
Antioxidant and Anti-Melanogenic Activities of Heat-Treated Licorice (Wongam, Glycyrrhiza glabra × G. uralensis) Extract

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Determination of Browning
2.3. Total Phenolic Content (TPC)
2.4. Determination of Antioxidant Capacity
2.5. High-Performance Liquid Chromatography (HPLC) Analysis
2.6. Mushroom Tyrosinase Inhibitory Activity Assay
2.7. Cell Culture
2.8. Cell Viability Assay
2.9. Measurement of Cellular Melanin Content
2.10. Cellular Tyrosinase Activity Assay
2.11. Real-Time Polymerase Chain Reaction (RT-PCR)
2.12. Preparation of Cell Lysates and Western Blot Analysis
2.13. Statistical Analysis
3. Results
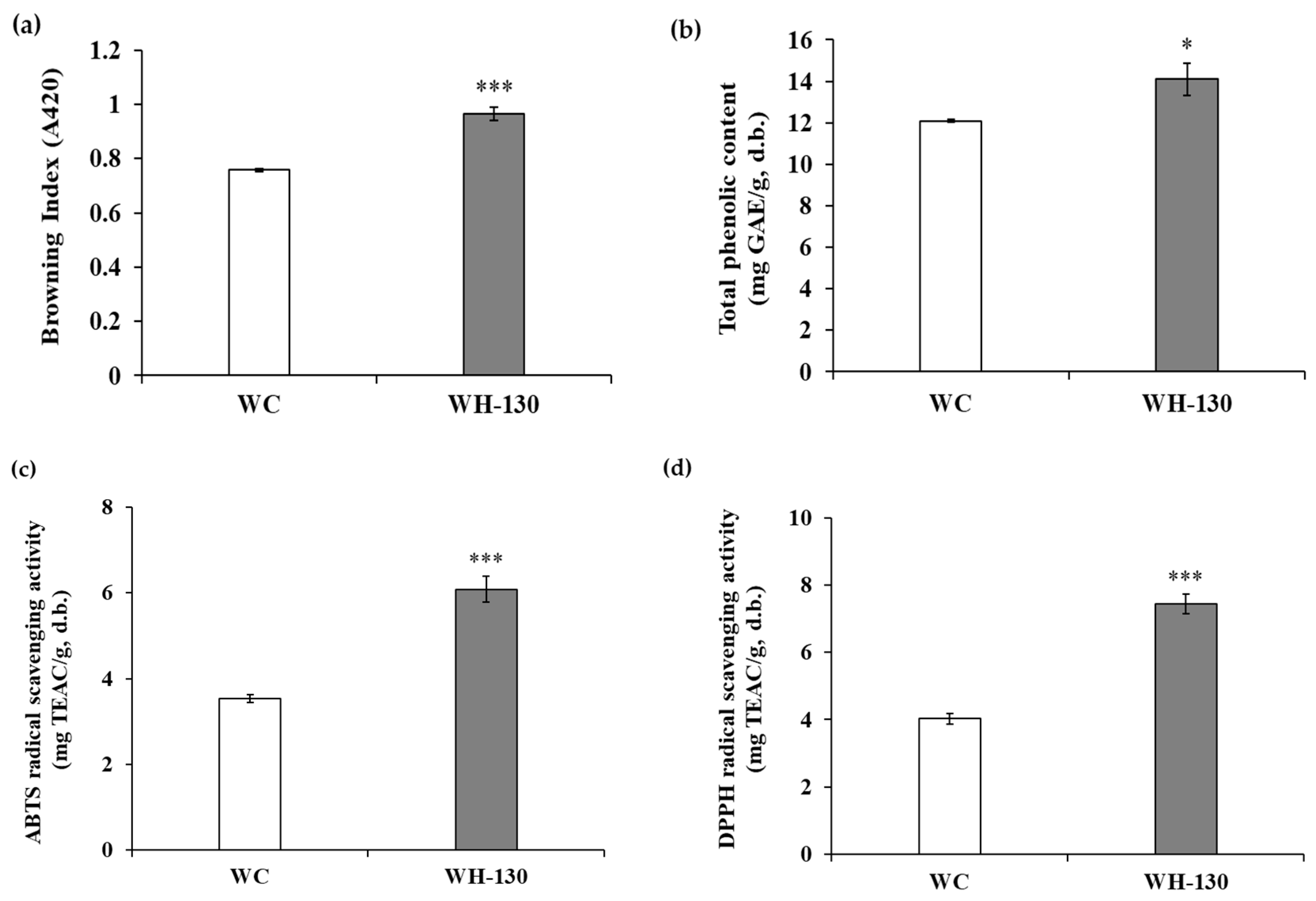
3.1. Browning Index of Licorice Extracts
3.2. Total Phenolic Content (TPC) of Licorice Extracts
3.3. Radical-Scavenging Activity of Licorice Extracts
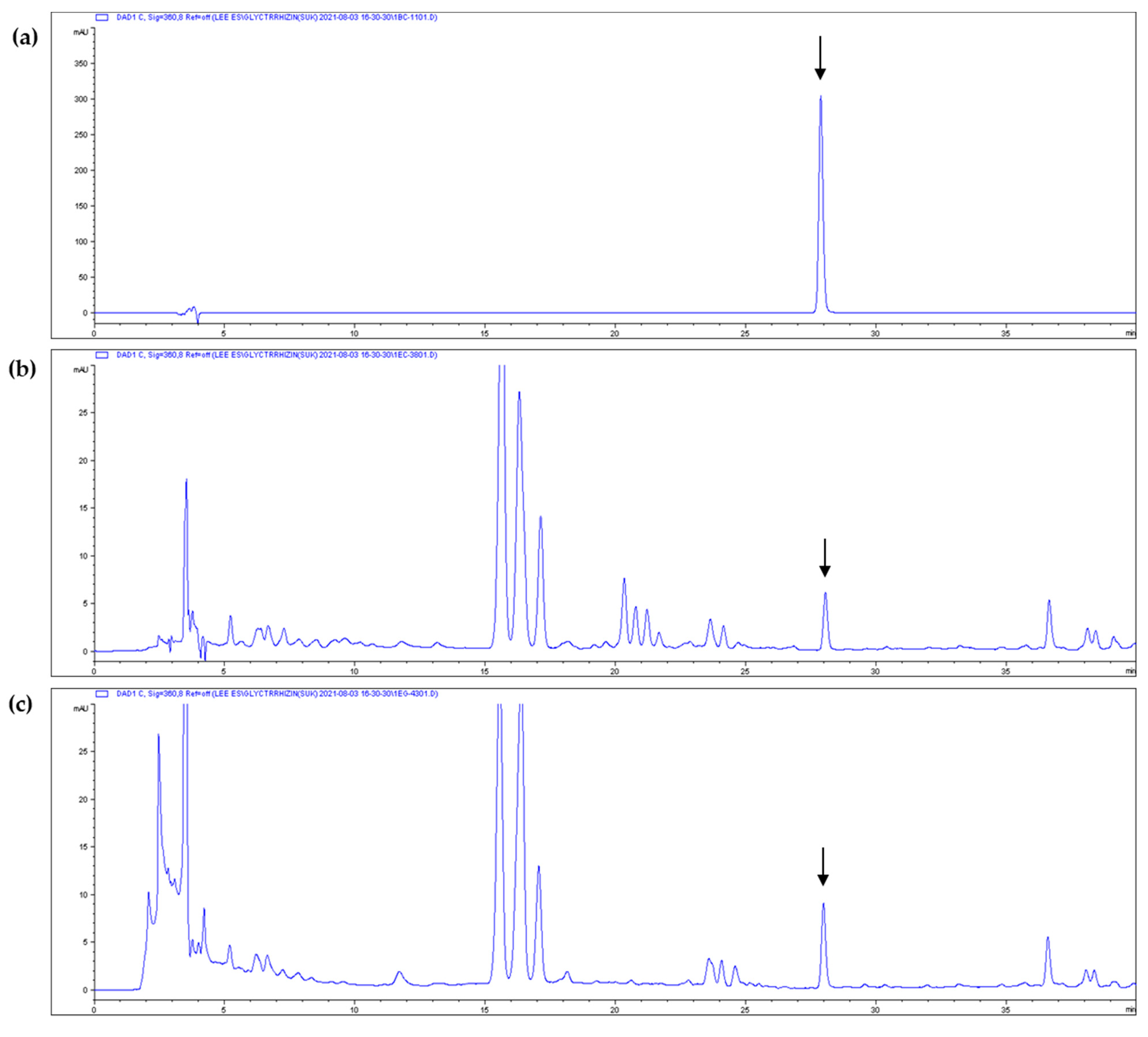
3.4. Isoliquiritigenin Content of Licorice Extracts
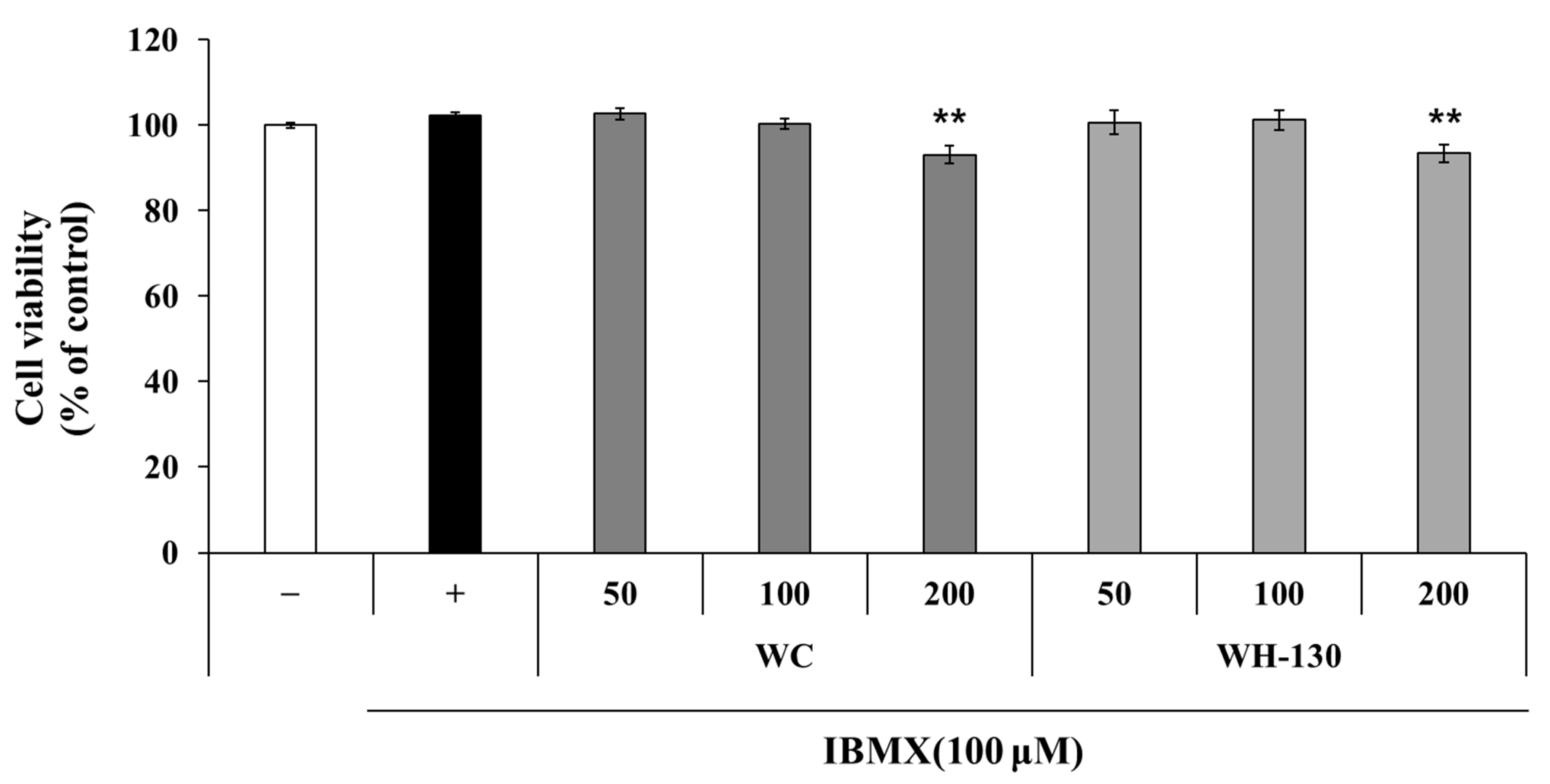
3.5. Cell Viability
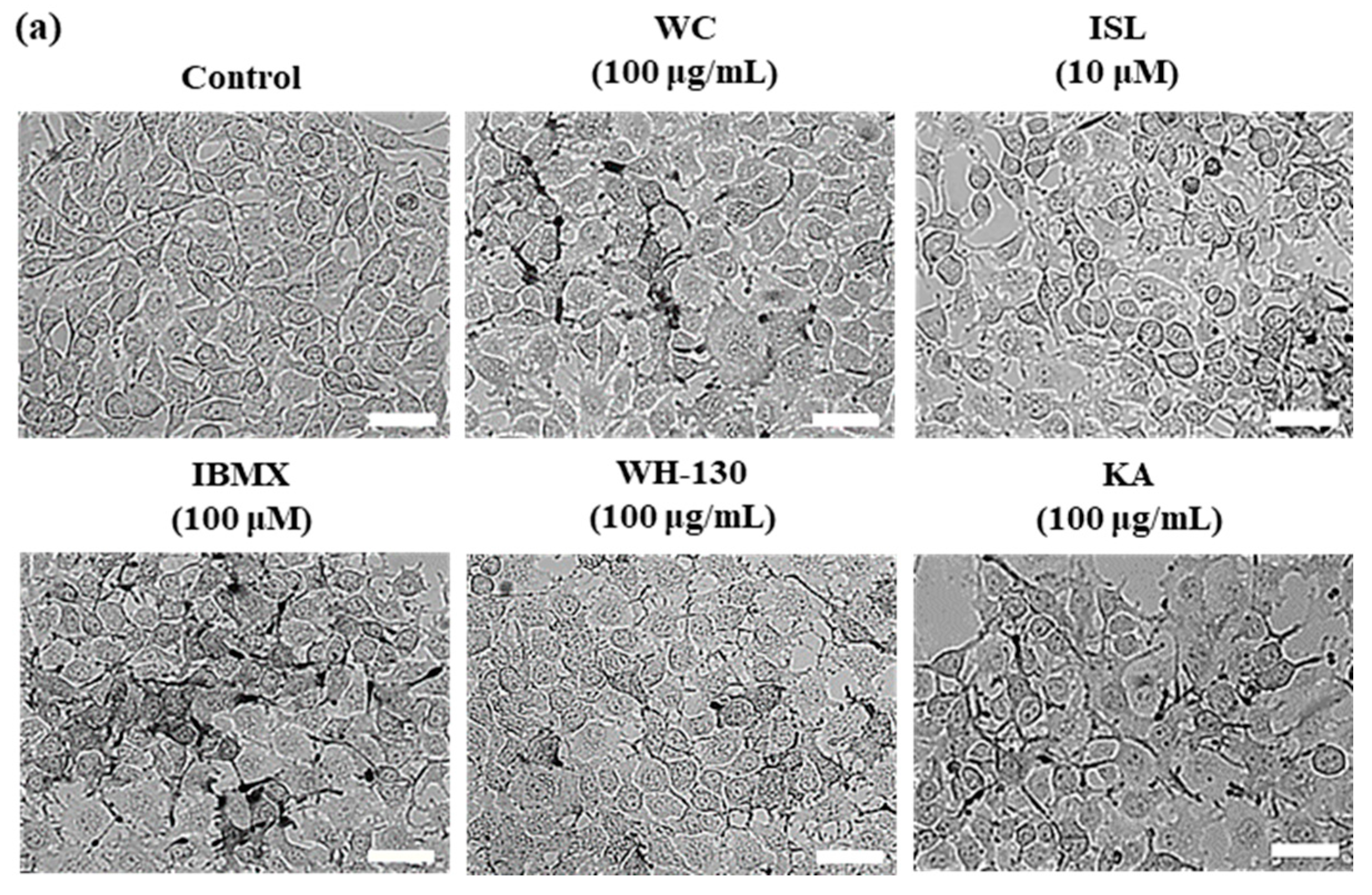
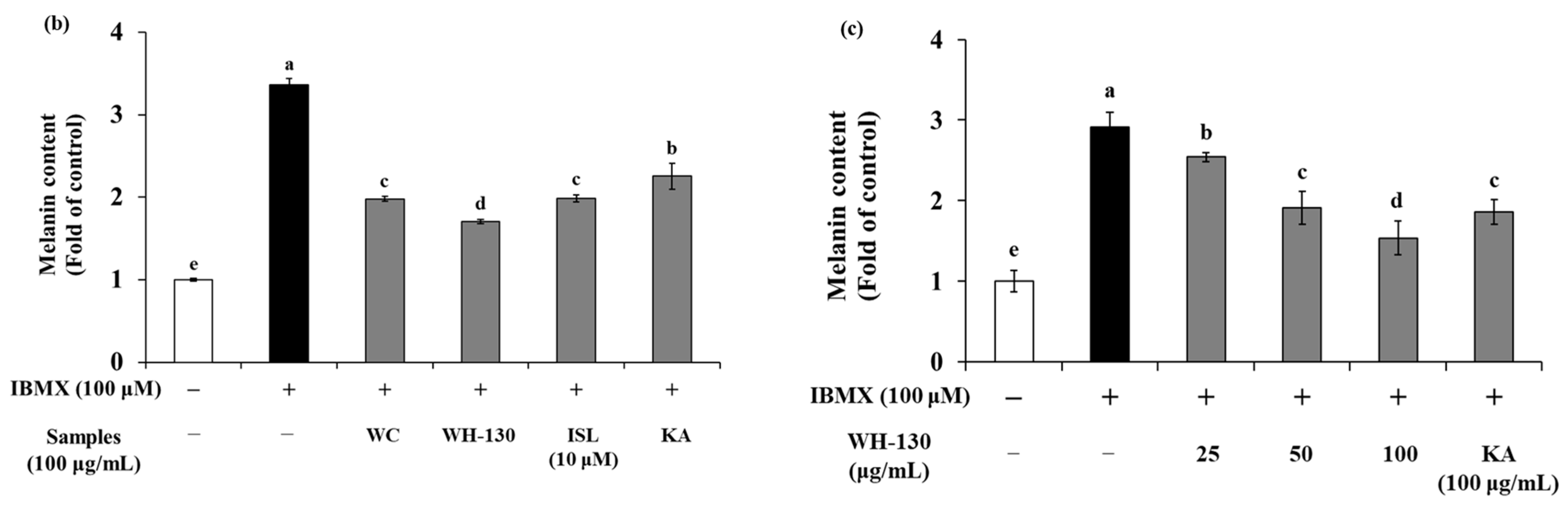
3.6. Effects of Licorice Extracts on Melanin Production in B16F10 Melanoma Cells
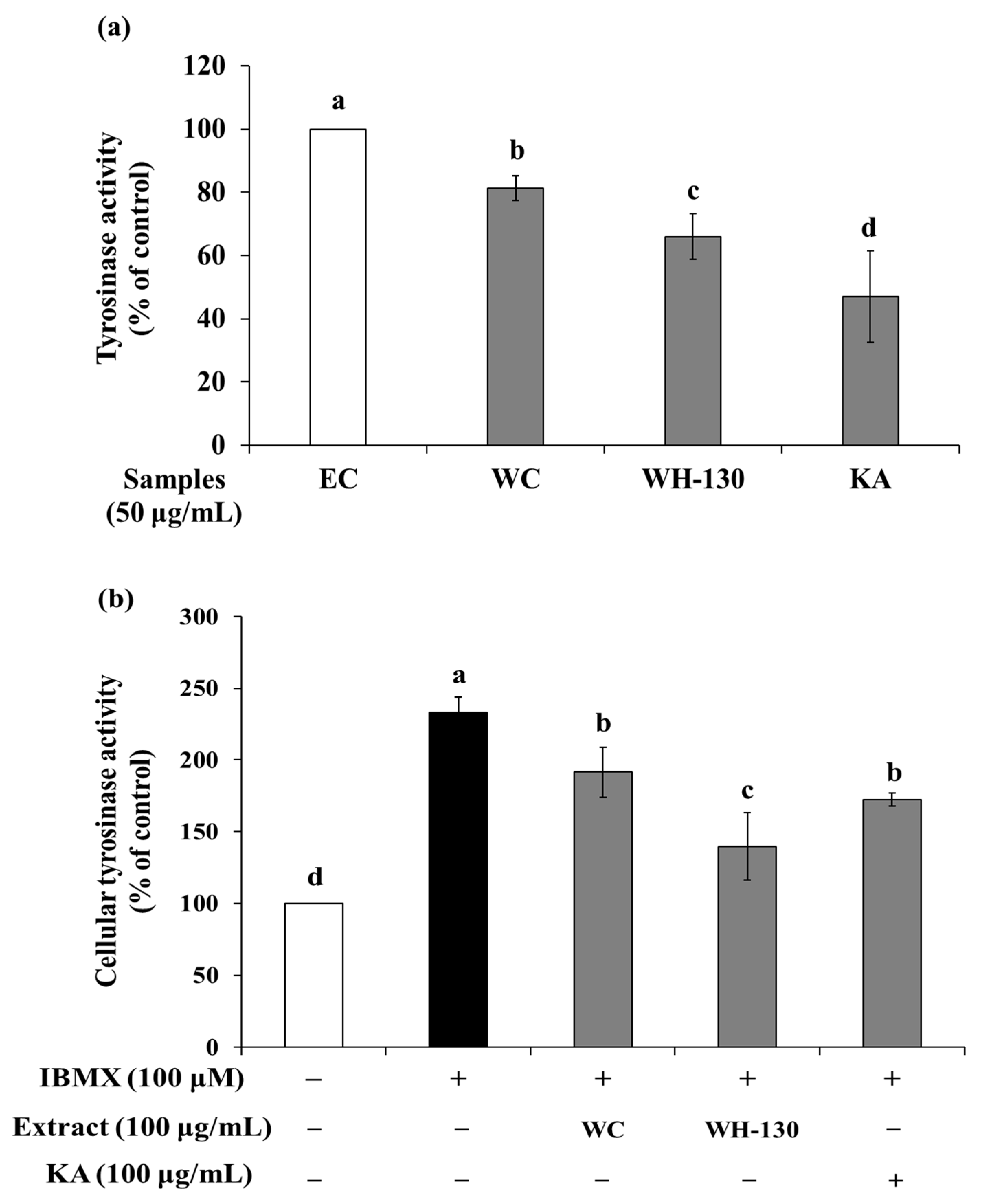
3.7. Effects of Licorice Extracts on Tyrosinase Activity
3.7.1. Mushroom Tyrosinase Activity
3.7.2. Cellular Tyrosinase Activity
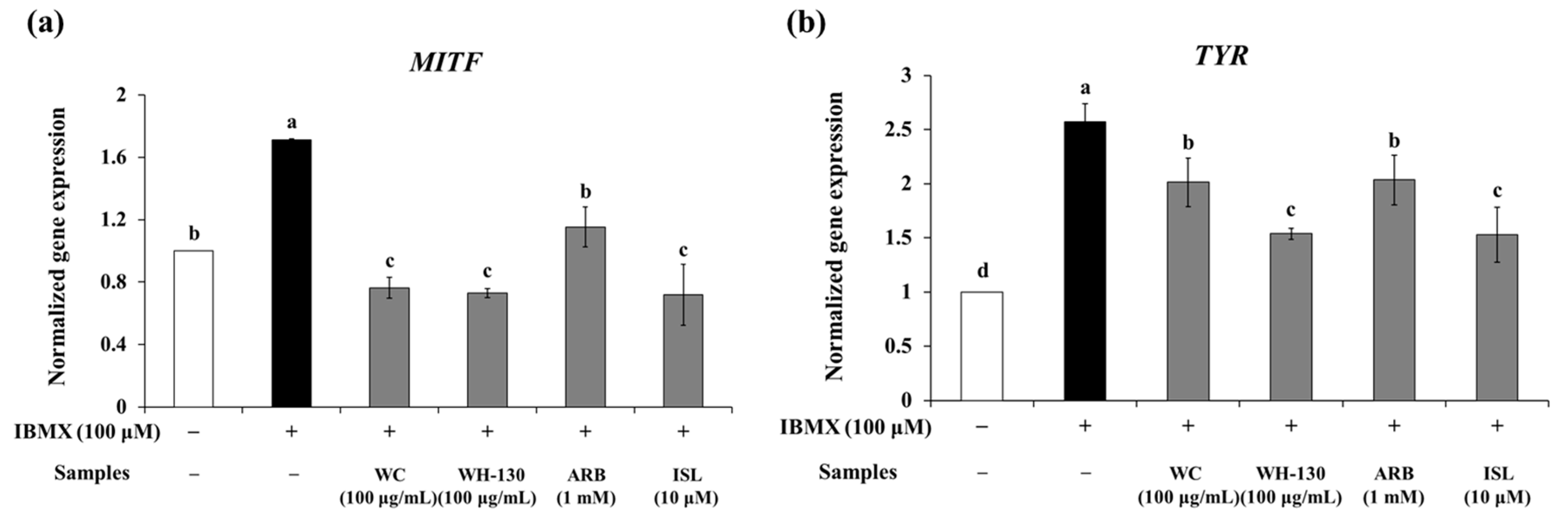
3.8. Effects of Licorice Extracts on mRNA Expression of Melanogenesis-Related Genes
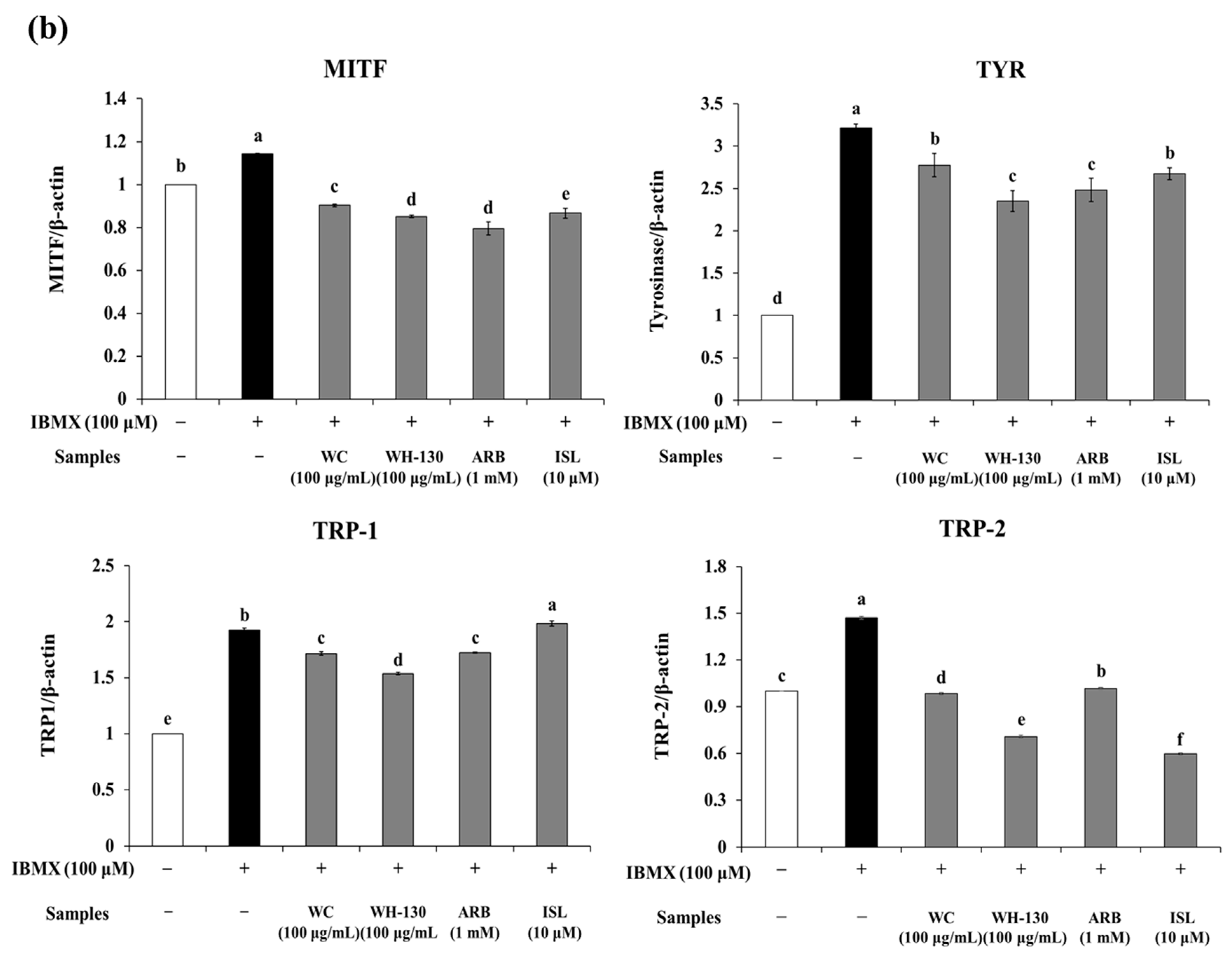
3.9. Effects of Licorice Extracts on Protein Expression of Melanogenesis-Related Genes
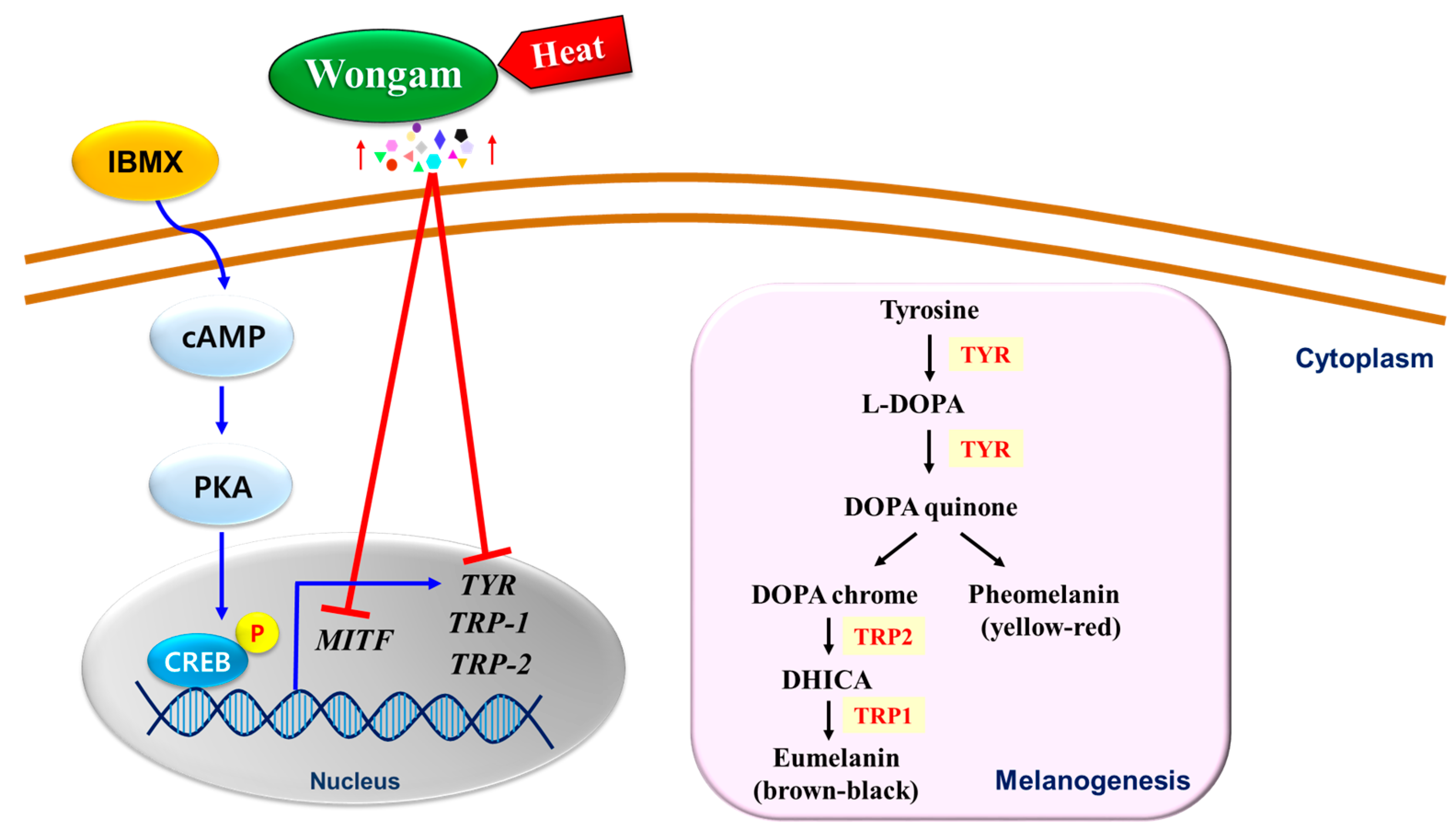
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lee, J.-H.; Oh, M.-W.; Lee, S.-H.; Park, C.-G.; Jeong, J.-T.; Han, J.-W.; Ma, K.-H.; Chang, J.-K. ‘Wongam’, a Licorice Interspecific Hybrid Cultivar with High Yield. Korean J. Breed. Sci. 2020, 52, 454–459. [Google Scholar] [CrossRef]
- Kim, D.G.; Lee, J.; Kim, W.; An, H.J.; Lee, J.H.; Chang, J.; Kang, S.H.; Song, Y.J.; Jeon, Y.D.; Jin, J.S. Assessment of General Toxicity of the Glycyrrhiza New Variety Extract in Rats. Plants 2021, 10, 1126. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.C.; Lin, Y.-H.; Chang, T.-M.; Huang, W.-Y. Identification of two licorice species, Glycyrrhiza uralensis and Glycyrrhiza glabra, based on separation and identification of their bioactive components. Food Chem. 2012, 132, 2188–2193. [Google Scholar] [CrossRef]
- Raoufinejad, K.; Rajabi, M.; Sarafian, G. Licorice in the Treatment of Acne Vulgaris and Postinflammatory Hyperpigmentation: A Review. J. Pharm. Care 2020, 8, 186–195. [Google Scholar] [CrossRef]
- Han, Y.J.; Kang, B.; Yang, E.J.; Choi, M.K.; Song, I.S. Simultaneous Determination and Pharmacokinetic Characterization of Glycyrrhizin, Isoliquiritigenin, Liquiritigenin, and Liquiritin in Rat Plasma Following Oral Administration of Glycyrrhizae Radix Extract. Molecules 2019, 24, 1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Zhu, L.; Liu, Y.; Zhou, Q.; Chen, H.; Yang, J. Isoliquiritigenin, a flavonoid from licorice, plays a dual role in regulating gastrointestinal motility in vitro and in vivo. Phytother. Res. 2009, 23, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Tanemoto, R.; Okuyama, T.; Matsuo, H.; Okumura, T.; Ikeya, Y.; Nishizawa, M. The constituents of licorice (Glycyrrhiza uralensis) differentially suppress nitric oxide production in interleukin-1beta-treated hepatocytes. Biochem. Biophys. Rep. 2015, 2, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.H.; Chiang, Y.F.; Shieh, T.M.; Chen, H.Y.; Shih, C.K.; Wang, T.H.; Wang, K.L.; Huang, T.C.; Hong, Y.H.; Li, S.C.; et al. Dietary Compound Isoliquiritigenin, an Antioxidant from Licorice, Suppresses Triple-Negative Breast Tumor Growth via Apoptotic Death Program Activation in Cell and Xenograft Animal Models. Antioxidants 2020, 9, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Huh, Y.; Lim, K.M. Anti-Pigmentary Natural Compounds and Their Mode of Action. Int. J. Mol. Sci. 2021, 22, 6206. [Google Scholar] [CrossRef]
- Lv, J.; Fu, Y.; Cao, Y.; Jiang, S.; Yang, Y.; Song, G.; Yun, C.; Gao, R. Isoliquiritigenin inhibits melanogenesis, melanocyte dendricity and melanosome transport by regulating ERK-mediated MITF degradation. Exp. Derm. 2020, 29, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Nerya, O.; Vaya, J.; Musa, R.; Izrael, S.; Ben-Arie, R.; Tamir, S. Glabrene and Isoliquiritigenin as Tyrosinase Inhibitors from Licorice Roots. J. Agric. Food Chem. 2003, 51, 1201–1207. [Google Scholar] [CrossRef]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tymińska, A. Skin melanocytes: Biology and development. Adv. Dermatol. Allergol. 2013, 1, 30–41. [Google Scholar] [CrossRef]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Park, G.; Son, H.J.; Lee, S.J. Suppression of alpha-MSH and IBMX-induced melanogenesis by cordycepin via inhibition of CREB and MITF, and activation of PI3K/Akt and ERK-dependent mechanisms. Int. J. Mol. Med. 2012, 29, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yu, X.; Huang, Y. Inhibitory mechanisms of glabridin on tyrosinase. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2016, 168, 111–117. [Google Scholar] [CrossRef]
- Alam, M.B.; Bajpai, V.K.; Lee, J.; Zhao, P.; Byeon, J.H.; Ra, J.S.; Majumder, R.; Lee, J.S.; Yoon, J.I.; Rather, I.A.; et al. Inhibition of melanogenesis by jineol from Scolopendra subspinipes mutilans via MAP-Kinase mediated MITF downregulation and the proteasomal degradation of tyrosinase. Sci. Rep. 2017, 7, 45858. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Tonissen, K.F.; Di Trapani, G. Modulating skin colour: Role of the thioredoxin and glutathione systems in regulating melanogenesis. Biosci. Rep. 2021, 41. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.H.; Yang, S.H.; Kim, D.S.; Kim, N.D.; Shin, H.J.; Liu, K. Novel Quercetin Derivative of 3,7-Dioleylquercetin Shows Less Toxicity and Highly Potent Tyrosinase Inhibition Activity. Int. J. Mol. Sci. 2021, 22, 4264. [Google Scholar] [CrossRef]
- Han, H.J.; Park, S.K.; Kang, J.Y.; Kim, J.M.; Yoo, S.K.; Heo, H.J. Anti-Melanogenic Effect of Ethanolic Extract of Sorghum bicolor on IBMX-Induced Melanogenesis in B16/F10 Melanoma Cells. Nutrients 2020, 12, 832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.J.; Malunga, L.N.; Eskin, M.; Eck, P.; Thandapilly, S.J.; Thiyam-Hollander, U. Valorization of Heat-Treated Brewers’ Spent Grain Through the Identification of Bioactive Phenolics by UPLC-PDA and Evaluation of Their Antioxidant Activities. Front. Nutr. 2021, 8, 634519. [Google Scholar] [CrossRef]
- Jeong, S.M.; Kim, S.Y.; Kim, D.R.; Jo, S.C.; Nam, K.C.; Ahn, D.U.; Lee, S.C. Effect of heat treatment on the antioxidant activity of extracts from citrus peels. J. Agric. Food Chem. 2004, 52, 3389–3393. [Google Scholar] [CrossRef]
- Park, J.-Y.; Ji, Y.-J.; Seo, K.H.; Lee, J.Y.; Kim, G.-S.; Kang, M.H.; Lee, J.H.; Jang, G.Y.; Kim, H.D. Heat Treatment Improves UV Photoprotective Effects of Licorice in Human Dermal Fibroblasts. Processes 2021, 9, 1040. [Google Scholar] [CrossRef]
- Echavarría, A.P.; Pagán, J.; Ibarz, A. Melanoidins Formed by Maillard Reaction in Food and Their Biological Activity. Food Eng. Rev. 2012, 4, 203–223. [Google Scholar] [CrossRef]
- Starowicz, M.; Ostaszyk, A.; Zielinski, H. The Relationship between the Browning Index, Total Phenolics, Color, and Antioxidant Activity of Polish-Originated Honey Samples. Foods 2021, 10, 967. [Google Scholar] [CrossRef] [PubMed]
- Blainski, A.; Lopes, G.C.; de Mello, J.C. Application and analysis of the folin ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilas-Boas, A.A.; Oliveira, A.; Ribeiro, T.B.; Ribeiro, S.; Nunes, C.; Gómez-García, R.; Nunes, J.; Pintado, M. Impact of Extraction Process in Non-Compliant ‘Bravo de Esmolfe’ Apples towards the Development of Natural Antioxidant Extracts. Appl. Sci. 2021, 11, 5916. [Google Scholar] [CrossRef]
- An, S.M.; Lee, S.I.; Choi, S.W.; Moon, S.W.; Boo, Y.C. p-Coumaric acid, a constituent of Sasa quelpaertensis Nakai, inhibits cellular melanogenesis stimulated by alpha-melanocyte stimulating hormone. Br. J. Derm. 2008, 159, 292–299. [Google Scholar] [CrossRef]
- Li, H.; DaSilva, N.A.; Liu, W.; Xu, J.; Dombi, G.W.; Dain, J.A.; Li, D.; Chamcheu, J.C.; Seeram, N.P.; Ma, H. Thymocid®, a Standardized Black Cumin (Nigella sativa) Seed Extract, Modulates Collagen Cross-Linking, Collagenase and Elastase Activities, and Melanogenesis in Murine B16F10 Melanoma Cells. Nutrients 2020, 12, 2146. [Google Scholar] [CrossRef] [PubMed]
- Colasanto, A.; Travaglia, F.; Bordiga, M.; Monteduro, S.; Arlorio, M.; Coisson, J.D.; Locatelli, M. Cooking of Artemide Black Rice: Impact on Proximate Composition and Phenolic Compounds. Foods 2021, 10, 824. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, J.Y.; Kim, J.-M.; Hong, I.K.; Oh, S.-G. Preparation of melanoidin/TiO2 composites with improved UV screening ability and suppressed photocatalytic activity for cosmetic application. J. Ind. Eng. Chem. 2020, 86, 186–193. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, J.; Chen, S.; Xue, J.; Huang, S.; Wang, Y.; Chen, O. Isoliquiritigenin attenuates lipopolysaccharide-induced cognitive impairment through antioxidant and anti-inflammatory activity. BMC Neurosci. 2019, 20, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Cortés, A.; Sánchez-Motta, T.; Tobar-Tosse, F.; Quimbaya, M. Spectrophotometric Estimation of Total Phenolic Content and Antioxidant Capacity of Molasses and Vinasses Generated from the Sugarcane Industry. Waste Biomass Valorization 2019, 11, 3453–3463. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Li, H.; Li, Y.; Li, M.; Chen, G. Effect of isoliquiritigenin for the treatment of atopic dermatitis-like skin lesions in mice. Arch. Derm. Res. 2017, 309, 805–813. [Google Scholar] [CrossRef]
- Jung, H.J.; Choi, D.C.; Noh, S.G.; Choi, H.; Choi, I.; Ryu, I.Y.; Chung, H.Y.; Moon, H.R. New Benzimidazothiazolone Derivatives as Tyrosinase Inhibitors with Potential Anti-Melanogenesis and Reactive Oxygen Species Scavenging Activities. Antioxidants 2021, 10, 1078. [Google Scholar] [CrossRef]
- Yokota, T.; Nishio, H.; Kubota, Y.; Mizoguchi, M. The Inhibitory Effect of Glabridin from Licorice Extracts on Melanogenesis and Inflammation. Pigment. Cell Res. 1998, 11, 355–361. [Google Scholar] [CrossRef]
- Chung, Y.C.; Ko, J.H.; Kang, H.K.; Kim, S.; Kang, C.I.; Lee, J.N.; Park, S.M.; Hyun, C.G. Antimelanogenic Effects of Polygonum tinctorium Flower Extract from Traditional Jeju Fermentation via Upregulation of Extracellular Signal-Regulated Kinase and Protein Kinase B Activation. Int. J. Mol. Sci. 2018, 19, 2895. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Alam, M.B.; An, H.; Choi, H.J.; Cha, Y.H.; Yoo, C.Y.; Kim, H.H.; Lee, S.H. Antimelanogenic Effect of an Oroxylum indicum Seed Extract by Suppression of MITF Expression through Activation of MAPK Signaling Protein. Int. J. Mol. Sci. 2018, 19, 760. [Google Scholar] [CrossRef] [Green Version]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.L.; Qadir, A.S.; Park, H.J.; Chung, E.; Lee, Y.S.; Woo, K.M.; Ryoo, H.M.; Kim, H.J.; Baek, J.H. cAMP/Protein Kinase A Signaling Inhibits Dlx5 Expression via Activation of CREB and Subsequent C/EBPbeta Induction in 3T3-L1 Preadipocytes. Int. J. Mol. Sci. 2018, 19, 3161. [Google Scholar] [CrossRef] [Green Version]
- Skroza, D.; Generalić Mekinić, I.; Svilović, S.; Šimat, V.; Katalinić, V. Investigation of the potential synergistic effect of resveratrol with other phenolic compounds: A case of binary phenolic mixtures. J. Food Compos. Anal. 2015, 38, 13–18. [Google Scholar] [CrossRef]
- Sathya, S.; Herath, H.M.D.R.; Amarasinghe, N.R.; Suraweera, R.K. Formulation development of cream incorporating extracts of Glycyrrhiza glabra (Licorice). Pharm. J. Sri Lanka 2017, 7, 44–50. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence | |
|---|---|---|
| MITF 1 | Forward | GGCCAAGGCAGAGCAACTT |
| Reverse | GCCCATGGTGGCAAGCT | |
| Tyrosinase | Forward | ATAGGTGCATTGGCTTCTGG |
| Reverse | CCAACGATCCCATTTTTCTT | |
| TRP-12 | Forward | GAGTGACATCCTGTGGCTCA |
| Reverse | CGATACCCTGGGAACACTTT | |
| TRP-2 | Forward | GCTCCAAGTGGCTGTAGACC |
| Reverse | AATGCAGTGGCTTGGAAATC | |
| β-actin | Forward | CCCACTCCTAAGAGGAGGATG |
| Reverse | AGGGAGACCAAAGCCTTCAT |
| Sample | Isoliquiritigenin (mg/g Extract, d.b.) |
|---|---|
| WC | 1.354 ± 0.144 a |
| WH-130 | 2.492 ± 0.152 b |
| Factors | TPC | ISL | ABTS+ | DPPH | TYR | Melanin |
|---|---|---|---|---|---|---|
| TPC 1 | 1.000 |  |  |  |  |  |
| ISL 2 | 0.827 * | 1.000 |  |  |  |  |
| ABTS+ 3 | 0.886 * | 0.977 *** | 1.000 |  |  |  |
| DPPH 3 | 0.896 * | 0.985 *** | 0.998 *** | 1.000 |  |  |
| TYR 4 | −0.662 | −0.834 * | −0.879 * | −0.856 * | 1.000 |  |
| Melanin 5 | −0.859 * | −0.995 *** | −0.974 ** | −0.984 *** | 0.821 * | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, M.H.; Jang, G.Y.; Ji, Y.-J.; Lee, J.H.; Choi, S.J.; Hyun, T.K.; Kim, H.D. Antioxidant and Anti-Melanogenic Activities of Heat-Treated Licorice (Wongam, Glycyrrhiza glabra × G. uralensis) Extract. Curr. Issues Mol. Biol. 2021, 43, 1171-1187. https://doi.org/10.3390/cimb43020083
Kang MH, Jang GY, Ji Y-J, Lee JH, Choi SJ, Hyun TK, Kim HD. Antioxidant and Anti-Melanogenic Activities of Heat-Treated Licorice (Wongam, Glycyrrhiza glabra × G. uralensis) Extract. Current Issues in Molecular Biology. 2021; 43(2):1171-1187. https://doi.org/10.3390/cimb43020083
Chicago/Turabian StyleKang, Min Hye, Gwi Yeong Jang, Yun-Jeong Ji, Jeong Hoon Lee, Su Ji Choi, Tae Kyung Hyun, and Hyung Don Kim. 2021. "Antioxidant and Anti-Melanogenic Activities of Heat-Treated Licorice (Wongam, Glycyrrhiza glabra × G. uralensis) Extract" Current Issues in Molecular Biology 43, no. 2: 1171-1187. https://doi.org/10.3390/cimb43020083