Two New Benthic Diatoms of the Genus Achnanthidium (Bacillariophyceae) from the Hangang River, Korea


1
Department of Environmental Science, Hanyang University, Seoul 04763, Korea
2
Biological Resource Center/Korean Collection for Type Cultures (KCTC), Korea Research Institute of Bioscience and Biotechnology, Jeongeup 56212, Korea
3
National Institute of Environmental Research, Incheon 22689, Korea
4
Department of Life Science and Research Institute for Natural Sciences, Hanyang University, Seoul 04763, Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally as co-first authors.
https://www.ipni.org/urn:lsid:ipni.org:authors:20045093-1
Diversity 2020, 12(7), 285; https://doi.org/10.3390/d12070285
Submission received: 25 June 2020
/
Revised: 17 July 2020
/
Accepted: 20 July 2020
/
Published: 21 July 2020
(This article belongs to the Special Issue Taxonomy, Ecology and Biogeography of Diatoms)
Abstract
:Two new benthic freshwater species belonging to the genus Achnanthidium were found in Korea. Achnanthidium ovale sp. nov. and A. cavitatum sp. nov. are described as new species based on light and scanning electron microscopy observations and molecular analyses. Both species are compared with the type material of morphologically similar taxa. Achnanthidium ovale differs from other species belonging to the A. pyrenaicum complex in outline, striation pattern, raphe central endings, and freestanding areolae at the apices. Achnanthidium cavitatum differs from other species in the A. minutissimum complex in outline, broad axial central area in the raphel ess valve, and slit-like areolae near the axial central area. We assessed their molecular characteristics by analyzing nuclear small subunit (SSU) rRNA and chloroplast-encoded rbcL gene sequences. Both the morphological comparison and the SSU and rbcL sequence analyses provide strong evidence to support the recognition of A. ovale and A. cavitatum as new species.
1. Introduction
The genus Achnanthidium Kütz. has been considered a subgenus of Achnanthes Bory [1]. Round et al. [2] restituted the genus rank to Achnanthidium and distinguished Achnanthes from Achnanthidium based on the areola, raphe, girdle, and plastid characteristics. Czarnecki [3] then transferred Achnanthes minutissima Kütz. to Achnanthidium as A. minutissimum (Kütz.) Czarn. The genus Achnanthidium was then redefined by Round and Bukhtiyarova [4].
The genus Achnanthidium currently includes freshwater monoraphid species with the following characteristics: (1) linear-lanceolate to lanceolate elliptic cells with length and width less than 30 μm and 5 μm, respectively, (2) concave raphe valve, uniseriate striae, and a wide central area; (3) a well-developed raphe that can be straight or turned to one side [4]. Because of their small size and inadequate morphological features, Achnanthidium species can be complicated to identify. Currently, species in the genus Achnanthidium can be divided into three major groups: (1) the A. minutissimum complex, which includes species with straight raphe fissures on the apical area; (2) the A. pyrenaicum complex, with species with hooked terminal raphe fissures; (3) the A. exiguum complex, with species that have terminal raphe endings deflecting to opposites sides [5,6,7].
Some new Achnanthidium species have been recently reported: A. sieminskae [8], A. barbei [9], and A. costei [9] in the A. minutissimum complex; A. rivulare by Potapova and Ponader [10], A. hoffmannii [11], and A. delmontii [12] in the A. pyrenaicum complex; A. initium [13] in the A. exiguum complex.
Species of the genus Achnanthidium have been reported to live in a wide variety of ecological conditions, from oligotrophic to eutrophic and from alkaline to acidic environments [14]. Species of Achnanthidium respond differently to water chemistry conditions. Therefore, species-level identification can produce more accurate bioassessments [15]. Because Achnanthidium species are both common and abundant, it is very important to identify the ecological environment of each species and propose an adequate classification of all the Achnanthidium species [16].
Diatom valves are perforated by areolae (pores) that allow communication between the diatom’s protoplast and the surrounding environment [17]. The development of internal hymenes (cf. Cox [18]) is the final stage in valve morphogenesis [19]. Areolae have two types of ultrastructure: poroid areolae are not markedly constricted on one surface of the valve, whereas loculate areolae are markedly constricted on one surface and occluded on the other [19]. Ross et al. [20] defined loculate areolae as a regular perforation through the basal siliceous valve, usually occluded by a velum (cribrum, rota, vola) or a rica. Mann [21] refined the definition of pore occlusion as a delicate siliceous membrane that crosses the entire pore, called a hymen (=rica of Ross et al. [20]). Cox [22] considered some structural characteristics of diatoms are highly consistent with raphid taxa—e.g., raphe construction, type of pore occlusion (hymenes, cribrum, etc.)—Whereas other characteristics, such as pore shape, degree of surface silicification, and raphe fissure path, could be more susceptible to variation. Yana and Mayama [23] proposed that, from the internal view, the areolae of Achnanthidium pseudoconspicuum var. yomensis Yana & Mayama [23] are loculate with incomplete vimines. Two types of areolae were detected on the valve face: round (various sizes in different taxa) and slit-like, which always appeared close to the valve margin in A. catenatum, A. dolomiticum, and A. saprophilum [24].
Sequences of a variety of genes such as ITS, COI, large subunit rRNA, small subunit rRNA (SSU), and ribulose-1, 5-bisphosphate carboxylase/oxygenase (rbcL), are currently being used to propose phylogenetic relationships among diatoms [25]. Of them, previous studies have shown that SSU is a powerful tool for inferring phylogenetic relationships at all taxonomic levels [26]. SSU rRNA gene sequences have been widely used to represent the major diatom lineages [27,28,29,30,31,32]. The rbcL gene is located in a single-copy region of the chloroplast genome [25,33]. Because it only rarely has insertions or deletions, the rbcL gene is considered better than the SSU gene for studies of diatom evolution from order to genus levels [34].
The sequences of some Achnanthidiaceae genera were not added to molecular phylogenetic datasets until 2016 [35]. Round et al. [2] described the family Achnanthidiaceae to include two genera, Achnanthidium and Eucocconeis Cleve ex Meister. However, according to Kulikovskiy et al. [35], genera such as Rossithidium Round & Bukhtiyarova, Psammothidium Bukhtiyarova & Round, and Lemnicola Round & Basson should also be included in this family. The taxonomy of achnanthoid diatoms has mostly been based on morphological data, and molecular studies with the SSU rRNA and rbcL genes should also be performed [36].
As part of documenting the distribution of diatoms from oligotrophic environments in Korea, we report the morphological characteristics of two Achnanthidium species and compare nuclear-encoded SSU rRNA and chloroplast-encoded rbcL gene sequences with those of related species. From those results, we describe two new species, Achnanthidium ovale sp. nov. and Achnanthidium cavitatum sp. nov. Upon addition of the two new species, 169 Achnanthidium species are known globally and 22 in Korea (AlgaeBase & NIBR).
2. Materials and Methods
2.1. Sample Collection, Isolation, and Culture
To collect diatoms, two or three pebbles were collected from the littoral zones (0.1 m depth) of rivers. The sampling points at which diatoms were collected are shown in Table 1 and located in the Hangang River, Republic of Korea (Figure 1). Epilithon was collected from the surfaces of the stones using a toothbrush. Single diatom cells were isolated using a Pasteur pipette (Hilgenberg GmbH, Germany) and the capillary method [37] under an Olympus CKX41 inverted microscope (Olympus, Tokyo, Japan). Cells were isolated and cultured in 96-well cell plates, and each well contained 160 μL of Diatom Medium (DM) [38]. After 10–14 days of isolation, diatoms reached the exponential growth stage [39]. The cells that grew and had a healthy aspect were transferred into 24-well cell plates with 1 mL of DM. Again, after 10–14 days, the cells that were in good condition were transferred to 50 cm3 culture flasks with 20 mL of DM. To maintain healthy cells, each strain was sub-cultured at 40-day intervals. All the strains were cultured at 20 °C, with an irradiance of c. 50 μmol quanta m−2 s−1, and a 12:12 h light: dark cycle with cool white fluorescent light. Two new diatom cultures were eventually established; of these, one culture was used for this study. The other two cultures were preserved at a lower temperature (<10 °C) and light intensity (<20 μmol m−2 s−1) for growth limitation.
2.2. Light Microscopy (LM)
For LM, both natural and cultured cells were fixed with Lugol solution. To remove organic compounds, HNO3 and H2SO4 (1:3) were added to the samples, which were then boiled at 100 °C for 2–3 min. To remove the acid from the oxidized cultures, the samples were washed four times with distilled water, following one day of sedimentation. Morphological characteristics were observed using an upright microscope (Nikon E600, Nikon, Tokyo, Japan). Slides of the washed frustules were mounted using Wako Mountmedia (Wako Pure Chemical Industries, Ltd., Osaka, Japan). Light micrographs were collected at 1000× magnification using an MSC-C5.0 microscope digital camera (SONY, Tokyo, Japan). Measurements of length and width of frustules were obtained from at least 50 diatom cells.
2.3. Scanning Electron Microscopy (SEM)
For SEM, the washed samples were gently filtered through a 0.2-μm pore-sized GTTP Millipore filter membrane (Millipore Filter Corporation, Cork, Ireland) using gravity. The membrane was then stuck to the SEM stubs with carbon tape (Shintron Enterprise CO., Ltd., Kaohsiung, Taiwan). Mounted specimens were dried for at least 12 h at room temperature. Finally, the specimens were coated for 120 s with platinum and examined using field emission SEM (Nova Nano SEM 450, FEI Inc., Hillsboro, OR, USA).
2.4. DNA Extraction, PCR Amplification, and Sequencing
Clonal cultures (10 mL) were prepared in the mid-logarithmic growth phase and centrifuged in a conical tube at 4000× g for 10 min. A DNeasy Plant Mini Kit (Qiagen, Valencia, CA, USA) was used for genomic DNA extraction. PCR reactions were performed in 40 μL reaction mixtures, and the primers used in the PCR amplification of SSU and rbcL genes are shown in Table 2. Each reaction mixture contained 23.8 μL of distilled water, 4 μL of 10× Ex PCR Buffer (TaKaRa, Tokyo, Japan), 4 μL of dNTP (deoxyribonucleotide triphosphate) (TaKaRa), 0.2 μL of ExTaq polymerase (TaKaRa), 2 μL of each primer, and 4 μL of DNA template. PCR amplification was carried out in a Bio-Rad iCycler (Bio-Rad, Hercules, CA, USA) using the following conditions: pre-denaturation at 94 °C for 4 min; 37 cycles at 94 °C for 20 s, 56 °C for 30 s, and 72 °C for 50 s; and a final extension at 72 °C for 5 min. The PCR products were separated by electrophoresis in 1% agarose gel with a staining solution (Genetics, Dueren, Germany) and then sent to the company BIONICS (ISO: 9001, Seoul, Korea) for SSU rRNA and rbcL gene sequencing.
2.5. Phylogenetic Analyses
Sequences were viewed and assembled in ContigExpress (Vector NTI version 1.6, Invitrogen, Grand Island, NY, USA). The SSU rRNA and rbcL sequences from this study were deposited in the National Center for Biotechnology Information (NCBI) GenBank (Table 3). Multiple sequence alignment between the sequences generated in this study and those obtained from the NCBI database was performed using ClustalW [41] in MEGA version 7.0 [42]. The alignments were manually edited, and ambiguously aligned characters were excluded using MEGA version 7.0 [42]. MEGA 7.0 was also used to calculate the genetic distance (p-distance) by means of a bootstrap method with 1000 replicates and a Kimura 2-parameter model [42]. The final alignment of the SSU rDNA dataset contained 39 taxa and 1628 characters (including gaps introduced for alignment), and the rbcL dataset contained 52 taxa and 1628 characters (1390 bp). Sequences of Aulacoseira granulata were used as outgroups for the SSU and rbcL phylogenetic trees.
Phylogenetic trees for the sequence alignments (SSU and rbcL) were inferred from maximum likelihood (ML) analyses (using RaxML version 8 [43]) and Bayesian inference (using MrBayes version 3.2: [44]). The general time-reversible model with parameters accounting for γ-distributed rate variation across sites was used in all analyses, taking into account a 6-class gamma. Bootstrap analyses for both datasets were carried out for ML with 1000 replicates to evaluate statistical reliability. The Markov chain Monte Carlo method was used with four runs for 10 million generations, sampling every 100 generations. A majority-rule consensus tree was created to examine the posterior probabilities of each clade. The final trees were visualized with MEGA version 7.0.
3. Results
3.1. Species Description
3.1.1. Achnanthidium ovale M. Miao & B.-H. Kim, sp. nov.
Description: Cells are elliptical, 6.3–7.7 µm long, and 3.8–4.1 µm wide. Striae density varies by location; 30–35 in 10 µm in the center, and up to 55 near the apices of the raphe valve. The number of striae on the primary side is higher than on the secondary side of the rapheless and raphe valves. Therefore, a “T” pattern can be seen in the LM images (Figure 2B–O).
At the external part of the raphe valve, the striae are parallel but radiate very slightly and curve near the apices. The shorter striae in the central part consist of 4–6 areolae. The terminal fissures of the raphe are hooked toward the same side (arrow in Figure 3A). Central raphe endings are laterally expanded (Figure 3C). The sternum is narrow and slightly broader in the central area (Figure 3A).
The rapheless valve is convex. The axial area is below the valve plane, and it forms a shallow V (arrow in Figure 4A). A row of slit-like areolae is present on the mantles of the raphe and rapheless valves (Figure 4A). In the oblique view, the helictoglossae are raised internally (Figure 4B, arrow). Areolae are occluded by hymens, which are connected in adjacent areolae. Pairs of unconnected vimines are present above the adjacent areolae (Figure 4B); however, the areolae near the margin area are separate from the neighboring areolae. The structure of the areolae from the inner parts of both valves is formed by hymenes with marginal slits; the thickness of the central disk differs from that of the marginal area (Figure 4C). Hymenes have perforations of the parallel array type (solid arrow) and the centric array type (dotted arrow) (Figure 4D).
Holotype: A slide of the isolate 180409KCB8B40511, illustrated in Figure 2A–N, was deposited at the Freshwater Bioresources Research Bureau, Nakdonggang National Institute of Biological Resources (slide number FBCC210015D).
Isotype: A slide of the isolate 180409KCB8B40511, illustrated in Figure 2O, was deposited at the Freshwater Bioresources Research Bureau, Nakdonggang National Institute of Biological Resources (slide number FBCC210015D).
Molecular characterization: Nucleotide sequences of the SSU rRNA and rbcL genes of strain 180409KCB8B40511 were deposited in GenBank (NCBI; accession numbers MK578710 and MK639354, respectively).
Locality: 37°31′58.90″ N, 128°00′42.00″ E; Gye Stream, Hoengseong, Korea. Minzi Miao collected the specimen on 4 April 2018.
Etymology: The epithet ovale refers to the diatoms’ ovate outline.
Habitat: This species is an epilithon diatom that lives in flowing freshwater. The environmental variables of this species’ habitat are shown in Table 1.
3.1.2. Achnanthidium cavitatum M. Miao & B.-H. Kim, sp. nov.
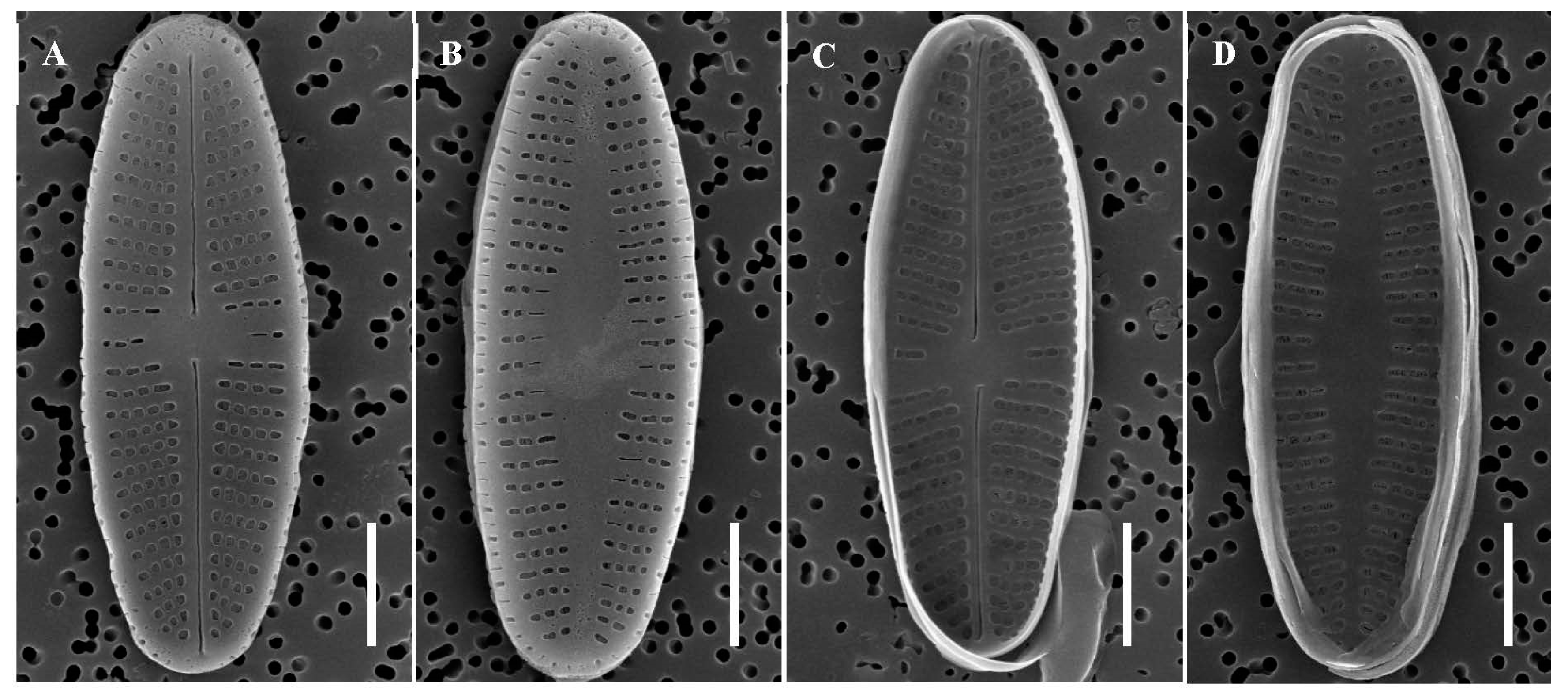
Description: Cells are linear-elliptical with slightly drawn-out ends, 8.8–13.0 µm long, and 3.0–3.5 µm wide. Striae density on the raphe valve is 25–28 in 10 µm in the center and 30–35 in 10 µm at the apices. Striae are radiate at the apices and weakly radiate in the central area. The raphe valve has a linear axial area that becomes wider in the central area. The raphe end is straight at the apices and teardrop-like in the central area. Raphe terminal fissures are absent. Most areolae are round, but some are elongate and slit-like in the central area (Figure 6A,B). Slit-like areolae are more numerous on the rapheless valve than on the raphe valve (Figure 6A,B). Striae are more numerous on the primary side of the raphe and rapheless valves than on the secondary side (Figure 6A,B). On the internal side of the raphe valve, the central raphe endings gently curve in opposite directions (Figure 6C). The central area of rapheless valves is broadly lanceolate to linear and narrow (Figure 6D).
The raphe valves are concave, and the rapheless valves are convex (Figure 7A,B). The axial area is below the valve plane in the rapheless valves (Figure 7B). Areolae on the valve mantle are elongated to slit-like on both raphe and rapheless valves (Figure 7A,B). Internally, there are two types of hymenes, including the valve mantle, in the raphe and rapheless valves (Figure 7C,D, solid arrow). The external valve of the cell can be seen through the broken hymen, which has slit-like openings. Areolae with slit-like openings are loculate (Figure 7D, solid arrow). On the other side, externally, two shapes of areolae can be seen in the raphe and rapheless valves: slit-like (arrow S) to elongate or round (arrow E) (Figure 7E,F). There are two types of areola occlusions: (1) round or elongate-round opening, with hymenes between the external and internal valves; and (2) slit-like opening on the external valve, covered by hymenes on the internal valve plate, different from the round or elongate areolae.
Holotype: A slide of the isolate 180419HG03C4C30524, illustrated in Figure 5A,C–AB, was deposited at the Freshwater Bioresources Culture Research Bureau, Nakdonggang National Institute of Biological Resources (slide number FBCC210016D).
Isotype: A slide of the isolate 180419HG03C4C30524, illustrated in Figure 5B, was deposited at the Freshwater Bioresources Culture Research Bureau, Nakdonggang National Institute of Biological Resources (slide number FBCC210016D).
Molecular characterization: Nucleotide sequences of the SSU rRNA and rbcL genes of strain 180419HG03C4C30524 were deposited in GenBank (NCBI; accession numbers MK578711 and MK639355, respectively).
Locality: 38°4′28″ N, 127°24′52″ E; Yeongpyeong Stream, Pocheon, Korea. Minzi Miao collected the specimens on 13 April 2018.
Etymology: The epithet cavitata refers to the specimens’ broad axial central area on the rapheless valve.
Habitat: This species is an epilithon diatom and lives in flowing freshwater. The environmental variables of its habitat are shown in Table 1.
3.2. Molecular Phylogeny
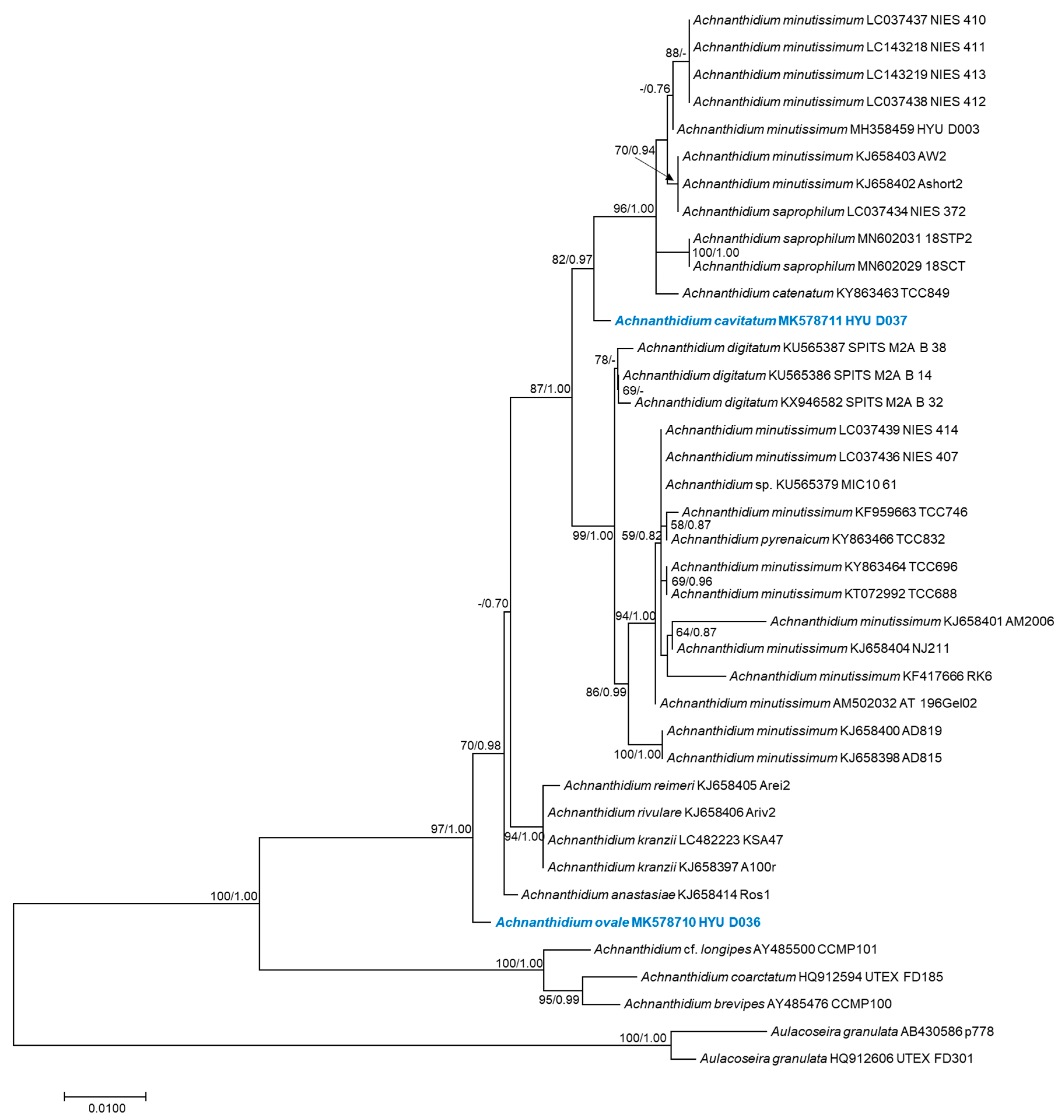
The phylogenetic positions of Achnanthidium ovale sp. nov. (HYU–D036) and Achnanthidium cavitatum sp. nov. (HYU–D037) were inferred using SSU rRNA and rbcL gene sequences (Figure 2G and Figure 8). ML and Bayesian analyses generated four similar trees that differed in only a few topological features. The results of the SSU-generated phylogenetic trees show that sequences of Achnanthidium species formed a monophyletic group with high statistical support (100% ML bootstrap support and 1.00 Bayesian posterior probability [PP]) (Figure 8). The phylogenetic positions of A. ovale (HYU–D036) and A. cavitatum (HYU–D037) were clearly different from those of other Achnanthidium species. The similarity scores based on the SSU rRNA data are shown in Table 4. The highest similarity score and lowest p-distance of A. ovale were found in the comparisons to A. reimeri (Arei2) (0.988) and A. anastasiae (Ros1) (0.006), respectively. The highest similarity score and lowest p-distance of A. cavitatum were found in the comparisons to A. catenatum (TCC849) (similarity score = 0.990; p-distance = 0.004).
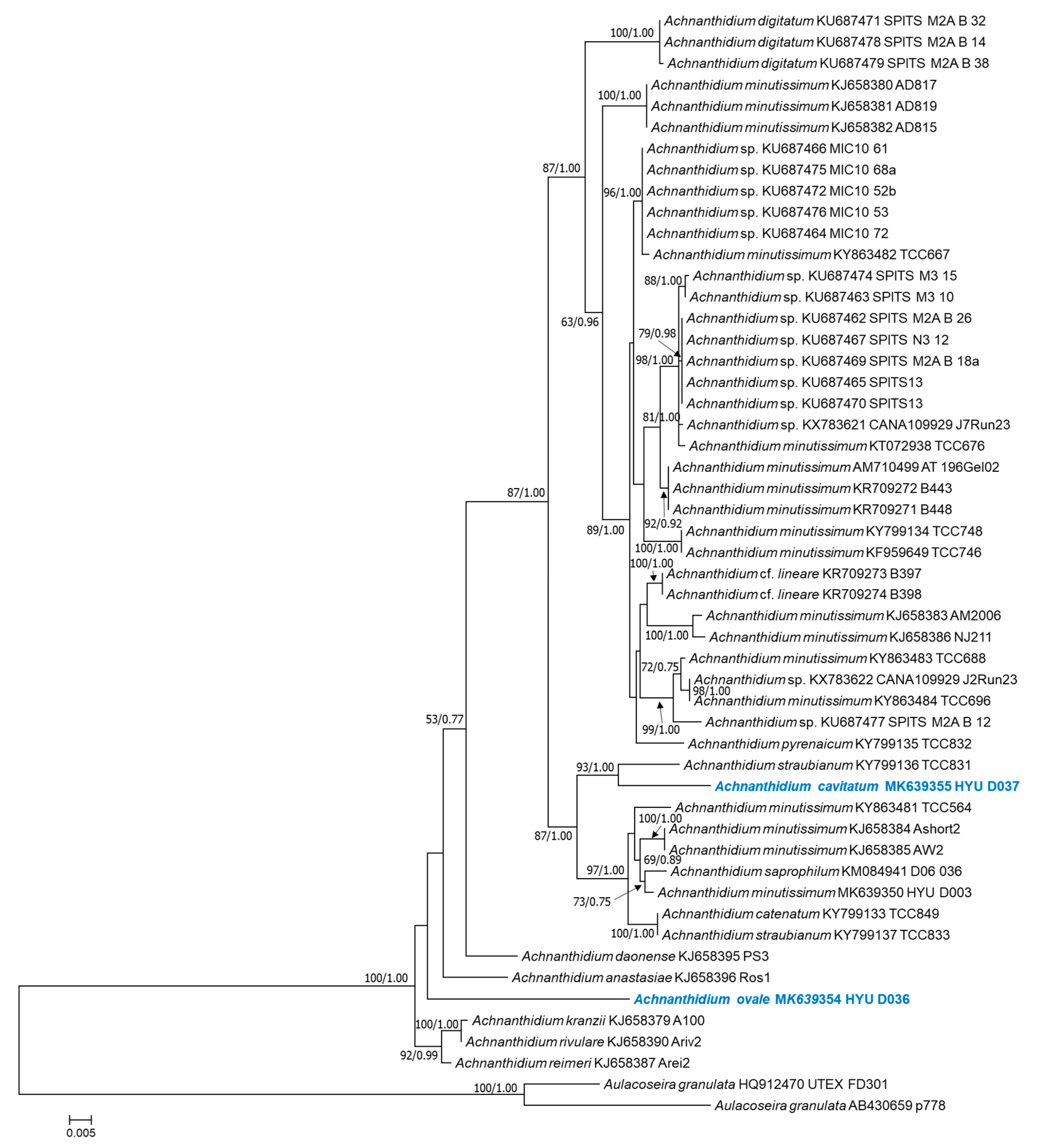
The rbcL-generated phylogenetic tree also shows that the position of A. ovale (HYU–D036) is distinct from those of other Achnanthidium species (Figure 9). A. cavitatum (HYU–D037) and A. straubianum (TCC831) form a single cluster with strong support (93% ML bootstrap and 1.00 Bayesian PP). The similarity scores based on rbcL gene sequences are shown in Table 5. Achnanthidium ovale had the highest similarity score (0.956) and lowest genetic distance (0.045) compared with A. anastasiae (Ros1), and A. cavitatum had the highest similarity score (0.971) and lowest genetic distance (0.028) compared with A. straubianum (TCC831).
4. Discussion
4.1. Achnanthidium ovale as a New Species
Achnanthidium ovale sp. nov. has terminal raphe endings that turn to the same side. This characteristic is typical of species belonging to the A. pyrenaicum complex. Table 6 shows a detailed comparison between A. ovale and similar species from the A. pyrenaicum complex taxa: A. rivulare Potapova & Ponader [10], A. pyrenaicum (Hustedt) Kobayasi (Karthick et al. [13]), and A. convergens Kobayasi [45]. Although the smaller A. rivulare is similar to A. ovale in valve outline, these species differ in (1) areola openings, (2) striation pattern on the raphe valve, and (3) internal raphe endings. The areola openings in A. ovale are mostly elongated or sometimes small and rounded, unlike the areolae in A. rivulare, which are mostly rounded. The striae on the apical area of the raphe valve in A. ovale are radiate, unlike the convergent striae of A. rivulare. Internally, A. ovale has deflected raphe central endings, whereas A. rivulare has hooked raphe central endings. In A. rivulare, the number of mantle areolae at the valve ends that do not correspond to the areolae on the valve face varies between 1 and 4 but is usually 2 or 3 ([10]; Figure 5E–K,M,S); however, in A. ovale, the number of areolae that do not have corresponding areolae on the valve face is as high as 5 (Figure 3A). According to Kobayasi [5], the number of freestanding areolae is a species-specific characteristic in Achnanthidium. Potapova & Ponader [10] considered that the number of areolae varied not only among species, but also within a single valve. Achnanthidium ovale differs from A. pyrenaicum in outline, as the latter has slightly drawn-out ends. Moreover, the outline and striation pattern of A. ovale differ from those of A. convergens. In both SSU rRNA and rbcL phylogenetic trees, A. ovale has a unique phylogenetic position, as appropriate to establish a new species. In addition, it is slightly related to A. daonense and A. anastasiae in the A. minutissimum complex, with less support.
4.2. Achnanthidium cavitatum as a New Species
Achnanthidium cavitatum has straight terminal raphe endings, which is characteristic of members of the A. minutissimum complex. Table 7 shows a detailed comparison between A. cavitatum and similar species from the A. minutissimum complex: A. minutissimum [6], A. saprophilum Round and Bukhtiyarova (Hlubikova et al. [24]), A. eutrophilum Lange-Bertalot (Hlubikova et al. [24]), and A. duriense Novais & Ector (Novais et al. [16]).
Achnanthidium cavitatum has two conspicuous characteristics that are typically observed in species from the A. minutissimum complex: (a) axial central area broadly lanceolate to linear and narrow, which can be observed under LM (Figure 5R–AB), and (b) slit-like areolae, mostly near the axial central area, which can be observed using SEM (Figure 6B and Figure 7B,F). On the other hand, A. cavitatum differs from A. minutissimum in outline and A. saprophilum in outline and number of areolae per stria. Moreover, A. cavitatum differs from A. eutrophilum in outline and central area of the raphe valve and from A. duriense in outline and areola arrangement on internal view [16].
The SSU rRNA and rbcL gene phylogenetic trees indicate that A. cavitatum is part of the clade containing Achnanthidium strains (Figure 2G and Figure 8). Therefore, based on its morphological characteristics and molecular data, it is correct to classify A. cavitatum in the genus Achnanthidium. The axial central area of A. cavitatum differs from those of other Achnanthidium species. In addition, the molecular data show that A. cavitatum has low genetic distance and similarity scores compared to sequences from the NCBI database ( Table 4; Table 5). Therefore, we propose A. cavitatum as a new Achnanthidium species based on morphological and molecular analyses.
4.3. Areolae Occlusions and Openings
Loculate areolae are markedly constricted at one surface and occluded [19] by a velum (cribrum, rota, vola) or a hymen at the other. Yana & Mayama [23] described A. pseudoconspicuum var. yomensis to have loculate type areolae and incomplete vimines through lost hymenes. In the present study, we found a similar arrangement in A. ovale: the ultrastructure of the loculate areolae can be seen from the broken valve. Moreover, most A. ovale vimines are incomplete, and the hymens have different thicknesses from the margin area to the central area. However, A. cavitatum also has a different ultrastructure of the valve on the internal view. In the external view, the areola openings are slit-like or round and elongate on both the valve face and mantle (Figure 7A,B). On the internal view, slit-like and elongate areola openings can be seen through broken hymenes, and vimines are complete between areolae. Around the rapheless valve, the slit-like areola openings occluded by hymenes differ from the hymenes occluded by elongate areolae openings, depending on the depth of the valve (Figure 7D). Thomas [46] recognized the areola type of A. minutissimum as poroid. However, using SEM, we observed that the areolae of Achnanthidium are loculated and internally covered by hymenes.
4.4. Ecological Characteristics of Two Achnanthidium Species
Achnanthidium ovale sp. nov. and A. cavitatum sp. nov. were recorded in the Gye Stream and the Yeongpyeong Stream (Korea), respectively. The summary of environmental data is shown in Table 1. Previous studies on Achnanthidium stated that species of this genus live in alkaline to acidic environments [14]; this is supported by the results of the present study. Achnanthidium ovale and A. minutissimum were collected from an alkaline environment, whereas A. cavitatum was collected from an acidic environment. The water velocity in the location of the two species was 0–80 cm/s (Table 1). Dissolved oxygen (5.93–9.86 mg/L), water temperature (4.10–11.71 °C), conductivity (57–148 μS/Cm), and turbidity (0.0–5.4 NTU) in the two locations differed widely. Studies have shown that water-quality assessment methods can be based on genus-level identifications because species within a genus can live in different ecological conditions [15,47,48,49]. The two species in our study, although they are from the same genus, live in different pH, dissolved oxygen, water temperature, velocity, conductivity, and turbidity conditions; therefore, our results support previous conclusions about the genus Achnanthidium.
Achnanthidium ovale was collected from a stone in Gye Stream, which has a fast flow and low conductivity and turbidity. Land use and cover conditions within a 1 km radius of this area are forest (50%) and agriculture (50%) [50]. The dominant species here is A. minutissimum (77.31%), a widespread species found in low abundance in polluted rivers [15].
Achnanthidium cavitatum was collected from a stone in the Yeongpyeong Stream, which is a slightly acidic environment with low conductivity and turbidity. Land use and cover conditions of this area are forest (80%) and urban (20%) [50]. The stream is fast-flowing and has a stony substratum. The dominant species in this area is Hannaea arcus var. recta Idei (41.33%), which is a saproxenous species [51]. The subdominant species is Achnanthidium minutissimum (22.45%).
Author Contributions
M.M. and B.-H.K. conceived the research; M.M., H.-K.K., H.L., E.A.-H., and B.-H.K. performed fieldwork; M.M. and Z.L. analyzed the data; M.M., Z.L., and B.-H.K. wrote and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partly supported by Hyperspectral Remote Sensing of Algal Distribution of Inherent Optical Properties (NIER-RP2018) and the KRIBB Research Initiative Program.
Acknowledgments
The authors thank the two anonymous referees for their valuable and constructive comments.
Conflicts of Interest
The authors report no potential conflict of interest.
References
- Patrick, R.; Reimer, C.W. The Diatoms of the United States Exclusive of Alaska and Hawaii; Academy of the Natural Sciences: Philadelphia, PA, USA, 1966; p. 688. [Google Scholar]
- Round, F.E.; Crawford, R.M.; Mann, D.G. Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Czarnecki, D.B. The freshwater diatoms culture collection at Loras College, Dubuque, Iowa. Mem. Calif. Acad. Sci. 1994, 17, 155–174. [Google Scholar]
- Round, F.E.; Bukhtiyarova, L. Four new genera based on Achnanthes (Achnanthidium) together with a re-definition of Achnanthidium. Diatom Res. 1996, 11, 345–361. [Google Scholar] [CrossRef]
- Kobayashi, H. Comparative Studies among Four Linear-Lanceolate Achnanthidium Species (Bacillariophyceae) with curved terminal raphe endings. Nova Hedwigia 1997, 65, 147–163. [Google Scholar] [CrossRef]
- Potapova, M.; Hamilton, P.B. Morphological and ecological variation within the Achnanthidium minutissimum (Bacillariophyceae) species complex. J. Phycol. 2007, 43, 561–575. [Google Scholar] [CrossRef]
- Yu, P.; Kociolek, J.P.; You, Q.M.; Wang, Q.X. Achnanthidium longissimum sp. nov. (Bacillariophyta), a new diatom species from Jiuzhai Valley, Southwestern China. Diatom Res. 2018, 33, 339–348. [Google Scholar] [CrossRef]
- Witkowski, A.; Kulikovskiy, M.; Riaux-Gobin, C. Achnanthidium sieminskae, a new diatom species from the Kerguelen Archipelago (Austral Islands). In Current Advances in Algal Taxonomy and Its Applications: Phylogenetic, Ecological and Applied Perspective; Wołowski, K., Kaczmarska, I., Ehrman, J.M., Wojtal, A.Z., Eds.; Polish Academy of Sciences: Kraków, Poland, 2012; pp. 61–68. [Google Scholar]
- Pérès, F.; Le Cohu, R.; Delmont, D. Achnanthidium barbei sp. nov. and Achnanthidium costei sp. nov., two new diatom species from French rivers. Diatom Res. 2014, 29, 387–397. [Google Scholar] [CrossRef]
- Potapova, M.G.; Ponader, K.C. Two common North American diatoms, Achnanthidium rivulare sp. nov. and A. deflexum (Reimer) Kingston: Morphology, ecology and comparison with related species. Diatom Res. 2004, 19, 33–57. [Google Scholar] [CrossRef]
- Wojtal, A.Z.; Ector, L.; Van de Vijver, B.; Morales, E.A.; Blanco, S.; Piątek, J.; Smieja, A. The Achnanthidium minutissimum complex (Bacillariophyceae) in southern Poland. Algol. Stud. 2011, 136, 211–238. [Google Scholar] [CrossRef]
- Pérès, F.; Barthès, A.; Ponton, E.; Coste, M.; Ten-Hage, L.; Le Cohu, R. Achnanthidium delmontii sp. nov., a new species from French rivers. Fottea 2012, 12, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Karthick, B.; Taylor, J.C.; Hamilton, P.B. Two new species of Achnanthidium Kützing (Bacillariophyceae) from Kolli Hills, Eastern Ghats, India. Fottea 2017, 17, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Martín, G.; Toja, J.; Sala, S.E.; De los Reyes Fernández, M.; Reyes, I.; Casco, M.A. Application of diatom biotic indices in the Guadalquivir River Basin, a Mediterranean basin. Which one is the most appropriated? Environ. Monit. Assess. 2010, 170, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Ponader, K.C.; Potapova, M.G. Diatoms from the genus Achnanthidium inflowing waters of the Appalachian Mountains (North America): Ecology, distribution and taxonomic notes. Limnologica 2007, 37, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Novais, M.H.; Juttner, I.; Van de Vijver, B.; Morais, M.M.; Hoffmann, L.; Ector, L. Morphological variability within the Achnanthidium minutissimum species complex (Bacillariophyta): Comparison between the type material of Achnanthes minutissima and related taxa, and new freshwater Achnanthidium species from Portugal. Phytotaxa 2015, 224, 101–139. [Google Scholar] [CrossRef]
- Cox, E.J. Ontogeny, homology, and terminology—Wall morphogenesis as an aid to character recognition and character state definition for pennate diatom systematics. J. Phycol. 2012, 48, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Cox, E.J. Symmetry and valve structure in naviculoid diatoms. Nova Hedwigia 1979, 64, 193–206. [Google Scholar]
- Cox, E.J. Variation in patterns of valve morphogenesis between representatives of six biraphid diatom genera (Bacillariophyceae). J. Phycol. 1999, 35, 1297–1312. [Google Scholar] [CrossRef]
- Ross, R.; Cox, E.J.; Karayeva, N.I.; Mann, D.G.; Paddock, T.B.B.; Simonsen, R.; Sims, P.A. An amended terminology for the siliceous components of the diatom cell. Nova Hedwigia 1979, 64, 513–533. [Google Scholar]
- Mann, D.G. Sieves and Flaps: Siliceous Minutiae in the Pores of Raphid Diatoms. In Proceedings of the 6th Symposium on Recent and Fossil Diatoms; Ross, R., Ed.; O. Koeltz: Koenigstein, Germany, 1981; pp. 279–300. [Google Scholar]
- Cox, E.J. Morphogenetic information and the selection of taxonomic characters for raphid diatom systematics. Plant Ecol. Evol. 2010, 143, 271–277. [Google Scholar] [CrossRef]
- Yana, E.; Mayama, S. Two new taxa of Achnanthidium and Encyonema (Bacillariophyceae) from the Yom River, Thailand, with special reference to the areolae occlusions implying ontogenetic relationship. Phycol. Res. 2015, 63, 239–252. [Google Scholar] [CrossRef]
- Hlubikova, D.; Ector, L.; Hoffmann, L. Examination of the type material of some diatom species related to Achnanthidium minutissimum (Kütz.) Czarn. (Bacillariophyceae). Algol. Stud. 2011, 136, 19–43. [Google Scholar] [CrossRef]
- Bruder, K.; Medlin, L.K. Molecular assessment of phylogenetic relationships in selected species/genera in the naviculoid diatoms (Bacillariophyta). I. The genus Placoneis. Nova Hedwigia 2007, 85, 331–352. [Google Scholar] [CrossRef]
- Medlin, L.K.; Kaczmarska, I. Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 2004, 43, 245–270. [Google Scholar] [CrossRef] [Green Version]
- Medlin, L.K.; Williams, D.M.; Sims, P.A. The evolution of the diatoms (Bacillariophyta). I. Origin of the group and assessment of the monophyly of its major divisions. Eur. J. Phycol. 1993, 28, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Medlin, L.K.; Cooper, A.; Hill, C.; Wrieden, S.; Wellbrock, U. Phylogenetic position of the Chromista plastids based on small subunit rRNA coding regions. Curr. Genet. 1995, 28, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L.K.; Kooistra, W.H.; Gersonde, R.; Wellbrock, U. Evolution of the diatoms (Bacillariophyta). II. Nuclear-encoded small-subunit rRNA sequence comparisons confirm a paraphyletic origin for the centric diatoms. Mol. Biol. Evol. 1996, 13, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Sorhannus, U. Diatom phylogenetics inferred based on direct optimization of nuclear-encoded SSU rRNA sequences. Cladistics 2004, 20, 487–497. [Google Scholar] [CrossRef]
- Sato, S.; Mann, D.G.; Matsumoto, S.; Medlin, L.K. Pseudostriatella (Bacillariophyta): A description of a new araphid diatom genus based on observations of frustule and auxospore structure and 18S rDNA phylogeny. Phycologia 2008, 47, 371–391. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Park, B.S.; Kim, J.H.; Kim, J.H.; Lee, H.O.; Han, M.S. Phylogenetic position of eight Amphora sensu lato (Bacillariophyceae) species and comparative analysis of morphological characteristics. Algae 2014, 29, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Sui, Z.; Zhang, S.; Ren, Y.; Liu, Y. Comparison of potential diatom ‘barcode’ genes (the 18S rRNA gene and ITS, COI, rbcL) and their effectiveness in discriminating and determining species taxonomy in the Bacillariophyta. Int. J. Syst. Evol. Microbiol. 2015, 65, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.G.; Simpson, G.E.; Sluiman, H.J.; Möller, M. rbcL gene tree of diatoms: A second large Data-Set for phylogenetic reconstruction. Phycologia 2001, 40, 1–2. [Google Scholar]
- Kulikovskiy, M.S.; Andreeva, S.A.; Gusev, E.S.; Kuznetsova, I.V.; Annenkova, N.V. Molecular phylogeny of monoraphid diatoms and raphe significance in evolution and taxonomy. Biol. Bull. 2016, 43, 398–407. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, P.; Kim, H.K.; Lee, H.; Han, M.S.; Kim, B.H. Lemnicola hungarica (Bacillariophyceae) and the new monoraphid diatom Lemnicola uniseriata sp. nov. (Bacillariophyceae) from Korea. Diatom Res. 2018, 33, 69–87. [Google Scholar] [CrossRef]
- Andersen, R.A. Algal Culturing Techniques; Academic Press: San Diego, CA, USA, 2005; p. 578. [Google Scholar]
- Beakes, G.W.; Canter, H.M.; Jaworski, G.H.M. Zoospore ultrastructure of Zygorhizidium affluens and Z. planktonicum, two chytrids parasitizing the diatom Asterionella formosa. Can. J. Bot. 1988, 66, 1054–1067. [Google Scholar] [CrossRef]
- Katano, T.; Lee, J.; Ki, J.S.; Kang, S.H.; Han, M.S. Effects of temperature and salinity on the growth and the optimum nitrogen to phosphorus ratio for the culture of Diatoma tenue isolated from a temporary Arctic pond in Svalbard, Norway. J. Freshw. Ecol. 2007, 22, 629–635. [Google Scholar] [CrossRef]
- Ki, J.S.; Han, M.S. Molecular analysis of complete SSU to LSU rRNA sequence in the harmful dinoflagellate Alexandrium tamarense (Korean isolate, HY970328M). Ocean Sci. J. 2005, 40, 155–166. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Fredrik, R.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar]
- Kobayasi, H.; Nagumo, T.; Mayama, S. Observation on the two rheophilic species of the genus Achnanthes (Bacillariophyceae), A. convergens H. Kob. And A. japonica H. Kob. Diatom 1986, 2, 83–93. [Google Scholar]
- Thomas, E.W. Exploring Species Boundaries in the Diatom Genus Rhoicosphenia Using Morphology, Phylogeny, Ecology, and Biogeography. Ph.D. Thesis, Department of Ecology & Evolutionary, University of Colorado, Boulder, Co, USA, 2016. [Google Scholar]
- Chessman, B.; Growns, I.; Currey, J.; Plunkett-Cole, N. Predicting diatom communities at the genus level for the rapid biological assessment of rivers. Freshw. Biol. 1999, 41, 317–331. [Google Scholar] [CrossRef]
- Wu, J.T. A generic index of diatom assemblages as bioindicator of pollution in the Keelung River of Taiwan. Hydrobiologia 1999, 397, 79–87. [Google Scholar] [CrossRef]
- Hill, B.H.; Herlihy, A.T.; Kaufmann, P.R.; Stevenson, R.J.; McCormick, F.H.; Johnson, C.B. Use of periphyton assemblage data as an index of biotic integrity. J. N. Am. Benth. Soc. 2000, 19, 50–67. [Google Scholar] [CrossRef]
- NGII (National Geographic Information Institute). Available online: https://www.ngii.go.kr/ (accessed on 11 June 2020).
- Kobayasi, H.; Idei, M.; Mayama, S.; Nagumo, T.; Osada, K. Kobayasi’s Atlas of Japanese Diatoms Based on Electron Microscopy; Uchida Rokakuho: Tokyo, Japan, 2006; p. 531. [Google Scholar]
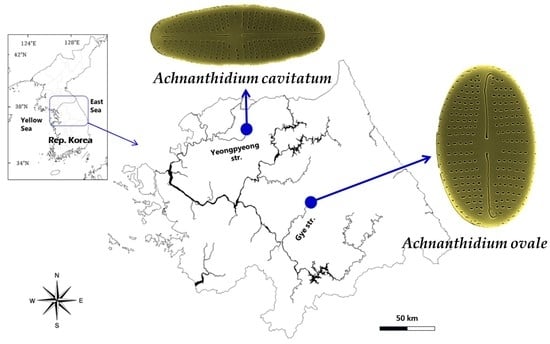
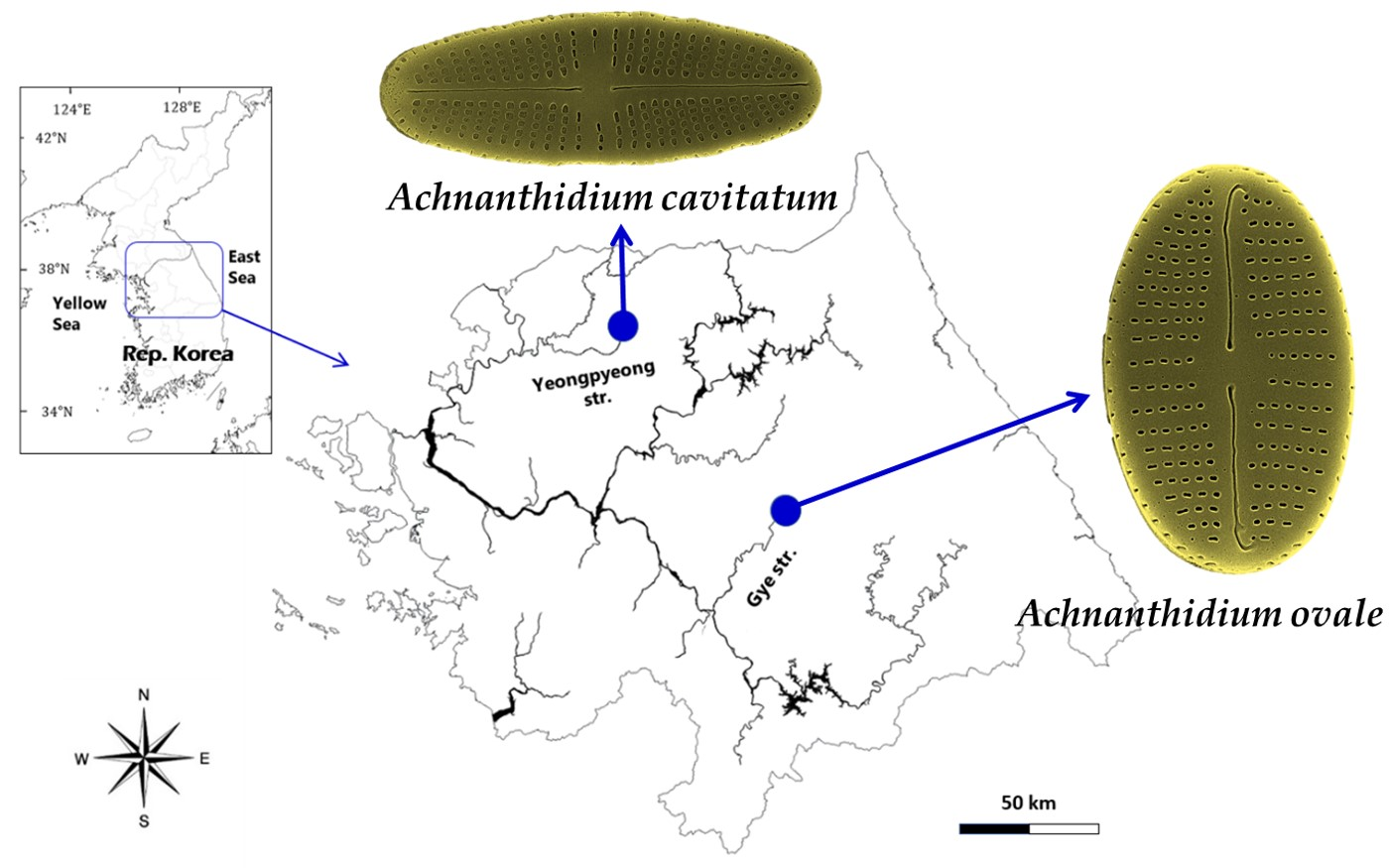
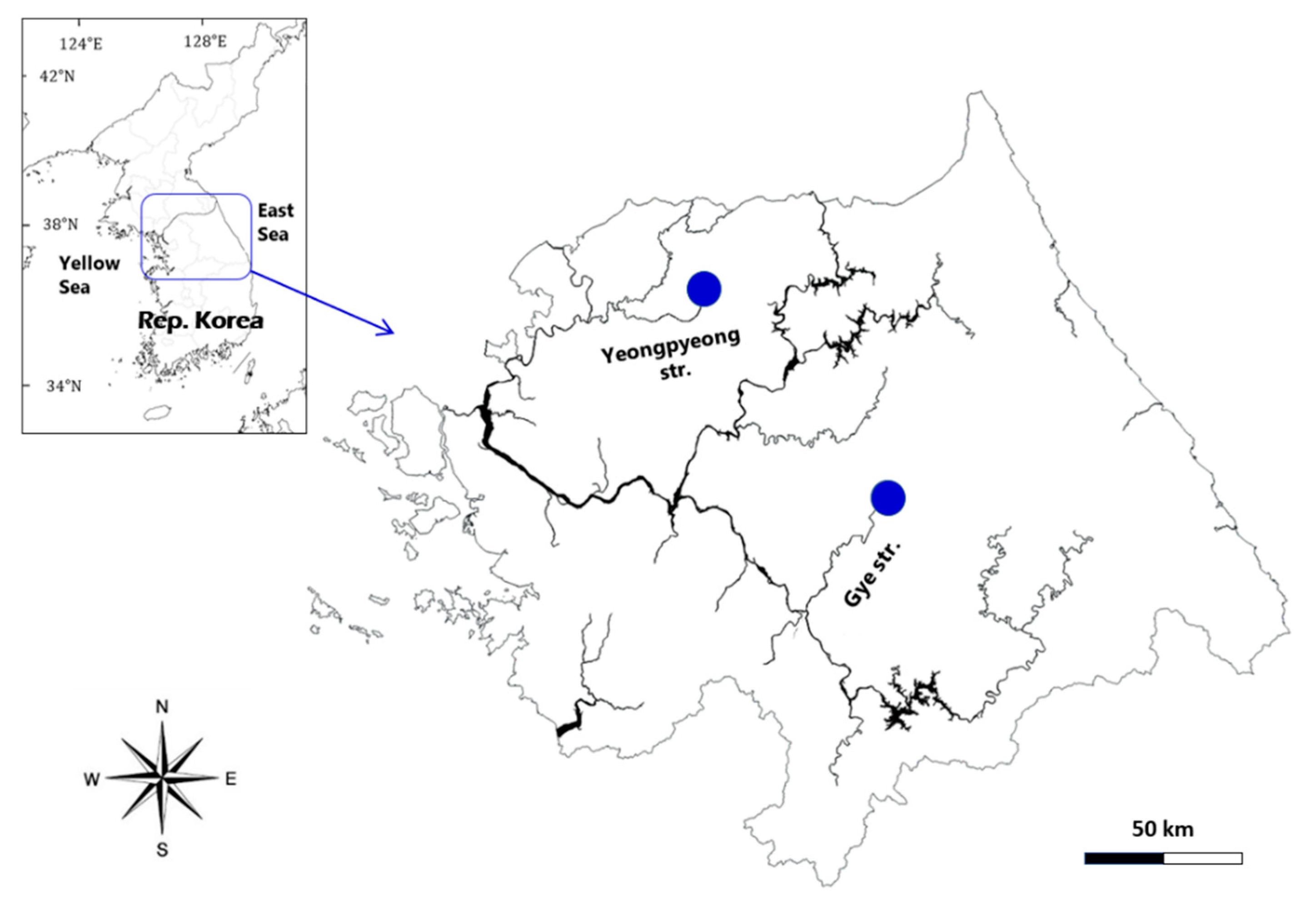
Figure 1.
A map showing the sampling sites of two diatoms as Achnanthidium ovale from Gye Stream in Hoengseong, Kangwon-do and A. cavitatum from the upstream of Yeonpyeong in Pocheon city, Gyeonggi-do, the Hangang River, Republic of Korea.
Figure 1.
A map showing the sampling sites of two diatoms as Achnanthidium ovale from Gye Stream in Hoengseong, Kangwon-do and A. cavitatum from the upstream of Yeonpyeong in Pocheon city, Gyeonggi-do, the Hangang River, Republic of Korea.

Figure 2.
LM of Achnanthidium ovale sp. nov. taken from the holotype population (180409KCB8B40511). Scale bar = 10 µm. (A). Girdle view of A. ovale. (B–H) Raphe view of cell valve. (I–O) Rapheless view of cell valve.
Figure 2.
LM of Achnanthidium ovale sp. nov. taken from the holotype population (180409KCB8B40511). Scale bar = 10 µm. (A). Girdle view of A. ovale. (B–H) Raphe view of cell valve. (I–O) Rapheless view of cell valve.

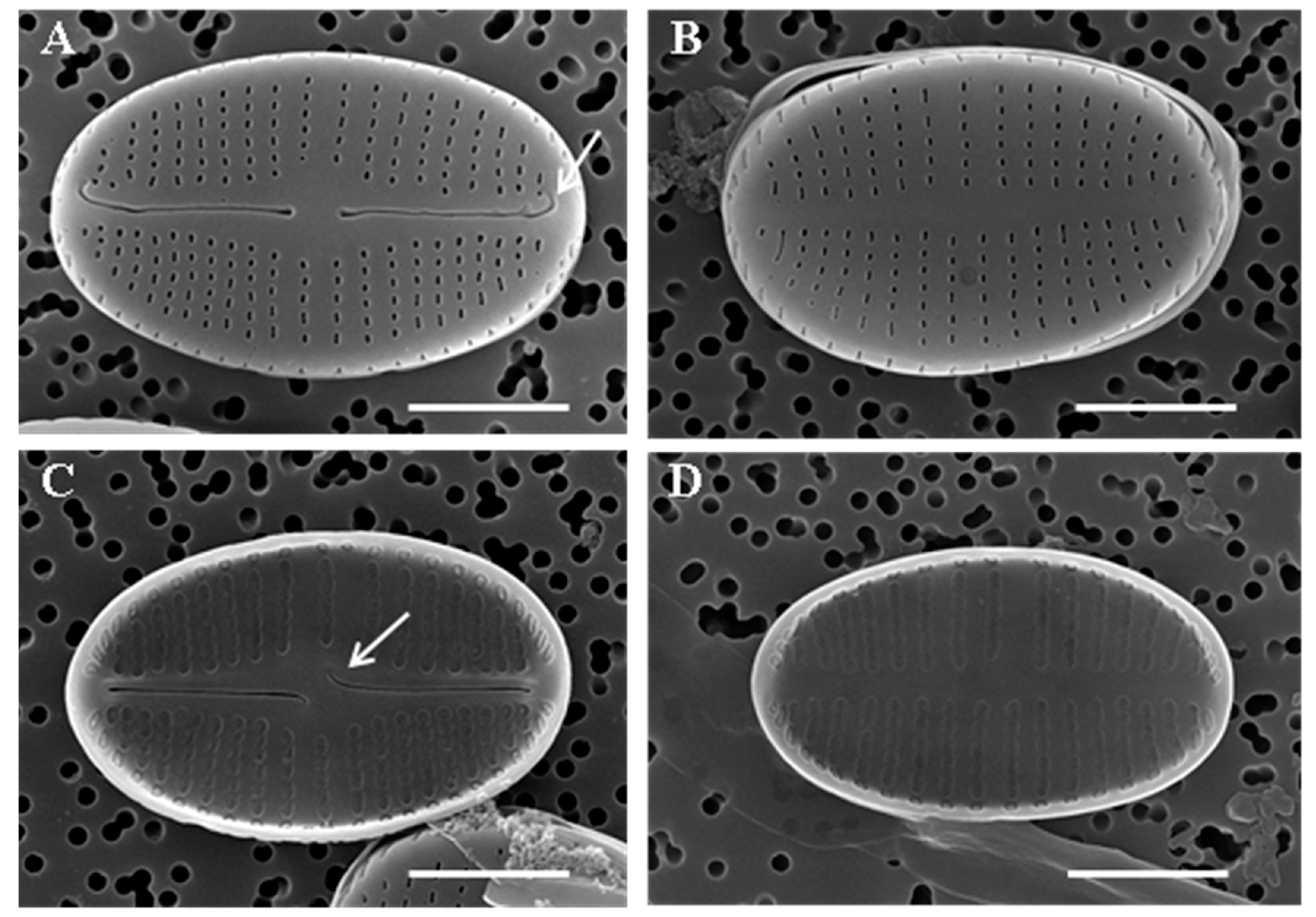
Figure 3.
SEM of Achnanthidium ovale sp. nov. taken from the holotype population (180409KCB8B40511). Scale bar = 2 µm. (A) External view of raphe valve. Hooked terminal raphe fissures (arrow). (B) External view of rapheless valve. (C) Internal view of raphe valve. Central raphe endings deflect in the opposite direction in internal view (arrow). (D) Internal view of rapheless valve.
Figure 3.
SEM of Achnanthidium ovale sp. nov. taken from the holotype population (180409KCB8B40511). Scale bar = 2 µm. (A) External view of raphe valve. Hooked terminal raphe fissures (arrow). (B) External view of rapheless valve. (C) Internal view of raphe valve. Central raphe endings deflect in the opposite direction in internal view (arrow). (D) Internal view of rapheless valve.

Figure 4.
SEM of Achnanthidium ovale sp. nov. taken from the holotype population (180409KCB8B40511). (A) Convex rapheless valve with shallow V axial area (arrow). Scale bar = 2 µm. (B) The raised helictoglossae (arrow) and unconnected vimines above the adjacent areolae in internal view. Scale bar = 2 µm. (C) Internally, areolae structures are hymenes with marginal slits which have different thickness (arrow). Scale bar = 0.5 µm. (D) The areolae close to the margin area separate from neighboring areolae. Hymenes with perforations of a parallel array type (solid arrow) and centric array type (dotted arrow). Scale bar = 0.2 µm.
Figure 4.
SEM of Achnanthidium ovale sp. nov. taken from the holotype population (180409KCB8B40511). (A) Convex rapheless valve with shallow V axial area (arrow). Scale bar = 2 µm. (B) The raised helictoglossae (arrow) and unconnected vimines above the adjacent areolae in internal view. Scale bar = 2 µm. (C) Internally, areolae structures are hymenes with marginal slits which have different thickness (arrow). Scale bar = 0.5 µm. (D) The areolae close to the margin area separate from neighboring areolae. Hymenes with perforations of a parallel array type (solid arrow) and centric array type (dotted arrow). Scale bar = 0.2 µm.

Figure 5.
LM of Achnanthidium cavitatum sp. nov. taken from the holotype population (180419HG03C4C30524). Scale bar = 10 µm. (A–P) Raphe view of cell valve. (Q) Girdle view of A. cavitatum. (R–AB) Rapheless view of cell valve.
Figure 5.
LM of Achnanthidium cavitatum sp. nov. taken from the holotype population (180419HG03C4C30524). Scale bar = 10 µm. (A–P) Raphe view of cell valve. (Q) Girdle view of A. cavitatum. (R–AB) Rapheless view of cell valve.

Figure 6.
SEM of Achnanthidium cavitatum sp. nov. taken from the holotype population (180419HG03C4C30524). Scale bar = 2 µm. (A) External view of raphe valve with straight terminal raphe fissures. (B) External view of rapheless valve with a broadly lanceolate axial central area. (C) Internal view of raphe valve. (D) Internal view of rapheless valve.
Figure 6.
SEM of Achnanthidium cavitatum sp. nov. taken from the holotype population (180419HG03C4C30524). Scale bar = 2 µm. (A) External view of raphe valve with straight terminal raphe fissures. (B) External view of rapheless valve with a broadly lanceolate axial central area. (C) Internal view of raphe valve. (D) Internal view of rapheless valve.

Figure 7.
SEM of Achnanthidium cavitatum sp. nov. taken from the holotype population (180419HG03C4C30524). (A) Concave raphe valve. Scale bar = 2 µm. (B) Convex rapheless valve with V shallow axial area. Scale bar = 2 µm. (C) Internal view on broken hymenes of rapheless valve. The external valve of the cell can be seen through the broken hymen, which has slit liked openings. The areolae with slit-liked openings are loculate areolae. Scale bar = 0.3 µm. (D) Different areolae occlusion on the internal view of rapheless valve and margin area of it (arrows). Scale bar = 0.3 µm. (E) Teardrop-shaped central raphe endings. Different shapes of areolae on the external view of the raphe valve and margin area. Elongated or rounded shape of areolae on valve plate and side (arrow E). Slit-liked areolae on valve plate and sides (arrow S). Scale bar = 1 µm. (F) Different shapes of areolae on the external view of the rapheless valve and margin area. Scale bar = 1 µm.
Figure 7.
SEM of Achnanthidium cavitatum sp. nov. taken from the holotype population (180419HG03C4C30524). (A) Concave raphe valve. Scale bar = 2 µm. (B) Convex rapheless valve with V shallow axial area. Scale bar = 2 µm. (C) Internal view on broken hymenes of rapheless valve. The external valve of the cell can be seen through the broken hymen, which has slit liked openings. The areolae with slit-liked openings are loculate areolae. Scale bar = 0.3 µm. (D) Different areolae occlusion on the internal view of rapheless valve and margin area of it (arrows). Scale bar = 0.3 µm. (E) Teardrop-shaped central raphe endings. Different shapes of areolae on the external view of the raphe valve and margin area. Elongated or rounded shape of areolae on valve plate and side (arrow E). Slit-liked areolae on valve plate and sides (arrow S). Scale bar = 1 µm. (F) Different shapes of areolae on the external view of the rapheless valve and margin area. Scale bar = 1 µm.

Figure 8.
Maximum likelihood phylogenetic tree based on the SSU rRNA gene dataset showing the phylogenetic positions of Achnanthidium ovale sp. nov. (HYU–D036) and A. cavitatum sp. nov. (HYU–D037). Sequences from Aulacoseira granulata were used as the outgroup. The numbers on each node are the bootstrap value (%) and the Bayesian posterior probability (PP). Only bootstrap values above 50 and PP above 0.7 are shown. The GenBank accession and strain numbers follow the taxon names. Scale bar = 0.01 nucleotide substitutions per site.
Figure 8.
Maximum likelihood phylogenetic tree based on the SSU rRNA gene dataset showing the phylogenetic positions of Achnanthidium ovale sp. nov. (HYU–D036) and A. cavitatum sp. nov. (HYU–D037). Sequences from Aulacoseira granulata were used as the outgroup. The numbers on each node are the bootstrap value (%) and the Bayesian posterior probability (PP). Only bootstrap values above 50 and PP above 0.7 are shown. The GenBank accession and strain numbers follow the taxon names. Scale bar = 0.01 nucleotide substitutions per site.

Figure 9.
Maximum likelihood phylogenetic tree based on the rbcL gene dataset showing the phylogenetic positions of Achnanthidium ovale sp. nov. (HYU–D036) and A. cavitatum sp. nov. (HYU–D037). Sequences from Aulacoseira granulata were used as the outgroup. The numbers on each node are the bootstrap value (%) and the Bayesian posterior probability (PP). Only bootstrap values above 50 and PP above 0.7 are shown. The GenBank accession and strain numbers follow the taxon names. Scale bar = 0.005 nucleotide substitutions per site.
Figure 9.
Maximum likelihood phylogenetic tree based on the rbcL gene dataset showing the phylogenetic positions of Achnanthidium ovale sp. nov. (HYU–D036) and A. cavitatum sp. nov. (HYU–D037). Sequences from Aulacoseira granulata were used as the outgroup. The numbers on each node are the bootstrap value (%) and the Bayesian posterior probability (PP). Only bootstrap values above 50 and PP above 0.7 are shown. The GenBank accession and strain numbers follow the taxon names. Scale bar = 0.005 nucleotide substitutions per site.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Environmental variables at the sampling sites of Achnanthidium ovale sp. nov. and A. cavitatum sp. nov.
Table 1.
Environmental variables at the sampling sites of Achnanthidium ovale sp. nov. and A. cavitatum sp. nov.
| Achnanthidium ovale | Achnanthidium cavitatum | |
|---|---|---|
| Coordinates | 37°31′59α″ N, 128°0′42″ E | 38°4′28″ N, 127°24′52″ E |
| pH | 7.07 | 6.72 |
| Dissolved oxygen | 5.93 | 7.93 |
| Temperature (°C) | 11.71 | 11.29 |
| Velocity (cm/sec) | 80 | 20 |
| Conductivity (µS/cm) | 57 | 148 |
| Turbidity (NTU) | 0.0 | 3.1 |
Table 2.
Primers used to amplify and sequence the SSU rRNA and rbcL genes.
| Gene | Primer | Nucleotide Sequence (5′ to 3′) | Reference |
|---|---|---|---|
| SSU rRNA | AT18F01 | YAC-CTG-GTT-GAT-CCT-GCC-AGT-AG | [40] |
| AT18R02 | GTT-TCA-GCC-TTG-CGA-CCA-TAC-TCC | [40] | |
| AT18F02 | AGA-ACG-AAA-GTT-AAG-GGA-TCG-AAG-ACG | [40] | |
| AT18R01 | GCT-TGA-TCC-TTC-TGC-AGG-TTC-ACC | [40] | |
| rbcL | F3 | GCT-TAC-CGT-GTA-GAT-CCA-GTT-CC | [25] |
| R3 | CCT-TCT-AAT-TTA-CCA-ACA-ACT-G | [25] |
Table 3.
Strains of Achnanthidium ovale sp. nov. and A. cavitatum sp. nov. isolated in the Hangang River, Republic of Korea.
Table 3.
Strains of Achnanthidium ovale sp. nov. and A. cavitatum sp. nov. isolated in the Hangang River, Republic of Korea.
| Species | Strain | Taxonomic Position | Gene Type | Locality | GenBank Accession No. |
|---|---|---|---|---|---|
| Achnanthidium ovale sp. nov. | HYU-D036 | Achnanthales; Achnanthidiaceae | SSU | Korea | MK578710, this study |
| Achnanthidium ovale sp. nov. | HYU-D036 | Achnanthales; Achnanthidiaceae | rbcL | Korea | MK639354, this study |
| Achnanthidium cavitatum sp. nov. | HYU-D037 | Achnanthales; Achnanthidiaceae | SSU | Korea | MK578711, this study |
| Achnanthidium cavitatum sp. nov. | HYU-D037 | Achnanthales; Achnanthidiaceae | rbcL | Korea | MK639355, this study |
Table 4.
Similarity scores and genetic distances (SSU) of Achnanthidium species based on 540 bp of nuclear-encoded SSU rRNA gene sequences. GenBank accession and strain numbers follow the taxon names.
Table 4.
Similarity scores and genetic distances (SSU) of Achnanthidium species based on 540 bp of nuclear-encoded SSU rRNA gene sequences. GenBank accession and strain numbers follow the taxon names.
| Sequence Name | Achnanthidium ovale MK578710 HYU-D036 | Achnanthidium cavitatum MK578711 HYU-D037 | ||
|---|---|---|---|---|
| Similarity | p-Distance | Similarity | p-Distance | |
| Achnanthidium ovale MK578710 HYU-D036 | - | - | 0.977 | 0.012 |
| Achnanthidium cavitatum MK578711 HYU-D037 | 0.977 | 0.012 | - | - |
| Achnanthidium brevipes AY485476 CCMP100 | 0.921 | 0.044 | 0.913 | 0.046 |
| Achnanthidium catenatum KY863463 TCC849 | 0.971 | 0.016 | 0.99 | 0.004 |
| Achnanthidium cf. longipes AY485500 CCMP101 | 0.912 | 0.050 | 0.904 | 0.052 |
| Achnanthidium coarctatum HQ912594 UTEX FD185 | 0.915 | 0.050 | 0.908 | 0.052 |
| Achnanthidium digitatum KU565386 SPITS-M2A+B-14 | 0.969 | 0.014 | 0.979 | 0.014 |
| Achnanthidium digitatum KU565387 SPITS-M2A+B-38 | 0.967 | 0.014 | 0.977 | 0.014 |
| Achnanthidium digitatum KX946582 SPITS-M2A+B-32 | 0.969 | 0.014 | 0.979 | 0.014 |
| Achnanthidium kranzii KJ658397 A100r | 0.986 | 0.008 | 0.979 | 0.010 |
| Achnanthidium kranzii LC482223 KSA47 | 0.986 | 0.008 | 0.979 | 0.010 |
| Achnanthidium minutissimum AM502032 AT-196Gel02 | 0.967 | 0.018 | 0.975 | 0.018 |
| Achnanthidium minutissimum KF417666 RK6 | 0.935 | 0.030 | 0.933 | 0.030 |
| Achnanthidium minutissimum KF959663 TCC746 | 0.967 | 0.018 | 0.975 | 0.018 |
| Achnanthidium minutissimum KJ658398 AD815 | 0.963 | 0.020 | 0.977 | 0.018 |
| Achnanthidium minutissimum KJ658400 AD819 | 0.963 | 0.020 | 0.977 | 0.018 |
| Achnanthidium minutissimum KJ658401 AM2006 | 0.965 | 0.018 | 0.973 | 0.018 |
| Achnanthidium minutissimum KJ658402 Ashort2 | 0.971 | 0.016 | 0.986 | 0.008 |
| Achnanthidium minutissimum KJ658403 AW2 | 0.971 | 0.016 | 0.986 | 0.008 |
| Achnanthidium minutissimum KJ658404 NJ211 | 0.963 | 0.018 | 0.971 | 0.018 |
| Achnanthidium minutissimum KT072992 TCC688 | 0.967 | 0.018 | 0.975 | 0.018 |
| Achnanthidium minutissimum KY863464 TCC696 | 0.967 | 0.018 | 0.975 | 0.018 |
| Achnanthidium minutissimum LC037436 NIES-407 | 0.967 | 0.018 | 0.975 | 0.018 |
| Achnanthidium minutissimum LC037437 NIES-410 | 0.973 | 0.014 | 0.98 | 0.010 |
| Achnanthidium minutissimum LC037438 NIES-412 | 0.973 | 0.014 | 0.98 | 0.010 |
| Achnanthidium minutissimum LC037439 NIES-414 | 0.967 | 0.018 | 0.975 | 0.018 |
| Achnanthidium minutissimum LC143218 NIES-411 | 0.973 | 0.014 | 0.98 | 0.010 |
| Achnanthidium minutissimum LC143219 NIES-413 | 0.973 | 0.014 | 0.98 | 0.010 |
| Achnanthidium minutissimum MH358459 HYU-D003 | 0.971 | 0.016 | 0.986 | 0.008 |
| Achnanthidium pyrenaicum KY863466 TCC832 | 0.967 | 0.018 | 0.975 | 0.018 |
| Achnanthidium reimeri KJ658405 Arei2 | 0.988 | 0.006 | 0.977 | 0.012 |
| Achnanthidium rivulare KJ658406 Ariv2 | 0.986 | 0.008 | 0.979 | 0.010 |
| Achnanthidium saprophilum LC037434 NIES-372 | 0.971 | 0.016 | 0.986 | 0.008 |
| Achnanthidium saprophilum MN602029 18SCT | 0.971 | 0.014 | 0.979 | 0.010 |
| Achnanthidium saprophilum MN602031 18STP2 | 0.971 | 0.014 | 0.979 | 0.010 |
| Achnanthidium anastasiae KJ658414 Ros1 | 0.988 | 0.006 | 0.975 | 0.010 |
| Achnanthidium sp. KU565379 MIC10 61 | 0.967 | 0.018 | 0.975 | 0.018 |
Table 5.
Similarity scores and genetic distances of Achnanthidium species sequences based on 594 bp of chloroplast-encoded rbcL gene sequences. GenBank accession and strain numbers follow the taxon names.
Table 5.
Similarity scores and genetic distances of Achnanthidium species sequences based on 594 bp of chloroplast-encoded rbcL gene sequences. GenBank accession and strain numbers follow the taxon names.
| Sequence Name | Achnanthidium ovale MK578710 HYU-D036 | Achnanthidium cavitatum MK578711 HYU-D037 | ||
|---|---|---|---|---|
| Similarity | p-Distance | Similarity | p-Distance | |
| Achnanthidium ovale MK639354 HYU-D036 | - | - | 0.924 | 0.075 |
| Achnanthidium cavitatum MK639355 HYU-D037 | 0.924 | 0.075 | - | - |
| Achnanthidium anastasiae KJ658396 Ros1 | 0.956 | 0.045 | 0.946 | 0.052 |
| Achnanthidium catenatum KY799133 TCC849 | 0.925 | 0.066 | 0.961 | 0.033 |
| Achnanthidium cf. lineare KR709273 B397 | 0.930 | 0.068 | 0.957 | 0.043 |
| Achnanthidium cf. lineare KR709274 B398 | 0.930 | 0.068 | 0.957 | 0.043 |
| Achnanthidium daonense KJ658395 PS3 | 0.937 | 0.064 | 0.941 | 0.057 |
| Achnanthidium digitatum KU687471 SPITS-M2A+B-32 | 0.936 | 0.063 | 0.954 | 0.047 |
| Achnanthidium digitatum KU687478 SPITS-M2A+B-14 | 0.936 | 0.063 | 0.954 | 0.047 |
| Achnanthidium digitatum KU687479 SPITS-M2A+B-38 | 0.934 | 0.064 | 0.952 | 0.049 |
| Achnanthidium kranzii KJ658379 A100 | 0.944 | 0.057 | 0.929 | 0.070 |
| Achnanthidium minutissimum AM710499 AT-196Gel02 | 0.936 | 0.063 | 0.959 | 0.042 |
| Achnanthidium minutissimum KF959649 TCC746 | 0.934 | 0.064 | 0.954 | 0.047 |
| Achnanthidium minutissimum KJ658380 AD817 | 0.936 | 0.063 | 0.954 | 0.047 |
| Achnanthidium minutissimum KJ658381 AD819 | 0.936 | 0.063 | 0.954 | 0.047 |
| Achnanthidium minutissimum KJ658382 AD815 | 0.936 | 0.063 | 0.954 | 0.047 |
| Achnanthidium minutissimum KJ658383 AM2006 | 0.934 | 0.064 | 0.954 | 0.047 |
| Achnanthidium minutissimum KJ658384 Ashort2 | 0.929 | 0.070 | 0.966 | 0.035 |
| Achnanthidium minutissimum KJ658385 AW2 | 0.929 | 0.070 | 0.966 | 0.035 |
| Achnanthidium minutissimum KJ658386 NJ211 | 0.937 | 0.061 | 0.956 | 0.045 |
| Achnanthidium minutissimum KR709271 B448 | 0.936 | 0.063 | 0.959 | 0.042 |
| Achnanthidium minutissimum KR709272 B443 | 0.936 | 0.063 | 0.959 | 0.042 |
| Achnanthidium minutissimum KT072938 TCC676 | 0.937 | 0.061 | 0.957 | 0.043 |
| Achnanthidium minutissimum KY799134 TCC748 | 0.932 | 0.064 | 0.951 | 0.047 |
| Achnanthidium minutissimum KY863481 TCC564 | 0.927 | 0.070 | 0.968 | 0.031 |
| Achnanthidium minutissimum KY863482 TCC667 | 0.936 | 0.063 | 0.962 | 0.038 |
| Achnanthidium minutissimum KY863483 TCC688 | 0.934 | 0.064 | 0.959 | 0.042 |
| Achnanthidium minutissimum KY863484 TCC696 | 0.930 | 0.068 | 0.956 | 0.045 |
| Achnanthidium minutissimum MK639350 HYU-D003 | 0.930 | 0.068 | 0.966 | 0.035 |
| Achnanthidium pyrenaicum KY799135 TCC832 | 0.919 | 0.070 | 0.942 | 0.049 |
| Achnanthidium reimeri KJ658387 Arei2 | 0.952 | 0.049 | 0.932 | 0.066 |
| Achnanthidium rivulare KJ658390 Ariv2 | 0.944 | 0.057 | 0.929 | 0.070 |
| Achnanthidium saprophilum KM084941 D06-036 | 0.927 | 0.071 | 0.962 | 0.038 |
| Achnanthidium straubianum KY799136 TCC831 | 0.925 | 0.073 | 0.971 | 0.028 |
| Achnanthidium straubianum KY799137 TCC833 | 0.922 | 0.066 | 0.957 | 0.033 |
| Achnanthidium sp. KU687462 SPITS-M2A+B-26 | 0.934 | 0.064 | 0.957 | 0.043 |
| Achnanthidium sp. KU687463 SPITS-M3-10 | 0.932 | 0.064 | 0.959 | 0.040 |
| Achnanthidium sp. KU687464 MIC10-72 | 0.936 | 0.063 | 0.962 | 0.038 |
| Achnanthidium sp. KU687465 SPITS13 | 0.934 | 0.064 | 0.957 | 0.043 |
| Achnanthidium sp. KU687466 MIC10-61 | 0.936 | 0.063 | 0.962 | 0.038 |
| Achnanthidium sp. KU687467 SPITS-N3-12 | 0.934 | 0.064 | 0.957 | 0.043 |
| Achnanthidium sp. KU687469 SPITS-M2A+B-18a | 0.934 | 0.064 | 0.957 | 0.043 |
| Achnanthidium sp. KU687470 SPITS13 | 0.934 | 0.064 | 0.957 | 0.043 |
| Achnanthidium sp. KU687472 MIC10-52b | 0.936 | 0.063 | 0.962 | 0.038 |
| Achnanthidium sp. KU687474 SPITS-M3-15 | 0.932 | 0.064 | 0.959 | 0.040 |
| Achnanthidium sp. KU687475 MIC10-68a | 0.936 | 0.063 | 0.962 | 0.038 |
| Achnanthidium sp. KU687476 MIC10-53 | 0.936 | 0.063 | 0.962 | 0.038 |
| Achnanthidium sp. KU687477 SPITS-M2A+B-12 | 0.930 | 0.068 | 0.956 | 0.045 |
| Achnanthidium sp. KX783621 CANA109929-J7Run23 | 0.936 | 0.063 | 0.959 | 0.042 |
| Achnanthidium sp. KX783622 CANA109929-J2Run23 | 0.930 | 0.068 | 0.956 | 0.045 |
Table 6.
Morphological comparison between Achnanthidium ovale sp. nov. and similar species.
| A. ovale M. Miao & B.-H. Kim sp. nov. | A. rivulare Potapova & Ponader | A. pyrenaicum (Hustedt) Kobayasi | A. convergens Kobayasi | |
|---|---|---|---|---|
| length (µm) | 6.3–7.7 | 5.4–21.3 | 10.0–16.0 | 10.0–25.0 |
| width (µm) | 3.8–4.1 | 2.6–4.4 | 2.5–4.0 | 4–4.5 |
| valve outline | elliptical | linear-elliptical | linear-lanceolate with slightly drawn-out ends | linear-lanceolate |
| external areolae | elongate and dot-like | small, round, or slightly elongated | elongate or circular (RV) | constricted in various degrees. |
| internal areolae | hymenes partially joined | elliptical internal openings occluded by hymenes | hymenes not joined | hymenes partially joined, linking bars between interstriae partly interrupted |
| areolae in valve mantle | slit-like | slit-like | elongate | elongate |
| raphe valve | ||||
| density of striae (in 10 µm) | 30–35 (up to 55 near apices) | 19–25 (up to 55 near apices) | center: 20–25 apices: 34–40 | center: 18, apices: 36–40 |
| striation pattern | parallel but slightly radiate at apices | parallel but convergent or parallel near apices | parallel or slightly radiate in the central area and slightly convergent at apices | densely convergent striae near the valve ends |
| external raphe endings | laterally expanded | teardrop-shaped | teardrop-shaped | laterally expanded |
| internal raphe endings | deflected in opposite directions | short, hook-shaped | slightly curved to opposite sides | deflected in opposite directions |
| rapheless valve | ||||
| density of striae (in 10 µm) | 30–33 (up to 50 near apices) | 19–28 (up to 43 near apices) | center: 20–28 apices: 32–38 | |
| striation pattern | parallel but slightly radiate near apices | parallel but slightly radiate near apices | parallel or slightly radiate in the central area and slightly convergent at apices | slightly radiate at the ends |
| source | this study | [10] | [13] | [45] |
Table 7.
Morphological comparison between Achnanthidium cavitatum sp. nov. and similar species.
| A. cavitatum M. Miao & B.-H. Kim sp. Nov. | A. minutissimum (Kützing) Czarnecki | A. saprophilum (Kobayashi & Mayama) Round & Bukhtiyarova | A. eutrophilum (Lange-Bertalot) Lange-Bertalot | A. duriense Novais & Ector | |
|---|---|---|---|---|---|
| length (µm) | 8.8–10.3 | 9.0–14.5 | 9.5–14.5 | 7.5–16.0 | 5.0–9.7 |
| width (µm) | 3.0–3.5 | 2.5–3.177 | 3.0–3.6 | 3.2–4.8 | 2.0–2.7 |
| valve outline | rhombic with slightly drawn-out ends | linear-elliptic to linear-lanceolate | broadly linear | narrowly rhombic | elliptic to linear-elliptic |
| striation pattern | radiate at apices and weakly radiate in central area | radiate, denser toward the apices | radiate at apices and weakly radiate in central area | radiate at apices and weakly radiate in central area | almost parallel near the center; slightly radiate elsewhere |
| areolae in the valve mantle | slit-like or elongate | slit-like | slit-like | elongate or slit-like | elongate or slit-like |
| raphe valve | |||||
| density of striae (in 10 µm) | 30–32 | 30–35 | 28–31 | 25–30 | 35 |
| areola openings | most areolae are rounded or elongate elliptic; some are slit-like in the central area | small and rounded; slit-like near the margin | rounded; slit-like near the margin | rounded to elongated areolae | quadrangular or rounded |
| central area | linear, becoming a little wider in the central area | small and lanceolate to rectangular | linear, becoming a little wider in the central area | small rhombic, almost absent | narrow linear axial area slightly expanded towards the center |
| rapheless valve | |||||
| density of striae (in 10 µm) | 28–40 | 32–35 | 28–31 | 25–30 | 35 |
| areola openings | most areolae are rounded or elongated elliptic, but some are slit-like; slit-like areolae are mostly in the axial central area and more than RV | small and rounded; slit-like near the margin | rounded; slit-like near the margin | rounded to elongated | quadrangular or rounded; sometimes slit-like near the margin |
| central area | broadly lanceolate to linear and narrow | narrowly lanceolate | broadly lanceolate to linear and narrow | narrowly rhombic to lanceolate, almost absent | narrow, linear axial area slightly widening toward the central area |
| Source | this study | [6] | [24] | [24] | [16] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Miao, M.; Li, Z.; Hwang, E.-A.; Kim, H.-K.; Lee, H.; Kim, B.-H. Two New Benthic Diatoms of the Genus Achnanthidium (Bacillariophyceae) from the Hangang River, Korea. Diversity 2020, 12, 285. https://doi.org/10.3390/d12070285
AMA Style
Miao M, Li Z, Hwang E-A, Kim H-K, Lee H, Kim B-H. Two New Benthic Diatoms of the Genus Achnanthidium (Bacillariophyceae) from the Hangang River, Korea. Diversity. 2020; 12(7):285. https://doi.org/10.3390/d12070285
Chicago/Turabian StyleMiao, Minzi, Zhun Li, Eun-A Hwang, Ha-Kyung Kim, Hyuk Lee, and Baik-Ho Kim. 2020. "Two New Benthic Diatoms of the Genus Achnanthidium (Bacillariophyceae) from the Hangang River, Korea" Diversity 12, no. 7: 285. https://doi.org/10.3390/d12070285
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.