
The First Genome from the Basal Monocot Family Has Been Misnamed: Taxonomic Identity of Acorus tatarinowii (Acoraceae), a Source of Numerous Chemical Compounds of Pharmaceutical Importance
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphology and Anatomy
2.2. Nomenclatural Analysis
3. Morphological Differences between the Acorus calamus Group and the A. gramineus Group
4. Nomenclatural Treatment of Acorus tatarinowii
4.1. Formal Nomenclature
4.2. Nomenclatural History
4.3. Original Material
4.4. Nomenclatural Discussion
5. Taxonomic Identity of Acorus tatarinowii
6. The Concept of Three Narrowly Defined Species in the A. gramineus Group Is Still Insufficiently Justified
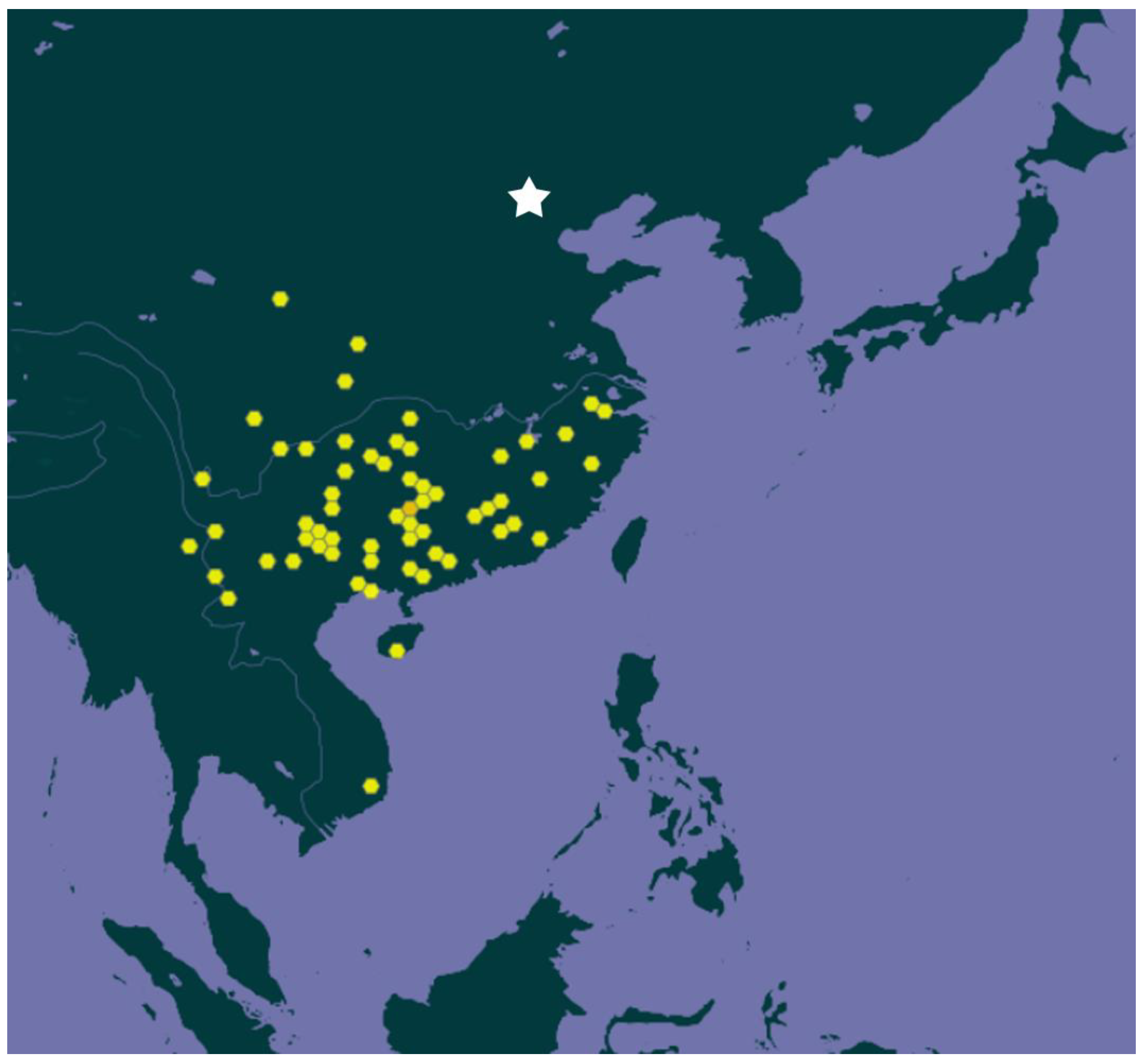
7. Distribution of Acorus in Eastern Indochina and Thailand
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code): Adopted by the Nineteenth International Botanical Congress, Shenzhen, China, July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; Stevens, P.F.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Bogner, J.; Mayo, S.J. Acoraceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin, Germany, 1998; Volume 4, pp. 7–11. [Google Scholar]
- Schott, H.W. Prodromus Systematis Aroidearum; Typis Congregationis Mechitharisticae: Vienna, Austria, 1860. [Google Scholar]
- Schott, H.W. Araceae. Pars altera. Nova genera et species, adiectis quibusdam adnotationibus. Ann. Mus. Bot. Lugduno-Batavi 1864, 1, 278–286. [Google Scholar]
- Engler, A. Araceae-Pothoideae. In Das Pflanzenreich, 21 (IV. 23B); Engler, A., Ed.; Engelmann: Leipzig, Germany, 1905; pp. 1–330. [Google Scholar]
- Li, H.; Zhu, G.; Bogner, J. Acoraceae. In Flora of China, Volume 2; Wu, Z., Raven, P., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2010; pp. 1–2. [Google Scholar]
- Bogner, J. Acoraceae. In Flora Malesiana, Series 1: Volume 20; Noteboom, H.P., Ed.; Nationaal Herbarium Nederland: Leiden, The Netherlands, 2011; pp. 1–13. [Google Scholar]
- Boyce, P. Acoraceae. In Flora of Thailand, Volume 11, Part 2; Santisuk, T., Larsen, K., Eds.; The Forest Herbarium, Department of National Parks, Wildlife and Plant Conservation: Bangkok, Thailand, 2012; pp. 322–325. [Google Scholar]
- Ryuk, J.A.; Kim, Y.S.; Lee, H.W.; Ko, B.S. Identification of Acorus gramineus, A. calamus, and A. tatarinowii using sequence characterized amplified regions (SCAR) primers for monitoring of Acori graminei rhizoma in Korean markets. Int. J. Clin. Exp. Med. 2014, 7, 2488. [Google Scholar] [PubMed]
- Xu, Z.; Chang, L. (Eds.) Acoraceae. In Identification and Control of Common Weeds; Springer: Singapore, 2017; Volume 3, pp. 787–789. [Google Scholar] [CrossRef]
- Kovtonyuk, N.K. Araceae. In Flora Sibiriae, Araceae–Orchidaceae; Malyshev, L.I., Peshkova, G.A., Eds.; Nauka: Novosibirsk, Russia, 1987; p. 14. (In Russian) [Google Scholar]
- Tzvelev, N.N. Araceae. In Plantae vasculares Orientis Extremi Sovietici, T. 8; Charkevicz, S.S., Ed.; Nauka: St.-Petersburg, Russia, 1996; pp. 357–364. (In Russian) [Google Scholar]
- Vinogradova, Y.K. Variability of Acorus calamus L. biological characteristics in natural and invasive populations. Byull. Glavn. Bot. Sada 2004, 187, 23–31. (In Russian) [Google Scholar]
- Chinov, P.S. (Ed.) Atlas of Distribution Ranges and Resources of Medicinal Plants of the USSR; Medgiz: Moscow, Russia, 1983. (In Russian) [Google Scholar]
- Sokoloff, D.D.; Skaptsov, M.V.; Vislobokov, N.A.; Smirnov, S.V.; Shmakov, A.I.; Remizowa, M.V. Morphological characterization of diploid and triploid Acorus calamus (Acoraceae) from southern Western Siberia, parthenocarpy in sterile plants and occurrence of aneuploidy. Bot. J. Linn. Soc. 2021, 195, 189–215. [Google Scholar] [CrossRef]
- Mücke, M. Über den Bau und die Entwicklung der Früchte und über die Herkunft von Acorus calamus L. Bot. Zeit. 1908, 66, 1–23. [Google Scholar]
- Packer, J.G.; Ringius, G.S. The distribution and status of Acorus (Araceae) in Canada. Can. J. Bot. 1984, 62, 2248–2252. [Google Scholar] [CrossRef]
- Thompson, S.A. Systematics and biology of the Araceae and Acoraceae of temperate North America. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 1995. [Google Scholar]
- Rajput, S.B.; Tonge, M.B.; Karuppayil, S.M. An overview on traditional uses and pharmacological profile of Acorus calamus Linn. (Sweet flag) and other Acorus species. Phytomedicine 2014, 21, 268–276. [Google Scholar] [CrossRef]
- Shu, H.; Zhang, S.; Ji, Y.; Luo, B.; Long, C. Acoraceae plants, potential ornamentals. Sb. Nauchnykh Tr. GNBS 2017, 145, 114–118. [Google Scholar]
- Gagnepain, F. Aracées. In Flore Générale de l’Indo-Chine, T. 6; Lecomte, H., Ed.; Masson: Paris, France, 1908; pp. 1075–1196. [Google Scholar]
- Li, H. Acorus. In Flora Reipublicae Popularis Sinicae Volume 13, 2; Wu, C., Li, H., Eds.; Science Press: Beijing, China, 1979; pp. 4–9. (In Chinese) [Google Scholar]
- Wei, F.N.; Li, Y.K. A new spice, Acorus macrospadiceus from south China. Guihaia 1985, 5, 179–182. (In Chinese) [Google Scholar]
- Ho, P.H. An Illustrated Flora of Vietnam, Volume 3; Nha Xuat Ban Tre: Ho Chi Minh, Vietnam, 2000; pp. 1–1020. (In Vietnamese) [Google Scholar]
- Shu, H.; Zhang, S.; Lei, Q.; Zhou, J.; Ji, Y.; Luo, B.; Hong, L.; Li, F.; Liu, B.; Long, C. Ethnobotany of Acorus in China. Acta Soc. Bot. Polon. 2018, 87, 3585. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Shu, H.; Zhang, S.; Luo, B.; Gu, R.; Zhang, R.; Ji, Y.; Li, F.; Long, C. From folk taxonomy to species confirmation of Acorus (Acoraceae): Evidences based on phylogenetic and metabolomic analyses. Front. Plant Sci. 2020, 11, 965. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z. Some new taxa of Acorus (Araceae) from Sichuan. Acta Bot. Bor.-Occ. Sinica 1985, 5, 118–121. (In Chinese) [Google Scholar]
- Liu, K. The studies of morphological variations and structure of the leaves on the six Acorus species. Acta Sci. Nat. Univ. Norm. Hunan 1990, 13, 146–152. (In Chinese) [Google Scholar]
- Zhang, W. Pharmacognostic study of Acorus—From a phylogenetic perspective. Ph.D. Thesis, Hong Kong Baptist University, Hong Kong, China, 2005. [Google Scholar]
- Yang, R.; Yan, Z.; Chen, C.; Liu, J. Constituents and activities of Acorus tatarinowii. Med. Res. Arch. 2017, 5, 1–14. [Google Scholar] [CrossRef]
- Lu, Y.; Xue, Y.; Chen, S.; Zhu, H.; Zhang, J.; Li, X.N.; Wang, J.; Liu, J.; Qi, C.; Du, G.; et al. Antioxidant lignans and neolignans from Acorus tatarinowii. Sci. Rep. 2016, 6, 22909. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Xue, Y.; Liu, J.; Yao, G.; Li, D.; Sun, B.; Zhang, J.; Liu, Y.; Qi, C.; Xiang, M.; et al. (±)-Acortatarinowins A–F, norlignan, neolignan, and lignan enantiomers from Acorus tatarinowii. J. Nat. Prod. 2015, 78, 2205–2214. [Google Scholar] [CrossRef]
- Tong, X.G.; Zhou, L.L.; Wang, Y.H.; Xia, C.; Wang, Y.; Liang, M.; Hou, F.F.; Cheng, Y.X. Acortatarins A and B, two novel antioxidative spiroalkaloids with a naturally unusual morpholine motif from Acorus tatarinowii. Org. Lett. 2010, 12, 1844–1847. [Google Scholar] [CrossRef]
- Hu, J.; Feng, X. Phenylpropanes from Acorus tatarinowii. Planta Med. 2000, 66, 662–664. [Google Scholar] [CrossRef]
- Wang, Z.J.; Zhu, Y.Y.; Yi, X.; Zhou, Z.S.; He, Y.J.; Zhou, Y.; Qi, Z.H.; Jin, D.N.; Zhao, L.X.; Luo, X.D. Bioguided isolation, identification and activity evaluation of antifungal compounds from Acorus tatarinowii Schott. J. Ethnopharmacol. 2020, 261, 113119. [Google Scholar] [CrossRef]
- Deng, C.; Li, N.; Zhang, X. Rapid determination of essential oil in Acorus tatarinowii Schott. by pressurized hot water extraction followed by solid-phase microextraction and gas chromatography–mass spectrometry. J. Chromatogr. A 2004, 1059, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Ali, I.; Hussain, H.; Hussain, M.; Wang, X.B.; Song, X.; Luo, G.; Zhang, Z.; Wang, Z.; Wang, D. Extraction and purification of cis/trans asarone from Acorus tatarinowii Schott: Accelerated solvent extraction and silver ion coordination high-speed counter-current chromatography. J. Chromatogr. A 2021, 1643, 462080. [Google Scholar] [CrossRef] [PubMed]
- Gao, E.; Zhou, Z.Q.; Zou, J.; Yu, Y.; Feng, X.L.; Chen, G.D.; He, R.R.; Yao, X.S.; Gao, H. Bioactive asarone-derived phenylpropanoids from the rhizome of Acorus tatarinowii Schott. J. Nat. Prod. 2017, 80, 2923–2929. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Mahady, G.; Qian, Y.; Song, P.; Jian, T.; Ding, X.; Guan, F.; Shan, Y.; Wei, M. The essential oil from Acori Tatarinowii Rhizome (the dried rhizome of Acorus tatarinowii Schott) prevents hydrogen peroxide-induced cell injury in PC12 cells: A signaling triggered by CREB/PGC-1α activation. Evid. Based Complement. Alternat. Med. 2020, 2020, 4845028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, Y.; Liang, Z.; Ho, A.; Chen, H.; Zhao, Z. Metabolite profiling of tissues of Acorus calamus and Acorus tatarinowii rhizomes by using LMD, UHPLC-QTOF MS, and GC-MS. Planta Med. 2015, 81, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Ni, G.; Shen, Z.F.; Lu, Y.; Wang, Y.H.; Tang, Y.B.; Chen, R.Y.; Hao, Z.Y.; Yu, D.Q. Glucokinase-activating sesquinlignans from the rhizomes of Acorus tatarinowii Schott. J. Org. Chem. 2011, 76, 2056–2061. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Bauer, R.; Melchart, D.; Xiao, P.-G.; Staudinger, A. (Eds.) Chromatographic fingerprint analysis of herbal medicines. In Thin-Layer and High Performance Liquid Chromatography of Chinese Drugs; Springer: Vienna, Austria, 2011; Volume 1. [Google Scholar]
- Han, P.; Han, T.; Peng, W.; Wang, X.R. Antidepressant-like effects of essential oil and asarone, a major essential oil component from the rhizome of Acorus tatarinowii. Pharm. Biol. 2013, 51, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.P.; Chen, L.; Yi, Y.H.; Sun, W.W.; Gao, M.M.; Su, T.; Yang, S.Q. Study of antiepileptic effect of extracts from Acorus tatarinowii Schott. Epilepsia 2005, 46, 21–24. [Google Scholar] [CrossRef]
- Shi, T.; Huneau, C.; Zhang, Y.; Li, Y.; Chen, J.; Salse, J.; Wang, Q. The slow-evolving Acorus tatarinowii genome sheds light on ancestral monocot evolution. Nature Plants 2022, 8, 764–777. [Google Scholar] [CrossRef]
- Aiton, W. Hortus Kewensis, or a Catalogue of the Plants Cultivated in the Royal Botanic Garden at Kew; George Nicol: London, UK, 1789; Volume 1. [Google Scholar]
- Schott. Aroideen-Skizzen. Oesterr. Bot. Z. 1859, 9, 98–101. [Google Scholar] [CrossRef]
- Ruprecht, F.J. An essay on the history of the Botanical Museum, Imperial Academy of Sciences. Zap. Imp. Akad. Nauk. 1864, 5, 139–162. (In Russian) [Google Scholar]
- Trautvetter, R.E. A brief overview of the history of the Imperial Botanical Garden in St.-Petersburg. Tr. Imp. S. Peterbg. Bot. Sada 1873, 2, 145–304. (In Russian) [Google Scholar]
- Bretschneider, E. History of European Botanical Discoveries in China; Sampson Low, Marston and Company: London, UK, 1898; Volumes 1 and 2. [Google Scholar]
- Lipsky, V.I. Herbarium of the Imperial Botanical Garden, 2nd ed.; Mattisen: Yuriev, Russia, 1908. (In Russian) [Google Scholar]
- Lipsky, V.I. Biographies and publications of botanists employed at and other persons relevant to the Imperial Botanical Garden. In Imperial Botanical Garden: 200 years of its history (1713–1913); Fischer von Waldheim, A.A., Ed.; Imperial Botanical Garden: St.-Petersburg, Russia, 1913; Volume 3, pp. 1–378. (In Russian) [Google Scholar]
- Lipschitz, S.Y.; Vasilchenko, I.T. The Central Herbarium of the USSR: A Historical Account; Nauka: St.-Petersburg, Russia, 1968. (In Russian) [Google Scholar]
- Sennikov, A.N. The history and nomenclatural significance of herbarium collections made by Alexander A. Tatarinov in North China and Mongolia in 1841–1850. Taxon, 2023; submitted. [Google Scholar]
- Kaplan, D.R. Comparative foliar histogenesis in Acorus calamus and its bearing on the phyllode theory of monocotyledonous leaves. Amer. J. Bot. 1970, 57, 331–361. [Google Scholar] [CrossRef]
- Thompson, S.A. Acoraceae. In Flora of North America North of Mexico, Volume 22. Magnoliophyta: Alismatidae, Arecidae, Commelinidae (in Part), and Zingiberidae; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2000; pp. 124–127. [Google Scholar]
- Keating, R.C. The Anatomy of the Monocotyledons: Volume IX: Acoraceae and Araceae; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Stafleu, F.A.; Cowan, R.S. Taxonomic Literature. A Selective Guide to Botanical Publications and Collections with Dates, Commentaries and Types, Volume 5; Bohn, Scheltema & Holkema; Dr. W. Junk b.v., Publishers: Utrecht, The Netherlands; The Hague, The Netherlands; Boston, MA, USA, 1985. [Google Scholar]
- Grubov, V.I. Plants of Central Asia, Volume 1; Academy of Sciences of the USSR: St. Petersburg, Russia, 1963. (In Russian) [Google Scholar]
- Schott, H.W. Aroideen-Skizzen. Oesterr. Bot. Wochenbl. 1857, 7, 341–342. [Google Scholar] [CrossRef]
- Schott, H.W. Aroideenskizzen. Bonplandia 1859, 7, 26–31. [Google Scholar] [CrossRef]
- Bunge, A. Enumeratio plantarum quas in China boreali collegit Dr. Al. Bunge. Mém. Acad. Imp. Sci. St. Pétersbourg Divers Savans 1833, 2, 75–148. [Google Scholar]
- Ma, L.; Jiang, S.-Z.; Lian, H.; Xiong, Y.-F.; Liu, Z.-J.; Chen, S.-P. The complete chloroplast genome sequence of Acorus tatarinowii (Araceae) from Fujian, China. Mitochondrial DNA Part B 2020, 5, 3159–3160. [Google Scholar] [CrossRef]
- Chen, J.; Hao, W.; Zhang, C.; Cui, M.; Sun, Y.; Zhang, Y.; Wang, J.; Mou, T.T.; Gu, S.; Zhao, T.T.; et al. Explore the therapeutic composition and mechanism of Schisandra chinensis-Acorus tatarinowii Schott on Alzheimer’s disease by using an integrated approach on chemical profile, network pharmacology, and UPLC-QTOF/MS-based metabolomics analysis. Oxid. Med. Cell. Longev. 2022, 2022, 6362617. [Google Scholar] [CrossRef]
- Hao, Z.Y.; Ni, G.; Liang, D.; Liu, Y.F.; Zhang, C.L.; Wang, Y.; Zhang, Q.J.; Chen, R.Y.; Yu, D.Q. A New brominated norsesquiterpene glycoside from the rhizomes of Acorus tatarinowii Schott. Nat. Prod. Commun. 2021, 16, 1–6. [Google Scholar] [CrossRef]
- Yamamoto, Y. Contributiones ad floram Kainanensem (‘Kainanensis’) I. Kainan Kaigun Tokumubu Seimukyoku: Taihoku, Japan, 1943. [Google Scholar]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2022. Available online: http://www.plantsoftheworldonline.org/ (accessed on 18 November 2022).
- GBIF. Global Biodiversity Information Facility. 2022. Available online: https://www.gbif.org/ (accessed on 18 November 2022).
- Newman, M.; Ketphanh, S.; Svengsuksa, B.; Thomas, P.; Sengdala, K.; Lamxay, V.; Armstrong, K. A Checklist of the Vascular Plants of Lao PDR; Royal Botanic Garden Edinburgh: Edinburgh, UK, 2007. [Google Scholar]
- Cho, S.H.; Chhang, P.; Kim, Y.D. A Checklist for the Seed Plants of Cambodia; National Institute of Biological Resources, Ministry of Environment: Incheon, Korea, 2016. [Google Scholar]
- Ito, M.Y.; Barfod, M.A.S. An updated checklist of aquatic plants of Myanmar and Thailand. Biodivers. Data J. 2014, 2, e1019. [Google Scholar] [CrossRef] [PubMed]
- Murata, J. Acoraceae. In Flora of Japan, Volume IVb; Iwatsuki, K., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 2016; p. 5. [Google Scholar]
- Wei, Y.G.; Do, T.V.; Wen, F. A Checklist to the Plants of Northern Vietnam; China Forestry Publishing House: Beijing, China, 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
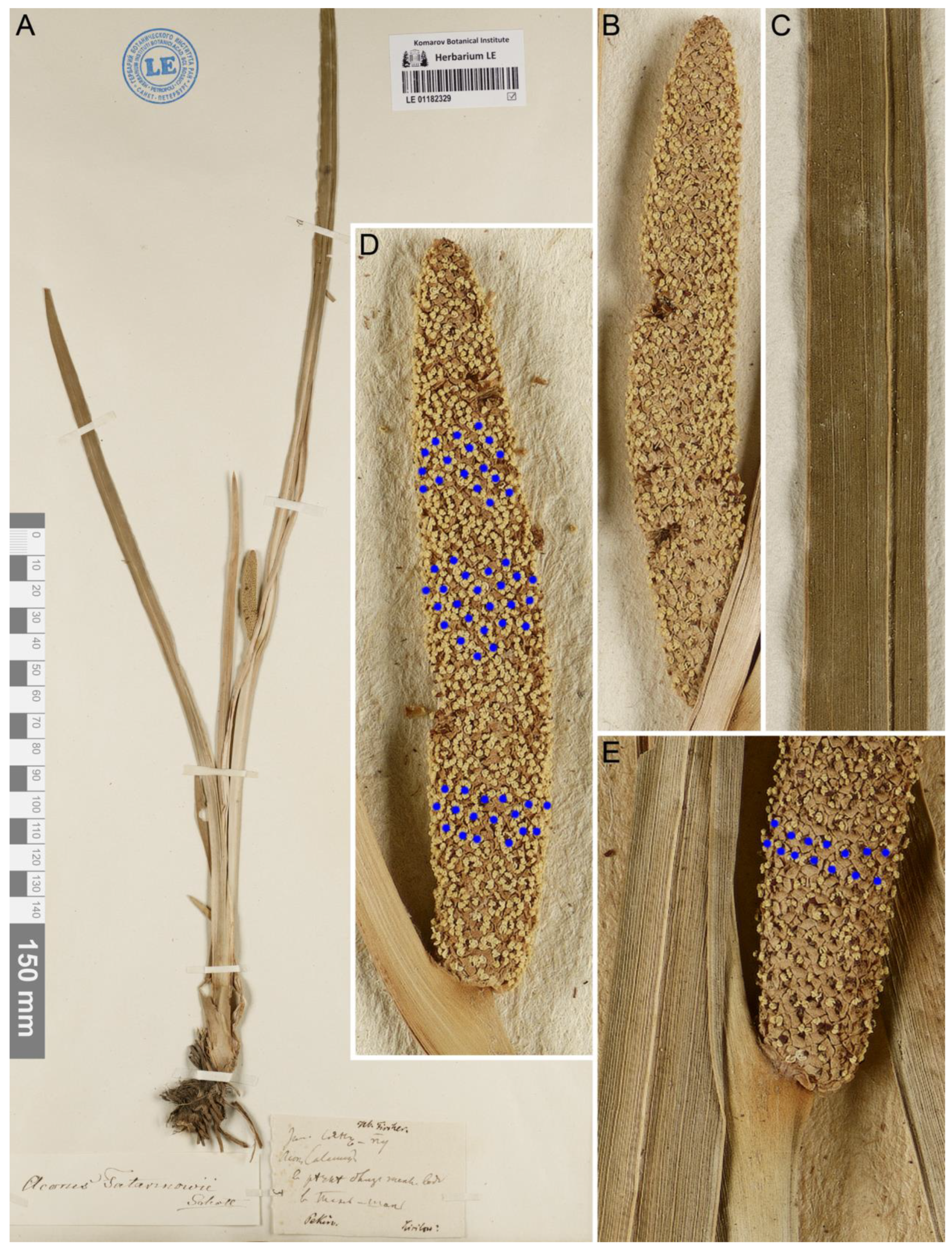
| Specimen Barcode | Script Verbatim | Translation or Transcription | Authority of Handwriting | Notes |
|---|---|---|---|---|
| LE 01182325 | Acorus Calamus L. (кит. cянъ-пy) Iюнь. близъ тeплыxъ вoдъ, въ 25 вepcтaxъ oтъ Пeкинa | Acorus Calamus L. (Changpu in Chinese) June, near hot springs, 25 versts [=26.5 km] from Beijing | unknown copyist | original label |
| Kиpилoв | P.Ye. Kirilov | V.L. Komarov | erroneous collector’s name, curatorial annotation | |
| LE 01182326 | non remitt[endum] | should not be given away | F.J. Ruprecht | curatorial annotation, applied by Ruprecht to most important specimens which cannot be used for exchange |
| Acorus Calamus var.? | F.J. Ruprecht | curatorial annotation | ||
| Herb. Acad. Petrop. Fl. Pekin. Dr. Tatarinow, acc. 1856. | Herbarium of the Imperial Academy of Sciences in St.-Petersburg Flora of Beijing Dr. A.A. Tatarinov, acquired in 1856 | printed curatorial label of the Botanical Museum, with collecting area, collector’s name, and accession year | ||
| LE 01182327 | Acorus Calamus L. | A.A. Tatarinov | taxonomic revision, identification label | |
| Herb. Acad. Petrop. Fl. Pekin. Dr. Tatarinow, acc. 1856. | Herbarium of the Imperial Academy of Sciences in St.-Petersburg Flora of Beijing Dr. A.A. Tatarinov, acquired in 1856 | printed curatorial label of the Botanical Museum, with collecting area, collector’s name, and accession year | ||
| cянъ-пy. Iюнь–в pѣчкѣ близъ тeплыx[ъ] вoдъ въ Taнъ-Шaнѣ | Changpu. June–in a small river near hot springs in Tangshan | A.A. Tatarinov | original label | |
| LE 01182329 | Acorus Tatarinowii Schott | H.W. Schott | taxonomic revision, identification label | |
| Hb. Fischer. Pekin. Kirilow? | Herbarium of F.E.L. von Fischer [Collected from the vicinity of] Beijing. By P.Ye. Kirilov? | F. Körnicke | curatorial annotation, Herbarium Fischer accession to the Botanical Garden in 1855 | |
| Jun. cянъ-пy Acorus Calamus? в pѣчкѣ близъ тeпл[ыxъ] вoдъ в[ъ] Taнъ-Шaнѣ | June. Changpu. Acorus calamus? In a small river near hot springs in Tangshan | A.A. Tatarinov | original label |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokoloff, D.D.; Remizowa, M.V.; Nuraliev, M.S.; Averyanov, L.V.; Sennikov, A.N. The First Genome from the Basal Monocot Family Has Been Misnamed: Taxonomic Identity of Acorus tatarinowii (Acoraceae), a Source of Numerous Chemical Compounds of Pharmaceutical Importance. Diversity 2023, 15, 176. https://doi.org/10.3390/d15020176
Sokoloff DD, Remizowa MV, Nuraliev MS, Averyanov LV, Sennikov AN. The First Genome from the Basal Monocot Family Has Been Misnamed: Taxonomic Identity of Acorus tatarinowii (Acoraceae), a Source of Numerous Chemical Compounds of Pharmaceutical Importance. Diversity. 2023; 15(2):176. https://doi.org/10.3390/d15020176
Chicago/Turabian StyleSokoloff, Dmitry D., Margarita V. Remizowa, Maxim S. Nuraliev, Leonid V. Averyanov, and Alexander N. Sennikov. 2023. "The First Genome from the Basal Monocot Family Has Been Misnamed: Taxonomic Identity of Acorus tatarinowii (Acoraceae), a Source of Numerous Chemical Compounds of Pharmaceutical Importance" Diversity 15, no. 2: 176. https://doi.org/10.3390/d15020176