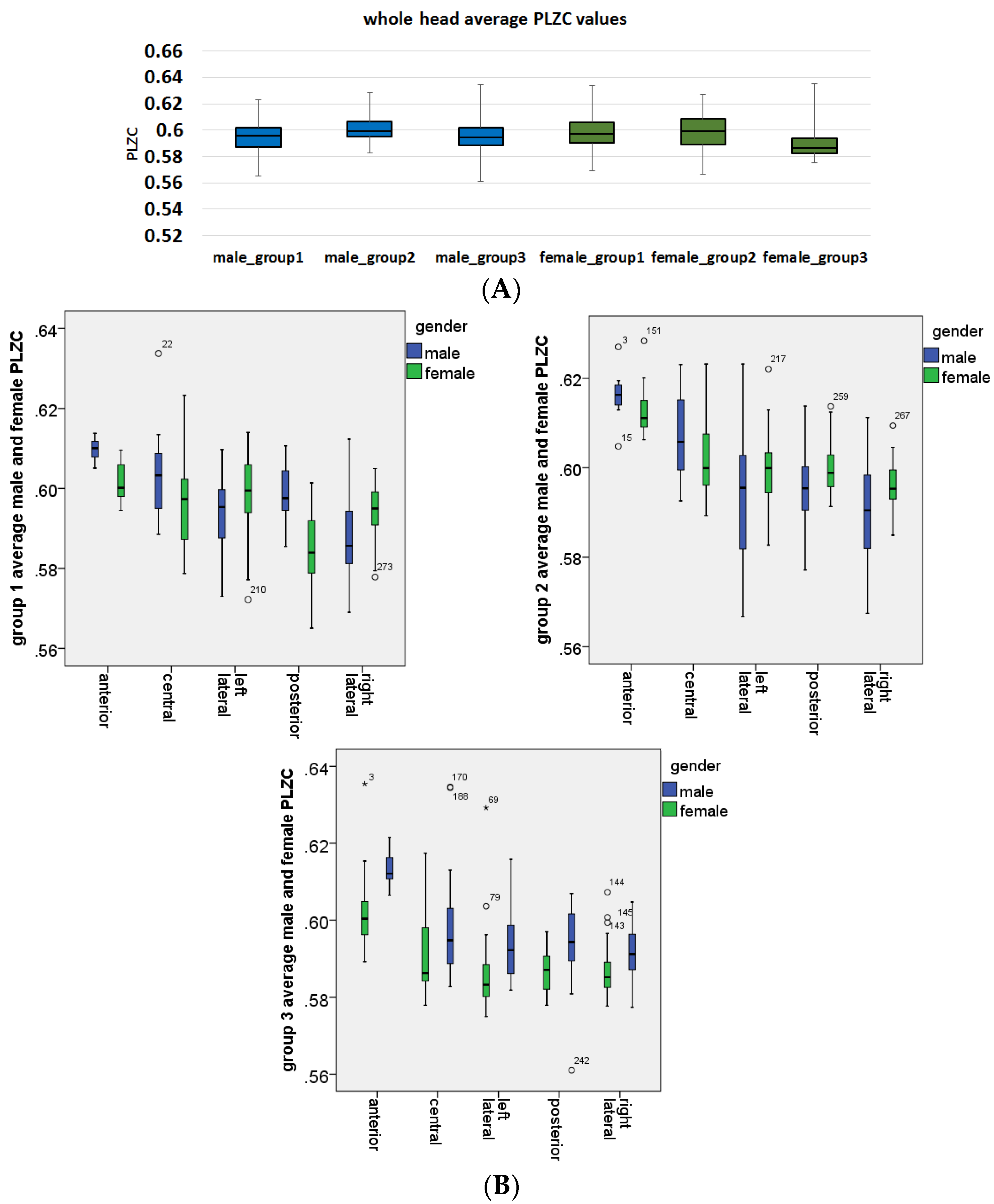
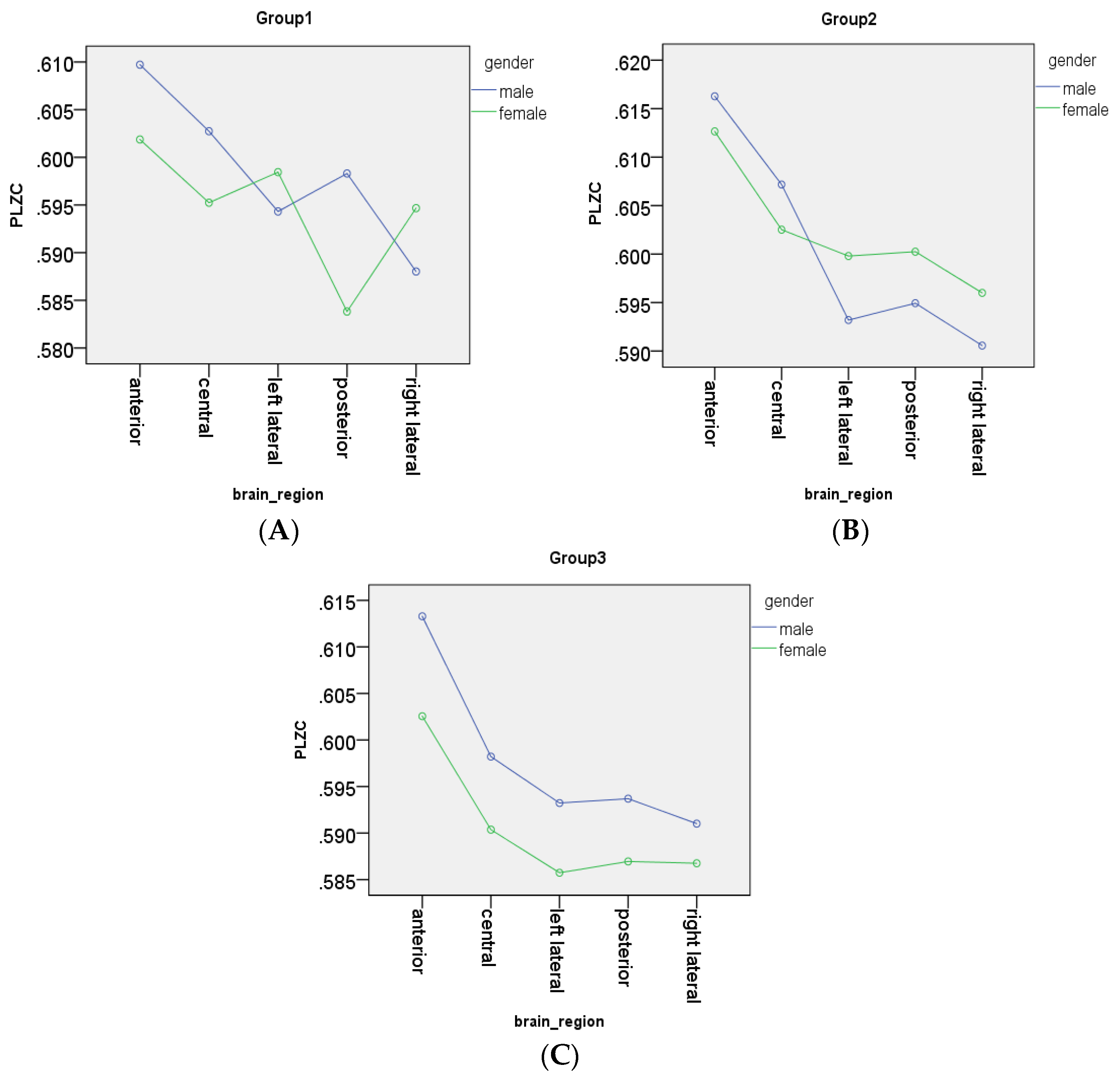
3.3. Analysis of Gender Effects
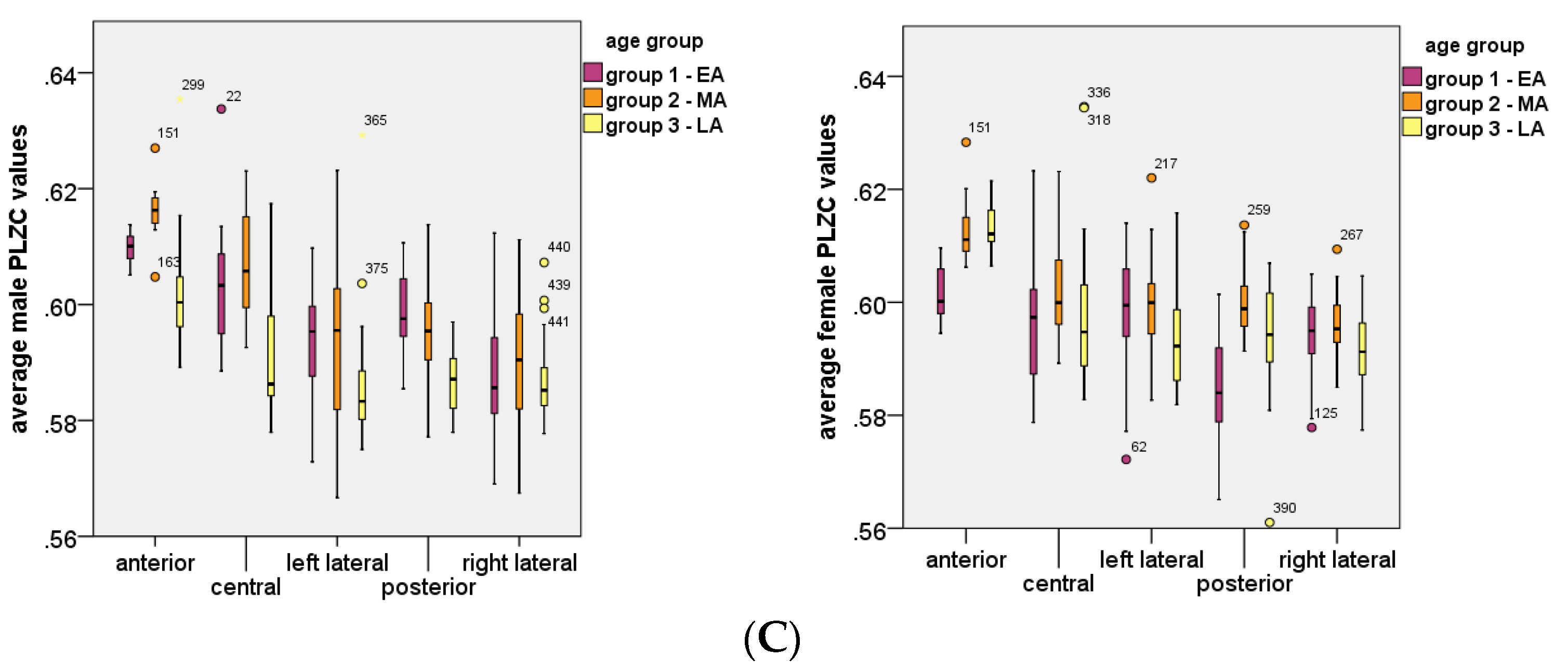
3.4. Analysis of Age Effects
3.5. Analysis of Age and Gender Interaction Effects
- In the anterior region, group 2 (p = 0.229) did not have a significant age and gender interaction effect on PLZC values., However, in group 1 (p = 0.0095) and group 3 (p = 0.000398) there was a significant interaction effect, which showed that PLZC values in this brain region (for both age groups) were influenced by gender, with complexity values being higher in males than in females.
- In the central region, group 1 (p = 0.0012), group 2 (p = 0.0441) and group 3 (p = 0.00717) all had significant interaction effects, which showed that males had higher PLZC values than females, and therefore gender influenced complexity values as a function of age for all three groups.
- In the left lateral region, group 2 (p = 0.002; females PLZC values were greater than males) and group 3 (p = 0.00047; male PLZC values were greater than females) had a significant interaction effect on PLZC values, while group 1 (p = 0.053) did not.
- In the posterior region, all groups had a significant interaction effect on PLZC values, with pgroup1 < 0.0001, pgroup2 = 0.0129, and pgroup3 = 0.00163. Thus, gender has a significant influence on PLZC values as a function of age, with males having higher complexity in group 1 and 3, and females having higher complexity in group 2.
- In the right lateral region, all groups had a significant age and gender interaction effect on PLZC values, with pgroup1 = 0.00189, pgroup2 = 0.0109, and pgroup3 = 0.0463. Gender has a significant effect as a function of age in all three groups, with females having higher complexity in group 1 and 2, while males had higher complexity values in group 2.
4. Discussion
4.1. Short Term Variability of Complexity Values
4.2. Effects of Age on Complexity Values
4.3. Effects of Gender on Complexity Values
4.4. Interaction Effects of Age and Gender on Complexity Values
4.5. Potential Clinical Singificance of Results
4.6. Limitations and Future Work
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Anokhin, A.; Birbaumer, N.; Lutzenberger, W.; Nikolaev, A.; Vogel, F. Age increases brain complexity. Electroencephalogr. Clin. Neurophysiol. 1996, 99, 63–68. [Google Scholar] [CrossRef]
- Meyer-Lindenber, A. The evolution of complexity in human brain development: An EEG study. Electroencephalogr. Clin. Neurophysiol. 1996, 99, 405–411. [Google Scholar] [CrossRef]
- Clarke, A.; Barry, R.; McCarthy, R.; Selikowitz, M. Age and sex effects in the EEG: Development of the normal child. Clin. Neurophysiol. 2001, 112, 806–814. [Google Scholar] [CrossRef]
- Fernandez, A.; Zuluaga, P.; Abasolo, D.; Gomez, C.; Serra, A.; Mendez, M.; Hornero, R. Brain oscillatory complexity across the life span. Clin. Neurophysiol. 2012, 123, 2154–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, E.; Thompson, P. Functional Brain Connectivity Using fMRI in Aging and Alzheimer’s Disease. Neuropsychol. Rev. 2014, 24, 49–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bivard, A.; Levi, C.; Krishnamurthy, V.; Hislop-Jambrich, J.; Salazar, P.; Jackson, B.; Davis, S.; Parsons, M. Defining acute ischemic stroke tissue pathophysiology with whole brain CT perfusion. J. Neuroradiol. 2014, 41, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Scholl, M.; Lockhart, S.; Schonhaut, D.; O’Neil, J.; Janabi, M.; Ossenkoppele, R.; Baker, S.; Vogel, J.; Faria, J.; Schwimmer, H.D.; et al. PET Imaging of Tau Deposition in the Aging Human Brain. Neuron 2016, 89, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Thut, G.; Bergmann, T.; Frohlich, F.; Soekadar, S.; Brittain, J.; Valero-Cabre, A.; Ack, A.S.; Miniussi, C.; Antal, A.; Siebner, H.; et al. Guiding transcranial brain stimulation by EEG/MEG to interact with ongoing brain activity and associated functions: A position paper. Clin. Neurophysiol. 2017, 128, 843–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torricelli, A.; Contini, D.; Pifferi, A.; Caffini, M.; Re, R.; Zucchelli, L.; Spinelli, L. Time domain functional NIRS imaging for human brain mapping. NeuroImage 2014, 85, 28–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, S.; Okada, Y. Contributions of principal neocortical neurons to magnetoencephalography and electroencephalography signals. J. Physiol. 2006, 575, 925–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stam, C.J.; van Straaten, E.C.; van Dellen, E.; Tewarie, P.; Gong, G.; Hillebrand, A.; Meier, J.; van Mieghem, P. The relation between structural and functional connectivity patterns in complex brain networks. Int. J. Psychophysiol. 2016, 103, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Stam, C.; Nolte, G.; Daffertshofer, A. Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Hum. Brain Mapp. 2007, 28, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- Englot, D.; Rolston, J.; Wang, D.; Kirsch, H.; Nagarajan, S.; Chang, E. 206 Spikes, Slowing, and Functional Connectivity: Multimodal Magnetoencephalography in Epilepsy Surgery. Neurosurgery 2016, 63, 181. [Google Scholar] [CrossRef]
- Escudero, J.; Hornero, R.; Abasolo, D.; Fernandez, A. Blind source separation to enhance spectral and non-linear features of magnetoencephalogram recordings. Application to Alzheimer’s disease. Med. Eng. Phys. 2009, 31, 872–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maestú, F.; Solesio-Jofre, E.; Bajo, R. Towards the Understanding of Healthy and Pathological Aging Through MEG. In International Review Neurobiology; Supek, S., Aine, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 609–640. [Google Scholar]
- Shumbayawonda, E.; Fernández, A.; Hughes, M.P.; Abasolo, D. Permutation Entropy for the Characterisation of Brain Activity Recorded with Magnetoencephalograms in Healthy Ageing Entropy. Entropy 2017, 19, 141. [Google Scholar] [CrossRef]
- Vicente, R.; Wibral, M.; Lindner, M.; Pipa, G. Transfer entropy-a model-free measure of effective connectivity for the neurosciences. J. Comput. Neurosci. 2011, 30, 45–67. [Google Scholar] [CrossRef] [PubMed]
- Pijnenburg, Y.; Made, Y.V.; van Walsum, A.V.; Knol, D.; Scheltens, P.; Stam, C. EEG synchronization likelihood in mild cognitive impairment and Alzheimer’s disease during a working memory task. Clin. Neurophysiol. 2004, 115, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Wang, J.; Dai, Z.; Cao, X.; Jiang, L.; Fan, F.; Song, X.; Xia, M.; Shu, N.; Dong, Q.; et al. Topological organization of the human brain functional connectome across the lifespan. Dev. Cognit. Neurosci. 2013, 7, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Edelman, G.M.; Sporns, O. Complexity and coherency: Integrating information in the brain. Trends Cognit. Sci. 1998, 2, 474–484. [Google Scholar] [CrossRef]
- Li, J.; Yan, J.; Liu, X.; Ouyang, G. Using Permutation Entropy to Measure the Changes in EEG Signals During Absence Seizures. Entropy 2014, 16, 3049–3061. [Google Scholar] [CrossRef] [Green Version]
- Bandt, C.; Pompe, B. Permutation entropy: A natural complexity measure for time series. Phys. Rev. Lett. 2002, 88, 174102. [Google Scholar] [CrossRef] [PubMed]
- Staniek, M.; Lehnertz, K. Symbolic Transfer Entropy. Phys. Rev. Lett. 2008, 100, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Lempel, A.; Ziv, J. On the complexity of finite sequence. IEEE Trans. Inf. Theory 1976, 22, 75–81. [Google Scholar] [CrossRef]
- Bai, Y.; Liang, Z.; Li, X. A permutation Lempel-Ziv complexity measure for EEG analysis. Biomed. Signal Process. 2015, 19, 102–114. [Google Scholar] [CrossRef]
- Boytsova, Y.A.; Danko, S.G. EEG differences between resting states with eyes open and closed in darkness. Hum. Physiol. 2010, 36, 367–369. [Google Scholar] [CrossRef]
- Talebinejad, M.; Chan, A.; Miri, A. A Lempel-Ziv complexity measure for muscle fatigue estimation. J. Electromyogr. Kinesiol. 2011, 21, 236–241. [Google Scholar] [CrossRef] [PubMed]
- López-Sanz, D.; Bruña, R.; Garcés, P.; Martín-Buro, M.; Walter, S.; Delgado, M.; Montenegro, M.; Higes, R.L.; Marcos, A.; Maestú, F. Functional Connectivity Disruption in Subjective Cognitive Decline and Mild Cognitive Impairment: A Common Pattern of Alterations. Front. Aging Neurosci. 2017, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Woodward, N.; Cascio, C. Resting-state functional connectivity in psychiatric disorders. JAMA Psychiatry 2015, 78, 743–744. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Grossman, R.; Babb, J.; Rabin, M.; Mannon, L.; Kolson, D. Age-Related Total Gray Matter and White Matter Changes in Normal Adult Brain. Part I: Volumetric MR Imaging Analysis. AJNR Am. J. Neuroradiol. 2002, 23, 1327–1333. [Google Scholar] [PubMed]
- Peters, R. Ageing and the brain. Postgrad. Med. J. 2005, 82, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Schafer, C.B.; Morgan, B.R.; Ye, A.X.; Taylor, M.; Doesburg, S.M. Oscillations, networks and their development: MEG connectivity changes with age. Hum. Brain Mapp. 2014, 35, 5249–5261. [Google Scholar] [CrossRef] [PubMed]
- Oppenheim, A.; Schafer, R. Discrete-Time Signal Processing; Pearson: Boston, MA, USA, 2013. [Google Scholar]
- Choi, S.; Cichocki, A.; Park, H.; Lee, S. Blind Source Separation and Independent Component Analysis: A Review. Neural Inf. Proc. 2004, 6, 1–57. [Google Scholar]
- Goldenberg, D.; Galvan, A. The use of functional and effective connectivity techniques to understand the developing brain. Dev. Cognit. Neurosci. 2015, 12, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Gao, J.; Principe, J. Analysis of biomedical signals by the Lempel–Ziv complexity: The effect of finite data size. IEEE Trans. Bio-Med. Eng. 2006, 53, 2606–2609. [Google Scholar]
- Olofsen, E.; Sleigh, J.; Dahan, A. Permutation entropy of the electroencephalogram: A measure of anaesthetic drug effect. Br. J. Anaesth. 2008, 101, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Bian, C.; Qin, C.; Ma, Q.; Shen, Q. Modified permutation-entropy analysis of heartbeat dynamics. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2012, 85, 021906. [Google Scholar] [CrossRef] [PubMed]
- Popov, A.; Avilov, O.; Kanaykin, O. Permutation entropy of EEG signals for different sampling rate and time lag combinations Serock. In Proceedings of the 2013 Signal Processing Symposium (SPS), Serock, Poland, 5–7 June 2013. [Google Scholar]
- Rosazza, C.; Minati, L. Resting-state brain networks: Literature review and clinical applications. Neurol. Sci. 2011, 32, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O.; Thivierge, J.; Breakspear, M. Neurobiologically realistic determinants of self-organized criticality in networks of spiking neurons. PLoS Comput. Biol. 2011, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Ponce-Alvarez, A.; Mantini, D.; Romani, G.; Hagmann, P.; Corbetta, M. Resting-state functional connectivity emerges from structurally and dynamically shaped slow linear fluctuations. J. Neurosci. 2013, 33, 11239–11252. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. The economy of brain network organization. Nat. Rev. Neurosci. 2012, 13, 336–349. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Appendix 14: Summary of the systematic review on maintaining normal body temperature (normothermia). In Global Guidelines for the Prevention of Surgical Site Infection; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Ramaekers, D.; Ector, H.; Aubert, A.; Rubens, A.; van de Werf, F. Heart rate variability and heart rate in healthy volunteers. Is the female autonomic nervous system cardioprotective? Eur. Heart J. 1998, 19, 1334–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- JNC VI Joint National Committee on Prevention Detection, Evaluation and Treatment of High Blood Pressure. The sixth report of the Joint National Committee on Prevention, Detection, Evaluation and Treatment of High Blood Pressure. Arch. Intern. Med. 1997, 157, 2413–2444. [Google Scholar]
- Lebel, C.; Walker, L.; Leemans, A.; Phillips, L.; Beauli, C. Microstructural maturation of the human brain from childhood to adulthood. NeuroImage 2007, 40, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Compton, J.; van amelsvoort, T.; Murphy, D. HRT and its effect on normal ageing of the brain and dementia. Br. J. Clin. Pharmacol. 2001, 52, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Sharma, P. Aging of Brain: Role of Estrogen. Neurochem. Res. 2006, 31, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Marner, L.; Nyengaard, J.; Tang, Y.; Pakkenberg, B. Marked loss of myelinated nerve fibers in the human brain with age. J. Comp. Neurol. 2003, 462, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Azami, H.; Abasolo, D.; Simons, S.; Escudero, J. Univariate and Multivariate Generalized Multiscale Entropy to Characterise EEG Signals in Alzheimer’s Disease. Entropy 2017, 19, 31. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Age | Subjects | Category | ||
|---|---|---|---|---|---|
| Male | Female | Total | |||
| 1 | 21–40 | 34 | 35 | 69 | Early Adulthood (EA) |
| 2 | 41–60 | 21 | 16 | 37 | Mid adulthood (MA) |
| 3 | 61–80 | 24 | 47 | 71 | Late adulthood (LA) |
| Comparisons | m = 3 and m = 5 | m = 5 and m = 7 | m = 3 and m = 7 | |
|---|---|---|---|---|
| Group 1 | M | Correlation = 0.615 | Correlation = 0.906 | Correlation = 0.417 |
| p < 0.0001 | p < 0.0001 | p < 0.0001 | ||
| F | Correlation = 0.381 | Correlation = 0.907 | Correlation = 0.118 | |
| p < 0.0001 | p < 0.0001 | p = 0.152 | ||
| Group 2 | M | Correlation = 0.354 | Correlation = 0.768 | Correlation = −0.108 |
| p < 0.0001 | p < 0.0001 | p = 0.191 | ||
| F | Correlation = 0.577 | Correlation = 0.940 | Correlation = 0.451 | |
| p < 0.0001 | p < 0.0001 | p < 0.0001 | ||
| Group 3 | M | Correlation = 0.354 | Correlation = 0.768 | Correlation = −0.108 |
| p < 0.0001 | p < 0.0001 | p = 0.191 | ||
| F | Correlation = 0.507 | Correlation = 0.762 | Correlation = 0.193 | |
| p < 0.0001 | p < 0.0001 | p = 0.019 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shumbayawonda, E.; Tosun, P.D.; Fernández, A.; Hughes, M.P.; Abásolo, D. Complexity Changes in Brain Activity in Healthy Ageing: A Permutation Lempel-Ziv Complexity Study of Magnetoencephalograms. Entropy 2018, 20, 506. https://doi.org/10.3390/e20070506
Shumbayawonda E, Tosun PD, Fernández A, Hughes MP, Abásolo D. Complexity Changes in Brain Activity in Healthy Ageing: A Permutation Lempel-Ziv Complexity Study of Magnetoencephalograms. Entropy. 2018; 20(7):506. https://doi.org/10.3390/e20070506
Chicago/Turabian StyleShumbayawonda, Elizabeth, Pinar Deniz Tosun, Alberto Fernández, Michael Pycraft Hughes, and Daniel Abásolo. 2018. "Complexity Changes in Brain Activity in Healthy Ageing: A Permutation Lempel-Ziv Complexity Study of Magnetoencephalograms" Entropy 20, no. 7: 506. https://doi.org/10.3390/e20070506