Yeast as a Model to Understand Actin-Mediated Cellular Functions in Mammals—Illustrated with Four Actin Cytoskeleton Proteins

 , , , , and
, , , , and
Abstract
:1. Introduction
1.1. Yeast as a Model Organism
1.2. Actin Cytoskeleton in Yeast
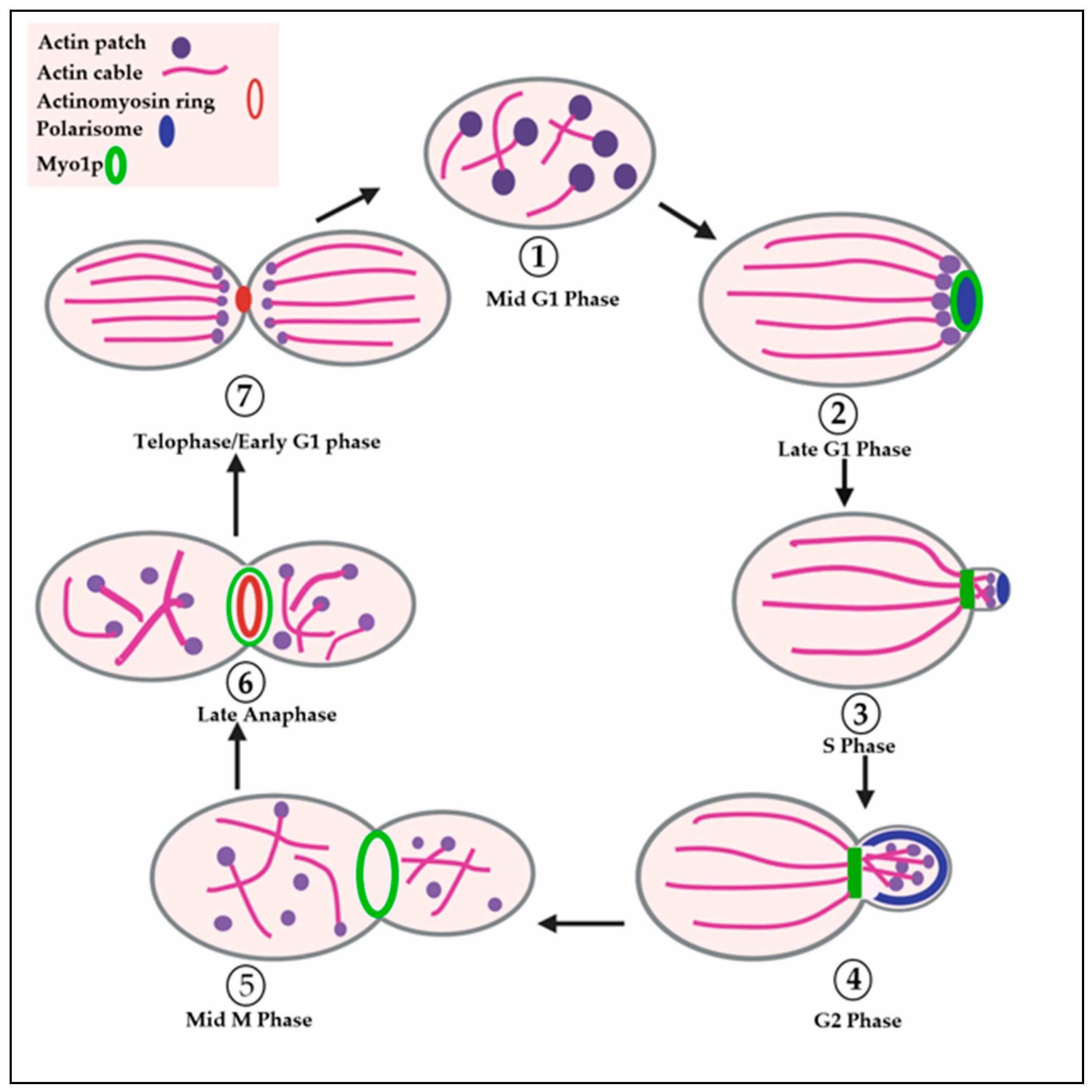
1.3. Polarization of the Actin Cytoskeleton during the Budding Cycle of Yeast
1.4. Role of Septins in Defining the Nascent Bud Site and Bud Neck in Yeast
1.5. Formation of Cortical Actin Patches and Their Function in Endocytosis
1.6. Overview of Cytokinesis in Yeast and Mammals
1.7. Initiation of Actomyosin Contractile Ring Assembly in Yeast
1.8. Role of the Formins Bni1p and Bnr1p in Actomyosin Ring Assembly in Yeast
1.9. Contraction of the Actomyosin Ring
1.10. Septum Formation during Cytokinesis in Yeast
1.11. Septum Degradation and Cell Separation in Yeast
2. Budding Yeast Hof1p and Human PSTPIP1
2.1. The Function of Budding Yeast Hof1p and Fission Yeast Cdc15 and Interactions of the Hof1p SH3 Domain
2.2. The Function of Mammalian PSTPIP1 and Interactions of the PSTPIP1 SH3 Domain
2.3. How the Yeast Model Can Provide Insight into the Function of PSTPIP1 in Mammals
3. Communication between the Actin Cytoskeleton and Protein Synthesis Machinery
3.1. Links between Actin and Translation
3.2. Gcn2 Function
3.3. Reciprocal Regulation of eEF1A and Actin, and the Link to Gcn2
3.4. Regulation of Gcn2 by Yih1p/IMPACT and the Link to Actin
3.5. Gcn2 is an Important Sensor of the State of the Actin Cytoskeleton
3.6. The GCN2-Actin Regulatory Axis May Have a Wide-Reaching Relevance
4. The Yeast and Human Amphiphysins and Their Link to Actin-Based Cellular Functions
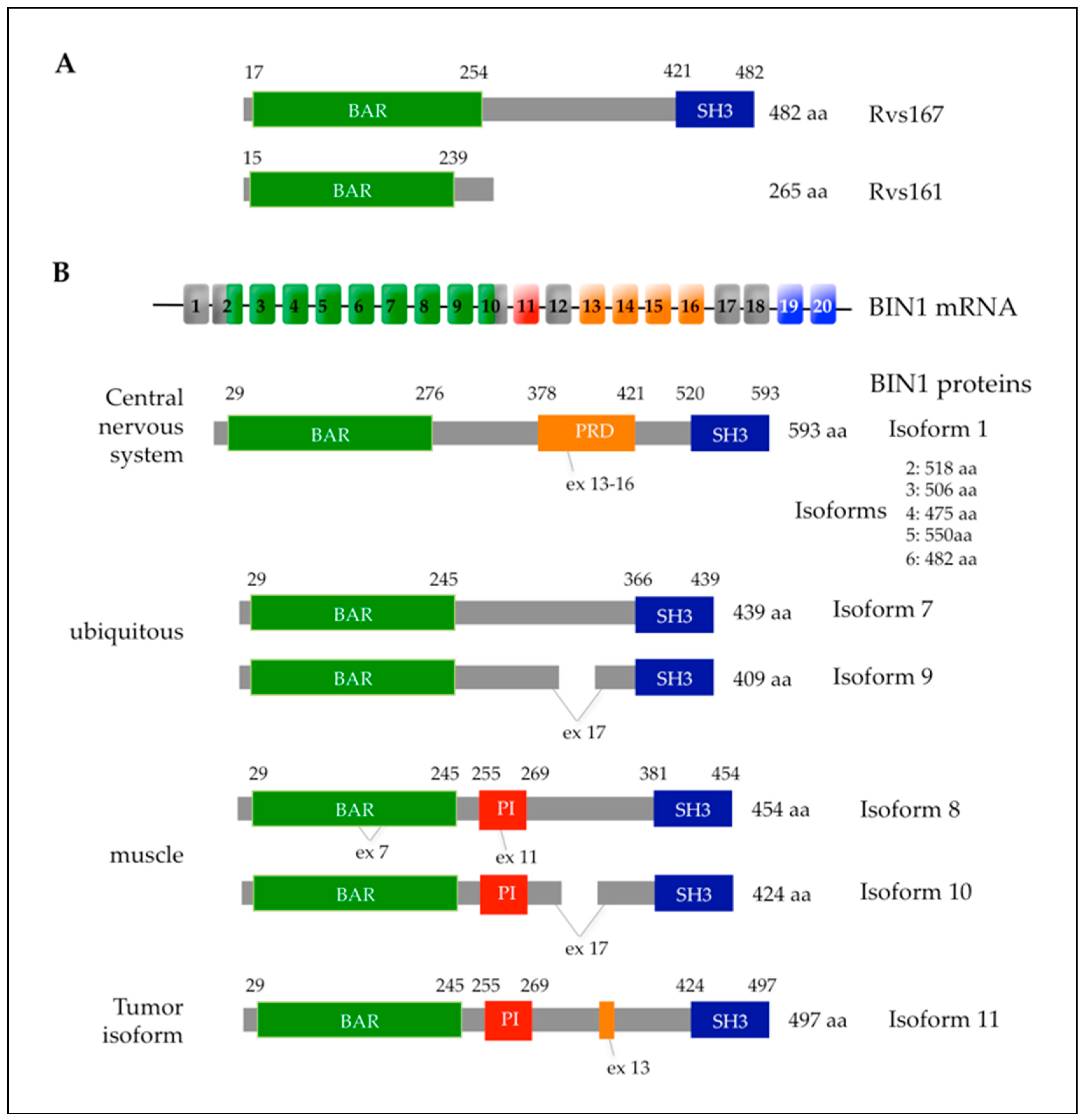
4.1. The Yeast Rvs161p and Rvs167p Amphiphysins, Key Regulators of Actin-Dependent Endocytosis
4.2. The AMPH1 and BIN1 Human Amphiphysins and Their Link to Actin Cytoskeleton
4.3. BIN1 Associated Diseases and Their Link with Actin-Based Functions
4.3.1. Cancer
4.3.2. Centronuclear Myopathies
4.3.3. Alzheimer’s Disease
5. Concluding Remarks and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Cherry, J.M.; Hong, E.L.; Amundsen, C.; Balakrishnan, R.; Binkley, G.; Chan, E.T.; Christie, K.R.; Costanzo, M.C.; Dwight, S.S.; Engel, S.R.; et al. Saccharomyces Genome Database: The genomics resource of budding yeast. Nucleic Acids Res. 2012, 40, D700–D705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broach, J.R.; Pringle, J.R.; Jones, E.W. (Eds.) The Molecular and Cellular Biology of the Yeast Saccharomyces; Volume 1 Genome Dynamics, Protein Synthesis, and Energetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1991. [Google Scholar]
- Jones, E.W.; Pringle, J.R.; Broach, J.R. (Eds.) The Molecular and Cellular Biology of the Yeast Saccharomyces; Volume II Gene Expression; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1992. [Google Scholar]
- Pringle, J.R.; Broach, J.R.; Jones, E.W. (Eds.) The Molecular and Cellular Biology of the Yeast Saccharomyces; Volume III Cell Cycle and Cell Biology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Guide to Yeast Genetics and Molecular Biology. Method Enzymol. 1991, 194, 1–863.
- Mnaimneh, S.; Davierwala, A.P.; Haynes, J.; Moffat, J.; Peng, W.-T.; Zhang, W.; Yang, X.; Pootoolal, J.; Chua, G.; Lopez, A. Exploration of essential gene functions via titratable promoter alleles. Cell 2004, 118, 31–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.-X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A. Genome evolution across 1011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.; Jacq, C.; Johnston, M. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [Green Version]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef]
- Chong, Y.T.; Koh, J.L.; Friesen, H.; Duffy, S.K.; Cox, M.J.; Moses, A.; Moffat, J.; Boone, C.; Andrews, B.J. Yeast proteome dynamics from single cell imaging and automated analysis. Cell 2015, 161, 1413–1424. [Google Scholar] [CrossRef] [Green Version]
- Huh, W.-K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef]
- Ball, C.A.; Jin, H.; Sherlock, G.; Weng, S.; Matese, J.C.; Andrada, R.; Binkley, G.; Dolinski, K.; Dwight, S.S.; Harris, M.A. Saccharomyces Genome Database provides tools to survey gene expression and functional analysis data. Nucleic Acids Res. 2001, 29, 80–81. [Google Scholar] [CrossRef] [Green Version]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [Green Version]
- Baryshnikova, A.; Costanzo, M.; Myers, C.L.; Andrews, B.; Boone, C. Genetic interaction networks: Toward an understanding of heritability. Annu. Rev. Genom. Hum. Genet. 2013, 14, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Bader, G.D.; Donaldson, I.; Wolting, C.; Ouellette, B.F.; Pawson, T.; Hogue, C.W. BIND—the biomolecular interaction network database. Nucleic Acids Res. 2001, 29, 242–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.; Gruhler, A.; Heilbut, A.; Bader, G.D.; Moore, L.; Adams, S.-L.; Millar, A.; Taylor, P.; Bennett, K.; Boutilier, K. Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. Nature 2002, 415, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavin, A.-C.; Aloy, P.; Grandi, P.; Krause, R.; Boesche, M.; Marzioch, M.; Rau, C.; Jensen, L.J.; Bastuck, S.; Dümpelfeld, B. Proteome survey reveals modularity of the yeast cell machinery. Nature 2006, 440, 631–636. [Google Scholar] [CrossRef]
- Gavin, A.-C.; Bösche, M.; Krause, R.; Grandi, P.; Marzioch, M.; Bauer, A.; Schultz, J.; Rick, J.M.; Michon, A.-M.; Cruciat, C.-M. Functional organization of the yeast proteome by systematic analysis of protein complexes. Nature 2002, 415, 141–147. [Google Scholar] [CrossRef]
- Fromont-Racine, M.; Rain, J.-C.; Legrain, P. Toward a functional analysis of the yeast genome through exhaustive two-hybrid screens. Nat. Genet. 1997, 16, 277–282. [Google Scholar] [CrossRef]
- Krogan, N.J.; Cagney, G.; Yu, H.; Zhong, G.; Guo, X.; Ignatchenko, A.; Li, J.; Pu, S.; Datta, N.; Tikuisis, A.P. Global landscape of protein complexes in the yeast Saccharomyces cerevisiae. Nature 2006, 440, 637–643. [Google Scholar] [CrossRef]
- Hwang, K.S.; Kim, H.U.; Charusanti, P.; Palsson, B.O.; Lee, S.Y. Systems biology and biotechnology of Streptomyces species for the production of secondary metabolites. Biotechnol. Adv. 2014, 32, 255–268. [Google Scholar] [CrossRef]
- Piotrowski, J.S.; Li, S.C.; Deshpande, R.; Simpkins, S.W.; Nelson, J.; Yashiroda, Y.; Barber, J.M.; Safizadeh, H.; Wilson, E.; Okada, H.; et al. Functional annotation of chemical libraries across diverse biological processes. Nat. Chem. Biol. 2017, 13, 982–993, Errata in 2017, 13, 1286. [Google Scholar] [CrossRef]
- Kachroo, A.H.; Laurent, J.M.; Yellman, C.M.; Meyer, A.G.; Wilke, C.O.; Marcotte, E.M. Systematic humanization of yeast genes reveals conserved functions and genetic modularity. Science 2015, 348, 921–925. [Google Scholar] [CrossRef] [Green Version]
- Harrison, A.F.; Shorter, J. RNA-binding proteins with prion-like domains in health and disease. Biochem. J. 2017, 474, 1417–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couthouis, J.; Hart, M.P.; Shorter, J.; DeJesus-Hernandez, M.; Erion, R.; Oristano, R.; Liu, A.X.; Ramos, D.; Jethava, N.; Hosangadi, D. A yeast functional screen predicts new candidate ALS disease genes. Proc. Natl. Acad. Sci. USA 2011, 108, 20881–20890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmetz, L.M.; Scharfe, C.; Deutschbauer, A.M.; Mokranjac, D.; Herman, Z.S.; Jones, T.; Chu, A.M.; Giaever, G.; Prokisch, H.; Oefner, P.J. Systematic screen for human disease genes in yeast. Nat. Genet. 2002, 31, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Sun, S.; Tan, G.; Costanzo, M.; Hill, D.E.; Vidal, M.; Andrews, B.J.; Boone, C.; Roth, F.P. Identifying pathogenicity of human variants via paralog-based yeast complementation. PLoS Genet. 2017, 13, e1006779. [Google Scholar] [CrossRef]
- Riezman, H.; Munn, A.; Geli, M.; Hicke, L. Actin-, myosin-and ubiquitin-dependent endocytosis. Cell. Mol. Life Sci. 1996, 52, 1033–1041. [Google Scholar] [CrossRef]
- Geli, M.I.; Riezman, H. Endocytic internalization in yeast and animal cells: Similar and different. J. Cell Sci. 1998, 111, 1031–1037. [Google Scholar]
- Munn, A.L. Molecular requirements for the internalisation step of endocytosis: Insights from yeast. BBA Mol. Basis Dis. 2001, 1535, 236–257. [Google Scholar] [CrossRef] [Green Version]
- Engqvist-Goldstein, Å.E.; Drubin, D.G. Actin assembly and endocytosis: From yeast to mammals. Annu. Rev. Cell Dev. Biol. 2003, 19, 287–332. [Google Scholar] [CrossRef] [Green Version]
- Moseley, J.B.; Goode, B.L. The yeast actin cytoskeleton: From cellular function to biochemical mechanism. Microbiol. Mol. Biol. Rev. 2006, 70, 605–645. [Google Scholar] [CrossRef] [Green Version]
- Kaksonen, M.; Toret, C.P.; Drubin, D.G. Harnessing actin dynamics for clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2006, 7, 404–414. [Google Scholar] [CrossRef]
- Sattlegger, E.; Chernova, T.A.; Gogoi, N.M.; Pillai, I.V.; Chernoff, Y.O.; Munn, A.L. Yeast studies reveal moonlighting functions of the ancient actin cytoskeleton. IUBMB Life 2014, 66, 538–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhavsar-Jog, Y.P.; Bi, E. Mechanics and regulation of cytokinesis in budding yeast. Semin. Cell Dev. Biol. 2017, 66, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.; Pringle, J.R. Relationship of actin and tubulin distribution to bud growth in wild-type and morphogenetic-mutant Saccharomyces cerevisiae. J. Cell Biol. 1984, 98, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Kilmartin, J.; Adams, A. Structural rearrangements of tubulin and actin during the cell cycle of the yeast Saccharomyces. J. Cell Biol. 1984, 98, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Shortle, D.; Novick, P.; Botstein, D. Construction and genetic characterization of temperature-sensitive mutant alleles of the yeast actin gene. Proc. Natl. Acad. Sci. USA 1984, 81, 4889–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandekerckhove, J.; Weber, K. At least six different actins are expressed in a higher mammal: An analysis based on the amino acid sequence of the amino-terminal tryptic peptide. J. Mol. Biol. 1978, 126, 783–802. [Google Scholar] [CrossRef]
- Harborth, J.; Elbashir, S.M.; Bechert, K.; Tuschl, T.; Weber, K. Identification of essential genes in cultured mammalian cells using small interfering RNAs. J. Cell Sci. 2001, 114, 4557–4565. [Google Scholar]
- Laporte, D.; Zhao, R.; Wu, J.-Q. Mechanisms of contractile-ring assembly in fission yeast and beyond. Semin. Cell Dev. Biol. 2010, 21, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Bi, E.; Maddox, P.; Lew, D.J.; Salmon, E.; McMillan, J.N.; Yeh, E.; Pringle, J.R. Involvement of an actomyosin contractile ring in Saccharomyces cerevisiae cytokinesis. J. Cell Biol. 1998, 142, 1301–1312. [Google Scholar] [CrossRef] [Green Version]
- Lippincott, J.; Li, R. Sequential assembly of myosin II, an IQGAP-like protein, and filamentous actin to a ring structure involved in budding yeast cytokinesis. J. Cell Biol. 1998, 140, 355–366. [Google Scholar] [CrossRef]
- Epp, J.A.; Chant, J. An IQGAP-related protein controls actin-ring formation and cytokinesis in yeast. Curr. Biol. 1997, 7, 921–929. [Google Scholar] [CrossRef] [Green Version]
- VerPlank, L.; Li, R. Cell cycle-regulated trafficking of Chs2 controls actomyosin ring stability during cytokinesis. Mol. Biol. Cell 2005, 16, 2529–2543. [Google Scholar] [CrossRef]
- Roh, D.-H.; Bowers, B.; Schmidt, M.; Cabib, E. The septation apparatus, an autonomous system in budding yeast. Mol. Biol. Cell 2002, 13, 2747–2759. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Bowers, B.; Varma, A.; Roh, D.-H.; Cabib, E. In budding yeast, contraction of the actomyosin ring and formation of the primary septum at cytokinesis depend on each other. J. Cell Sci. 2002, 115, 293–302. [Google Scholar]
- Dobbelaere, J.; Barral, Y. Spatial coordination of cytokinetic events by compartmentalization of the cell cortex. Science 2004, 305, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Barr, F.A.; Gruneberg, U. Cytokinesis: Placing and making the final cut. Cell 2007, 131, 847–860. [Google Scholar] [CrossRef] [Green Version]
- Meitinger, F.; Palani, S. Actomyosin ring driven cytokinesis in budding yeast. Semin. Cell Dev. Biol. 2016, 53, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolliday, N.; VerPlank, L.; Li, R. Rho1 directs formin-mediated actin ring assembly during budding yeast cytokinesis. Curr. Biol. 2002, 12, 1864–1870. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, M.K.; Bi, E.; Glotzer, M. Comparative analysis of cytokinesis in budding yeast, fission yeast and animal cells. Curr. Biol. 2004, 14, R806–R818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huckaba, T.M.; Gay, A.C.; Pantalena, L.F.; Yang, H.-C.; Pon, L.A. Live cell imaging of the assembly, disassembly, and actin cable–dependent movement of endosomes and actin patches in the budding yeast, Saccharomyces cerevisiae. J. Cell Biol. 2004, 167, 519–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehrenbacher, K.L.; Yang, H.-C.; Gay, A.C.; Huckaba, T.M.; Pon, L.A. Live cell imaging of mitochondrial movement along actin cables in budding yeast. Curr. Biol. 2004, 14, 1996–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruyne, D.; Bretscher, A. Polarization of cell growth in yeast. J. Cell Sci. 2000, 113, 571–585. [Google Scholar] [PubMed]
- Pruyne, D.W.; Schott, D.H.; Bretscher, A. Tropomyosin-containing actin cables direct the Myo2p-dependent polarized delivery of secretory vesicles in budding yeast. J. Cell Biol. 1998, 143, 1931–1945. [Google Scholar] [CrossRef] [Green Version]
- Gonsalvez, G.B.; Urbinati, C.R.; Long, R.M. RNA localization in yeast: Moving towards a mechanism. Biol. Cell 2005, 97, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takizawa, P.A.; Sil, A.; Swedlow, J.R.; Herskowitz, I.; Vale, R.D. Actin-dependent localization of an RNA encoding a cell-fate determinant in yeast. Nature 1997, 389, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Pruyne, D.; Gao, L.; Bi, E.; Bretscher, A. Stable and dynamic axes of polarity use distinct formin isoforms in budding yeast. Mol. Biol. Cell 2004, 15, 4971–4989. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Liu, W.; Bretscher, A. The yeast formin Bnr1p has two localization regions that show spatially and temporally distinct association with septin structures. Mol. Biol. Cell 2010, 21, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Karpova, T.S.; McNally, J.G.; Moltz, S.L.; Cooper, J.A. Assembly and function of the actin cytoskeleton of yeast: Relationships between cables and patches. J. Cell Biol. 1998, 142, 1501–1517. [Google Scholar] [CrossRef]
- Mulholland, J.; Preuss, D.; Moon, A.; Wong, A.; Drubin, D.; Botstein, D. Ultrastructure of the yeast actin cytoskeleton and its association with the plasma membrane. J. Cell Biol. 1994, 125, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Kaksonen, M.; Sun, Y.; Drubin, D.G. A pathway for association of receptors, adaptors, and actin during endocytic internalization. Cell 2003, 115, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Kaksonen, M.; Toret, C.P.; Drubin, D.G. A modular design for the clathrin-and actin-mediated endocytosis machinery. Cell 2005, 123, 305–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpova, T.S.; Reck-Peterson, S.L.; Elkind, N.B.; Mooseker, M.S.; Novick, P.J.; Cooper, J.A. Role of actin and Myo2p in polarized secretion and growth of Saccharomyces cerevisiae. Mol. Biol. Cell 2000, 11, 1727–1737. [Google Scholar] [CrossRef] [PubMed]
- Rodal, A.A.; Kozubowski, L.; Goode, B.L.; Drubin, D.G.; Hartwig, J.H. Actin and septin ultrastructures at the budding yeast cell cortex. Mol. Biol. Cell 2005, 16, 372–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.E.; Cooper, J.A.; Bridgman, P.C. Yeast actin patches are networks of branched actin filaments. J. Cell Biol. 2004, 166, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Goley, E.D.; Welch, M.D. The ARP2/3 complex: An actin nucleator comes of age. Nat. Rev. Mol. Cell Biol. 2006, 7, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Evangelista, M.; Blundell, K.; Longtine, M.S.; Chow, C.J.; Adames, N.; Pringle, J.R.; Peter, M.; Boone, C. Bni1p, a yeast formin linking cdc42p and the actin cytoskeleton during polarized morphogenesis. Science 1997, 276, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Imamura, H.; Tanaka, K.; Hihara, T.; Umikawa, M.; Kamei, T.; Takahashi, K.; Sasaki, T.; Takai, Y. Bni1p and Bnr1p: Downstream targets of the Rho family small G-proteins which interact with profilin and regulate actin cytoskeleton in Saccharomyces cerevisiae. EMBO J. 1997, 16, 2745–2755. [Google Scholar] [CrossRef] [Green Version]
- Graziano, B.R.; DuPage, A.G.; Michelot, A.; Breitsprecher, D.; Moseley, J.B.; Sagot, I.; Blanchoin, L.; Goode, B.L. Mechanism and cellular function of Bud6 as an actin nucleation–promoting factor. Mol. Biol. Cell 2011, 22, 4016–4028. [Google Scholar] [CrossRef]
- Park, E.; Graziano, B.R.; Zheng, W.; Garabedian, M.; Goode, B.L.; Eck, M.J. Structure of a Bud6/Actin complex reveals a novel WH2-like actin monomer recruitment motif. Structure 2015, 23, 1492–1499. [Google Scholar] [CrossRef] [Green Version]
- Pruyne, D.; Evangelista, M.; Yang, C.; Bi, E.; Zigmond, S.; Bretscher, A.; Boone, C. Role of formins in actin assembly: Nucleation and barbed-end association. Science 2002, 297, 612–615. [Google Scholar] [CrossRef]
- Evangelista, M.; Pruyne, D.; Amberg, D.C.; Boone, C.; Bretscher, A. Formins direct Arp2/3-independent actin filament assembly to polarize cell growth in yeast. Nat. Cell Biol. 2002, 4, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Buttery, S.M.; Yoshida, S.; Pellman, D. Yeast formins Bni1 and Bnr1 utilize different modes of cortical interaction during the assembly of actin cables. Mol. Biol. Cell 2007, 18, 1826–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Linardopoulou, E.V.; Osborn, G.E.; Parkhurst, S.M. Formins in development: Orchestrating body plan origami. BBA Mol. Cell Res. 2010, 1803, 207–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drgonova, J.; Drgon, T.; Tanaka, K.; Kollár, R.; Chen, G.-C.; Ford, R.A.; Chan, C.S.; Takai, Y.; Cabib, E. Rho1p, a yeast protein at the interface between cell polarization and morphogenesis. Science 1996, 272, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Drgonová, J.; Drgon, T.; Roh, D.-H.; Cabib, E. The GTP-binding protein Rho1p is required for cell cycle progression and polarization of the yeast cell. J. Cell Biol. 1999, 146, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Roemer, T.; Vallier, L.; Sheu, Y.-J.; Snyder, M. The Spa2-related protein, Sph1p, is important for polarized growth in yeast. J. Cell Sci. 1998, 111, 479–494. [Google Scholar]
- Tcheperegine, S.E.; Gao, X.-D.; Bi, E. Regulation of cell polarity by interactions of Msb3 and Msb4 with Cdc42 and polarisome components. Mol. Cell. Biol. 2005, 25, 8567–8580. [Google Scholar] [CrossRef] [Green Version]
- Sheu, Y.-J.; Santos, B.; Fortin, N.; Costigan, C.; Snyder, M. Spa2p interacts with cell polarity proteins and signaling components involved in yeast cell morphogenesis. Mol. Cell. Biol. 1998, 18, 4053–4069. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Pruyne, D.; Bretscher, A. Formin-dependent actin assembly is regulated by distinct modes of Rho signaling in yeast. J. Cell Biol. 2003, 161, 1081–1092. [Google Scholar] [CrossRef] [Green Version]
- Ozaki-Kuroda, K.; Yamamoto, Y.; Nohara, H.; Kinoshita, M.; Fujiwara, T.; Irie, K.; Takai, Y. Dynamic localization and function of Bni1p at the sites of directed growth in Saccharomyces cerevisiae. Mol. Cell. Biol. 2001, 21, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Kuo, C.-C.; Kang, H.; Howell, A.S.; Zyla, T.R.; Jin, M.; Lew, D.J. Cdc42p regulation of the yeast formin Bni1p mediated by the effector Gic2p. Mol. Biol. Cell 2012, 23, 3814–3826. [Google Scholar] [CrossRef] [PubMed]
- Kamei, T.; Tanaka, K.; Hihara, T.; Umikawa, M.; Imamura, H.; Kikyo, M.; Ozaki, K.; Takai, Y. Interaction of Bnr1p with a novel Src Homology 3 domain-containing Hof1p implication in cytokinesis in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 28341–28345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikyo, M.; Tanaka, K.; Kamei, T.; Ozaki, K.; Fujiwara, T.; Inoue, E.; Takita, Y.; Ohya, Y.; Takai, Y. An FH domain-containing Bnr1p is a multifunctional protein interacting with a variety of cytoskeletal proteins in Saccharomyces cerevisiae. Oncogene 1999, 18, 7046–7054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.B.; Haarer, B.K.; Pringle, J.R. Cellular morphogenesis in the Saccharomyces cerevisiae cell cycle: Localization of the CDC3 gene product and the timing of events at the budding site. J. Cell Biol. 1991, 112, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Frazier, J.A.; Wong, M.L.; Longtine, M.S.; Pringle, J.R.; Mann, M.; Mitchison, T.J.; Field, C. Polymerization of purified yeast septins: Evidence that organized filament arrays may not be required for septin function. J. Cell Biol. 1998, 143, 737–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wloka, C.; Nishihama, R.; Onishi, M.; Oh, Y.; Hanna, J.; Pringle, J.R.; Krauß, M.; Bi, E. Evidence that a septin diffusion barrier is dispensable for cytokinesis in budding yeast. Biol. Chem. 2011, 392, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Kadota, J.; Yamamoto, T.; Yoshiuchi, S.; Bi, E.; Tanaka, K. Septin ring assembly requires concerted action of polarisome components, a PAK kinase Cla4p, and the actin cytoskeleton in Saccharomyces cerevisiae. Mol. Biol. Cell 2004, 15, 5329–5345. [Google Scholar] [CrossRef]
- Glomb, O.; Gronemeyer, T. Septin Organization and functions in budding yeast. Front. Cell Dev. Biol. 2016, 4, 123. [Google Scholar] [CrossRef] [Green Version]
- Bertin, A.; Nogales, E. Septin filament organization in Saccharomyces cerevisiae. Commun. Integr. Biol. 2012, 5, 503–505. [Google Scholar] [CrossRef]
- Iwase, M.; Luo, J.; Nagaraj, S.; Longtine, M.; Kim, H.B.; Haarer, B.K.; Caruso, C.; Tong, Z.; Pringle, J.R.; Bi, E. Role of a Cdc42p effector pathway in recruitment of the yeast septins to the presumptive bud site. Mol. Biol. Cell 2006, 17, 1110–1125. [Google Scholar] [CrossRef]
- Gladfelter, A.S.; Bose, I.; Zyla, T.R.; Bardes, E.S.; Lew, D.J. Septin ring assembly involves cycles of GTP loading and hydrolysis by Cdc42p. J. Cell Biol. 2002, 156, 315–326. [Google Scholar] [CrossRef]
- Takizawa, P.A.; DeRisi, J.L.; Wilhelm, J.E.; Vale, R.D. Plasma membrane compartmentalization in yeast by messenger RNA transport and a septin diffusion barrier. Science 2000, 290, 341–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barral, Y.; Mermall, V.; Mooseker, M.S.; Snyder, M. Compartmentalization of the cell cortex by septins is required for maintenance of cell polarity in yeast. Mol. Cell 2000, 5, 841–851. [Google Scholar] [CrossRef]
- Caviston, J.P.; Longtine, M.; Pringle, J.R.; Bi, E. The role of Cdc42p GTPase-activating proteins in assembly of the septin ring in yeast. Mol. Biol. Cell 2003, 14, 4051–4066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, T.; Botstein, D. Movement of yeast cortical actin cytoskeleton visualized in vivo. Proc. Natl Acad. Sci. USA 1996, 93, 3886–3891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waddle, J.A.; Karpova, T.S.; Waterston, R.H.; Cooper, J.A. Movement of cortical actin patches in yeast. J. Cell Biol. 1996, 132, 861–870. [Google Scholar] [CrossRef]
- Smith, M.G.; Swamy, S.R.; Pon, L.A. The life cycle of actin patches in mating yeast. J. Cell Sci. 2001, 114, 1505–1513. [Google Scholar]
- Lu, R.; Drubin, D.G.; Sun, Y. Clathrin-mediated endocytosis in budding yeast at a glance. J. Cell Sci. 2016, 129, 1531–1536. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Martin, A.C.; Drubin, D.G. Endocytic internalization in budding yeast requires coordinated actin nucleation and myosin motor activity. Dev. Cell 2006, 11, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Jonsdottir, G.A.; Li, R. Dynamics of yeast myosin I: Evidence for a possible role in scission of endocytic vesicles. Curr. Biol. 2004, 14, 1604–1609. [Google Scholar] [CrossRef] [Green Version]
- Smaczynska-de Rooij, I.I.; Allwood, E.G.; Mishra, R.; Booth, W.I.; Aghamohammadzadeh, S.; Goldberg, M.W.; Ayscough, K.R. Yeast dynamin Vps1 and amphiphysin Rvs167 function together during endocytosis. Traffic 2012, 13, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Vallen, E.A.; Caviston, J.; Bi, E. Roles of Hof1p, Bni1p, Bnr1p, and Myo1p in cytokinesis in Saccharomyces cerevisiae. Mol. Biol. Cell 2000, 11, 593–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wloka, C.; Vallen, E.A.; Fang, X.; Oh, Y.; Bi, E. Immobile myosin-II plays a scaffolding role during cytokinesis in budding yeast. J. Cell Biol. 2013, 200, 271–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cid, V.C.J.; Adamiková, L.; Sánchez, M.; Molina, M.A.; Nombela, C. Cell cycle control of septin ring dynamics in the budding yeast. Microbiology 2001, 147, 1437–1450. [Google Scholar] [CrossRef] [Green Version]
- Lippincott, J.; Shannon, K.B.; Shou, W.; Deshaies, R.J.; Li, R. The Tem1 small GTPase controls actomyosin and septin dynamics during cytokinesis. J. Cell Sci. 2001, 114, 1379–1386. [Google Scholar]
- Yoshida, S.; Kono, K.; Lowery, D.M.; Bartolini, S.; Yaffe, M.B.; Ohya, Y.; Pellman, D. Polo-like kinase Cdc5 controls the local activation of Rho1 to promote cytokinesis. Science 2006, 313, 108–111. [Google Scholar] [CrossRef]
- Atkins, B.D.; Yoshida, S.; Saito, K.; Wu, C.-F.; Lew, D.J.; Pellman, D. Inhibition of Cdc42 during mitotic exit is required for cytokinesis. J. Cell Biol. 2013, 202, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Meitinger, F.; Richter, H.; Heisel, S.; Hub, B.; Seufert, W.; Pereira, G. A safeguard mechanism regulates Rho GTPases to coordinate cytokinesis with the establishment of cell polarity. PLoS Biol. 2013, 11, e1001495. [Google Scholar] [CrossRef]
- Osman, M.A.; Cerione, R.A. Iqg1p, a yeast homologue of the mammalian IQGAPs, mediates cdc42p effects on the actin cytoskeleton. J. Cell Biol. 1998, 142, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Shannon, K.B.; Li, R. The multiple roles of Cyk1p in the assembly and function of the actomyosin ring in budding yeast. Mol. Biol. Cell 1999, 10, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Vallen, E.A.; Dravis, C.; Tcheperegine, S.E.; Drees, B.; Bi, E. Identification and functional analysis of the essential and regulatory light chains of the only type II myosin Myo1p in Saccharomyces cerevisiae. J. Cell Biol. 2004, 165, 843–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes Pinto, I.; Rubinstein, B.; Kucharavy, A.; Unruh, J.R.; Li, R. Actin depolymerization drives actomyosin ring contraction during budding yeast cytokinesis. Dev. Cell 2012, 22, 1247–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardin, A.J.; Amon, A. Men and sin: What’s the difference? Nat. Rev. Mol. Cell Biol. 2001, 2, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Luca, F.C.; Mody, M.; Kurischko, C.; Roof, D.M.; Giddings, T.H.; Winey, M. Saccharomyces cerevisiae Mob1p is required for cytokinesis and mitotic exit. Mol. Cell. Biol. 2001, 21, 6972–6983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menssen, R.; Neutzner, A.; Seufert, W. Asymmetric spindle pole localization of yeast Cdc15 kinase links mitotic exit and cytokinesis. Curr. Biol. 2001, 11, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Teh, E.M.; Chai, C.C.; Yeong, F.M. Retention of Chs2p in the ER requires N-terminal CDK1-phosphorylation sites. Cell Cycle 2009, 8, 2965–2976. [Google Scholar] [CrossRef] [Green Version]
- Chuang, J.S.; Schekman, R.W. Differential trafficking and timed localization of two chitin synthase proteins, Chs2p and Chs3p. J. Cell Biol. 1996, 135, 597–610. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Kashimshetty, R.; Ng, K.E.; Tan, H.B.; Yeong, F.M. Exit from mitosis triggers Chs2p transport from the endoplasmic reticulum to mother-daughter neck via the secretory pathway in budding yeast. J. Cell Biol. 2006, 174, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.F.; Bennett, A.M.; Ma, W.K.; Hall, M.C.; Yeong, F.M. Dependence of Chs2 ER export on dephosphorylation by cytoplasmic Cdc14 ensures that septum formation follows mitosis. Mol. Biol. Cell 2012, 23, 45–58. [Google Scholar] [CrossRef]
- Sburlati, A.; Cabib, E. Chitin synthetase 2, a presumptive participant in septum formation in Saccharomyces cerevisiae. J. Biol. Chem. 1986, 261, 15147–15152. [Google Scholar]
- Shaw, J.A.; Mol, P.C.; Bowers, B.; Silverman, S.J.; Valdivieso, M.H.; Durán, A.; Cabib, E. The function of chitin synthases 2 and 3 in the Saccharomyces cerevisiae cell cycle. J. Cell Biol. 1991, 114, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulawa, C.E. Genetics and molecular biology of chitin synthesis in fungi. Annu. Rev. Microbiol. 1993, 47, 505–534. [Google Scholar] [CrossRef] [PubMed]
- Orlean, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMay, B.S.; Bai, X.; Howard, L.; Occhipinti, P.; Meseroll, R.A.; Spiliotis, E.T.; Oldenbourg, R.; Gladfelter, A.S. Septin filaments exhibit a dynamic, paired organization that is conserved from yeast to mammals. J. Cell Biol. 2011, 193, 1065–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrabioiu, A.M.; Mitchison, T.J. Structural insights into yeast septin organization from polarized fluorescence microscopy. Nature 2006, 443, 466. [Google Scholar] [CrossRef] [PubMed]
- Onishi, M.; Ko, N.; Nishihama, R.; Pringle, J.R. Distinct roles of Rho1, Cdc42, and Cyk3 in septum formation and abscission during yeast cytokinesis. J. Cell Biol. 2013, 202, 311–329. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, M.K.; Cheng, Z.; Lam, S.K.; Roth-Johnson, E.; Barfield, R.M.; Schekman, R. Phosphorylation of Chs2p regulates interaction with COPII. J. Cell Sci. 2013, 126, 2151–2156. [Google Scholar] [CrossRef] [Green Version]
- Meitinger, F.; Petrova, B.; Lombardi, I.M.; Bertazzi, D.T.; Hub, B.; Zentgraf, H.; Pereira, G. Targeted localization of Inn1, Cyk3 and Chs2 by the mitotic-exit network regulates cytokinesis in budding yeast. J. Cell Sci. 2010, 123, 1851–1861. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Chang, K.-J.; Orlean, P.; Wloka, C.; Deshaies, R.; Bi, E. Mitotic exit kinase Dbf2 directly phosphorylates chitin synthase Chs2 to regulate cytokinesis in budding yeast. Mol. Biol. Cell 2012, 23, 2445–2456. [Google Scholar] [CrossRef]
- Cabib, E.; Schmidt, M. Chitin synthase III activity, but not the chitin ring, is required for remedial septa formation in budding yeast. FEMS Microbiol. Lett. 2003, 224, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Izumikawa, T.; Kanagawa, N.; Watamoto, Y.; Okada, M.; Saeki, M.; Sakano, M.; Sugahara, K.; Sugihara, K.; Asano, M.; Kitagawa, H. Impairment of embryonic cell division and glycosaminoglycan biosynthesis in glucuronyltransferase-I-deficient mice. J. Biol. Chem. 2010, 285, 12190–12196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Vogel, B.E. A secreted protein promotes cleavage furrow maturation during cytokinesis. Curr. Biol. 2011, 21, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, C.M.; Foor, F.; Marrinan, J.A.; Morin, N.; Nielsen, J.B.; Dahl, A.M.; Mazur, P.; Baginsky, W.; Li, W.; El-Sherbeini, M. The Saccharomyces cerevisiae FKS1 (ETG1) gene encodes an integral membrane protein which is a subunit of 1, 3-beta-D-glucan synthase. Proc. Natl. Acad. Sci. USA 1994, 91, 12907–12911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, S.B.; Takewaki, N.; Takasuka, T.; Mio, T.; Adachi, M.; Fujii, Y.; Miyamoto, C.; Arisawa, M.; Furuichi, Y.; Watanabe, T. Characterization and gene cloning of 1, 3-β-D-glucan synthase from Saccharomyces cerevisiae. FEBS J. 1995, 231, 845–854. [Google Scholar] [CrossRef]
- Yoshida, S.; Bartolini, S.; Pellman, D. Mechanisms for concentrating Rho1 during cytokinesis. Genes Dev. 2009, 23, 810–823. [Google Scholar] [CrossRef] [Green Version]
- Mazur, P.; Baginsky, W. In vitro activity of 1, 3-β-D-glucan synthase requires the GTP-binding protein Rho1. J. Biol. Chem. 1996, 271, 14604–14609. [Google Scholar] [CrossRef] [Green Version]
- Qadota, H.; Python, C.P.; Inoue, S.B.; Arisawa, M.; Anraku, Y.; Zheng, Y.; Watanabe, T.; Levin, D.E.; Ohya, Y. Identification of yeast Rho1p GTPase as a regulatory subunit of 1, 3-β-glucan synthase. Science 1996, 272, 279–281. [Google Scholar] [CrossRef]
- Sekiya-Kawasaki, M.; Abe, M.; Saka, A.; Watanabe, D.; Kono, K.; Minemura-Asakawa, M.; Ishihara, S.; Watanabe, T.; Ohya, Y. Dissection of upstream regulatory components of the Rho1p effector, 1, 3-β-glucan synthase, in Saccharomyces cerevisiae. Genetics 2002, 162, 663–676. [Google Scholar]
- Baladrón, V.; Ufano, S.; Dueñas, E.; Martín-Cuadrado, A.B.; del Rey, F.; de Aldana, C.R.V. Eng1p, an endo-1, 3-β-glucanase localized at the daughter side of the septum, is involved in cell separation in Saccharomyces cerevisiae. Eukaryot. Cell 2002, 1, 774–786. [Google Scholar] [CrossRef] [Green Version]
- Kuranda, M.J.; Robbins, P.W. Chitinase is required for cell separation during growth of Saccharomyces cerevisiae. J. Biol. Chem. 1991, 266, 19758–19767. [Google Scholar]
- Colman-Lerner, A.; Chin, T.E.; Brent, R. Yeast Cbk1 and Mob2 activate daughter-specific genetic programs to induce asymmetric cell fates. Cell 2001, 107, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Fankhauser, C.; Reymond, A.; Cerutti, L.; Utzig, S.; Hofmann, K.; Simanis, V. The S. pombe cdc15 gene is a key element in the reorganization of F-actin at mitosis. Cell 1995, 82, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Lippincott, J.; Li, R. Dual function of Cyk2, a cdc15/PSTPIP family protein, in regulating actomyosin ring dynamics and septin distribution. J. Cell Biol. 1998, 143, 1947–1960. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.; Dowbenko, D.; Cheng, J.; Li, W.; Brush, J.; Utzig, S.; Simanis, V.; Lasky, L.A. PSTPIP: A tyrosine phosphorylated cleavage furrow–associated protein that is a substrate for a PEST tyrosine phosphatase. J. Cell Biol. 1997, 138, 845–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, M.K.; McCollum, D.; Chang, L.; Wong, K.C.; Naqvi, N.I.; He, X.; Sazer, S.; Gould, K.L. Isolation and characterization of new fission yeast cytokinesis mutants. Genetics 1998, 149, 1265–1275. [Google Scholar] [PubMed]
- Rechsteiner, M. PEST sequences are signals for rapid intracellular proteolysis. Semin. Cell Biol. 1990, 1, 433–440. [Google Scholar] [PubMed]
- Itoh, T.; Erdmann, K.S.; Roux, A.; Habermann, B.; Werner, H.; De Camilli, P. Dynamin and the actin cytoskeleton cooperatively regulate plasma membrane invagination by BAR and F-BAR proteins. Dev. Cell 2005, 9, 791–804. [Google Scholar] [CrossRef]
- Tsujita, K.; Suetsugu, S.; Sasaki, N.; Furutani, M.; Oikawa, T.; Takenawa, T. Coordination between the actin cytoskeleton and membrane deformation by a novel membrane tubulation domain of PCH proteins is involved in endocytosis. J. Cell Biol. 2006, 172, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Aspenström, P. Roles of F-BAR/PCH proteins in the regulation of membrane dynamics and actin reorganization. Int. Rev. Cell Mol. Biol. 2008, 272, 1–31. [Google Scholar]
- Itoh, T.; De Camilli, P. BAR, F-BAR (EFC) and ENTH/ANTH domains in the regulation of membrane–cytosol interfaces and membrane curvature. BBA Mol. Cell Biol. Lipids 2006, 1761, 897–912. [Google Scholar] [CrossRef]
- Takano, K.; Toyooka, K.; Suetsugu, S. EFC/F-BAR proteins and the N-WASP–WIP complex induce membrane curvature-dependent actin polymerization. EMBO J. 2008, 27, 2817–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, C.A.; Gillum, J.D.; Seidman, C.E.; Lindor, N.M.; Veile, R.; Bashiardes, S.; Lovett, M. Mutations in CD2BP1 disrupt binding to PTP PEST and are responsible for PAPA syndrome, an autoinflammatory disorder. Hum. Mol. Genet. 2002, 11, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Mayer, B.J.; Hanafusa, H. Mutagenic analysis of the v-crk oncogene: Requirement for SH2 and SH3 domains and correlation between increased cellular phosphotyrosine and transformation. J. Virol. 1990, 64, 3581–3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feller, S.M.; Ren, R.; Hanafusa, H.; Baltimore, D. SH2 and SH3 domains as molecular adhesives: The interactions of Crk and Abl. Trends Biochem. Sci. 1994, 19, 453–458. [Google Scholar] [CrossRef]
- Mayer, B.J.; Eck, M.J. SH3 domains: Minding your p’s and q’s. Curr. Biol. 1995, 5, 364–367. [Google Scholar] [CrossRef] [Green Version]
- Bar-Sagi, D.; Rotin, D.; Batzer, A.; Mandiyan, V.; Schlessinger, J. SH3 domains direct cellular localization of signaling molecules. Cell 1993, 74, 83–91. [Google Scholar] [CrossRef]
- Koch, C.A.; Anderson, D.; Moran, M.F.; Ellis, C.; Pawson, T. SH2 and SH3 domains: Elements that control interactions of cytoplasmic signaling proteins. Science 1991, 252, 668–675. [Google Scholar] [CrossRef]
- Blondel, M.; Bach, S.; Bamps, S.; Dobbelaere, J.; Wiget, P.; Longaretti, C.; Barral, Y.; Meijer, L.; Peter, M. Degradation of Hof1 by SCFGrr1 is important for actomyosin contraction during cytokinesis in yeast. EMBO J. 2005, 24, 1440–1452. [Google Scholar] [CrossRef] [Green Version]
- Graziano, B.R.; Hoi-Ying, E.Y.; Alioto, S.L.; Eskin, J.A.; Ydenberg, C.A.; Waterman, D.P.; Garabedian, M.; Goode, B.L. The F-BAR protein Hof1 tunes formin activity to sculpt actin cables during polarized growth. Mol. Biol. Cell 2014, 25, 1730–1743. [Google Scholar] [CrossRef]
- Meitinger, F.; Boehm, M.E.; Hofmann, A.; Hub, B.; Zentgraf, H.; Lehmann, W.D.; Pereira, G. Phosphorylation-dependent regulation of the F-BAR protein Hof1 during cytokinesis. Genes Dev. 2011, 25, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Meitinger, F.; Palani, S.; Hub, B.; Pereira, G. Dual function of the NDR-kinase Dbf2 in the regulation of the F-BAR protein Hof1 during cytokinesis. Mol. Biol. Cell 2013, 24, 1290–1304. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Wang, J.; Brinkworth, R.; Winsor, B.; Kobe, B.; Munn, A.L. Verprolin cytokinesis function mediated by the hof one trap domain. Traffic 2005, 6, 575–593. [Google Scholar] [CrossRef] [PubMed]
- Labedzka, K.; Tian, C.; Nussbaumer, U.; Timmermann, S.; Walther, P.; Müller, J.; Johnsson, N. Sho1p connects the plasma membrane with proteins of the cytokinesis network through multiple isomeric interaction states. J. Cell Sci. 2012, 125, 4103–4113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishihama, R.; Schreiter, J.H.; Onishi, M.; Vallen, E.A.; Hanna, J.; Moravcevic, K.; Lippincott, M.F.; Han, H.; Lemmon, M.A.; Pringle, J.R. Role of Inn1 and its interactions with Hof1 and Cyk3 in promoting cleavage furrow and septum formation in S. cerevisiae. J. Cell Biol. 2009, 185, 995–1012. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Nishizawa, K.; An, W.; Hussey, R.E.; Lialios, F.E.; Salgia, R.; Sunder-Plassmann, R.; Reinherz, E.L. A cdc15-like adaptor protein (CD2BP1) interacts with the CD2 cytoplasmic domain and regulates CD2-triggered adhesion. EMBO J. 1998, 17, 7320–7336. [Google Scholar] [CrossRef] [Green Version]
- Angers-Loustau, A.; Côté, J.-F.; Charest, A.; Dowbenko, D.; Spencer, S.; Lasky, L.A.; Tremblay, M.L. Protein tyrosine phosphatase-PEST regulates focal adhesion disassembly, migration, and cytokinesis in fibroblasts. J. Cell Biol. 1999, 144, 1019–1031. [Google Scholar] [CrossRef]
- Côté, J.-F.; Chung, P.L.; Théberge, J.-F.; Hallé, M.; Spencer, S.; Lasky, L.A.; Tremblay, M.L. PSTPIP is a substrate of PTP-PEST and serves as a scaffold guiding PTP-PEST toward a specific dephosphorylation of WASP. J. Biol. Chem. 2002, 277, 2973–2986. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Spencer, S.D.; Lasky, L.A. Tyrosine phosphorylation regulates the SH3-mediated binding of the Wiskott-Aldrich syndrome protein to PSTPIP, a cytoskeletal-associated protein. J. Biol. Chem. 1998, 273, 5765–5770. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, N.; Antón, I.M.; Hartwig, J.H.; Geha, R.S. WIP, a protein associated with Wiskott–Aldrich syndrome protein, induces actin polymerization and redistribution in lymphoid cells. Proc. Natl. Acad. Sci. USA 1997, 94, 14671–14676. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.M.; Tian, L.; Nelson, D.L. Mutations that cause the Wiskott-Aldrich syndrome impair the interaction of Wiskott-Aldrich syndrome protein (WASP) with WASP interacting protein. J. Immunol. 1999, 162, 5019–5024. [Google Scholar]
- Machesky, L.M.; Atkinson, S.J.; Ampe, C.; Vandekerckhove, J.; Pollard, T.D. Purification of a cortical complex containing two unconventional actins from Acanthamoeba by affinity chromatography on profilin-agarose. J. Cell Biol. 1994, 127, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Machesky, L.M.; Insall, R.H. Scar1 and the related Wiskott–Aldrich syndrome protein, WASP, regulate the actin cytoskeleton through the Arp2/3 complex. Curr. Biol. 1998, 8, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Pollard, T.D.; Borisy, G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, R.; Ma, L.; Miki, H.; Lopez, M.; Kirchhausen, T.; Takenawa, T.; Kirschner, M.W. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 1999, 97, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Miki, H.; Miura, K.; Takenawa, T. N-WASP, a novel actin-depolymerizing protein, regulates the cortical cytoskeletal rearrangement in a PIP2-dependent manner downstream of tyrosine kinases. EMBO J. 1996, 15, 5326. [Google Scholar] [CrossRef]
- Suetsugu, S.; Miki, H.; Yamaguchi, H.; Obinata, T.; Takenawa, T. Enhancement of branching efficiency by the actin filament-binding activity of N-WASP/WAVE2. J. Cell Sci. 2001, 114, 4533–4542. [Google Scholar]
- Symons, M.; Derry, J.M.; Karlak, B.; Jiang, S.; Lemahieu, V.; McCormick, F.; Francke, U.; Abo, A. Wiskott–Aldrich syndrome protein, a novel effector for the GTPase CDC42Hs, is implicated in actin polymerization. Cell 1996, 84, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Badour, K.; Zhang, J.; Shi, F.; Leng, Y.; Collins, M.; Siminovitch, K.A. Fyn and PTP-PEST–mediated regulation of Wiskott-Aldrich syndrome protein (WASp) tyrosine phosphorylation is required for coupling T cell antigen receptor engagement to WASp effector function and T cell activation. J. Exp. Med. 2004, 199, 99–112. [Google Scholar] [CrossRef]
- Badour, K.; Zhang, J.; Shi, F.; McGavin, M.K.; Rampersad, V.; Hardy, L.A.; Field, D.; Siminovitch, K.A. The Wiskott-Aldrich syndrome protein acts downstream of CD2 and the CD2AP and PSTPIP1 adaptors to promote formation of the immunological synapse. Immunity 2003, 18, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Gallego, M.D.; Miguel, A.; Anton, I.M.; Snapper, S.; Fuhlbrigge, R.; Geha, R.S. WIP and WASP play complementary roles in T cell homing and chemotaxis to SDF-1α. Int. Immunol. 2006, 18, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; George, B.; Thanabalu, T. Wiskott–Aldrich Syndrome causing mutation, Pro373Ser restricts conformational changes essential for WASP activity in T-cells. BBA Mol. Basis Dis. 2014, 1842, 623–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenney, D.; Cairns, L.; Remold-O’Donnell, E.; Peterson, J.; Rosen, F.S.; Parkman, R. Morphological abnormalities in the lymphocytes of patients with the Wiskott-Aldrich syndrome. Blood 1986, 68, 1329–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snapper, S.B.; Rosen, F.S. The Wiskott-Aldrich syndrome protein (WASP): Roles in signaling and cytoskeletal organization. Annu. Rev. Immunol. 1999, 17, 905–929. [Google Scholar] [CrossRef]
- Wada, T.; Jagadeesh, G.J.; Nelson, D.L.; Candotti, F. Retrovirus-mediated WASP gene transfer corrects Wiskott-Aldrich syndrome T-cell dysfunction. Hum. Gene Ther. 2002, 13, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.N.; Zahn, R.; Mitchell, D.A.; Stevenson, B.J.; Munn, A.L. The WASp homologue Las17p functions with the WIP homologue End5p/verprolin and is essential for endocytosis in yeast. Curr. Biol. 1998, 8, 959–962. [Google Scholar] [CrossRef] [Green Version]
- Thanabalu, T.; Munn, A.L. Functions of Vrp1p in cytokinesis and actin patches are distinct and neither requires a WH2/V domain. EMBO J. 2001, 20, 6979–6989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naqvi, S.N.; Feng, Q.; Boulton, V.J.; Zahn, R.; Munn, A.L. Vrp1p functions in both actomyosin ring-dependent and Hof1p-dependent pathways of cytokinesis. Traffic 2001, 2, 189–201. [Google Scholar] [CrossRef]
- Vaduva, G.; Martinez-Quiles, N.; Anton, I.M.; Martin, N.C.; Geha, R.S.; Hopper, A.K.; Ramesh, N. The human WASP-interacting protein, WIP, activates the cell polarity pathway in yeast. J. Biol. Chem. 1999, 274, 17103–17108. [Google Scholar] [CrossRef] [Green Version]
- Rajmohan, R.; Meng, L.; Yu, S.; Thanabalu, T. WASP suppresses the growth defect of Saccharomyces cerevisiae las17Δ strain in the presence of WIP. Biochem. Biophys. Res. Commun. 2006, 342, 529–536. [Google Scholar] [CrossRef]
- Endris, V.; Wogatzky, B.; Leimer, U.; Bartsch, D.; Zatyka, M.; Latif, F.; Maher, E.R.; Tariverdian, G.; Kirsch, S.; Karch, D. The novel Rho-GTPase activating gene MEGAP/srGAP3 has a putative role in severe mental retardation. Proc. Natl. Acad. Sci. USA 2002, 99, 11754–11759. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Coulombe, P.A. Emerging role for the cytoskeleton as an organizer and regulator of translation. Nat. Rev. Mol. Cell Biol. 2010, 11, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, A.G. Translational regulation of GCN4 and the general amino acid control of yeast. Annu. Rev. Microbiol. 2005, 59, 407–450. [Google Scholar] [CrossRef] [PubMed]
- Castilho, B.A.; Shanmugam, R.; Silva, R.C.; Ramesh, R.; Himme, B.M.; Sattlegger, E. Keeping the eIF2 alpha kinase Gcn2 in check. BBA Mol. Cell Res. 2014, 1843, 1948–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, R.C.; Sattlegger, E.; Castilho, B.A. Perturbations in actin dynamics reconfigure protein complexes that modulate GCN2 activity and promote an eIF2 response. J. Cell Sci. 2016, 129, 4521–4533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattlegger, E.; Swanson, M.J.; Ashcraft, E.A.; Jennings, J.L.; Fekete, R.A.; Link, A.J.; Hinnebusch, A.G. YIH1 is an actin-binding protein that inhibits protein kinase GCN2 and impairs general amino acid control when overexpressed. J. Biol. Chem. 2004, 279, 29952–29962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visweswaraiah, J.; Lageix, S.; Castilho, B.A.; Izotova, L.; Kinzy, T.G.; Hinnebusch, A.G.; Sattlegger, E. Evidence that eukaryotic translation elongation factor 1A (eEF1A) binds the Gcn2 protein C terminus and inhibits Gcn2 activity. J. Biol. Chem. 2011, 286, 36568–36579. [Google Scholar] [CrossRef] [Green Version]
- Kubota, H.; Sakaki, Y.; Ito, T. GI domain-mediated association of the eukaryotic initiation factor 2α kinase GCN2 with its activator GCN1 is required for general amino acid control in budding yeast. J. Biol. Chem. 2000, 275, 20243–20246. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.M.; Sattlegger, E.; Jiang, H.-Y.; Longo, B.M.; Jaqueta, C.B.; Hinnebusch, A.G.; Wek, R.C.; Mello, L.E.; Castilho, B.A. IMPACT, a protein preferentially expressed in the mouse brain, binds GCN1 and inhibits GCN2 activation. J. Biol. Chem. 2005, 280, 28316–28323. [Google Scholar] [CrossRef] [Green Version]
- Mateyak, M.K.; Kinzy, T.G. eEF1A: Thinking outside the ribosome. J. Biol. Chem. 2010, 285, 21209–21213. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Qiu, H.; Garcia-Barrio, M.; Anderson, J.; Hinnebusch, A.G. Uncharged tRNA activates GCN2 by displacing the protein kinase moiety from a bipartite tRNA-binding domain. Mol. Cell 2000, 6, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Arenz, S.; Abdelshahid, M.; Sohmen, D.; Payoe, R.; Starosta, A.L.; Berninghausen, O.; Hauryliuk, V.; Beckmann, R.; Wilson, D.N. The stringent factor RelA adopts an open conformation on the ribosome to stimulate ppGpp synthesis. Nucleic Acids Res. 2016, 44, 6471–6481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.; Fernández, I.S.; Gordiyenko, Y.; Ramakrishnan, V. Ribosome-dependent activation of stringent control. Nature 2016, 534, 277–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattlegger, E.; Hinnebusch, A.G. Separate domains in GCN1 for binding protein kinase GCN2 and ribosomes are required for GCN2 activation in amino acid-starved cells. EMBO J. 2000, 19, 6622–6633. [Google Scholar] [CrossRef] [PubMed]
- Marton, M.J.; De Aldana, C.V.; Qiu, H.; Chakraburtty, K.; Hinnebusch, A.G. Evidence that GCN1 and GCN20, translational regulators of GCN4, function on elongating ribosomes in activation of eIF2alpha kinase GCN2. Mol. Cell. Biol. 1997, 17, 4474–4489. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.; Wek, R.C.; Hinnebusch, A.G. Ribosome association of GCN2 protein kinase, a translational activator of the GCN4 gene of Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 3027–3036. [Google Scholar] [CrossRef] [Green Version]
- Dey, M.; Cao, C.; Sicheri, F.; Dever, T.E. Conserved intermolecular salt bridge required for activation of protein kinases PKR, GCN2, and PERK. J. Biol. Chem. 2007, 282, 6653–6660. [Google Scholar] [CrossRef] [Green Version]
- Gárriz, A.; Qiu, H.; Dey, M.; Seo, E.-J.; Dever, T.E.; Hinnebusch, A.G. A network of hydrophobic residues impeding helix αC rotation maintains latency of kinase Gcn2, which phosphorylates the α subunit of translation initiation factor 2. Mol. Cell. Biol. 2009, 29, 1592–1607. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, A.G. eIF2 [alpha] kinases provide a new solution to the puzzle of substrate specificity. Nat. Struct. Mol. Biol. 2005, 12, 835–839. [Google Scholar] [CrossRef]
- Lageix, S.; Rothenburg, S.; Dever, T.E.; Hinnebusch, A.G. Enhanced interaction between pseudokinase and kinase domains in Gcn2 stimulates eIF2α phosphorylation in starved cells. PLoS Genet. 2014, 10, e1004326. [Google Scholar] [CrossRef] [Green Version]
- Lageix, S.; Zhang, J.; Rothenburg, S.; Hinnebusch, A.G. Interaction between the tRNA-binding and C-terminal domains of yeast Gcn2 regulates kinase activity in vivo. PLoS Genet. 2015, 11, e1004991. [Google Scholar] [CrossRef] [Green Version]
- Padyana, A.K.; Qiu, H.; Roll-Mecak, A.; Hinnebusch, A.G.; Burley, S.K. Structural basis for autoinhibition and mutational activation of eukaryotic initiation factor 2α protein kinase GCN2. J. Biol. Chem. 2005, 280, 29289–29299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, H.P.; Ordonez, A.; Allen, F.; Parts, L.; Inglis, A.J.; Williams, R.L.; Ron, D. The ribosomal P-stalk couples amino acid starvation to GCN2 activation in mammalian cells. Elife 2019, 8, e50149. [Google Scholar] [CrossRef] [PubMed]
- Inglis, A.J.; Masson, G.R.; Shao, S.; Perisic, O.; McLaughlin, S.H.; Hegde, R.S.; Williams, R.L. Activation of GCN2 by the ribosomal P-stalk. Proc. Natl. Acad. Sci. USA 2019, 116, 4946–4954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Diaz, A.; Remacha, M.; Ballesta, J.P.; Berlanga, J.J. Phosphorylation of initiation factor eIF2 in response to stress conditions is mediated by acidic ribosomal P1/P2 proteins in Saccharomyces cerevisiae. PLoS ONE 2013, 8, e84219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dever, T.E.; Kinzy, T.G.; Pavitt, G.D. Mechanism and regulation of protein synthesis in Saccharomyces cerevisiae. Genetics 2016, 203, 65–107. [Google Scholar] [CrossRef] [Green Version]
- Kilberg, M.S.; Shan, J.; Su, N. ATF4-dependent transcription mediates signaling of amino acid limitation. Trends Endocrinol. Metab. 2009, 20, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Rakesh, R.; Krishnan, R.; Sattlegger, E.; Srinivasan, N. Recognition of a structural domain (RWDBD) in Gcn1 proteins that interacts with the RWD domain containing proteins. Biol. Direct 2017, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Cambiaghi, T.D.; Pereira, C.M.; Shanmugam, R.; Bolech, M.; Wek, R.C.; Sattlegger, E.; Castilho, B.A. Evolutionarily conserved IMPACT impairs various stress responses that require GCN1 for activating the eIF2 kinase GCN2. Biochem. Biophys. Res. Commun. 2014, 443, 592–597. [Google Scholar] [CrossRef]
- Baird, T.D.; Wek, R.C. Eukaryotic initiation factor 2 phosphorylation and translational control in metabolism. Adv. Nutr. 2012, 3, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The many roles of the eukaryotic elongation factor 1 complex. Wiley Interdiscip. Rev. RNA 2012, 3, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Edmonds, B.T.; Bell, A.; Wyckoff, J.; Condeelis, J.; Leyh, T.S. The effect of F-actin on the binding and hydrolysis of guanine nucleotide by Dictyostelium elongation factor 1A. J. Biol. Chem. 1998, 273, 10288–10295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edmonds, B.T.; Murray, J.; Condeelis, J. pH regulation of the F-actin binding properties of Dictyostelium elongation factor 1α. J. Biol. Chem. 1995, 270, 15222–15230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Tang, J.; Edmonds, B.T.; Murray, J.; Levin, S.; Condeelis, J. F-actin sequesters elongation factor 1alpha from interaction with aminoacyl-tRNA in a pH-dependent reaction. J. Cell Biol. 1996, 135, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Astekar, M.; Soi, S.; Manjunatha, B.S.; Shetty, D.C.; Radhakrishnan, R. pH gradient reversal: An emerging hallmark of cancers. Recent Pat. Anticancer. Drug Discov. 2015, 10, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Namkung, W.; Yoon, J.S.; Jo, M.J.; Lee, S.H.; Kim, K.H.; Kim, J.Y.; Lee, M.G. The role of translation elongation factor eEF1A in intracellular alkalinization-induced tumor cell growth. Lab. Investig. 2009, 89, 867–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, Y.; Hirai, M.; Nishiyama, K.; Kanazawa, I.; Ueda, T.; Sakaki, Y.; Ito, T. Screening for imprinted genes by allelic message display: Identification of a paternally expressed gene impact on mouse chromosome 18. Proc. Natl. Acad. Sci. USA 1997, 94, 9249–9254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattlegger, E.; Barbosa, J.A.; Moraes, M.C.S.; Martins, R.M.; Hinnebusch, A.G.; Castilho, B.A. Gcn1 and actin binding to Yih1 implications for activation of the eIF2 kinase Gcn2. J. Biol. Chem. 2011, 286, 10341–10355. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.C.; Dautel, M.; Di Genova, B.M.; Amberg, D.C.; Castilho, B.A.; Sattlegger, E. The Gcn2 regulator Yih1 interacts with the cyclin dependent kinase Cdc28 and promotes cell cycle progression through G2/M in budding yeast. PLoS ONE 2015, 10, e0131070. [Google Scholar] [CrossRef] [Green Version]
- Waller, T.; Lee, S.J.; Sattlegger, E. Evidence that Yih1 resides in a complex with ribosomes. FEBS J. 2012, 279, 1761–1776. [Google Scholar] [CrossRef]
- Roffé, M.; Hajj, G.N.; Azevedo, H.F.; Alves, V.S.; Castilho, B.A. IMPACT is a developmentally regulated protein in neurons that opposes the eukaryotic initiation factor 2α kinase GCN2 in the modulation of neurite outgrowth. J. Biol. Chem. 2013, 288, 10860–10869. [Google Scholar] [CrossRef] [Green Version]
- Bittencourt, S.; Pereira, C.M.; Avedissian, M.; Delamano, A.; Mello, L.E.; Castilho, B.A. Distribution of the protein IMPACT, an inhibitor of GCN2, in the mouse, rat, and marmoset brain. J. Comp. Neurol. 2008, 507, 1811–1830. [Google Scholar] [CrossRef] [PubMed]
- Zharikov, S.I.; Sigova, A.A.; Chen, S.; Bubb, M.R.; Block, E.R. Cytoskeletal regulation of the L-arginine/NO pathway in pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2001, 280, L465–L473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminska, M.; Havrylenko, S.; Decottignies, P.; Le Marechal, P.; Negrutskii, B.; Mirande, M. Dynamic organization of aminoacyl-tRNA synthetase complexes in the cytoplasm of human cells. J. Biol. Chem. 2009, 284, 13746–13754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillon, C.; Goda, Y. The actin cytoskeleton: Integrating form and function at the synapse. Annu. Rev. Neurosci. 2005, 28, 25–55. [Google Scholar] [CrossRef]
- Lamprecht, R. The actin cytoskeleton in memory formation. Prog. Neurobiol. 2014, 117, 1–19. [Google Scholar] [CrossRef]
- Costa-Mattioli, M.; Gobert, D.; Harding, H.; Herdy, B.; Azzi, M.; Bruno, M.; Bidinosti, M.; Mamou, C.B.; Marcinkiewicz, E.; Yoshida, M. Translational control of hippocampal synaptic plasticity and memory by the eIF2α kinase GCN2. Nature 2005, 436, 1166–1173. [Google Scholar] [CrossRef] [Green Version]
- Tsokas, P.; Grace, E.A.; Chan, P.; Ma, T.; Sealfon, S.C.; Iyengar, R.; Landau, E.M.; Blitzer, R.D. Local protein synthesis mediates a rapid increase in dendritic elongation factor 1A after induction of late long-term potentiation. J. Neurosci. 2005, 25, 5833–5843. [Google Scholar] [CrossRef]
- Grallert, B.; Boye, E. GCN2, an old dog with new tricks. Biochem. Soc. Trans. 2013, 41, 1687–1691. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, G.C.; Smith, C.; Thomas, S.; Mandik-Nayak, L.; Laury-Kleintop, L.; Metz, R.; Muller, A.J. Indoleamine 2, 3-dioxygenase pathways of pathogenic inflammation and immune escape in cancer. Cancer Immunol. Immun. 2014, 63, 721–735. [Google Scholar] [CrossRef]
- Bauer, F.; Urdaci, M.; Aigle, M.; Crouzet, M. Alteration of a yeast SH3 protein leads to conditional viability with defects in cytoskeletal and budding patterns. Mol. Cell. Biol. 1993, 13, 5070–5084. [Google Scholar] [CrossRef] [Green Version]
- Amberg, D.C.; Basart, E.; Botstein, D. Defining protein interactions with yeast actin in vivo. Nat. Struct. Biol. 1995, 2, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Munn, A.L.; Stevenson, B.J.; Geli, M.I.; Riezman, H. end5, end6, and end7: Mutations that cause actin delocalization and block the internalization step of endocytosis in Saccharomyces cerevisiae. Mol. Biol. Cell 1995, 6, 1721–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivadon, P.; Bauer, F.; Aigle, M.; Crouzet, M. Actin cytoskeleton and budding pattern are altered in the yeast rvs161 mutant: The Rvs161 protein shares common domains with the brain protein amphiphysin. Mol. Gen. Genet. 1995, 246, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Vajjhala, P.; Lee, J.S.; Winsor, B.; Munn, A.L. The BAR domain proteins: Molding membranes in fission, fusion, and phagy. Microbiol. Mol. Biol. Rev. 2006, 70, 37–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallop, J.L.; McMahon, H.T. BAR domains and membrane curvature: Bringing your curves to the BAR. Biochem. Soc. Symp. 2005, 72, 223–231. [Google Scholar]
- Youn, J.-Y.; Friesen, H.; Kishimoto, T.; Henne, W.M.; Kurat, C.F.; Ye, W.; Ceccarelli, D.F.; Sicheri, F.; Kohlwein, S.D.; McMahon, H.T. Dissecting BAR domain function in the yeast Amphiphysins Rvs161 and Rvs167 during endocytosis. Mol. Biol. Cell 2010, 21, 3054–3069. [Google Scholar] [CrossRef] [Green Version]
- Colwill, K.; Field, D.; Moore, L.; Friesen, J.; Andrews, B. In vivo analysis of the domains of yeast Rvs167p suggests Rvs167p function is mediated through multiple protein interactions. Genetics 1999, 152, 881–893. [Google Scholar]
- Zhao, H.; Michelot, A.; Koskela, E.V.; Tkach, V.; Stamou, D.; Drubin, D.G.; Lappalainen, P. Membrane-sculpting BAR domains generate stable lipid microdomains. Cell Rep. 2013, 4, 1213–1223. [Google Scholar] [CrossRef] [Green Version]
- Sivadon, P.; Crouzet, M.; Aigle, M. Functional assessment of the yeast Rvs161 and Rvs167 protein domains. FEBS Lett. 1997, 417, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Balguerie, A.; Sivadon, P.; Bonneu, M.; Aigle, M. Rvs167p, the budding yeast homolog of amphiphysin, colocalizes with actin patches. J. Cell Sci. 1999, 112, 2529–2537. [Google Scholar]
- Brizzio, V.; Gammie, A.E.; Rose, M.D. Rvs161p interacts with Fus2p to promote cell fusion in Saccharomyces cerevisiae. J. Cell Biol. 1998, 141, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Feyder, S.; De Craene, J.-O.; Bär, S.; Bertazzi, D.L.; Friant, S. Membrane trafficking in the yeast Saccharomyces cerevisiae model. Int. J. Mol. Sci. 2015, 16, 1509–1525. [Google Scholar] [CrossRef] [Green Version]
- Idrissi, F.-Z.; Blasco, A.; Espinal, A.; Geli, M.I. Ultrastructural dynamics of proteins involved in endocytic budding. Proc. Natl. Acad. Sci. USA 2012, 109, E2587–E2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, M.D.; Ryazantsev, S.; Hicke, L.; Payne, G.S. Calmodulin promotes N-BAR domain-mediated membrane constriction and endocytosis. Dev. Cell 2016, 37, 162–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichte, B.; Veh, R.W.; Meyer, H.E.; Kilimann, M.W. Amphiphysin, a novel protein associated with synaptic vesicles. EMBO J. 1992, 11, 2521. [Google Scholar] [CrossRef] [PubMed]
- Sakamuro, D.; Elliott, K.J.; Wechsler-Reya, R.; Prendergast, G.C. BIN1 is a novel MYC-interacting protein with features of a tumour suppressor. Nat. Genet. 1996, 14, 69–77. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, M.; Hnia, K.; Gache, V.; Koch, C.; Gavriilidis, C.; Rodriguez, D.; Nicot, A.-S.; Romero, N.B.; Schwab, Y.; Gomes, E. Amphiphysin 2 orchestrates nucleus positioning and shape by linking the nuclear envelope to the actin and microtubule cytoskeleton. Dev. Cell 2015, 35, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Fernando, P.; Sandoz, J.S.; Ding, W.; de Repentigny, Y.; Brunette, S.; Kelly, J.F.; Kothary, R.; Megeney, L.A. Bin1 SRC homology 3 domain acts as a scaffold for myofiber sarcomere assembly. J. Biol. Chem. 2009, 284, 27674–27686. [Google Scholar] [CrossRef] [Green Version]
- Falcone, S.; Roman, W.; Hnia, K.; Gache, V.; Didier, N.; Laine, J.; Aurade, F.; Marty, I.; Nishino, I.; Charlet-Berguerand, N.; et al. N-WASP is required for Amphiphysin-2/BIN1-dependent nuclear positioning and triad organization in skeletal muscle and is involved in the pathophysiology of centronuclear myopathy. EMBO Mol. Med. 2014, 6, 1455–1475. [Google Scholar] [CrossRef]
- Demonbreun, A.R.; Quattrocelli, M.; Barefield, D.Y.; Allen, M.V.; Swanson, K.E.; McNally, E.M. An actin-dependent annexin complex mediates plasma membrane repair in muscle. J. Cell Biol. 2016, 213, 705–718. [Google Scholar] [CrossRef]
- Drager, N.M.; Nachman, E.; Winterhoff, M.; Bruhmann, S.; Shah, P.; Katsinelos, T.; Boulant, S.; Teleman, A.A.; Faix, J.; Jahn, T.R. Bin1 directly remodels actin dynamics through its BAR domain. EMBO Rep. 2017, 18, 2051–2066. [Google Scholar] [CrossRef] [PubMed]
- Wechsler-Reya, R.; Elliott, K.; Herlyn, M.; Prendergast, G.C. The putative tumor suppressor BIN1 is a short-lived nuclear phosphoprotein, the localization of which is altered in malignant cells. Cancer Res. 1997, 57, 3258–3263. [Google Scholar] [PubMed]
- Butler, M.H.; David, C.; Ochoa, G.-C.; Freyberg, Z.; Daniell, L.; Grabs, D.; Cremona, O.; De Camilli, P. Amphiphysin II (SH3P9; BIN1), a member of the amphiphysin/Rvs family, is concentrated in the cortical cytomatrix of axon initial segments and nodes of ranvier in brain and around T tubules in skeletal muscle. J. Cell Biol. 1997, 137, 1355–1367. [Google Scholar] [CrossRef] [PubMed]
- Casal, E.; Federici, L.; Zhang, W.; Fernandez-Recio, J.; Priego, E.-M.; Miguel, R.N.; DuHadaway, J.B.; Prendergast, G.C.; Luisi, B.F.; Laue, E.D. The crystal structure of the BAR domain from human Bin1/amphiphysin II and its implications for molecular recognition. Biochemistry 2006, 45, 12917–12928. [Google Scholar] [CrossRef] [Green Version]
- Wigge, P.; Kohler, K.; Vallis, Y.; Doyle, C.A.; Owen, D.; Hunt, S.P.; McMahon, H.T. Amphiphysin heterodimers: Potential role in clathrin-mediated endocytosis. Mol. Biol. Cell 1997, 8, 2003–2015. [Google Scholar] [CrossRef] [Green Version]
- Nicot, A.-S.; Toussaint, A.; Tosch, V.; Kretz, C.; Wallgren-Pettersson, C.; Iwarsson, E.; Kingston, H.; Garnier, J.-M.; Biancalana, V.; Oldfors, A. Mutations in amphiphysin 2 (BIN1) disrupt interaction with dynamin 2 and cause autosomal recessive centronuclear myopathy. Nat. Genet. 2007, 39, 1134–1139. [Google Scholar] [CrossRef]
- Wu, T.; Baumgart, T. BIN1 membrane curvature sensing and generation show autoinhibition regulated by downstream ligands and PI (4, 5) P2. Biochemistry 2014, 53, 7297–7309. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Marcucci, M.; Daniell, L.; Pypaert, M.; Weisz, O.A.; Ochoa, G.-C.; Farsad, K.; Wenk, M.R.; De Camilli, P. Amphiphysin 2 (Bin1) and T-tubule biogenesis in muscle. Science 2002, 297, 1193–1196. [Google Scholar] [CrossRef] [Green Version]
- Picas, L.; Viaud, J.; Schauer, K.; Vanni, S.; Hnia, K.; Fraisier, V.; Roux, A.; Bassereau, P.; Gaits-Iacovoni, F.; Payrastre, B. BIN1/M-Amphiphysin 2 induces clustering of phosphoinositides to recruit its downstream partner dynamin. Nat. Commun. 2014, 5, 5647. [Google Scholar] [CrossRef] [Green Version]
- Cowling, B.S.; Prokic, I.; Tasfaout, H.; Rabai, A.; Humbert, F.; Rinaldi, B.; Nicot, A.S.; Kretz, C.; Friant, S.; Roux, A.; et al. Amphiphysin (BIN1) negatively regulates dynamin 2 for normal muscle maturation. J. Clin. Investig. 2017, 127, 4477–4487. [Google Scholar] [CrossRef] [Green Version]
- Muller, A.J.; Baker, J.F.; DuHadaway, J.B.; Ge, K.; Farmer, G.; Donover, P.S.; Meade, R.; Reid, C.; Grzanna, R.; Roach, A.H. Targeted disruption of the murine Bin1/Amphiphysin II gene does not disable endocytosis but results in embryonic cardiomyopathy with aberrant myofibril formation. Mol. Cell. Biol. 2003, 23, 4295–4306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyrskyluoto, A.; Vartiainen, M.K. Regulation of nuclear actin dynamics in development and disease. Curr. Opin. Cell Biol. 2020, 64, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Lewinska, A.; Klukowska-Rotzler, J.; Deregowska, A.; Adamczyk-Grochala, J.; Wnuk, M. C-myc activation promotes cofilin-mediated F-actin cytoskeleton remodeling and telomere homeostasis as a response to oxidant-based DNA damage in medulloblastoma cells. Redox Biol. 2019, 24, 101163. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.; Sakamuro, D.; Basu, A.; Du, W.; Wunner, W.; Staller, P.; Gaubatz, S.; Zhang, H.; Prochownik, E.; Eilers, M. Bin1 functionally interacts with Myc and inhibits cell proliferation via multiple mechanisms. Oncogene 1999, 18, 3564–3573. [Google Scholar] [CrossRef] [Green Version]
- Pineda-Lucena, A.; Ho, C.S.; Mao, D.Y.; Sheng, Y.; Laister, R.C.; Muhandiram, R.; Lu, Y.; Seet, B.T.; Katz, S.; Szyperski, T. A structure-based model of the c-Myc/Bin1 protein interaction shows alternative splicing of Bin1 and c-Myc phosphorylation are key binding determinants. J. Mol. Biol. 2005, 351, 182–194. [Google Scholar] [CrossRef]
- Elliott, K.; Ge, K.; Du, W.; Prendergast, G.C. The c-Myc-interacting adaptor protein Bin1 activates a caspase-independent cell death program. Oncogene 2000, 19, 4669. [Google Scholar] [CrossRef] [Green Version]
- Lundgaard, G.L.; Daniels, N.E.; Pyndiah, S.; Cassimere, E.K.; Ahmed, K.M.; Rodrigue, A.; Kihara, D.; Post, C.B.; Sakamuro, D. Identification of a novel effector domain of BIN1 for cancer suppression. J. Cell. Biochem. 2011, 112, 2992–3001. [Google Scholar] [CrossRef] [Green Version]
- Prokic, I.; Cowling, B.S.; Laporte, J. Amphiphysin 2 (BIN1) in physiology and diseases. J. Mol. Med. 2014, 92, 453–463. [Google Scholar] [CrossRef]
- Al-Qusairi, L.; Laporte, J. T-tubule biogenesis and triad formation in skeletal muscle and implication in human diseases. Skelet. Muscle 2011, 1, 26. [Google Scholar] [CrossRef]
- Bitoun, M.; Maugenre, S.; Jeannet, P.-Y.; Lacene, E.; Ferrer, X.; Laforet, P.; Martin, J.-J.; Laporte, J.; Lochmüller, H.; Beggs, A.H. Mutations in dynamin 2 cause dominant centronuclear myopathy. Nat. Genet. 2005, 37, 1207–1209. [Google Scholar] [CrossRef]
- Lionello, V.M.; Nicot, A.S.; Sartori, M.; Kretz, C.; Kessler, P.; Buono, S.; Djerroud, S.; Messaddeq, N.; Koebel, P.; Prokic, I.; et al. Amphiphysin 2 modulation rescues myotubular myopathy and prevents focal adhesion defects in mice. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Harold, D.; Abraham, R.; Hollingworth, P.; Sims, R.; Gerrish, A.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; Dowzell, K.; Williams, A. Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat. Genet. 2009, 41, 1088–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belbin, O.; Carrasquillo, M.M.; Crump, M.; Culley, O.J.; Hunter, T.A.; Ma, L.; Bisceglio, G.; Zou, F.; Allen, M.; Dickson, D.W. Investigation of 15 of the top candidate genes for late-onset Alzheimer’s disease. Hum. Genet. 2011, 129, 273–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holler, C.J.; Davis, P.R.; Beckett, T.L.; Platt, T.L.; Webb, R.L.; Head, E.; Murphy, M.P. Bridging integrator 1 (BIN1) protein expression increases in the Alzheimer’s disease brain and correlates with neurofibrillary tangle pathology. J. Alzheimers Dis. 2014, 42, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapuis, J.; Hansmannel, F.; Gistelinck, M.; Mounier, A.; Van Cauwenberghe, C.; Kolen, K.; Geller, F.; Sottejeau, Y.; Harold, D.; Dourlen, P. Increased expression of BIN1 mediates Alzheimer genetic risk by modulating tau pathology. Mol. Psychiatr. 2013, 18, 1225–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sottejeau, Y.; Bretteville, A.; Cantrelle, F.-X.; Malmanche, N.; Demiaute, F.; Mendes, T.; Delay, C.; Dos Alves, H.A.; Flaig, A.; Davies, P. Tau phosphorylation regulates the interaction between BIN1′s SH3 domain and Tau’s proline-rich domain. Acta Neuropathol. Commun. 2015, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Sartori, M.; Mendes, T.; Desai, S.; Lasorsa, A.; Herledan, A.; Malmanche, N.; Makinen, P.; Marttinen, M.; Malki, I.; Chapuis, J.; et al. BIN1 recovers tauopathy-induced long-term memory deficits in mice and interacts with Tau through Thr(348) phosphorylation. Acta Neuropathol. 2019, 138, 631–652. [Google Scholar] [CrossRef] [Green Version]
- Calafate, S.; Flavin, W.; Verstreken, P.; Moechars, D. Loss of bin1 promotes the propagation of tau pathology. Cell Rep. 2016, 17, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Miyagawa, T.; Ebinuma, I.; Morohashi, Y.; Hori, Y.; Young Chang, M.; Hattori, H.; Maehara, T.; Yokoshima, S.; Fukuyama, T.; Tsuji, S.; et al. BIN1 regulates BACE1 intracellular trafficking and amyloid-beta production. Hum. Mol. Genet. 2016, 25, 2948–2958. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mammals | S. cerevisiae |
|---|---|
| PSTPIP1 | Hof1p (cdc15p in S. pombe) |
| BIN1 | Rvs167p |
| IMPACT | Yih1p |
| GCN2 (in humans also eIF2AK4) | Gcn2p |
| eEF1A1 and eEF1A2 | eEF1a (or Tef1p) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akram, Z.; Ahmed, I.; Mack, H.; Kaur, R.; Silva, R.C.; Castilho, B.A.; Friant, S.; Sattlegger, E.; Munn, A.L. Yeast as a Model to Understand Actin-Mediated Cellular Functions in Mammals—Illustrated with Four Actin Cytoskeleton Proteins. Cells 2020, 9, 672. https://doi.org/10.3390/cells9030672
Akram Z, Ahmed I, Mack H, Kaur R, Silva RC, Castilho BA, Friant S, Sattlegger E, Munn AL. Yeast as a Model to Understand Actin-Mediated Cellular Functions in Mammals—Illustrated with Four Actin Cytoskeleton Proteins. Cells. 2020; 9(3):672. https://doi.org/10.3390/cells9030672
Chicago/Turabian StyleAkram, Zain, Ishtiaq Ahmed, Heike Mack, Ramandeep Kaur, Richard C. Silva, Beatriz A. Castilho, Sylvie Friant, Evelyn Sattlegger, and Alan L. Munn. 2020. "Yeast as a Model to Understand Actin-Mediated Cellular Functions in Mammals—Illustrated with Four Actin Cytoskeleton Proteins" Cells 9, no. 3: 672. https://doi.org/10.3390/cells9030672