The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis


Department of Cytophysiology, Faculty of Biology and Environmental Protection, University of Lodz, 90-236 Lódź, Poland
Cells 2020, 9(3), 606; https://doi.org/10.3390/cells9030606
Submission received: 4 February 2020
/
Revised: 21 February 2020
/
Accepted: 27 February 2020
/
Published: 3 March 2020
(This article belongs to the Section Plant, Algae and Fungi Cell Biology)
{kind=link}
{kind=link}
{kind=link}
Abstract
:In plants, the first asymmetrical division of a zygote leads to the formation of two cells with different developmental fates. The establishment of various patterns relies on spatial and temporal gene expression, however the precise mechanism responsible for embryonic patterning still needs elucidation. Auxin seems to be the main player which regulates embryo development and controls expression of various genes in a dose-dependent manner. Thus, local auxin maxima and minima which are provided by polar auxin transport underlie cell fate specification. Diverse auxin concentrations in various regions of an embryo would easily explain distinct cell identities, however the question about the mechanism of cellular patterning in cells exposed to similar auxin concentrations still remains open. Thus, specification of cell fate might result not only from the cell position within an embryo but also from events occurring before and during mitosis. This review presents the impact of auxin on the orientation of the cell division plane and discusses the mechanism of auxin-dependent cytoskeleton alignment. Furthermore, close attention is paid to auxin-induced calcium fluxes, which regulate the activity of MAPKs during postembryonic development and which possibly might also underlie cellular patterning during embryogenesis.
Keywords:
MAPK; embryogenesis; cellular patterning; microtubules; actin filaments; auxin; calcium ions1. Introduction
Embryogenesis is driven by sophisticated and orchestrated processes which are kept under the control of various molecular pathways. Phenotypic analyses of mutants and studies with the use of protein inhibitors provide clear evidence that three molecular pathways are indispensable for a correct embryo development, and these include polar auxin transport, mitogen-activated protein kinases and DNA methylation. Dysfunction of these molecular pathways leads to embryogenesis disorders and may be lethal for the developing embryo [1,2,3]. In angiosperms after fertilization, a zygote elongates in micropylar-chalzal axis and divides asymmetrically. In this way the extraordinary process begins which leads to development of the functional embryo [4,5,6]. Since auxin was found to be responsible for cell extension according to the direction of auxin flow [7], it seems that the polar auxin transport may determine the direction of the zygote elongation. Although the disruption of the polar auxin transport manifests itself mainly during the apical-basal axis formation, some disorders of embryogenesis are also noticeable at earlier stages [1,8,9]. The process of zygote elongation is crucial for the first asymmetrical division, as a result of which a small apical and a large basal cells are created, both already with different gene expression [10,11,12]. Detailed research on the plant embryogenesis indicates that the asymmetrical division of the zygote relies both on microtubules, which control the zygote elongation, and on actin filaments, which regulate polar migration of nuclei [13,14].
After early cell divisions, the embryo starts to develop specific tissues. Different tissues were found to have unique profiles of activated genes [10,15,16,17], however the precise mechanism responsible for the diverse gene expression needs elucidation and the question when and how cellular patterning is established still remains open. The general plane of embryogenesis and the accuracy of this process incline to state that not only molecular pathways mentioned above but also mutual relationships between them and other events should be taken into consideration. Since auxin is a plant phytohormone controlling the cell polarity, the orientation of the cell division and expression of auxin-dependent genes, one could ask how auxin regulates cytoskeleton alignment and establishment of various gene expression during embryogenesis. This review presents cytophysiological responses to auxin which affect cytoskeleton alignment, and show that gene expression may contribute to cellular patterning.
2. Role of Cytoskeleton in Embryo Development
With the first division of a zygote, embryogenesis follows the patterns which allow it to develop a final profile of an embryo. Thus, mechanisms which regulate cell shape and interactions between them must be taken into consideration. Replication and cell division are energy consuming processes, therefore it is not surprising that cells tend to reduce energy expenditure during their lifespan. The shortest wall rule assumes that cell wall after mitosis is created according to a surface which needs minimum energy [18,19,20,21].
Throughout anisotropic growth cells display transverse pattern of cortical microtubules. This alignment was found in developing trichomes [22,23] and in a root elongation zone [24]. Thus, rectangular cells usually divide transversely, however, there are exceptions to this pattern during morphogenesis, and response to mechanical stress is a good example when the shortest wall rule is broken [25,26,27]. Mechanical stress regulates microtubule orientation at subcellular and tissue levels and they were found to align along the direction of maximal stress in epidermal cells of Arabidopsis thaliana [27]. Interestingly, orientation of cortical microtubules was found to be parallel to localization of PIN-FORMED (PIN) proteins, one of the plasma membrane auxin transporters. Mechanical stress induces both circumferential localization of microtubules around the site of cell ablation and translocation of PIN proteins to membranes which are away from the place of damage, however parallel alignment of microtubules and PINs was maintained [28,29]. During morphogenesis, mechanical stress within tissues may be induced when neighboring cells grow faster and PINs seem to be recruited to the membrane with the highest tensile stress [30]. Explanation of this behavior may come from the mechanism of PINs cycling between cell membranes which utilize endocytosis and exocytosis. It was found that high tension of plasma membrane inhibited endocytosis, however exocytosis might reduce the emerging tension. Therefore, PINs density was found to increase when plasma membrane tension enlarges [31]. Thus, it is very plausible that tension stress regulates microtubule alignment during embryogenesis and might be responsible for breaking the rule of division along the shortest cell wall. Furthermore, parallel alignment of microtubules and PIN proteins indicates that auxin must cooperate in this process.
Actin filaments (F-actin), another cytoskeleton component, were found to play a role in the directional cell growth as well. Two fractions of F-actin bundles exist inside cells, one which decorates plasma membrane (cortical F-actin) and the other which polymerizes in cytoplasm [32]. Longitudinal bundling of cytoplasmic F-actin takes part in trichomes development, root hair growth [22,23,33], pollen tube expansion [32,34] and anisotropy growth of hypocotyl cells [24]. Depending on localization, F-actin bundles display different impact on cellular molecules, vesicle transport and finally cell polarization. Cortical F-actin bundles seem to act as a physical barrier for vesicle docking and transport of molecules through plasma membrane. On the other hand, directional growth of cytoplasmic F-actin bundles was found to be responsible for polarized trafficking. Thus, cortical and cytoplasmic F-actin bundles co-regulate directional transport of molecules, creating a physical barrier and triggering polar localization of plasma membrane transporters [35,36,37,38,39,40]. Interestingly, cortical microtubules and actin filaments were found to coalign [32,41], which may indicate their mutual dependence [24,42]. It seems to be very plausible that the alignment of cortical microtubules and F-actin bundles by plasma membrane reduces transport at the site of their polymerization and together with cytoplasmic F-actin, they support directional transfer of molecules and finally anisotropic growth. However, the question of whether the opposite alignment of cytoplasmic F-actin and cortical microtubules depends on each other still remains open.
It has been previously shown that microtubule alignment is strictly connected with the division plane between 2- and 16-cell stages of plant embryogenesis [43,44]. Thus, it seems that elements of cytoskeleton with their impact on mechanical properties of cells and signaling pathways which control cytoskeleton alignment must play a prominent role in the determination of the division plane during embryogenesis. Studies on embryo development indicate that similarly to post-embrionic growth, it relies on mitoses which are beyond the shortest wall rule, and the first two divisions of the small apical cell in A. thaliana embryos take place in the longitudinal plane [21]. It is likely that tensile stress impacting the alignment of microtubules and actin filaments regulates the directional transport, the cell polarization and finally the direction of future division. This mechanism may be conservative and takes part both in embryo development and in post-embryonic growth. However, it seems that the response to mechanical stress solely may not be the only mechanism responsible for regulation of cell division plane and cellular patterning.
Microtubule interactions such as zippering, catastrophe or crossover, which underlie changes in cytoskeleton alignment, seem to impact the correct embryo development as well. The first term describes the state when two microtubules meet at an angle smaller than 40° and continue to growth next to each other, the second when microtubules shrink after meeting at an angle larger than 40° and the last when two microtubules slip over each other. Computational analyses indicate that the probability of transverse and longitudinal alignment is nearly equal, however the situation dramatically changes when edge-catastrophe and enhanced stability of microtubules at new division site are taken into consideration [43,44]. The edge-catastrophe, i.e., shrinking of the microtubules which reached cell edge and high probability of their collision make molecular apparatus regulating microtubule dynamics indispensable during growth along the shortest cell wall. Due to the limited amount of free tubulin and the dynamic instability [45], short microtubules with short lifespan would be more favorable when edge-catastrophe is diminished. Thus, transverse alignment seems to be the default orientation of microtubules in cells revealing anisotropic growth.
3. Cytoskeleton Elements are Regulated by Auxin
Auxin was postulated to be the main player which regulates microtubule growth, however some opposing results have been demonstrated so far. Although exogenous auxin triggers the transverse position of cortical microtubules in hypocotyl cells [46,47], contradictory studies indicate that an increase in auxin concentration induces inhibition of cell elongation [48] which is accompanied by reorientation of cortical microtubules from transverse to longitudinal alignment. This behavior was observed in the transition zone of root and in the elongation zone of hypocotyl. Inhibition of cell elongation in response to auxin relies on auxin binding protein 1 (ABP1) [49] and it was found to be non-transcriptional-dependent [46,49,50]. On the other hand, aberrant horizontal divisions of an apical cell and embryogenesis disorders in mp (auxin response factor) and bdl (Aux-IAA repressor) mutants [51,52] may lead to the conclusion that regulation of microtubule alignment is transcriptional-dependent. However, it must be taken into consideration that embryos with those mutant proteins display impaired polar auxin transport which prevents establishment of the correct auxin maxima. Thus, it appears that microtubule alignment depends indirectly on the expression of auxin-inducible genes which are responsible for final auxin localization, and the direct regulation of microtubule ordering seems not to depend on transcription.
A high auxin level induces cortical microtubule disorganization during anisotropic expansion and supports the shift into isotropic cell growth [18,53,54]. Microtubule alignment along the longest cell wall after exogenous auxin application might be extrapolated to embryogenesis and would explain orientation of the division plane in embryonic cells. In A. thaliana after the first division of a zygote, auxin is transported toward the apical cell due to polar localization of PIN7 proteins [55]. The apical cell elongates slightly in the direction of auxin flow [56,57,58,59], however progressive auxin accumulation [60] would break default transverse alignment of microtubules and promote longitudinal division in this way. Interestingly, anisotropic growth of the apical and basal cells seems to be necessary for correct embryogenesis. A mutation in genes encoding polygalacturonases, enzymes degrading pectins, reduces the length of those cells leading to slower embryogenesis [59]. Furthermore, the regulation of a division plane orientation through the creation of auxin maxima may be supported indirectly by the studies which indicate longitudinal mitoses of the apical cell regenerated from a suspensor [61,62].
Auxin regulates not only microtubule alignment but also the growth of cytoplasmic F-actin filaments. Although there are no studies showing the impact of auxin level on direction of F-actin growth, auxin was found to play a role in actin filament rearrangements or actin-dependent transport. It was suggested that auxin regulated the organization of fine F-actin bundles, providing an efficient transport of signaling proteins toward the cell pole, and reduction of polar auxin transport was found to initiate actin filament bundling which may detain polar distribution of auxin transporters [34,63,64,65,66]. Thus, dissociation of actin bundles might be necessary for redistribution of PIN proteins to the plasma membrane [63,64]. Furthermore, auxin induces the production of phosphatidic acid which may regulate the organization of actin filaments and the vesicle trafficking, therefore it might have impact on the final PIN localization [67].
Since auxin regulates alignment of cytoskeleton elements and exogenous auxin was found to induce the microtubule reorientation from transverse to longitudinal during postembryonic development [46,49,50], analogous role of auxin is possible during embryogenesis. Thus, one could ask what are the players which stand behind the shift from transverse to longitudinal plane of an embryonic cell division?
4. The Players Regulating Auxin-Dependent Microtubule Alignment During Embryogenesis
Computational analyses show that the auxin-mediated microtubule stability at a new division site and the CLASP-mediated reduction of edge-catastrophe are the key factors regulating the division plane orientation during early stages of embryogenesis [43,44]. Nevertheless, the authors of these papers propose that alignment of cortical microtubules may also depend on the direction of mechanical stress at further stages of embryogenesis and they do not exclude both mechanisms acting synergistically. Enhanced microtubule stability at a new division site may explain their longitudinal alignment and the vertical division of the apical cell in A. thaliana, however it does not seem to be the general rule since the apical cell in some species divides horizontally [68,69,70]. Transverse microtubule bundles relay on the CLASP proteins which stabilizes plus-end of severed microtubules [71] and protects them from a catastrophe when they reach cell cortex [46,50]. However, transverse microtubule alignment is maintained in clasp-1 mutants during postembryonic growth, and only their abundance was found to be reduced [72]. Furthermore, a dysfunction of CLASP protein does not affect vertical divisions of the apical cell at the beginning of embryogenesis [43,44]. Thus, it seems reasonable to ask how auxin regulates microtubule alignment and the orientation of the division plane in embryonic cells.
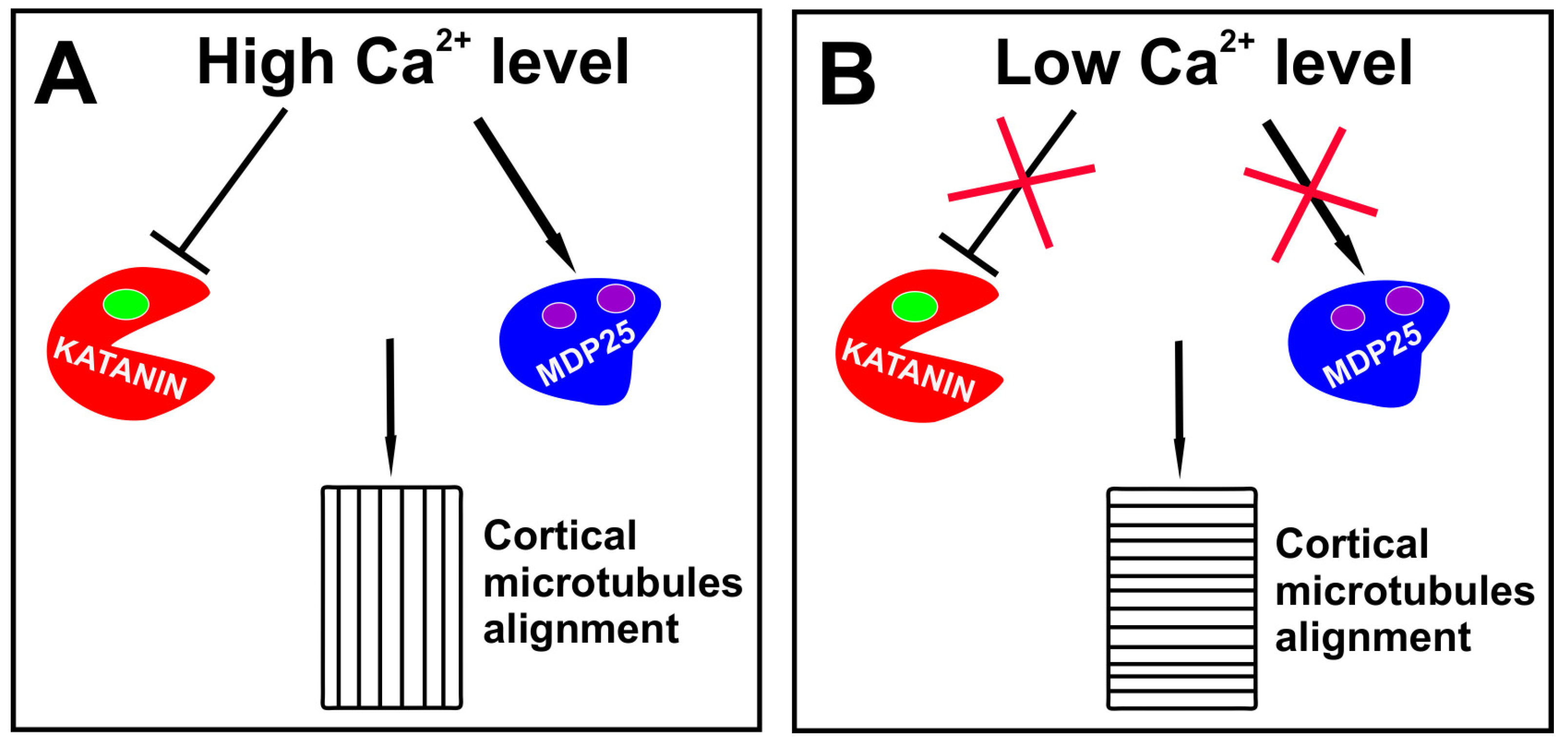
It is possible that other proteins regulating microtubule behavior are responsible for cytoskeleton rearrangements during embryo development and I concentrate on two proteins whose specific functions during embryogenesis still need elucidation. Katanin, the microtubule-severing protein, was found to be indispensable for the transverse microtubule alignment during postembryonic development [73,74] and disorders of embryogenesis in katanin mutants were previously shown [75]. Although auxin activates the katanin activity via the ROP6-RIC1 pathway [54,73,76,77], a high auxin level induces calcium release from intracellular reservoirs and the external space [78] which at an elevated concentration may inhibit the katanin activity without changing its ability to bind substrates [79]. Therefore, it is possible that due to high auxin level in the apical cell, katanin activity is reduced and the transverse alignment of microtubules is not supported. Calcium ions were also found to accelerate the activity of MDP25, one of the microtubule destabilizing proteins. Consequently, a high auxin level both reduces the severing activity of katanin and intensifies the MDP25 destabilizing action, leading to a higher frequency of an encounter-based catastrophe of transversely growing microtubules, and therefore making the longitudinal ordering more favorable (Figure 1A). Furthermore, the depolimerization of short microtubules increases the amount of free tubulin, which may be utilized by those microtubules growing longitudinally and having a smaller chance for collision. An oblique or longitudinal array of microtubules in the cells overexpressing MDP25 may support the role of this protein in the shift from transverse to longitudinal alignment [80,81]. Auxin was also found to increase the level of phosphatidic acid which mediates through MAP65 (one of microtubule bundling protein) attachment of microtubules to plasma membrane [82,83], and therefore it could promote higher stability of longitudinally growing microtubules.
Although some studies indicate that PIN localization and microtubule alignment are independent of auxin in specific situations [28,29], a transcriptional response to auxin and polar auxin transport were found to be necessary both for overriding the shortest wall rule and for asymmetric division, which ensure the correct embryo development [21]. A mechanical stress and an auxin-dependent response were proposed as the two mechanisms responsible for changes of cell shape in a shoot apical meristem [84]. It seems that auxin plays a crucial role in correct embryogenesis and mechanical stress may act as a complementary mechanism modulating the orientation of the division plane or might be switched on when necessary.
If the mechanism employing katanin and MDP25 protein is responsible for longitudinal alignment of microtubules during embryogenesis, one could ask how microtubule ordering is changed to drive a horizontal plane of the cell divisions. Since high intracellular level of calcium ions is temporary [85], it is possible that after two longitudinal divisions of the apical cell, the level of free calcium ions declines or the activation of calcium ion channels is not intensive enough, so that the severing function of katanin is maintained and the destabilizing activity of MDP25 is silenced. This may increase the stability of short microtubules and support the transverse microtubule alignment (Figure 1B).
5. Calcium Ions at the Service of Auxin
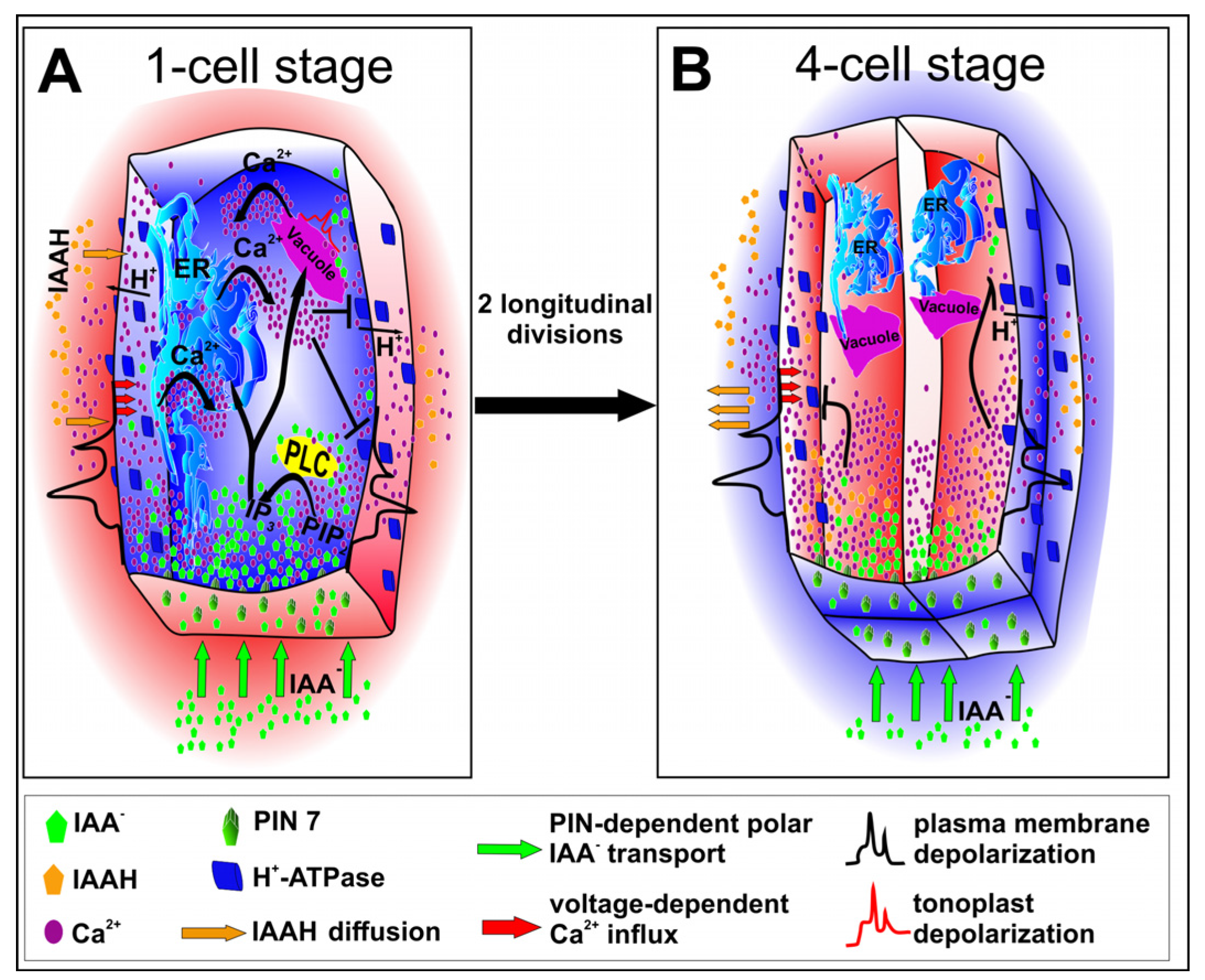
Auxin induces a dose-dependent depolarization of plasma membrane which results in calcium influx. It was also found to activate a plasma membrane H+-ATPase which pumps H+ ions to apoplast, and thereby provides a low pH in the extracellular space. Since apoplast acidification increases the membrane potential, the activity of the H+-ATPase may be considered as one of the mechanisms which regulate the extent of calcium influx. Interestingly, although the membrane depolarization continued, calcium concentration was found to return to the baseline level [86,87,88,89]. Elevation of calcium ion levels in cytoplasm results in their subsequent release from vacuole [90,91]. Auxin seems to participate in amplification of calcium signaling, since it activates a phospholipase C (PLC) [82] whose optimal activity depends on calcium ions [92,93]. The PLC hydrolizes phosphatidylinositol 4,5-bisphosphate (PIP2) to inositol-1,4,5-triphosphate (IP3) which is a messenger molecule. IP3 by binding to receptors in the endoplasmic reticulum (ER) induces calcium efflux from the lumen [67,94,95]. Furthermore, starting from one cell embryo, small vacuoles are present in cytoplasm [96,97] and they may also be a source of calcium ions. The tonoplast is decorated with voltage-dependent slow vacuolar (SV) channels and ligand-gated channels. The former may be activated by calcium ions and auxin was found to enhance these channels activity. The latter, similarly to ER, are activated by IP3 [98]. Interestingly, calmodulin, an adaptor of calcium signals, was found to modulate SV channels activity [99]. Calcium signaling during plant embryogenesis is not studied in detail, however it has been shown so far that calcium ions [100] and calmodulin [101,102] are indispensable during somatic embryogenesis. Thus, considering calcium oscillation during postembryonic development, it is worth asking whether two steps of calcium ions accumulation could take place also during embryogenesis. A hypothetical model of calcium influx which could impact cytoskeleton alignment during embryogenesis is presented in Figure 2. First, the calcium level elevates due to fluxes through plasma membrane and after it reaches an adequate level inside a cell, calcium efflux from vacuoles and ER is activated. Thus, it is possible that due to a progressive accumulation of auxin in a apical cell and in daughter cells after the first division, the concentration of calcium ions and the time of their action are sufficient to impact microtubule regulating proteins and to induce the longitudinal alignment of microtubules (Figure 1A and Figure 2A).
This model might also explain the shift to transverse alignment at further stages of embryogenesis. Following two longitudinal mitosis, the volume of an apical cell becomes nearly 4-fold smaller [21]. Theoretically, the surface of the basal membrane, which ensures directional auxin transport, is also 4-fold smaller, however, the surface of vertical membranes of daughter cells is only halved. Thus, it seems possible that the level of calcium ions is reached much faster in daughter cells after the second division of the apical cell and their high concentration in cytoplasm triggers the inhibition of plasma membrane H+-ATPase much earlier, leading to the cytosol acidification and apoplast alkalization, similarly as it happens during postembryonic development [103,104,105]. Under low intracellular pH, both voltage-dependent channels in a vacuole are inhibited [106] and auxin appears in the protonated form (IAAH) which, due to lipophilic properties, may cross lipid bilayer [107]. It is possible that the cytosol acidification decreases the auxin concentration and reduces the activation of calcium channels in ER or vacuole, thereby prevents further accumulation of calcium ions and their long-term action (Figure 2B). Thus, under low calcium level severing katanin activity is maintained in contrast to MDP25 activity. This provides the transverse microtubule alignment, which could result in the horizontal direction of the third division (Figure 1B).
6. Asymmetric Division Depends on Auxin
The asymmetric division which generates daughter cells with unequal volumes may have an impact on the cell fate during embryogenesis. At a 16-cell embryo stage, the asymmetrical mitosis occurs in an outer and inner cell of the lower tier. The former divides periclinally, giving a larger apical and smaller basal cell, while the latter divides anticlinally, generating a larger outer ground tissue precursor and a smaller inner vascular precursor [21]. The orientation of cell division, especially at further stages of embryogenesis may depend on auxin gradient and tensions generated between cells [44]. As far as both mechanisms explain the direction of cell division, could they elucidate the choice between symmetrical and asymmetrical divisions?
Although alignment of cortical microtubules and the formation of preprophase band were found to dictate the direction of cell division, actin filaments seem to regulate the polar migration of nuclei which heralds upcoming unequal mitosis [13,108]. Furthermore, in animal and yeast cells, actin filaments were found to position the mitotic spindle, and thus to have an impact on the symmetry of cell division [14]. Unequal mitosis during stomata or root development depends on auxin and PIN proteins, and inhibitors of a polar auxin transport were found to restrain the polar migration of nuclei in plant cells [109,110,111]. It seems that the choice between symmetrical or asymmetrical mitosis depends on actin filaments whose alignment is regulated by auxin. Although the direction and the symmetry of cell division depend on cytoskeleton alignment and auxin transport, this does not explain directly the mechanism in which the spatiotemporal expression of specific genes is established during embryo development.
7. New Mechanism Regulating Intracellular Asymmetry and Cellular Patterning
Cellular patterning starts very early during embryogenesis and already after the first division of a zygote different genes are expressed in apical and basal cells. Many genes are known to be differently expressed in various cells and tissues and their impact on the cellular patterning is well established [4,6,10,112]. Following two longitudinal divisions of an apical cell, the same genes were found to be activated in embryo proper, however, after transverse divisions, when an embryo reaches 8-cell stage, upper and bottom tiers display different gene expression which is controlled by auxin threshold [60,113,114]. Thus, auxin transporters, whose expression is induced by high auxin level [115,116], regulate polar auxin transport and thereby generate auxin maxima in different tissues. A family of auxin transporters is a good example of genes differently expressed in various cells. LAX1, one of the auxin influx carriers, is restricted to embryo proper and once an embryo reaches 32-cell stage, it shows higher expression in the upper tier. LAX2 was not detected at globular stage, however following the 32-cell stage, this protein is restricted to provascular cells and hypophysis. PIN proteins, auxin efflux carriers, also display cell-specific localization. For example, PIN7 is restricted to suspensor, PIN4 to hypophysis and PIN1 was generally found expressed in embryo proper. However, PIN1 displays no polar localization before 16-cell stage and its polarity is established during the next steps of embryogenesis. Interestingly, PIN1 is mainly localized in a basal membrane of provascular cells and in an apical membrane in outermost cells [1,5,8,9,10,11,55]. Another example of genes differently expressed in various cells is the WOX gene family which encodes transcription factors that regulate plant development. WOX9 and WOX2 were found to be expressed in an apical and central domain but WOX8 in hypophysis and suspensor only [6,16,60].
Expression of various genes in particular regions of a developing embryo raises a question about the timing and the way in which these cellular patterns are established. One could ask whether they are set up after the division as a result of the cell position and hence different auxin level or maybe before mitosis as a result of intracellular auxin gradients.
The existence of embryo regions with different auxin levels may lead to the conclusion that the cell position within an embryo plays a major role during cellular patterning. Furthermore, inversion of root meristem to shoot meristem or changing the shoot meristem patterns to root meristem which depend on auxin level [117,118,119,120] may support this idea. Although cell position and the resulting exposure to different auxin concentrations explain well cell fate at the level of organs, it does not explicate the development of cells with different identities, which are located in a region displaying homogenous auxin level.
It was previously described that MAPKs were indispensable during embryogenesis and that mutations in genes of MAPK pathway lead to reduced elongation of zygote and symmetrical cell division [4]. MAPKs control expression of different genes through interaction with transcription factors or regulate other cellular processes by targeting cytoplasmic proteins, e.g., in rice MAPKs were found to interact with Aux/Lax1 [121,122,123]. Studies on Vicia faba embryos revealed a new and so far not described localization of MAPKs. The phosphorylated form of these kinases was found in the vicinity of chromosomes and auxin seems to regulate the number of mitoses with MAPKs and the number of spots with activated kinases localized on the surface of chromosomes. Interestingly, the number of spots differs between groups of separating chromatids during anaphase and telophase [124]. This phenomenon raises the question about the mechanism which could regulate unequal distribution of MAPKs in the vicinity of chromosomes. During postembryonic development and stress response, phosphorylation of MAPKs is controlled by dual-specificity phosphatases (DUSPs). Calmodulin, which is a calcium sensor, was found to increase the activity of dual-specificity phosphatases (DUSPs) indicating a link between calcium ions oscillation and MAPKs phosphorylation [125,126]. Asymmetrically distributed calcium ions were well documented in highly elongated cells such as growing pollen tubes [127,128], and exogenous auxin was found to induce calcium ions release to cytoplasm [78]. However, intracellular gradients of calcium and auxin are difficult to follow during plant embryogenesis. Nevertheless, intracellular auxin gradient was detected directly in cells during proliferative period of Chara vulgaris [129] and this gradient might exist in embryonic cells as well.
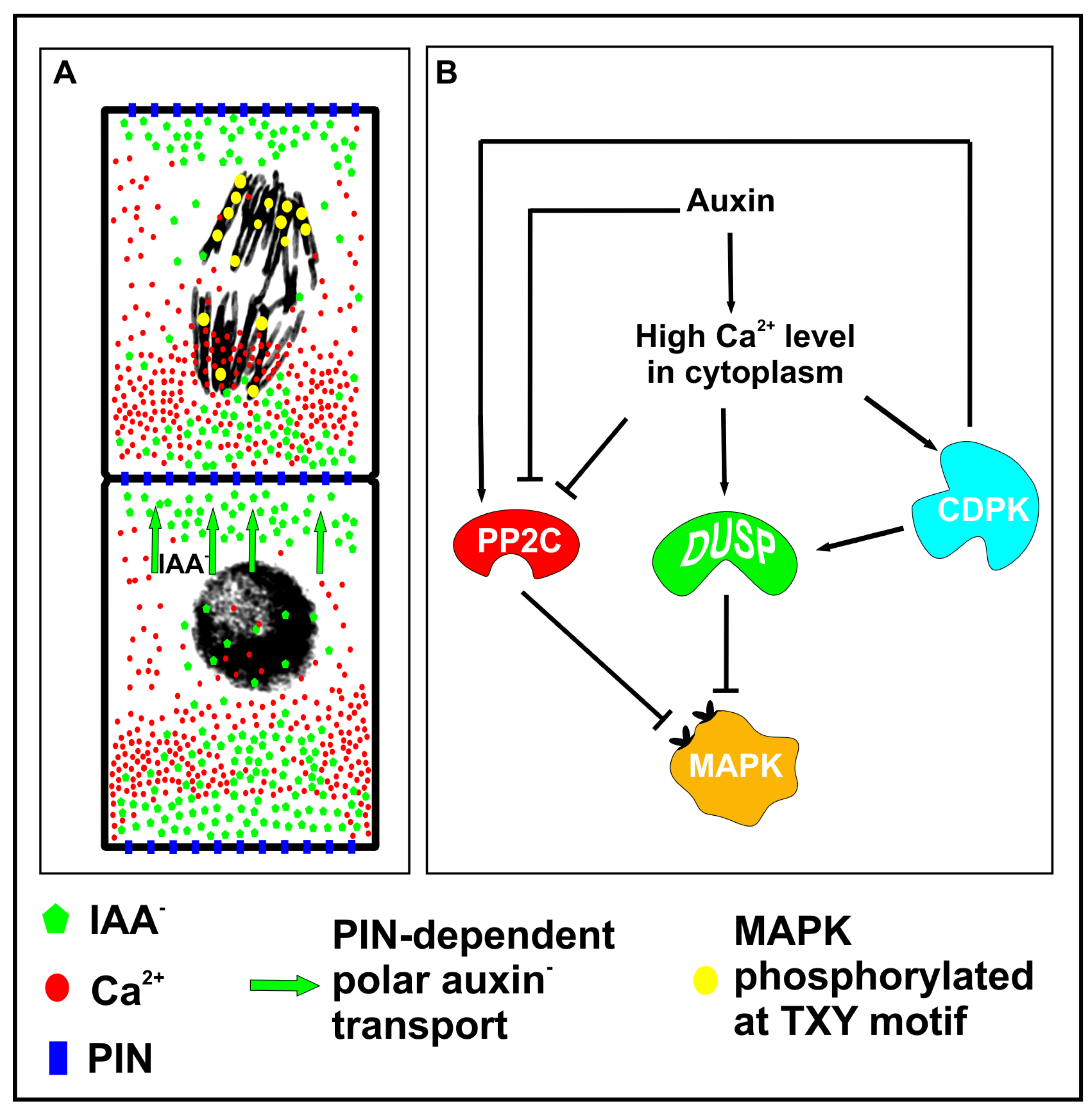
It is possible that, due to auxin gradient, locally activated DUSPs dephosphorylate MAPKs in their conservative TXY motif, and thereby reduce the number of activated MAPKs in the vicinity of those chromatids which are exposed to a high calcium content. Calcium dependent dephosphorylation of MAPKs may also be supported by studies showing that the function of PP2C kinase, another potential MPAKs phosphatase, is regulated by calcium-dependent protein kinase (CDPK) [130]. However, some studies indicate a reduced activity of PP2C under a high concentration of auxin or calcium ions (Figure 3A,B) [131,132,133]. Another explanation of asymmetric MAPK distribution may come from regulation of chromatin structure or segregation of molecules in cytoplasm. Chromosomes exposed to calcium ions display compact structure [134], thus it is possible that the level of chromatin condensation might differ between groups of chromatids due to calcium gradients, and in this way calcium ions could facilitate or reduce DNA accessibility for MAPKs. Thus, auxin-induced calcium fluxes might regulate the number of phosphorylated MPAKs in the vicinity of chromosomes, as they might trigger the activation of phosphatases or they might control chromatin compaction which reduces accessibility of DNA for MAPKs. Interestingly, calcium ion gradient might induce an electrical field which would segregate molecules by electrophoresis [135,136] and this could be another mechanism regulating protein composition at both ends of a dividing cell. It is very plausible that MAPKs participate in cellular patterning before cell division is completed, and due to their unequal distribution in separating chromatids, they execute the asymmetry of intracellular gradients of auxin and calcium molecules. Mammalian transcription factors (TFs) were found to associate with mitotic chromosomes and their role in regulation of cell fate by controlling post-mitotic gene reactivation was previously postulated [137,138]. It seems that MAPKs may act in the same way as pioneer TFs and mark the genes which will be activated in daughter cells early during mitotic exit.
8. Conclusions
Cellular patterning, which starts with the first division of a zygote, seems to be executed not only by regulation of different gene expression but also through factors which control the direction and the symmetry of cell division. Auxin, which is multifunctional plant phytohormone, regulates post-embryonic development and response to stress stimuli. They were also found to play a considerable role in cellular patterning during embryogenesis. Although many genes and molecular pathways regulated by auxin are well known, precise mechanisms which underlie auxin-dependent control of embryogenesis still need elucidation. Auxin was found to regulate the release of calcium ions to cytoplasm and it is possible that their accumulation in cytoplasm takes place in two stages. Thus, the direction and the symmetry of cell division which is controlled by cytoskeleton alignment might depend not only on high level of calcium ions, but also on the duration of their action. Furthermore, chromosomal localization of MAPK might indicate a new function of these kinases during cellular patterning. Nevertheless, exactly which of many MAPKs identified in a plant genome play the prominent role in this process and how MAPKs in the vicinity of chromosome impact gene expression is yet to be determined.
Funding
This research received no external funding.
Acknowledgments
This work is dedicated to the memories of my father and professor Maria Kwiatkowska, a great scientist fascinated with ultrastructure of plant cells, cytoskeleton components and the nature of lipotuboloids, the organizer of Department of Cytophysiology and the former Dean of the Faculty of Biology and Environmental Protection. I thank M. Fronczak for English language corrections.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Robert, H.S.; Grunewald, W.; Sauer, M.; Cannoot, B.; Soriano, M.; Swarup, R.; Weijers, D.; Bennett, M.; Boutilier, K.; Friml, J. Plant embryogenesis requires aux/lax-mediated auxin influx. Development 2015, 142, 702–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiwongsar, S.; Strohm, A.K.; Su, S.-H.; Krysan, P.J. Genetic analysis of the Arabidopsis protein kinases MAP3Kε1 and MAP3Kε2 indicates roles in cell expansion and embryo development. Front. Plant Sci. 2012, 3, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckardt, N.A. Genetic and epigenetic regulation of embryogenesis. Plant Cell Online 2006, 18, 781–784. [Google Scholar] [CrossRef]
- Bayer, M.; Slane, D.; Jürgens, G. Early plant embryogenesis—Dark ages or dark matter? Curr. Opin. Plant Biol. 2017, 35, 30–36. [Google Scholar] [CrossRef]
- Capron, A.; Chatfield, S.; Provart, N.; Berleth, T. Embryogenesis: Pattern formation from a single cell. In The Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2009. [Google Scholar]
- Zhao, P.; Shi, D.Q.; Yang, W.C. Patterning the embryo in higher plants: Emerging pathways and challenges. Front. Biol. China 2011, 6, 3–11. [Google Scholar] [CrossRef]
- Zaban, B.; Liu, W.; Jiang, X.; Nick, P. Plant cells use auxin efflux to explore geometry. Sci. Rep. 2014, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jürgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Möller, B.; Weijers, D. Auxin control of embryo patterning. Cold Spring Harb. Perspect. Biol. 2009, 1, a001545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.; Bayer, M.; Lukowitz, W. Taking the very first steps: From polarity to axial domains in the early Arabidopsis embryo. J. Exp. Bot. 2011, 62, 1687–1697. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Dhonukshe, P.; Brewer, P.B.; Friml, J. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell. Mol. Life Sci. 2006, 63, 2738–2754. [Google Scholar] [CrossRef]
- Spitzer, C.; Reyes, F.C.; Buono, R.; Sliwinski, M.K.; Haas, T.J.; Otegui, M.S. The ESCRT-Related CHMP1A and B proteins mediate multivesicular body sorting of auxin carriers in Arabidopsis and are required for plant development. Plant Cell 2009, 21, 749–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimata, Y.; Higaki, T.; Kawashima, T.; Kurihara, D.; Sato, Y.; Yamada, T.; Hasezawa, S.; Berger, F.; Higashiyama, T.; Ueda, M. Cytoskeleton dynamics control the first asymmetric cell division in Arabidopsis zygote. Proc. Natl. Acad. Sci. USA 2016, 113, 14157–14162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunda, P.; Baum, B. The actin cytoskeleton in spindle assembly and positioning. Trends Cell Biol. 2009, 19, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Wendrich, J.R.; Weijers, D. The Arabidopsis embryo as a miniature morphogenesis model. New Phytol. 2013, 199, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Laux, T. The asymmetric division of the Arabidopsis zygote: From cell polarity to an embryo axis. Sex. Plant Reprod. 2011, 24, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Begcy, K.; Dresselhaus, T.; Sun, M.-X. Does early embryogenesis in eudicots and monocots involve the same mechanism and molecular players? Plant Physiol. 2017, 173, 130–142. [Google Scholar] [CrossRef] [Green Version]
- Sablowski, R. Coordination of plant cell growth and division: Collective control or mutual agreement? Curr. Opin. Plant Biol. 2016, 34, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Besson, S.; Dumais, J. Universal rule for the symmetric division of plant cells. Proc. Natl. Acad. Sci. USA 2011, 108, 6294–6299. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, B.E.; Tobin, C.; Mjolsness, E.; Meyerowitz, E.M. Analysis of cell division patterns in the Arabidopsis shoot apical meristem. Proc. Natl. Acad. Sci. USA 2015, 112, 4815–4820. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; BarbierdeReuille, P.; Lane, B.; Bassel, G.W.; Prusinkiewicz, P.; Smith, R.S.; Weijers, D. Genetic control of plant development by overriding a geometric division rule. Dev. Cell 2014, 29, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Han, L.; Feng, Z.; Wang, G.; Liu, W.; Ma, Y.; Yu, Y.; Kong, Z. Orchestration of microtubules and the actin cytoskeleton in trichome cell shape determination by a plant-unique kinesin. Elife 2015, 4, e09351. [Google Scholar] [CrossRef] [PubMed]
- Sambade, A.; Findlay, K.; Schäffner, A.R.; Lloyd, C.W.; Buschmann, H. Actin-dependent and -independent functions of cortical microtubules in the differentiation of Arabidopsis leaf trichomes. Plant Cell 2014, 26, 1629–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blancaflor, E.B. Cortical actin filaments potentially interact with cortical microtubules in regulating polarity of cell expansion in primary roots of maize (Zea mays L.). J. Plant Growth Regul. 2000, 19, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Louveaux, M.; Julien, J.D.; Mirabet, V.; Boudaoud, A.; Hamant, O. Cell division plane orientation based on tensile stress in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, E4294–E4303. [Google Scholar] [CrossRef] [Green Version]
- Verger, S.; Long, Y.; Boudaoud, A.; Hamant, O. A tension-adhesion feedback loop in plant epidermis. Elife 2018, 7, e34460. [Google Scholar] [CrossRef] [PubMed]
- Sampathkumar, A.; Krupinski, P.; Wightman, R.; Milani, P.; Berquand, A.; Boudaoud, A.; Hamant, O.; Jönsson, H.; Meyerowitz, E.M. Subcellular and supracellular mechanical stress prescribes cytoskeleton behavior in Arabidopsis cotyledon pavement cells. Elife 2014, 3, e01967. [Google Scholar] [CrossRef]
- Boutté, Y.; Crosnier, M.T.; Carraro, N.; Traas, J.; Satiat-Jeunemaitre, B. The plasma membrane recycling pathway and cell polarity in plants: Studies on PIN proteins. J. Cell Sci. 2006, 119, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Heisler, M.G.; Hamant, O.; Krupinski, P.; Uyttewaal, M.; Ohno, C.; Jönsson, H.; Traas, J.; Meyerowitz, E.M. Alignment between PIN1 polarity and microtubule orientation in the shoot apical meristem reveals a tight coupling between morphogenesis and auxin transport. PLoS Biol. 2010, 8, e1000516. [Google Scholar] [CrossRef] [Green Version]
- Landrein, B.; Hamant, O. How mechanical stress controls microtubule behavior and morphogenesis in plants: History, experiments and revisited theories. Plant J. 2013, 75, 324–338. [Google Scholar] [CrossRef]
- Asnacios, A.; Hamant, O. The mechanics behind cell polarity. Trends Cell Biol. 2012, 22, 584–591. [Google Scholar] [CrossRef]
- Smith, L.G. Cytoskeletal control of plant cell shape: Getting the fine points. Curr. Opin. Plant Biol. 2003, 6, 63–73. [Google Scholar] [CrossRef]
- Peremyslov, V.V.; Prokhnevsky, A.I.; Dolja, V.V. Class XI myosins are required for development, cell expansion, and F-actin organization in Arabidopsis. Plant Cell 2010, 22, 1883–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovaleva, L.V.; Voronkov, A.S.; Zakharova, E.V. Role of auxin and cytokinin in the regulation of the actin cytoskeleton in the in vitro germinating male gametophyte of petunia. Russ. J. Plant Physiol. 2015, 62, 179–186. [Google Scholar] [CrossRef]
- Lee, Y.J.; Szumlanski, A.; Nielsen, E.; Yang, Z. Rho-GTPase-dependent filamentous actin dynamics coordinate vesicle targeting and exocytosis during tip growth. J. Cell Biol. 2008, 181, 1155–1168. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Zhang, R.; Zhang, M.; Diao, M.; Xue, Y.; Huang, S. Organizational innovation of apical actin filaments drives rapid pollen tube growth and turning. Mol. Plant 2017, 10, 930–947. [Google Scholar] [CrossRef] [Green Version]
- Eitzen, G. Actin remodeling to facilitate membrane fusion. Biochim. Biophys. Acta Mol. Cell Res. 2003, 1641, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Bademosi, A.T.; Luo, J.; Meunier, F.A. Actin remodeling in regulated exocytosis: Toward a mesoscopic view. Trends Cell Biol. 2018, 28, 685–697. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, F.; Vogel, S.K.; Schwille, P. Lateral membrane diffusion modulated by a minimal actin cortex. Biophys. J. 2013, 104, 1465–1475. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Shi, H.; Chen, B.; Zhang, R.; Huang, S.; Fu, Y. Arabidopsis RIC1 severs actin filaments at the apex to regulate pollen tube growth. Plant Cell 2015, 27, 1140–1161. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, M.; Staehelin, L.A.; Mineyuki, Y. Actin-Microtubule Interaction in Plants, Cytoskeleton–Structure, Dynamics, Function and Disease; Jimenez-Lopez, J.C., Ed.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Sampathkumar, A.; Lindeboom, J.J.; Debolt, S.; Gutierrez, R.; Ehrhardt, D.W.; Ketelaar, T.; Persson, S. Live cell imaging reveals structural associations between the actin and microtubule cytoskeleton in arabidopsis. Plant Cell 2011, 23, 2302–2313. [Google Scholar] [CrossRef] [Green Version]
- Chakrabortty, B.; Blilou, I.; Scheres, B.; Mulder, B.M. A computational framework for cortical microtubule dynamics in realistically shaped plant cells. PLoS Comput. Biol. 2018, 14, e1005959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabortty, B.; Willemsen, V.; de Zeeuw, T.; Liao, C.Y.; Weijers, D.; Mulder, B.; Scheres, B. A plausible microtubule-based mechanism for cell division orientation in plant embryogenesis. Curr. Biol. 2018, 28, 3031–3043.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun, L.; Rupp, B.; Ward, J.J.; Nédélec, F. A theory of microtubule catastrophes and their regulation. Proc. Natl. Acad. Sci. USA 2009, 106, 21173–21178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, A.; Shaw, S.L. Update: Plant cortical microtubule arrays. Plant Physiol. 2018, 176, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Vineyard, L.; Elliott, A.; Dhingra, S.; Lucas, J.R.; Shaw, S.L. Progressive transverse microtubule array organization in hormone-induced Arabidopsis hypocotyl cells. Plant Cell 2013, 25, 662–676. [Google Scholar] [CrossRef] [Green Version]
- Ivanchenko, M.G.; Napsucialy-Mendivil, S.; Dubrovsky, J.G. Auxin-induced inhibition of lateral root initiation contributes to root system shaping in Arabidopsis thaliana. Plant J. 2010, 64, 740–752. [Google Scholar] [CrossRef]
- Chen, X.; Grandont, L.; Li, H.; Hauschild, R.; Paque, S.; Abuzeineh, A.; Rakusová, H.; Benkova, E.; Perrot-Rechenmann, C.; Friml, J. Inhibition of cell expansion by rapid ABP1-mediated auxin effect on microtubules. Nature 2014, 516, 90–93. [Google Scholar] [CrossRef]
- Oda, Y. Cortical microtubule rearrangements and cell wall patterning. Front. Plant Sci. 2015, 6, 236. [Google Scholar] [CrossRef] [Green Version]
- Hamann, T.; Mayer, U.; Jürgens, G. The auxin-insensitive bodenlos mutation affects primary root formation and apical-basal patterning in the Arabidopsis embryo. Development 1999, 126, 1387–1395. [Google Scholar]
- Hamann, T.; Benkova, E.; Bäurle, I.; Kientz, M.; Jürgens, G. The Arabidopsis BODENLOS gene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning. Genes Dev. 2002, 16, 1610–1615. [Google Scholar] [CrossRef] [Green Version]
- Sassi, M.; Traas, J. New insights in shoot apical meristem morphogenesis: Isotropy comes into play. Plant Signal. Behav. 2015, 10, e1000150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassi, M.; Ali, O.; Boudon, F.; Cloarec, G.; Abad, U.; Cellier, C.; Chen, X.; Gilles, B.; Milani, P.; Friml, J.; et al. An auxin-mediated shift toward growth isotropy promotes organ formation at the shoot meristem in arabidopsis. Curr. Biol. 2014, 24, 2335–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wabnik, K.; Robert, H.S.; Smith, R.S.; Friml, J. Modeling framework for the establishment of the apical-basal embryonic axis in plants. Curr. Biol. 2013, 23, 2513–2518. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.Y.; Shi, D.Q.; Jia, P.F.; Tang, J.; Li, H.J.; Liu, J.; Yang, W.C. The arabidopsis receptor kinase ZAR1 is required for zygote asymmetric division and its daughter cell fate. PLoS Genet. 2016, 12, e1005933. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Guo, Z.; Song, L.; Wang, Y.; Cheng, Y. NCP1/AtMOB1A plays key roles in auxin-mediated arabidopsis development. PLoS Genet. 2016, 12, e1005923. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Shi, C.; Zhao, P.; Sun, M. Isolation of living apical and basal cell lineages of early proembryos for transcriptome analysis. Plant Reprod. 2019, 32, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Babu, Y.; Musielak, T.; Henschen, A.; Bayer, M. Suspensor length determines developmental progression of the embryo in Arabidopsis. Plant Physiol. 2013, 162, 1448–1458. [Google Scholar] [CrossRef] [Green Version]
- Jenik, P.D.; Barton, M.K. Surge and destroy: The role of auxin in plant embryogenesis. Development 2005, 132, 3577–3585. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, X.; Zhao, J.; Tang, X.; Tian, S.; Chen, J.; Shi, C.; Wang, W.; Zhang, L.; Feng, X.; et al. Direct evidence that suspensor cells have embryogenic potential that is suppressed by the embryo proper during normal embryogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, 12432–12437. [Google Scholar] [CrossRef] [Green Version]
- Gooh, K.; Ueda, M.; Aruga, K.; Park, J.; Arata, H.; Higashiyama, T.; Kurihara, D. Live-cell imaging and optical manipulation of Arabidopsis early embryogenesis. Dev. Cell 2015, 34, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Nick, P.; Han, M.J.; An, G. Auxin Stimulates its own transport by shaping actin filaments. Plant Physiol. 2009, 151, 155–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nick, P. Probing the actin-auxin oscillator. Plant Signal. Behav. 2010, 5, 94–98. [Google Scholar] [CrossRef]
- Waller, F.; Riemann, M.; Nick, P. A role for actin-driven secretion in auxin-induced growth. Protoplasma 2002, 219, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Geisler, M. Keeping it all together: Auxin-actin crosstalk in plant development. J. Exp. Bot. 2015, 66, 4983–4998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.W.; Chen, X.; Mei, Y. Function and regulation of phospholipid signalling in plants. Biochem. J. 2009, 421, 145–156. [Google Scholar] [CrossRef]
- Qu, L.H.; Zhou, X.; Li, X.; Li, S.S.; Zhao, J.; Zhao, P.; Liu, Y.; Sun, M.X. The autonomous cell fate specification of basal cell lineage: The initial round of cell fate specification occurs at the two-celled proembryo stage. Plant J. 2017, 91, 1051–1063. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Guo, Y.; Zhao, P.; Sun, M.X. Comparative analysis of WUSCHEL-related homeobox genes revealed their parent-of-origin and cell type-specific expression pattern during early embryogenesis in tobacco. Front. Plant Sci. 2018, 9, 311. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Sun, M.X. NtDRP is necessary for accurate zygotic division orientation and differentiation of basal cell lineage toward suspensor formation. New Phytol. 2016, 212, 598–612. [Google Scholar] [CrossRef] [Green Version]
- Lindeboom, J.J.; Nakamura, M.; Saltini, M.; Hibbel, A.; Walia, A.; Ketelaar, T.; Emons, A.M.C.; Sedbrook, J.C.; Kirik, V.; Mulder, B.M.; et al. CLASP stabilization of plus ends created by severing promotes microtubule creation and reorientation. J. Cell Biol. 2019, 218, 190–205. [Google Scholar] [CrossRef]
- Ambrose, J.C.; Shoji, T.; Kotzer, A.M.; Pighin, J.A.; Wasteneys, G.O. The Arabidopsis CLASP gene encodes a microtubule-associated protein involved in cell expansion and division. Plant Cell 2007, 19, 2763–2775. [Google Scholar] [CrossRef] [Green Version]
- Luptovčiak, I.; Komis, G.; Takáč, T.; Ovečka, M.; Šamaj, J. Katanin: A sword cutting microtubules for cellular, developmental, and physiological purposes. Front. Plant Sci. 2017, 8, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burk, D.H.; Ye, Z.H. Alteration of oriented deposition of cellulose microfibrils by mutation of a katanin-like microtubule-severing protein. Plant Cell 2002, 14, 2145–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luptovciak, I.; Samakovli, D.; Komis, G.; Šamaj, J. KATANIN 1 is essential for embryogenesis and seed formation in Arabidopsis. Front. Plant Sci. 2017, 8, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Dong, J. Focusing on the focus: What else beyond the master switches for polar cell growth? Mol. Plant 2015, 8, 582–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takáč, T.; Šamajová, O.; Pechan, T.; Luptovčiak, I.; Šamaj, J. Feedback microtubule control and microtubule-actin cross-talk in arabidopsis revealed by integrative proteomic and cell biology analysis of KATANIN 1 mutants. Mol. Cell. Proteomics 2017, 16, 1591–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanneste, S.; Friml, J. Calcium: The missing link in auxin action. Plants 2013, 2, 650–675. [Google Scholar] [CrossRef] [Green Version]
- Iwaya, N.; Akiyama, K.; Goda, N.; Tenno, T.; Fujiwara, Y.; Hamada, D.; Ikura, T.; Shirakawa, M.; Hiroaki, H. Effect of Ca 2+ on the microtubule-severing enzyme p60-katanin. Insight into the substrate-dependent activation mechanism. FEBS J. 2012, 279, 1339–1352. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Qin, T.; Zhang, Y.; Liu, X.; Sun, J.; Zhou, Y.; Zhu, L.; Zhang, Z.; Yuan, M.; et al. MDP25, a novel calcium regulatory protein, mediates hypocotyl cell elongation by destabilizing cortical microtubules in Arabidopsis. Plant Cell 2011, 23, 4411–4427. [Google Scholar] [CrossRef] [Green Version]
- Qin, T.; Li, J.; Yuan, M.; Mao, T. Characterization of the role of calcium in regulating the microtubule-destabilizing activity of MDP25. Plant Signal. Behav. 2012, 7, 708–710. [Google Scholar] [CrossRef] [Green Version]
- Scherer, G.F.E. AUXIN-BINDING-PROTEIN1, the second auxin receptor: What is the significance of a two-receptor concept in plant signal transduction? J. Exp. Bot. 2011, 62, 3339–3357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhang, W. Regulation of developmental and environmental signaling by interaction between microtubules and membranes in plant cells. Protein Cell 2016, 7, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamant, O.; Heisler, M.G.; Jönsson, H.; Krupinski, P.; Uyttewaal, M.; Bokov, P.; Corson, F.; Sahlin, P.; Boudaoud, A.; Meyerowitz, E.M.; et al. Developmental patterning by mechanical signals in Arabidopsis. Science 2008, 322, 1650–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarbreck, S.M.; Colaço, R.; Davies, J.M. Plant calcium-permeable channel. Plant Physiol. 2013, 163, 514–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma membrane H+-ATPase regulation in the center of plant physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [CrossRef] [Green Version]
- Dindas, J.; Scherzer, S.; Roelfsema, M.R.G.; Von Meyer, K.; Müller, H.M.; Al-Rasheid, K.A.S.; Palme, K.; Dietrich, P.; Becker, D.; Bennett, M.J.; et al. AUX1-mediated root hair auxin influx governs SCFTIR1/AFB-type Ca2+ signaling. Nat. Commun. 2018, 9, 1174. [Google Scholar] [CrossRef]
- Ephritikhine, G.; Barbier-Brygoo, H.; Muller, J.-F.; Guern, J. Auxin effect on the transmembrane potential difference of wild-type and mutant tobacco protoplasts exhibiting a differential sensitiity to auxin. Plant Physiol. 1987, 83, 801–804. [Google Scholar] [CrossRef]
- Felle, H.; Peters, W.; Palme, K. The electrical response of maize to auxins. BBA Biomembr. 1991, 1064, 199–204. [Google Scholar] [CrossRef]
- Krebs, M.; Waadt, R.; Wang, Y.; Dindas, J.; Rienmu, F.; Schumacher, K.; Wu, W.; Hedrich, R.; Roelfsema, M.R.G. Cytosolic Ca 2+ signals enhance the vacuolar ion conductivity of bulging Arabidopsis root hair cells. Mol. Plant 2015, 8, 1665–1674. [Google Scholar]
- Schönknecht, G. Calcium signals from the vacuole. Plants 2013, 2, 589–614. [Google Scholar] [CrossRef] [Green Version]
- Kopka, J.; Pical, C.; Gray, J.E.; Müller-Röber, B. Molecular and enzymatic characterization of three phosphoinositide-specific phospholipase C isoforms from potato. Plant Physiol. 1998, 116, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Gonzales, R.A.; Bhattacharyya, M.K. Characterization of a plasma membrane-associated phosphoinositide-specific phospholipase C from soybean. Plant J. 1995, 8, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling: An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinke, O.; Novotná, Z.; Valentová, O.; Martinec, J. Inositol trisphosphate receptor in higher plants: Is it real? J. Exp. Bot. 2007, 58, 361–376. [Google Scholar] [CrossRef] [Green Version]
- Kimata, Y.; Kato, T.; Higaki, T.; Kurihara, D.; Yamada, T.; Segami, S.; Morita, M.T.; Maeshima, M.; Hasezawa, S.; Higashiyama, T.; et al. Polar vacuolar distribution is essential for accurate asymmetric division of Arabidopsis zygotes. Proc. Natl. Acad. Sci. USA 2019, 116, 2338–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.Y.; Weijers, D. A toolkit for studying cellular reorganization during early embryogenesis in Arabidopsis thaliana. Plant J. 2018, 93, 963–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottosin, I.I.; Schönknecht, G. Vacuolar calcium channels. J. Exp. Bot. 2007, 58, 1559–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethke, P.C.; Jones, R.L. Ca2+-calmodulin modulates lon channel activity in storage protein vacuoles of barley aleurone cells. Plant Cell 1994, 6, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Anil, V.S.; Rao, K.S. Calcium-mediated signaling during sandalwood somatic embryogenesis. Role for exogenous calcium as second messenger. Plant Physiol. 2000, 123, 1301–1311. [Google Scholar] [CrossRef] [Green Version]
- Mahalakshmi, A.; Singla, B.; Khurana, J.P.; Khurana, P. Role of calcium-calmodulin in auxin-induced somatic embryogenesis in leaf base cultures of wheat (Triticum aestivum var. HD 2329). Plant Cell Tissue Organ Cult. 2007, 88, 167–174. [Google Scholar] [CrossRef]
- Zeng, F.; Zhang, X.; Cheng, L.; Hu, L.; Zhu, L.; Cao, J.; Guo, X. A draft gene regulatory network for cellular totipotency reprogramming during plant somatic embryogenesis. Genomics 2007, 90, 620–628. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Nishimura, M.; Shimazaki, K.I. Cytosolic concentration of Ca2+ regulates the plasma membrane H+-ATPase in guard cells of fava bean. Plant Cell 1995, 7, 1333–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Nisi, P.; Dell’Orto, M.; Pirovano, L.; Zocchi, G. Calcium-dependent phosphorylation regulates the plasma-membrane H+-ATPase activity of maize (Zea mays L.) roots. Planta 1999, 209, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Brault, M.; Amiar, Z.; Pennarun, A.-M.; Monestiez, M.; Zhang, Z.; Cornel, D.; Dellis, O.; Knight, H.; Bouteau, F.; Rona, J.-P. Plasma membrane depolarization induced by abscisic acid in Arabidopsis suspension cells involves reduction of proton pumping in addition to anion channel activation, which are both Ca21 dependent. Plant Physiol. 2004, 135, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz-Lessdorf, B.; Hedrich, R. Protons and calcium modulate SV-type channels in the vacuolar-lysosomal compartment-channel interaction with calmodulin inhibitors. Planta 1995, 197, 655–671. [Google Scholar] [CrossRef]
- Balzan, S.; Johal, G.S.; Carraro, N. The role of auxin transporters in monocots development. Front. Plant Sci. 2014, 5, 393. [Google Scholar] [CrossRef] [Green Version]
- Lucas, J.R.; Nadeau, J.A.; Sack, F.D. Microtubule arrays and Arabidopsis stomatal development. J. Exp. Bot. 2006, 57, 71–79. [Google Scholar] [CrossRef]
- Le, J.; Liu, X.G.; Yang, K.Z.; Chen, X.L.; Zou, J.J.; Wang, H.Z.; Wang, M.; Vanneste, S.; Morita, M.; Tasaka, M.; et al. Auxin transport and activity regulate stomatal patterning and development. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, E.; Belcram, K.; Uyttewaal, M.; Duroc, Y.; Goussot, M.; Legland, D.; Laruelle, E.; De Tauzia-Moreau, M.L.; Pastuglia, M.; Bouchez, D. The preprophase band of microtubules controls the robustness of division orientation in plants. Science 2017, 356, 186–189. [Google Scholar] [CrossRef]
- Livanos, P.; Giannoutsou, E.; Apostolakos, P.; Galatis, B. Auxin as an inducer of asymmetrical division generating the subsidiary cells in stomatal complexes of Zea mays. Plant Signal. Behav. 2015, 10, e984531. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Tucker, E.; Hermann, M.; Laux, T. A molecular framework for the embryonic initiation of shoot meristem stem cells. Dev. Cell 2017, 40, 264–277.e4. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; Park, J.W.; Lee, H.W.; Kim, J. Genome-wide analysis of the auxin-responsive transcriptome downstream of iaa1 and its expression analysis reveal the diversity and complexity of auxin-regulated gene expression. J. Exp. Bot. 2009, 60, 3935–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paponov, I.A.; Paponov, M.; Teale, W.; Menges, M.; Chakrabortee, S. Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol. Plant 2008, 1, 321–337. [Google Scholar] [CrossRef]
- Habets, M.E.J.; Offringa, R. PIN-driven polar auxin transport in plant developmental plasticity: A key target for environmental and endogenous signals. New Phytol. 2014, 203, 362–377. [Google Scholar] [CrossRef]
- He, P.; Zhao, P.; Wang, L.; Zhang, Y.; Wang, X.; Xiao, H.; Yu, J.; Xiao, G. The PIN gene family in cotton (Gossypium hirsutum): Genome-wide identification and gene expression analyses during root development and abiotic stress responses. BMC Genom. 2017, 18, 507. [Google Scholar] [CrossRef] [Green Version]
- Smith, Z.R.; Long, J.A. Control of Arabidopsis apical-basal embryo polarity by antagonistic transcription factors. Nature 2010, 464, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Rosspopoff, O.; Chelysheva, L.; Saffar, J.; Lecorgne, L.; Gey, D.; Caillieux, E.; Colot, V.; Roudier, F.; Hilson, P.; Berthomé, R.; et al. Direct conversion of root primordium into shoot meristem relies on timing of stem cell niche development. Development 2017, 144, 1187–1200. [Google Scholar] [CrossRef] [Green Version]
- Peres, L.E.P.; Kerbauy, G.B. High cytokinin accumulation following root tip excision changes the endogenous auxin-to-cytokinin ratio during root-to-shoot conversion in Catasetum fimbriatum Lindl (Orchidaceae). Plant Cell Rep. 1999, 18, 1002–1006. [Google Scholar] [CrossRef]
- Kareem, A.; Radhakrishnan, D.; Wang, X.; Bagavathiappan, S.; Trivedi, Z.B.; Sugimoto, K.; Xu, J.; Mähönen, A.P.; Prasad, K. Protocol: A method to study the direct reprogramming of lateral root primordia to fertile shoots. Plant Methods 2016, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Bigeard, J.; Hirt, H. Nuclear signaling of plant MAPKs. Front. Plant Sci. 2018, 9, 469. [Google Scholar] [CrossRef] [Green Version]
- Dóczi, R.; Bögre, L. The quest for MAP kinase substrates: Gaining momentum. Trends Plant Sci. 2018, 23, 918–932. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Mohanta, N.; Parida, P.; Bae, H. Mitogen Activated Protein Kinase (MPK) interacts with auxin influx carrier (OsAux/LAX1) involved in auxin signaling in plant. Biol. Proced. Online 2015, 17, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnicki, K.; Żabka, A.; Polit, J.T.; Maszewski, J. Mitogen-activated protein kinases concentrate in the vicinity of chromosomes and may regulate directly cellular patterning in Vicia faba embryos. Planta 2018, 248, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Chen, Y.; Luo, L.; Peck, S.C. Central roles and regulatory mechanisms of dual-specificity MAPK phosphatases in developmental and stress signaling. Front. Plant Sci. 2018, 871, 1697. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.; Besteiro, M.A.G.; Lang, D.; Ulm, R. Emerging functions for plant MAP kinase phosphatases. Trends Plant Sci. 2010, 15, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.D.; Callaham, D.A.; Gross, D.J.; Hepler, P.K. Free Ca2+ gradient in growing pollen tubes of lilium. J. Cell Sci. 1992, 101, 7–12. [Google Scholar]
- Jaffe, L.A.; Weisenseel, M.H.; Jaffe, L.F. Calcium accumulations within the growing tips of pollen tubes. J. Antibiot. (Tokyo) 1975, 67, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Zabka, A.; Polit, J.T.; Winnicki, K.; Paciorek, P.; Juszczak, J.; Nowak, M.; Maszewski, J. PIN2-like proteins may contribute to the regulation of morphogenetic processes during spermatogenesis in Chara vulgaris. Plant Cell Rep. 2016, 35, 1655–1669. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Sun, H.L.; Mei, C.; Wang, X.J.; Yan, L.; Liu, R.; Zhang, X.F.; Wang, X.F.; Zhang, D.P. The Arabidopsis Ca 2+-dependent protein kinase CPK12 negatively regulates abscisic acid signaling in seed germination and post-germination growth. New Phytol. 2011, 192, 61–73. [Google Scholar] [CrossRef]
- Inoue, S.I.; Takahashi, K.; Okumura-Noda, H.; Kinoshita, T. Auxin influx carrier AUX1 confers acid resistance for arabidopsis root elongation through the regulation of plasma membrane H+-ATPase. Plant Cell Physiol. 2016, 57, 2194–2201. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, P.; Du, H.; Sheen, J.; Assmann, S.M.; Albert, R. Model-driven discovery of calcium-related protein-phosphatase inhibition in plant guard cell signaling. PLoS Comput. Biol. 2019, 15, e1007429. [Google Scholar] [CrossRef]
- Rodriguez, P.L. Protein phosphatase 2C (PP2C) function in higher plants. Plant Mol. Biol. 1998, 38, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Phengchat, R.; Takata, H.; Morii, K.; Inada, N.; Murakoshi, H.; Uchiyama, S.; Fukui, K. Calcium ions function as a booster of chromosome condensation. Sci. Rep. 2016, 6, 38281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, L.F.; Robinson, K.R.; Nuccitelli, R. Local cation entry and self-electrophoresis as an intracellular localization mechanism. Ann. N. Y. Acad. Sci. 1974, 238, 372–389. [Google Scholar] [CrossRef] [PubMed]
- Raccaud, M.; Friman, E.T.; Alber, A.B.; Agarwal, H.; Deluz, C.; Kuhn, T.; Gebhardt, J.C.M.; Suter, D.M. Mitotic chromosome binding predicts transcription factor properties in interphase. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raccaud, M.; Suter, D.M. Transcription factor retention on mitotic chromosomes: Regulatory mechanisms and impact on cell fate decisions. FEBS Lett. 2018, 592, 878–887. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The role of calcium ions in the regulation of the direction of cell division during embryogenesis. (A) A high concentration of calcium ions negatively regulates functioning of katanin and activate destabilizing properties of MDP25, finally supporting longitudinal alignment of microtubules. (B) Low concentration of calcium ions has limited impact on katanin or MDP25. In this situation katanin activity is maintained and function of MDP25 is stopped, which results in transverse alignment of microtubules.
Figure 1.
The role of calcium ions in the regulation of the direction of cell division during embryogenesis. (A) A high concentration of calcium ions negatively regulates functioning of katanin and activate destabilizing properties of MDP25, finally supporting longitudinal alignment of microtubules. (B) Low concentration of calcium ions has limited impact on katanin or MDP25. In this situation katanin activity is maintained and function of MDP25 is stopped, which results in transverse alignment of microtubules.

Figure 2.
A hypothetical model of calcium ion release. (A) Transport of auxin ions to the apical cell is provided by polar localization of PIN7 proteins. The auxin influx induces plasma membrane depolarization, followed by an opening of calcium channels. Auxin and calcium ions regulate the activity of phospholipase C (PLC) which catalyzes conversion of PIP2 to IP3. IP3 induces opening ligand-gated calcium channels by binding to specific receptors in ER or vacuole. An elevated concentration of calcium ions activates also voltage-dependent channels in a vacuole. On the other hand, high concentration of calcium ions triggers inhibition of plasma membrane H+-ATPase which results in apoplast alkalization. High pH of an apoplast reduces auxin-induced plasma membrane depolarization and may act as self-attenuating mechanism of auxin impact. (B) After two longitudinal divisions the cell volume is 4-fold smaller but the surface of vertical membranes of daughter cells is only halved. All this results in faster achievement of high calcium concentration and following inhibition of plasma membrane H+-ATPase. Apoplast alkalization reduces auxin-induced depolarization of plasma membrane. In turn, cytosol acidification results in auxin protonation which allows for its diffusion from cytoplasm. Thus, both auxin concentration and calcium ion release from ER and vacuole are diminished.
Figure 2.
A hypothetical model of calcium ion release. (A) Transport of auxin ions to the apical cell is provided by polar localization of PIN7 proteins. The auxin influx induces plasma membrane depolarization, followed by an opening of calcium channels. Auxin and calcium ions regulate the activity of phospholipase C (PLC) which catalyzes conversion of PIP2 to IP3. IP3 induces opening ligand-gated calcium channels by binding to specific receptors in ER or vacuole. An elevated concentration of calcium ions activates also voltage-dependent channels in a vacuole. On the other hand, high concentration of calcium ions triggers inhibition of plasma membrane H+-ATPase which results in apoplast alkalization. High pH of an apoplast reduces auxin-induced plasma membrane depolarization and may act as self-attenuating mechanism of auxin impact. (B) After two longitudinal divisions the cell volume is 4-fold smaller but the surface of vertical membranes of daughter cells is only halved. All this results in faster achievement of high calcium concentration and following inhibition of plasma membrane H+-ATPase. Apoplast alkalization reduces auxin-induced depolarization of plasma membrane. In turn, cytosol acidification results in auxin protonation which allows for its diffusion from cytoplasm. Thus, both auxin concentration and calcium ion release from ER and vacuole are diminished.

Figure 3.
A hypothetical model of mitogen-activated protein kinase (MAPK)-dependent cellular patterning during embryogenesis. (A) The asymmetrical distribution of auxin, calcium ions and phosphorylated MAPKs in the vicinity of chromosomes. (B) High concentration of calcium ions at one end activates dual-specificity phosphatases (DUSPs) which dephosphorylate MAPKs and diminish the number of activated kinases in the vicinity of chromosomes. However, low calcium level at the opposite end might result in reduced activity of phosphatases, and therefore the number of chromatids with activated MAPKs might be greater at this end. Interestingly calcium ions activate CDPKs (calcium-dependent protein kinases) which may positively regulate PP2C phosphatase, however high calcium concentration was found to directly inhibit the activity of PP2C phosphatase.
Figure 3.
A hypothetical model of mitogen-activated protein kinase (MAPK)-dependent cellular patterning during embryogenesis. (A) The asymmetrical distribution of auxin, calcium ions and phosphorylated MAPKs in the vicinity of chromosomes. (B) High concentration of calcium ions at one end activates dual-specificity phosphatases (DUSPs) which dephosphorylate MAPKs and diminish the number of activated kinases in the vicinity of chromosomes. However, low calcium level at the opposite end might result in reduced activity of phosphatases, and therefore the number of chromatids with activated MAPKs might be greater at this end. Interestingly calcium ions activate CDPKs (calcium-dependent protein kinases) which may positively regulate PP2C phosphatase, however high calcium concentration was found to directly inhibit the activity of PP2C phosphatase.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Winnicki, K. The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis. Cells 2020, 9, 606. https://doi.org/10.3390/cells9030606
AMA Style
Winnicki K. The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis. Cells. 2020; 9(3):606. https://doi.org/10.3390/cells9030606
Chicago/Turabian StyleWinnicki, Konrad. 2020. "The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis" Cells 9, no. 3: 606. https://doi.org/10.3390/cells9030606
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.