Actin Cytoskeleton Straddling the Immunological Synapse between Cytotoxic Lymphocytes and Cancer Cells


1
Cytoskeleton and Cancer Progression; Laboratory of Experimental Cancer Research, Department of Oncology 84 Val Fleuri, L-1526 Luxembourg City, Luxembourg
2
University of Luxembourg, Faculty of Science, Technology and Communication, 2 Avenue de l’Université, L-4365 Esch-sur-Alzette, Luxembourg
3
University of Strasbourg, 67081 Strasbourg, France
*
Author to whom correspondence should be addressed.
Cells 2019, 8(5), 463; https://doi.org/10.3390/cells8050463
Submission received: 15 April 2019
/
Revised: 8 May 2019
/
Accepted: 13 May 2019
/
Published: 16 May 2019
(This article belongs to the Special Issue Tumor Microenvironment: Interaction and Metabolism)
{kind=link}
Abstract
:The immune system is a fundamental part of the tumor microenvironment. In particular, cytotoxic lymphocytes, such as cytolytic T cells and natural killer cells, control tumor growth and disease progression by interacting and eliminating tumor cells. The actin cytoskeleton of cytotoxic lymphocytes engaged in an immunological synapse has received considerable research attention. It has been recognized as a central mediator of the formation and maturation of the immunological synapse, and its signaling and cytolytic activities. In comparison, fewer studies have explored the organization and function of actin filaments on the target cancer cell side of the immunological synapse. However, there is growing evidence that the actin cytoskeleton of cancer cells also undergoes extensive remodeling upon cytotoxic lymphocyte attack, and that such remodeling can alter physical and functional interactions at the immunological synapse. In this article, we review the current knowledge of actin organization and functions at both sides of the immunological synapse between cytotoxic lymphocytes and cancer cells, with particular focus on synapse formation, signaling and cytolytic activity, and immune evasion.
1. Introduction
The tumor microenvironment (TME) plays multiple and central roles in cancer progression, e.g., by promoting tumor invasion, chemo and radiation-resistance, and by modulating the antitumor immune response. Ongoing research in the field of tumor immunobiology has identified immune escape as a classical hallmark of cancer and characterized different escape strategies elaborated by cancer cells [1,2,3]. The immune cell population with the best studied anti-tumor effector functions are cytotoxic lymphocytes cells, including cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells. The concept of immunoediting is highly dependent on these key cells of the adaptive and innate immune systems. That the immune system and cancer cells exist in a delicate balance was already described more than a hundred years ago by Paul Ehrlich, when he suggested that host defense may prevent neoplastic cells from developing into tumors [4]. Sir Frank Mac Farlane Burnet hypothesized later that tumor cell neo-antigens induce an immunological reaction against cancer and subsequently formulated the immune surveillance theory [5]. The elimination phase of the immunoediting process is characterized by successful immunosurveillance of cancer cells that are recognized and eradicated by cytotoxic lymphocytes [6]. Constant immune selection pressure allows tumor clones to emerge, which escape immune cell-mediated elimination in the so-called equilibrium phase. During this phase, tumor cells develop different stratagems to escape immune surveillance, such as altered expression of surface markers, immune cell inhibition and establishment of an immunosuppressive TME [7,8,9,10]. Finally, tumors progress into the third phase of the immunoediting process, the escape phase, which leads to faster disease progression and poorly immunogenic tumors. Tumor cells have the ability to change their surroundings to their benefit, a feature that is accentuated by the characteristics of the TME, such as hypoxia—as a result of poor vascularization and rapid proliferation of cancer cells [11,12,13]. Hypoxia creates a hostile environment for cytotoxic immune cells that hinders their activation and effectiveness, while promoting activity of immune suppressive cell populations [11,14,15,16,17].
The actin cytoskeleton mainly consists in a complex network of polarized actin filaments (AFs) that contributes to nearly all fundamental cellular processes, including morphogenesis, motility, differentiation, division, membrane trafficking and signaling and the reader is invited to read the following review suggestions referring to these fields [18,19,20,21,22,23,24,25]. The actin cytoskeleton is subject to the activity of over 100 actin-binding proteins (ABPs) that regulate the organization and dynamics of AFs [26,27]. Broadly, ABPs can be distinguished according to their functions as actin nucleators, AF severing and capping proteins, and AF crosslinkers [25,26]. Actin polymerization occurs by polymerization of globular actin monomers, a process facilitated by actin nucleators, such as the Arp2/3 complex and formins, which promote the assembly of branched and linear arrays of AFs, respectively [26,28,29,30]. Further organization of AFs into higher-order structures, such as parallel bundles and three-dimensional networks, is mediated by crosslinking proteins of differing structural properties [31]. Severing proteins, such as actin depolymerizing factors (ADFs) and cofilin, play important roles in regulating AF dynamics by either increasing the amount of fast-growing barbed ends available for polymerization or by accelerating depolymerization from AF pointed ends [26,32].
A fundamental process underlying cytotoxic lymphocyte-mediated cancer cell killing is the formation of a specialized cell-cell junction, referred to as the immunological synapse (IS), between the immune cell and its prospective target. Different types of ISs can be formed, postulated to execute several functions including but not limited to the directed secretion of cytokines or lytic granules in a lytic IS with the goal of target cell lysis. The formation of a lytic IS requires activation of CTLs and NK cells through interaction of the T cell receptor (TCR) with peptide-loaded major histocompatibility complex-I (pMHC-I) on the target cell surface, or recognition of increased stress-induced ligands or loss of inhibitory MHC-I on the target cell surface, respectively. Initiation, formation and maturation of a lytic IS between cytotoxic lymphocytes and targets is a delicate process associated with major rearrangements of the actin cytoskeleton in cytotoxic lymphocytes [33,34,35]. In addition, the directed secretion of cytolytic granules containing perforin and granzymes, which ultimately promotes osmotic cell death and caspase-mediated apoptosis, also critically rely on actin cytoskeleton remodeling on the immune cell side of the IS [36,37,38,39,40]. On healthy cells, self-antigens presented on the MHC-I are not recognized by the TCR and the amount of stress-induced ligands is below the threshold required for NK cell activation, resulting in the formation of an inhibitory IS that protects the target cell from lymphocyte-mediated cytotoxicity. Another function executed by the IS formed between CTLs and dendritic cells (DCs) is the regulation of lymphocyte activation and maturation. A similar type of IS has been reported for the cell-cell-contact between NK cells and mature DCs and this interaction was termed regulatory IS [35,41,42,43]. Intercellular communication in a regulatory synapse can induce CTL activation and priming, increase the capacity of NK cells to release cytokines, and promote DC maturation. The different types of ISs are depicted in Figure 1.
Specific changes in actin cytoskeleton organization and dynamics are critically required for the initiation and stabilization of an IS, as well as for the activation and efficacy of associated immune cell effector functions. Such changes are the subject of much ongoing research [35]. Beside the well-defined actin cytoskeleton organization and roles in cytotoxic lymphocytes cells during IS formation and killing activity, there is increasing evidence that the actin cytoskeleton of cancer cells is also quickly remodeled in response to immune cell attack [44]. This review aims to discuss the role of the actin cytoskeleton in both, immune cells and cancer cells, during their interaction in an IS. The formation of an IS follows well-orchestrated spatiotemporal rearrangements of cytoskeletal components, that we will describe in detail. Particular attention will be paid to the organization and functions of the actin cytoskeleton during IS formation, signaling, and activity. In addition, we will discuss recent studies supporting that actin cytoskeleton remodeling in cancer cells can alter IS activity and lead to cancer immune evasion.
2. Actin Cytoskeleton and Immunological Synapse Architecture
2.1. Actin Assemblies at the Immunological Synapse
In CTLs, immune cell activation is triggered by interaction of the T cell receptor (TCR) with a non-self-antigen presented on the major histocompatibility complex class I (MHC-I) and engagement of further costimulatory receptors trigger [46,47,48,49]. In contrast, NK cell activity is regulated by the balance between activating and inhibiting ligands presented on the target cells surface. A vast array of inhibitory and activating receptors are co-expressed on the surface of NK cells, and activation occurs when signals from activating receptors dominate over the counterbalancing signals from inhibitory receptors [50,51]. In both CTLs and NK cells, engagement of activating receptors induces phosphorylation of membrane proximal signaling molecules and formation of a signalosome that includes many signaling and adapter molecules. Sustained signaling stimulates actin polymerization and generation of a branched AF network at the synapse periphery. The architecture of an IS is commonly divided in three regions defined by distinct actin assemblies and with specific functions in relation to signaling, adhesion, and cytolytic activities. The so-called supramolecular activation clusters (SMAC) are organized in a bulls-eye shape with a central SMAC (cSMAC), a peripheral SMAC (pSMAC), and a distal SMAC (dSMAC) from the inside out [35,52,53]. Live imaging of AF assembly and organization using fluorescent reporter, such as the small actin peptide Lifeact or GFP-actin, revealed a radially symmetric actin organization with differently assembled structures [54,55,56,57,58].
Initial engagement of the TCR–MHC-I complex activates actin polymerization in CTLs leading to a ring-shaped branched actin network in the dSMAC, shortly after contact with the target cell [48,59,60,61]. This Arp2/3-complex-mediated branched actin network, which exhibits high similarities to that of the lamellipodium of migrating cells, allows CTLs to spread across the surface of their targets [56,62], and provides support for the symmetric retrograde actin flow towards the cSMAC (see below). The pSMAC is composed of a lamella-like actin arc network that defines the radial symmetric contractile capability of this part of the IS [63]. Mechanistic studies have established that the assembly of the pSMAC actin arc is mediated by formins and organized by myosin II into antiparallel concentric arcs [63,64]. Indeed, selective depletion of the formin mDia prevents formation of the linear AF arrays required to generate the typical concentric myosin IIA-rich arcs [63,65,66]. The innermost part of the IS, the cSMAC, is referred to as an actin poor or actin hypodense region, which contains a fine AF network, which can only be visualized using super resolution imaging [53,66,67,68,69]. Moreover, the surface of CTLs it not flat, but is dominated by highly flexible and dynamic microvilli that show a high degree of AF polymerization at their core [70]. A recent study showed that using super resolution fluorescence imaging techniques and subsequent 3D reconstruction of the CTL membrane topography makes the tips of these membrane protrusions enriched for TCR clusters, which facilitate antigen screening. In addition, AF depolymerization not only inhibits microvilli formation, but also prevents the selective localization of TCR clusters [71]. Moreover, intercellular communication between CTLs and APCs is thought to be facilitated in parts by large, TCR-enriched membrane particles that are generated at the tip of these microvilli [72]. Delivery of these CTL-derived particles to cognate APCs was reported to increase DC activation status. Assembly of TCR microclusters was suggested to be a sequential process to which the actin cytoskeleton contributes by lowering the dissociation rates due to increased pulling forces at the IS, which maintain the different parts of the TCR signalosome together [73]. Intact actin dynamics may accordingly not only be involved in the retrograde movement of surface proteins towards the cSMAC but could also account for the observation that TCR microclusters are segregated from other protein clusters. Maintenance of the TCR signalosome composition during the lateral movement within the immune cell membrane is also subject to actin dynamics [74,75].
The integration of several different actin regulatory pathways underlying the formation of a functional IS makes this structure highly vulnerable to cytoskeletal alterations. The majority of actin cytoskeleton defects leading to primary immunodeficiencies (PIDs) are associated with a disruption of assembly and disassembly of AFs in multiple immune cell subtypes altering cell migration and adhesion, intercellular communication, intracellular signaling, and IS formation [76,77,78,79,80,81]. The architecture of the IS is subject to the spatiotemporal distinct activity of different actin polymerizing and regulatory proteins that result in topographical differences in AF organization. Engagement of activating receptors triggers Arp2/3 complex-dependent polymerization of branched AF, as illustrated in Figure 1. Polymerization activity of the Arp2/3 complex is regulated by different actin nucleation promoting factors (NPFs), such as WASp or WASp-verprolin homolog 2 (WAVE2). Actin regulation mediated by these NPFs does not occur on the effector immune cell side of the IS, but also plays an important role in DCs. Activity of WASp in target cells increases the overall IS area, and restricted knockdown of WASp in target DCs affects IS stability by inhibiting integrin polarization [82]. Actin cytoskeleton remodeling in target cells consequently represents a key component of normal IS formation through association of adhesive interaction and modulation of immunological synapse stability. These NPFs have to be associated with distinct functions corresponding to their localization within the IS. WAVE2 has been found primarily at the edges of the IS, and its activity has been associated with CTL adhesion and spreading over the antigen-presenting surface [83,84]. In addition to its role in modulation of IS stability, WASp is involved in the formation of actin-rich membrane protrusions at the secretory area, which is consistent with its predominant localization at the center of the IS [85,86,87].
Gene-expression profiling of peripheral CTLs from chronic lymphocytic leukemia (CLL) patients revealed a set of dysregulated actin regulatory proteins that was upregulated after cell-cell contact with leukemia cells [88]. Interestingly, the same cytoskeletal defects could be reproduced with CTLs from healthy donors that were cocultured with patient-derived CLL cells, indicating that cancer cells can promote cytoskeletal changes in cytotoxic lymphocytes. Further evaluation of the underlying mechanism identified several upregulated inhibitory ligands on leukemic cells that compromised the activation of key actin regulators including the Ras homolog (Rho) GTPases RhoA and Rac1, while promoting Cdc42 activity [89,90]. The combination of reduced RhoA/Rac1 signaling and increased Cdc42 activation resulted in the negative regulation of integrin activity in CTLs and subsequent adhesion and motility defects. Interestingly, defective actin polymerization was not limited to the IS between CLL cells and CTLs, but was also observed between APCs and CTLs, indicating that the cytoskeletal modifications in CTLs preventing functional IS formation were sustained over an extended period of time. A possible explanation for these effects comes from an elegant study in which it was demonstrated that leukemia cell-mediated changes in Rho GTPase activation result in impaired integrin signaling in CTLs, and that these defects could be rescued by the administration of the immunomodulatory drug lenalidomide, which resulted in the normalization of Rho GTPase activation signaling [90,91].
The ability of tumor cells to have a lasting negative impact on actin cytoskeletal dynamics in CTLs by compromising regulation of Rho GTPases shows a potential Achilles´ heel of cytotoxic lymphocytes. A variety of actin cytoskeleton regulators are involved at different spatiotemporal points during the formation and activity of a lytic IS, some of them were mentioned above. Dysregulation of one or two of these proteins can have a significant impact on anti-tumor immunity by inhibiting IS formation or affecting related functions, such as directed degranulation.
2.2. Actin Cytoskeleton Organization and Integrin Signaling at the Immunological Synapse
Binding of cytotoxic lymphocytes to target cells and IS maturation are dependent on the interaction of integrins on both sides of the IS. Cytotoxicity and polarized degranulation rely on a tight connection between CTLs and their prospective target cell. The integrin lymphocyte function-associated antigen 1 (LFA-1) was recently reported to be organized in nanometer-scaled clusters around the secretory domain of cytotoxic CD8+ T lymphocytes to allow confined delivery of cytotoxic granules [64,92,93]. The pSMAC shows the highest density of integrins, with a further increased LFA-1 density at the pSMAC/cSMAC boundary [94,95,96]. This integrin-mediated adhesive ring around the cSMAC was also suggested to help to seal the directed degranulation of lytic vesicles in the direction of the target cell [97,98]. Additionally, the sheer forces generated by the retrograde actin flow support binding of LFA-1 to its ligand intercellular adhesion molecule 1 (ICAM-1) by inducing a conformational change to its high affinity form. Beside this adhesion function, ICAM-1-bound LFA-1 also serves as a costimulatory signal for CTL and NK cell activation [96,99,100]. Topographical positioning and clustering of LFA-1 at the IS was recently shown to be confined to interstices within the actin cytoskeleton meshwork and underlie the activity of the actin nucleator WASp in CTLs [101]. The mobility of ICAM-1 in turn influences the actin flow-dependent signaling and cytotoxic lymphocyte functions. The actin cytoskeleton of antigen-presenting cells (APCs) engaged in an IS with CTLs was reported to limit the mobility of ICAM-1 on the cell surface. Release of this restriction by actin depolymerization in APCs negatively influences LFA-1 activation on T cells and adhesion properties [102]. Resistance of non-small cell lung cancer cells to CTL-mediated lysis was associated with the overexpression of the actin cytoskeleton regulators scinderin and ephrin-1A [103]. Scinderin is an actin severing protein and its overexpression is associated with increased actin dynamics, while signals of the receptor tyrosine kinases ephrin-A1 are translated into increased actin dynamics via activation of the N-WASp–Arp2/3 complex-pathway [104,105]. Silencing these genes using RNA interference restores both actin dynamics and cancer cell susceptibility to CTL-mediated lysis, giving indirect evidence that altered actin dynamics in cancer cells promotes immune evasion [103]. Detailed analysis of the IS formed between resistant non-small cell lung cancer (NSCLC) cells and CTLs revealed significant structural changes with sparse areas of tight cell-cell junctions and a wide IS cleft, contrasting with the tight IS formed with susceptible NSCLC cells. In a follow up study, the focal adhesion kinase (FAK) signaling pathway and Rho GTPase signaling were found to play a role in NSCLC resistance to CTLs [106]. FAK is a non-receptor tyrosine kinase that is activated by phosphorylation in response to integrin signaling [107]. Activated FAK can be found where cells attach to extracellular matrix components and at cell-cell contact areas, where it plays an important role in regulating AF turnover [108]. Expression of FAK has been reported to be upregulated in certain types of cancer, including ovarian and breast cancer [109]. Experimental silencing of FAK in NSCLC induces both, adhesion deficiency and increased resistance to CTL-mediated lysis by impairing IS formation [103,106].
These studies highlight that adhesion of CTLs to their prospective target is a crucial prerequisite for effective target cell lysis. Actin cytoskeletal dynamics drive the segregation of transmembrane protein clusters, such as integrins and induce their maturation by exertion of sheer forces to further increase their ligand affinity. Cancer cells can exploit the dependence of integrins for immobilized ligands by increasing actin cytoskeletal dynamics and increased ligands motility to prevent formation of the sealing ring at the pSMAC/cSMAC border to reduce efficiency of cytotoxic granule delivery.
3. Actin Cytoskeleton and Immunological Synapse Signaling
3.1. Actin Cytoskeleton and Receptor Signaling
The majority of engaged TCRs assemble in microclusters with their associated costimulatory receptors in the so-called TCR signalosome and can be found within the pSMAC. Signalosomes can also be found in the NK cell IS, although their composition differs from the TCR signalosome [110,111]. From the periphery, signalosomes move following the retrograde actin flow until they reach the pSMAC/cSMAC boundary [56,64]. Treatment with actin depolymerizing drugs, such as latrunculin, prevents microcluster-formation and consequently CTL and NK cell activation, highlighting the importance of actin dynamics in signaling at the IS [58,95]. Robust signaling of TCR microclusters occurs during the transition from the dSMAC to the cSMAC, with an abrupt stop upon reaching of the cSMAC [58]. As part of the TCR signalosome, linker of activation of T cells (LAT) recruits other adaptor proteins such as Nck and Vav1 that integrate TCR signaling into the rearrangement of the actin cytoskeleton by modulating WASp activity [60,66,112]. In the actin hypodense cSMAC, the connection between the signalosome and the actin cytoskeleton is lost, and thus TCR signaling comes to an abrupt stop after transition of TCR microclusters into the cSMAC, where exhausted TCRs are internalized in a Rho-dependent manner [35,58,113]. The strength of TCR signaling is directly correlated to the time it takes TCR microclusters to transit through the dSMAC/pSMAC region of the IS and, in primary CTLs, this depends on the speed of the retrograde actin flow [63,65]. Integrin signaling slows the retrograde actin flow resulting in activating signal by the TCR over longer periods of time. The actin-based molecular motor myosin IIA is not only involved in CTL motility and spreading over an antigen-coated surface, but has also been suggested to take part in the formation of the pSMAC and cSMAC, maturation of the IS, and persistence of TCR signaling [114]. Formation of TCR microclusters and their retrograde flow were both associated with recruitment and sustained activity of myosin IIA [115]. However, the exact role and importance of myosin IIA in IS formation and TCR signaling remain a matter of debate [114,116].
Although the role of activating receptor microclusters in activation of cytotoxic lymphocytes has been well established, much less is known about the distribution and stabilization of activating and inhibitory ligands on target cells. Cytotoxic lymphocytes do not only form an IS with infected or cancer cells, but also with APCs, such as DCs, with the aim to promote immune functions on both sides of the IS. In this regard, recent studies have shown that DCs also exhibit prominent actin polymerization in the region of the IS upon conjugation with CTLs and NK cells [43,82]. Such cytoskeletal remodeling was also shown to stabilize inhibitory ligands at the IS, and thereby prevent activation of immune cell effector functions. Preventing actin cytoskeleton polymerization in DCs was sufficient to increase the release of cytokines and cytotoxicity of NK cells against conjugates DCs. Such an IS was termed “regulatory synapse” [45] and was also reported between CTLs and DCs [82,117]. All in all, these works support that the actin cytoskeleton plays an important role in bridging TCR signaling to AF polymerization and that actin polymerization in DCs are the basis for the strong inhibitory signals in a regulatory IS.
Classification of functionally different ISs was proposed based on the status of actin polymerization on both sides of the IS, including the inhibitory synapse with a healthy target cell (AF accumulation on either side), the lytic synapse with a transformed target cell (AF accumulation in the immune cell only), and the regulatory synapse between DC and cytotoxic immune cells (AF accumulation in the APC only, as seen in Figure 1) [45]. Recently, we provided evidence that cancer cells use a similar mechanism as the regulatory synapse to prevent the activation of interacting NK cells [44,118]. Using high resolution confocal microscopy and high-throughput techniques, we found that resistant breast cancer cells in conjugation with cytotoxic NK cells accumulate AFs at their side of the IS, while susceptible cells do not [44]. As observed with DCs [43], the inhibition of actin polymerization using RNA interference was sufficient to abolish cancer cell resistance to NK cell-mediated lysis [44]. Moreover, actin accumulation in resistant cancer cells was also associated with increased levels of inhibitory ligands, including PD-L1 and MHC-I, at the IS. Both types of ligands induce negative receptor signaling in NK cells and can tip the balance between activating and inhibitory signaling in favor of cancer cells. This suggests that resistant cancer cells can mimic an increased expression of inhibitory ligands and thereby escape killing by NK cells, and possibly, by CTLs (through PD-L1) [118]. Accordingly, we propose a fourth type of IS that is formed between NK cells (possibly CTLs) and resistant cancer cells, which is characterized by actin polymerization on the target cell side (Figure 1). We named it “evasion synapse”.
3.2. Actin Cytoskeleton and Mechanopotentiation of Signaling and Killing at the Immunological Synapse
Over the last years, it has become increasingly clear that the actin cytoskeleton is not only a platform, but actively involved in the integration of spatial, molecular, and biochemical signals, as well as mechanical features into the formation and maintenance of the IS [119,120,121,122]. The forces submitted in the context of the IS are the result of membrane dynamics, cell spreading, and rearrangement of the actomyosin cytoskeleton.
Recently, direct evidence was given that the TCR acts as a mechanoreceptor and that activation of TCR signaling only occurs during applied tangential force, indicating that TCR–MHC-I interaction alone is not sufficient for CTL activation [123,124]. Following its activation, the TCR induces force changes at the IS by triggering dynamic actin cytoskeleton reorganization that translates into biochemical signals. As a result of antigen recognition, integrin-mediated adhesion force increases, with a direct correlation between TCR specificity and the strength of adhesion forces [125,126,127]. Forces in the frame of the IS have been shown to influence affinity maturation of integrins and strengthening of LFA-1/ICAM-1 interaction at the IS [128,129]. In addition, it was demonstrated that the cytotoxicity of CTLs is directly correlated to the mechanical forces applied on the conjugated target cell [130]. Furthermore, the actin cytoskeleton plays a key role as a main driver of force generation at the IS with regard to the retrograde translocation of receptor clusters originating from pushing and pulling forces at the dSMAC and pSMAC, respectively. The previously mentioned integrin-mediated adhesive ring around the cSMAC region; helps to apply the actin-based pulling force that supports pore formation by perforin [64,130]. In turn, actin remodeling on the target cell side of the IS locally modifies membrane properties, e.g., by inducing a substantial increase in cellular stiffness that can alter CTL activation [120,121,126,131]. CTLs interact with a variety of different cells, including endothelial cells, APCs, and target cells. The viscoelastic properties of endothelial cells and APCs change in response to inflammatory conditions and these changes are associated with myosin activity and actin polymerization dynamics [132,133]. Interestingly, mature APCs exhibit a higher degree of actin filament reorganization and a polarization of AFs towards the IS region [134,135]. This actin rearrangement in APCs was found to be critical for functional IS formation and T cell priming. Furthermore, it has been reported that ex vivo activation, expansion, and differentiation of CTLs is dependent on substrate rigidity [136]. CTLs interacting with immobilized stimulatory ligands show increased activation and proliferation under conditions of variable substrate stiffness as a result of increased signaling by the mechanosensitive TCR and other receptors [119,136]. The increased actin accumulation observed in APCs is thus sufficient to directly influence mechanotransduction by stabilizing inhibitory ligands at the IS, but also important for the maintenance of TCR signaling by providing a rigid substrate [122]. Additionally, mechanical forces at the IS can potentiate CTL-mediated killing of target cells. A recent study reported, that actin-enriched membrane protrusion in the dSMAC and cSMAC of the IS facilitate directed delivery of cytotoxic granules while also inducing the physical deformation of the target cell membrane [87]. These protrusions were dependent on WASp and Arp2/3 complex activity at the central parts of the IS and were a crucial requirement for force exertion in the context of the IS and potentiated target cell lysis.
4. Actin Cytoskeleton and Immunological Synapse Cytolytic Activity
In addition to signaling function, the IS is also the place of directed secretion of cytolytic granules toward cancer cells. Following interaction with a target in a lytic IS, CTLs and NK cells both show dynein-mediated polarization of the microtubule organizing center (MTOC) towards the IS [137,138,139]. The docking of the MTOC to the plasma membrane at the boundary between the actin hypodense cSMAC and the secretory domain of the IS is required for intracellular trafficking of lytic granules to the secretory domain of the IS [97] as seen in Figure 1. The content of lytic granules induces target cell death by osmotic cell lysis or via activation of caspases or pro-apoptotic proteins [140,141]. The mechanical force necessary for MTOC reorganization towards the membrane has been reported to involve different actin regulatory proteins, in particular formin family members that bind to microtubule plus ends. Accordingly, depletion of FMNL-1 and mDIA-1 has been shown to result in irregular MTOC positioning at the lytic IS [142,143]. It was initially suggested that the cytolytic granules can passively pass through the loose mesh of cortical AFs in the cSMAC [67,68]. However, recent high-resolution imaging-based studies established that, while AF dynamics at the cSMAC are reduced as compared to the directional actin flow in the dSMAC and the pSMAC, a fine network of AFs at the cSMAC shows dynamics at nanoscale level [69,144].
In vitro studies using cover slips or planar lipid bilayers coated with activating ligands or antibodies mimicking the target cell surface showed that in NK cells and CTLs in interaction with an activating surface, results in increased AF network mesh size at the cSMAC. The actin network mesh size reaches its peak at the same time the highest value of directed degranulation is observed. This granule-permissive clearance of actin is conserved in lytic ISs of both, CTLs and NK cells, although their actin clearance kinetics differ. A recent study found that in an IS formed by primary CTLs, actin clearance at the cSMAC appeared earlier, reached its peak faster and was short-lived compared to actin meshwork depolymerization observed in primary NK cells [69]. Using lattice light-sheet imaging, it could be shown that the MTOC and lytic granules clustered around it were positioned at the center of the cSMAC [53,139]. Based on the above results, it has been proposed that the actin network at the cSMAC serves as a barrier for lytic granules, and that transitory AF depletion decreases actin network density to initiate degranulation events [139]. Additionally, actin dynamics and myosin contractility have been shown to support the fusion of lytic granules with the plasma membrane by providing the required membrane tension. Furthermore, while myosin IIA activity was shown to be dispensable for NK cell adhesion to target cells, it was reported to interact directly with lytic granules, and facilitate the approach to the cell membrane at the secretory domain of the cSMAC [145,146]. Accordingly, mutations in the heavy chain of non-muscle myosin IIA were reported to be associated with immunodeficiency syndromes characterized by decreased CTL-mediated cytotoxicity despite otherwise normal IS formation [147,148,149]. The approach of lytic granules to the CTL membrane is accompanied by coronin 1A-mediated deconstruction of AFs at the cSMAC [53]. Another recent study also showed that secretion of perforin was associated with the formation of actin-rich protrusions in the secretory domain of CTLs, additionally highlighting that the cSMAC is not simply an actin-poor region as previously suggested, but exhibits highly regulated actin dynamics [87]. The termination of degranulation events is also actin cytoskeleton-dependent, as replenishment of AFs at the cSMAC acts as barrier for further granule degranulation and results in retraction of the MTOC from its position close to the IS area [150]. Therefore, rapid actin cytoskeleton remodeling seems to be crucial for initiation, but also the termination of directed secretion. Treatment of CTLs with actin depolymerizing drugs, such as Latrunculin A, increases duration of lytic granule secretion, underlining the role of AFs in creating a physical barrier for granule release.
Although only a few studies have directly investigated the organization and functions of the actin cytoskeleton in cancer cells during cytotoxic lymphocyte attack, there is growing evidence that actin remodeling plays a central role in mediating tumor immune evasion by altering either IS formation and function [44,103,106,151]. As previously discussed, we recently reported a prominent accumulation of AFs on the tumor cell side of the IS between intrinsically resistant breast tumor cells and NK cells, and that such process was correlated to immune evasion [44]. Selective targeting of actin cytoskeleton remodeling in cancer cells by targeted downregulation of the ARP2/3 complex regulators Cdc42 or N-WASP was sufficient to substantially increase target cell susceptibility to NK cell-induced lysis, as well as to restore high levels of NK cell derived-granzyme B levels in target cells. Similar to the regulatory synapse between DCs and NK cells, key inhibitory ligands, including HLA-A,B,C and PD-L1, were observed to accumulate at the evasion synapse together with actin accumulation in resistant cancer cells (Figure 1). A central role for the actin cytoskeleton in tumor immune evasion is further supported by an immune escape screen that identified Cdc42 as a mediator of increased resistance to antigen-specific CTL-mediated cytotoxicity [152]. Importantly, the expression of a constitutively active form of Cdc42 in mouse fibrosarcoma and human colorectal cancer cells was sufficient to induce resistance against CTLs and NK cell cytotoxicity in vitro, and promoted tumor growth in different mouse models, while it did not affect cell proliferation in vitro. Finally, additional evidence that actin remodeling promotes tumor immune evasion comes from the multiple links between the epithelial-to-mesenchymal transition (EMT), a process involving extensive modification of actin cytoskeleton organization and dynamics [153], and acquisition of cancer cell resistance to cytotoxic lymphocytes [154,155,156,157,158]. Such links have been recently reviewed and the reader is invited to refer to the following article [3].
5. Conclusions
Actin remodeling plays central roles in the three main processes underlying cytotoxic lymphocyte-mediated tumor cell killing, including target cell recognition, immune cell activation, and cancer cell killing. Actin remodeling at the cancer cell side of IS is emerging as an important mechanism of tumor immune evasion; however, our knowledge regarding this aspect remains fragmented. Additional work is required to identify clinically relevant targets to selectively impair actin organization and/or the dynamics in tumor cells and thereby restore an effective anti-tumor immune response. If and how the actin cytoskeleton of cancer cells could serve as a drug target for immunotherapy should be further investigated.
Funding
A.A.A. and H.W. are recipients of PhD fellowships from the National Research Fund (FNR), Luxembourg (AFR7892325 and PRIDE15/10675146/CANBIO, respectively). Work in C. Thomas’s group is supported by the Cancer Foundation Luxembourg (ACTIMMUNE).
Acknowledgments
The authors thank J. Michalska for her constructive criticism and proofreading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Kim, R.; Emi, M.; Tanabe, K. Cancer immunoediting from immune surveillance to immune escape. Immunology 2007, 121, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, S.; Savagner, P.; Ortiz-Cuaran, S.; Mahjoubi, L.; Saintigny, P.; Thiery, J.P.; Chouaib, S. New insights into the role of EMT in tumor immune escape. Mol. Oncol. 2017, 11, 824–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, P. Ueber den jetzigen Stand der Karzinomforschung. 1908. Available online: https://www.pei.de/SharedDocs/Downloads/institut/veroeffentlichungen-von-paul-ehrlich/1906-1914/1909-karzinomforschung.pdf;jsessionid=C5C5AF872FC8ED888657A08D4BBB4B1B.1_cid344?__blob=publicationFile&v=1 (accessed on 15 April 2019).
- Burnet, F.M. Immunological surveillance in neoplasia. Transplant. Rev. 1971, 7, 3–25. [Google Scholar] [CrossRef]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The three Es of cancer immunoediting. Annu. Rev. Immunol. 2004, 22, 329–360. [Google Scholar] [CrossRef] [PubMed]
- Bubenik, J. MHC class I down-regulation: Tumour escape from immune surveillance? (review). Int. J. Oncol. 2004, 25, 487–491. [Google Scholar] [CrossRef]
- Fiegler, N.; Textor, S.; Arnold, A.; Rolle, A.; Oehme, I.; Breuhahn, K.; Moldenhauer, G.; Witzens-Harig, M.; Cerwenka, A. Downregulation of the activating NKp30 ligand B7-H6 by HDAC inhibitors impairs tumor cell recognition by NK cells. Blood 2013, 122, 684–693. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.; Emi, M.; Tanabe, K.; Arihiro, K. Tumor-driven evolution of immunosuppressive networks during malignant progression. Cancer Res. 2006, 66, 5527–5536. [Google Scholar] [CrossRef]
- Crespo, J.; Sun, H.; Welling, T.H.; Tian, Z.; Zou, W. T cell anergy, exhaustion, senescence, and stemness in the tumor microenvironment. Curr. Opin. Immunol. 2013, 25, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Gabrilovich, D.I. Hypoxia-inducible factors in regulation of immune responses in tumour microenvironment. Immunology 2014, 143, 512–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhard, A.; Kahlert, S.; Goede, V.; Hemmerlein, B.; Plate, K.H.; Augustin, H.G. Heterogeneity of angiogenesis and blood vessel maturation in human tumors: Implications for antiangiogenic tumor therapies. Cancer Res. 2000, 60, 1388–1393. [Google Scholar]
- Azab, A.K.; Hu, J.; Quang, P.; Azab, F.; Pitsillides, C.; Awwad, R.; Thompson, B.; Maiso, P.; Sun, J.D.; Hart, C.P.; et al. Hypoxia promotes dissemination of multiple myeloma through acquisition of epithelial to mesenchymal transition-like features. Blood 2012, 119, 5782–5794. [Google Scholar] [CrossRef] [PubMed]
- Baginska, J.; Viry, E.; Paggetti, J.; Medves, S.; Berchem, G.; Moussay, E.; Janji, B. The critical role of the tumor microenvironment in shaping natural killer cell-mediated anti-tumor immunity. Front. Immunol. 2013, 4, 490. [Google Scholar] [CrossRef]
- Baudry, N.; Danialou, G.; Boczkowski, J.; Vicaut, E. In vivo study of the effect of systemic hypoxia on leukocyte-endothelium interactions. Am. J. Respir. Crit. Care Med. 1998, 158, 477–483. [Google Scholar] [CrossRef]
- Siemens, D.R.; Hu, N.; Sheikhi, A.K.; Chung, E.; Frederiksen, L.J.; Pross, H.; Graham, C.H. Hypoxia increases tumor cell shedding of MHC class I chain-related molecule: Role of nitric oxide. Cancer Res. 2008, 68, 4746–4753. [Google Scholar] [CrossRef]
- Yang, M.; Ma, C.; Liu, S.; Sun, J.; Shao, Q.; Gao, W.; Zhang, Y.; Li, Z.; Xie, Q.; Dong, Z.; et al. Hypoxia skews dendritic cells to a T helper type 2-stimulating phenotype and promotes tumour cell migration by dendritic cell-derived osteopontin. Immunology 2009, 128, e237–e249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartsock, A.; Nelson, W.J. Adherens and tight junctions: Structure, function and connections to the actin cytoskeleton. Biochim Biophys Acta 2008, 1778, 660–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Heng, Y.W.; Koh, C.G. Actin cytoskeleton dynamics and the cell division cycle. Int. J. Biochem. Cell Biol. 2010, 42, 1622–1633. [Google Scholar] [CrossRef]
- Freeman, S.A.; Grinstein, S. Phagocytosis: Receptors, signal integration, and the cytoskeleton. Immunol. Rev. 2014, 262, 193–215. [Google Scholar] [CrossRef]
- Gurel, P.S.; Hatch, A.L.; Higgs, H.N. Connecting the cytoskeleton to the endoplasmic reticulum and Golgi. Curr. Biol. 2014, 24, R660–R672. [Google Scholar] [CrossRef]
- Chugh, P.; Clark, A.G.; Smith, M.B.; Cassani, D.A.D.; Dierkes, K.; Ragab, A.; Roux, P.P.; Charras, G.; Salbreux, G.; Paluch, E.K. Actin cortex architecture regulates cell surface tension. Nat. Cell Biol. 2017, 19, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kast, D.J.; Dominguez, R. The Cytoskeleton-Autophagy Connection. Curr. Biol. 2017, 27, R318–R326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Dominguez, R. Regulation of actin cytoskeleton dynamics in cells. Mol. Cells 2010, 29, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D. Actin and Actin-Binding Proteins. Cold Spring Harb Perspect Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C. Bundling actin filaments from membranes: Some novel players. Front. Plant. Sci. 2012, 3, 188. [Google Scholar] [CrossRef]
- Mullins, R.D.; Heuser, J.A.; Pollard, T.D. The interaction of Arp2/3 complex with actin: Nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proc. Natl. Acad. Sci. USA 1998, 95, 6181. [Google Scholar] [CrossRef] [PubMed]
- Pruyne, D.; Evangelista, M.; Yang, C.; Bi, E.; Zigmond, S.; Bretscher, A.; Boone, C. Role of Formins in Actin Assembly: Nucleation and Barbed-End Association. Science 2002, 297, 612. [Google Scholar] [CrossRef]
- Romero, S.; Le Clainche, C.; Didry, D.; Egile, C.; Pantaloni, D.; Carlier, M.-F. Formin Is a Processive Motor that Requires Profilin to Accelerate Actin Assembly and Associated ATP Hydrolysis. Cell 2004, 119, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Tholl, S.; Moes, D.; Dieterle, M.; Papuga, J.; Moreau, F.; Steinmetz, A. Actin bundling in plants. Cell Motil. Cytoskelet. 2009, 66, 940–957. [Google Scholar] [CrossRef] [Green Version]
- Carlier, M.F.; Laurent, V.; Santolini, J.; Melki, R.; Didry, D.; Xia, G.X.; Hong, Y.; Chua, N.H.; Pantaloni, D. Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: Implication in actin-based motility. J. Cell Biol. 1997, 136, 1307–1322. [Google Scholar] [CrossRef]
- Piragyte, I.; Jun, C.D. Actin engine in immunological synapse. Immune Netw. 2012, 12, 71–83. [Google Scholar] [CrossRef]
- Kumari, S.; Curado, S.; Mayya, V.; Dustin, M.L. T cell antigen receptor activation and actin cytoskeleton remodeling. Biochim Biophys Acta 2014, 1838, 546–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dustin, M.L. The immunological synapse. Cancer Immunol. Res. 2014, 2, 1023–1033. [Google Scholar] [CrossRef] [Green Version]
- Thiery, J.; Keefe, D.; Boulant, S.; Boucrot, E.; Walch, M.; Martinvalet, D.; Goping, I.S.; Bleackley, R.C.; Kirchhausen, T.; Lieberman, J. Perforin pores in the endosomal membrane trigger the release of endocytosed granzyme B into the cytosol of target cells. Nat. Immunol. 2011, 12, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Pahl, J.; Cerwenka, A. Tricking the balance: NK cells in anti-cancer immunity. Immunobiology 2017, 222, 11–20. [Google Scholar] [CrossRef]
- Chowdhury, D.; Lieberman, J. Death by a Thousand Cuts: Granzyme Pathways of Programmed Cell Death. Annu. Rev. Immunol. 2008, 26, 389–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, J.A.; Susanto, O.; Jenkins, M.R.; Lukoyanova, N.; Sutton, V.R.; Law, R.H.P.; Johnston, A.; Bird, C.H.; Bird, P.I.; Whisstock, J.C.; et al. Perforin forms transient pores on the target cell plasma membrane to facilitate rapid access of granzymes during killer cell attack. Blood 2013, 121, 2659. [Google Scholar] [CrossRef]
- Pardo, J.; Bosque, A.; Brehm, R.; Wallich, R.; Naval, J.; Müllbacher, A.; Anel, A.; Simon, M.M. Apoptotic pathways are selectively activated by granzyme A and/or granzyme B in CTL-mediated target cell lysis. J. Cell Biol. 2004, 167, 457. [Google Scholar] [CrossRef]
- Brossard, C.; Feuillet, V.; Schmitt, A.; Randriamampita, C.; Romao, M.; Raposo, G.; Trautmann, A. Multifocal structure of the T cell - dendritic cell synapse. Eur. J. Immunol. 2005, 35, 1741–1753. [Google Scholar] [CrossRef] [Green Version]
- Tseng, S.Y.; Waite, J.C.; Liu, M.; Vardhana, S.; Dustin, M.L. T cell-dendritic cell immunological synapses contain TCR-dependent CD28-CD80 clusters that recruit protein kinase C theta. J. Immunol. 2008, 181, 4852–4863. [Google Scholar] [CrossRef] [PubMed]
- Barreira da Silva, R.; Graf, C.; Munz, C. Cytoskeletal stabilization of inhibitory interactions in immunologic synapses of mature human dendritic cells with natural killer cells. Blood 2011, 118, 6487–6498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Absi, A.; Wurzer, H.; Guerin, C.; Hoffmann, C.; Moreau, F.; Mao, X.; Brown-Clay, J.; Petrolli, R.; Casellas, C.P.; Dieterle, M.; et al. Actin Cytoskeleton Remodeling Drives Breast Cancer Cell Escape from Natural Killer-Mediated Cytotoxicity. Cancer Res. 2018, 78, 5631–5643. [Google Scholar] [CrossRef] [PubMed]
- Mace, E.M.; Orange, J.S. Multiple distinct NK-cell synapses. Blood 2011, 118, 6475–6476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkhardt, J.K.; Carrizosa, E.; Shaffer, M.H. The actin cytoskeleton in T cell activation. Annu. Rev. Immunol. 2008, 26, 233–259. [Google Scholar] [CrossRef] [PubMed]
- Smith-Garvin, J.E.; Koretzky, G.A.; Jordan, M.S. T Cell Activation. Annu. Rev. Immunol. 2009, 27, 591–619. [Google Scholar] [CrossRef]
- Valitutti, S.; Dessing, M.; Aktories, K.; Gallati, H.; Lanzavecchia, A. Sustained signaling leading to T cell activation results from prolonged T cell receptor occupancy. Role of T cell actin cytoskeleton. J. Exp. Med. 1995, 181, 577. [Google Scholar] [CrossRef] [PubMed]
- Nolz, J.C.; Gomez, T.S.; Zhu, P.; Li, S.; Medeiros, R.B.; Shimizu, Y.; Burkhardt, J.K.; Freedman, B.D.; Billadeau, D.D. The WAVE2 Complex Regulates Actin Cytoskeletal Reorganization and CRAC-Mediated Calcium Entry during T Cell Activation. Curr. Biol. 2006, 16, 24–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauriat, C.; Long, E.O.; Ljunggren, H.G.; Bryceson, Y.T. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood 2010, 115, 2167–2176. [Google Scholar] [CrossRef]
- Bryceson, Y.T.; Chiang, S.C.; Darmanin, S.; Fauriat, C.; Schlums, H.; Theorell, J.; Wood, S.M. Molecular mechanisms of natural killer cell activation. J. Innate Immun. 2011, 3, 216–226. [Google Scholar] [CrossRef]
- Davis, D.M.; Chiu, I.; Fassett, M.; Cohen, G.B.; Mandelboim, O.; Strominger, J.L. The human natural killer cell immune synapse. Proc. Natl. Acad. Sci. USA 1999, 96, 15062–15067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mace, E.M.; Orange, J.S. Lytic immune synapse function requires filamentous actin deconstruction by Coronin 1A. Proc. Natl. Acad. Sci. USA 2014, 111, 6708–6713. [Google Scholar] [CrossRef] [Green Version]
- Riedl, J.; Crevenna, A.H.; Kessenbrock, K.; Yu, J.H.; Neukirchen, D.; Bista, M.; Bradke, F.; Jenne, D.; Holak, T.A.; Werb, Z.; et al. Lifeact: A versatile marker to visualize F-actin. Nat. Methods 2008, 5, 605. [Google Scholar] [CrossRef] [PubMed]
- Bunnell, S.C.; Kapoor, V.; Trible, R.P.; Zhang, W.; Samelson, L.E. Dynamic actin polymerization drives T cell receptor-induced spreading: A role for the signal transduction adaptor LAT. Immunity 2001, 14, 315–329. [Google Scholar] [CrossRef]
- DeMond, A.L.; Mossman, K.D.; Starr, T.; Dustin, M.L.; Groves, J.T. T cell receptor microcluster transport through molecular mazes reveals mechanism of translocation. Biophys. J. 2008, 94, 3286–3292. [Google Scholar] [CrossRef] [PubMed]
- Kaizuka, Y.; Douglass, A.D.; Varma, R.; Dustin, M.L.; Vale, R.D. Mechanisms for segregating T cell receptor and adhesion molecules during immunological synapse formation in Jurkat T cells. Proc. Natl. Acad. Sci. USA 2007, 104, 20296–20301. [Google Scholar] [CrossRef] [Green Version]
- Varma, R.; Campi, G.; Yokosuka, T.; Saito, T.; Dustin, M.L. T cell receptor-proximal signals are sustained in peripheral microclusters and terminated in the central supramolecular activation cluster. Immunity 2006, 25, 117–127. [Google Scholar] [CrossRef]
- Saito, T.; Yokosuka, T. Immunological synapse and microclusters: The site for recognition and activation of T cells. Curr. Opin. Immunol. 2006, 18, 305–313. [Google Scholar] [CrossRef]
- Rohatgi, R.; Nollau, P.; Ho, H.Y.; Kirschner, M.W.; Mayer, B.J. Nck and phosphatidylinositol 4,5-bisphosphate synergistically activate actin polymerization through the N-WASP-Arp2/3 pathway. J. Biol. Chem. 2001, 276, 26448–26452. [Google Scholar] [CrossRef]
- Fischer, K.D.; Kong, Y.Y.; Nishina, H.; Tedford, K.; Marengère, L.E.M.; Kozieradzki, I.; Sasaki, T.; Starr, M.; Chan, G.; Gardener, S.; et al. Vav is a regulator of cytoskeletal reorganization mediated by the T-cell receptor. Curr. Biol. 1998, 8, 554–562. [Google Scholar] [CrossRef]
- Jankowska, K.I.; Burkhardt, J.K. Analyzing Actin Dynamics at the Immunological Synapse. Methods Mol. Biol. 2017, 1584, 7–29. [Google Scholar] [PubMed]
- Yi, J.; Wu, X.S.; Crites, T.; Hammer, J.A. Actin retrograde flow and actomyosin II arc contraction drive receptor cluster dynamics at the immunological synapse in Jurkat T cells. Mol. Biol Cell. 2012, 23, 834–852. [Google Scholar] [CrossRef] [Green Version]
- Murugesan, S.; Hong, J.; Yi, J.; Li, D.; Beach, J.R.; Shao, L.; Meinhardt, J.; Madison, G.; Wu, X.; Betzig, E.; et al. Formin-generated actomyosin arcs propel T cell receptor microcluster movement at the immune synapse. J. Cell Biol. 2016, 215, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Babich, A.; Li, S.; O’Connor, R.S.; Milone, M.C.; Freedman, B.D.; Burkhardt, J.K. F-actin polymerization and retrograde flow drive sustained PLCgamma1 signaling during T cell activation. J. Cell Biol. 2012, 197, 775–787. [Google Scholar] [CrossRef]
- Hammer, J.A.; Wang, J.C.; Saeed, M.; Pedrosa, A.T. Origin, Organization, Dynamics, and Function of Actin and Actomyosin Networks at the T Cell Immunological Synapse. Annu. Rev. Immunol. 2019, 37, 201–224. [Google Scholar] [CrossRef]
- Brown, A.C.; Oddos, S.; Dobbie, I.M.; Alakoskela, J.M.; Parton, R.M.; Eissmann, P.; Neil, M.A.; Dunsby, C.; French, P.M.; Davis, I.; et al. Remodelling of cortical actin where lytic granules dock at natural killer cell immune synapses revealed by super-resolution microscopy. PLoS Biol. 2011, 9, e1001152. [Google Scholar] [CrossRef] [PubMed]
- Rak, G.D.; Mace, E.M.; Banerjee, P.P.; Svitkina, T.; Orange, J.S. Natural killer cell lytic granule secretion occurs through a pervasive actin network at the immune synapse. PLoS Biol. 2011, 9, e1001151. [Google Scholar] [CrossRef]
- Carisey, A.F.; Mace, E.M.; Saeed, M.B.; Davis, D.M.; Orange, J.S. Nanoscale Dynamism of Actin Enables Secretory Function in Cytolytic Cells. Curr. Biol. 2018, 28, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Nijhara, R.; van Hennik, P.B.; Gignac, M.L.; Kruhlak, M.J.; Hordijk, P.L.; Delon, J.; Shaw, S. Rac1 mediates collapse of microvilli on chemokine-activated T lymphocytes. J. Immunol. 2004, 173, 4985–4993. [Google Scholar] [CrossRef]
- Jung, Y.; Riven, I.; Feigelson, S.W.; Kartvelishvily, E.; Tohya, K.; Miyasaka, M.; Alon, R.; Haran, G. Three-dimensional localization of T-cell receptors in relation to microvilli using a combination of superresolution microscopies. Proc. Natl. Acad. Sci. USA 2016, 113, E5916–E5924. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-R.; Mun, Y.; Lee, K.-S.; Park, Y.-J.; Park, J.-S.; Park, J.-H.; Jeon, B.-N.; Kim, C.-H.; Jun, Y.; Hyun, Y.-M.; et al. T cell microvilli constitute immunological synaptosomes that carry messages to antigen-presenting cells. Nat. Commun. 2018, 9, 3630. [Google Scholar] [CrossRef]
- Yi, J.; Balagopalan, L.; Nguyen, T.; McIntire, K.M.; Samelson, L.E. TCR microclusters form spatially segregated domains and sequentially assemble in calcium-dependent kinetic steps. Nat. Commun. 2019, 10, 277. [Google Scholar] [CrossRef]
- Lillemeier, B.F.; Mortelmaier, M.A.; Forstner, M.B.; Huppa, J.B.; Groves, J.T.; Davis, M.M. TCR and Lat are expressed on separate protein islands on T cell membranes and concatenate during activation. Nat. Immunol. 2010, 11, 90–96. [Google Scholar] [CrossRef]
- Feigelson, S.W.; Pasvolsky, R.; Cemerski, S.; Shulman, Z.; Grabovsky, V.; Ilani, T.; Sagiv, A.; Lemaitre, F.; Laudanna, C.; Shaw, A.S.; et al. Occupancy of lymphocyte LFA-1 by surface-immobilized ICAM-1 is critical for TCR- but not for chemokine-triggered LFA-1 conversion to an open headpiece high-affinity state. J. Immunol. 2010, 185, 7394–7404. [Google Scholar] [CrossRef]
- Westerberg, L.S.; Meelu, P.; Baptista, M.; Eston, M.A.; Adamovich, D.A.; Cotta-de-Almeida, V.; Seed, B.; Rosen, M.K.; Vandenberghe, P.; Thrasher, A.J.; et al. Activating WASP mutations associated with X-linked neutropenia result in enhanced actin polymerization, altered cytoskeletal responses, and genomic instability in lymphocytes. J. Exp. Med. 2010, 207, 1145–1152. [Google Scholar] [CrossRef]
- Moulding, D.A.; Record, J.; Malinova, D.; Thrasher, A.J. Actin cytoskeletal defects in immunodeficiency. Immunol. Rev. 2013, 256, 282–299. [Google Scholar] [CrossRef] [Green Version]
- Burns, S.O.; Zarafov, A.; Thrasher, A.J. Primary immunodeficiencies due to abnormalities of the actin cytoskeleton. Curr. Opin. Hematol. 2017, 24, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Dahlberg, C.I.; Torres, M.L.; Petersen, S.H.; Baptista, M.A.; Keszei, M.; Volpi, S.; Grasset, E.K.; Karlsson, M.C.; Walter, J.E.; Snapper, S.B.; et al. Deletion of WASp and N-WASp in B cells cripples the germinal center response and results in production of IgM autoantibodies. J. Autoimmun. 2015, 62, 81–92. [Google Scholar] [CrossRef]
- Baptista, M.A.; Keszei, M.; Oliveira, M.; Sunahara, K.K.; Andersson, J.; Dahlberg, C.I.; Worth, A.J.; Lieden, A.; Kuo, I.C.; Wallin, R.P.; et al. Deletion of Wiskott-Aldrich syndrome protein triggers Rac2 activity and increased cross-presentation by dendritic cells. Nat. Commun. 2016, 7, 12175. [Google Scholar] [CrossRef]
- Keszei, M.; Record, J.; Kritikou, J.S.; Wurzer, H.; Geyer, C.; Thiemann, M.; Drescher, P.; Brauner, H.; Kocher, L.; James, J.; et al. Constitutive activation of WASp in X-linked neutropenia renders neutrophils hyperactive. J. Clin. Investig. 2018, 128, 4115–4131. [Google Scholar] [CrossRef]
- Malinova, D.; Fritzsche, M.; Nowosad, C.R.; Armer, H.; Munro, P.M.G.; Blundell, M.P.; Charras, G.; Tolar, P.; Bouma, G.; Thrasher, A.J. WASp-dependent actin cytoskeleton stability at the dendritic cell immunological synapse is required for extensive, functional T cell contacts. J. Leukoc. Biol. 2016, 99, 699–710. [Google Scholar] [CrossRef]
- Nolz, J.C.; Nacusi, L.P.; Segovis, C.M.; Medeiros, R.B.; Mitchell, J.S.; Shimizu, Y.; Billadeau, D.D. The WAVE2 complex regulates T cell receptor signaling to integrins via Abl- and CrkL-C3G-mediated activation of Rap1. J. Cell Biol. 2008, 182, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, P.A.; Bunnell, S.C.; Witherow, D.S.; Gu, J.J.; Chislock, E.M.; Ring, C.; Pendergast, A.M. Role for the Abi/wave protein complex in T cell receptor-mediated proliferation and cytoskeletal remodeling. Curr. Biol. 2006, 16, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Depoil, D.; Martinelli, R.; Judokusumo, E.; Carmona, G.; Gertler, F.B.; Kam, L.C.; Carman, C.V.; Burkhardt, J.K.; Irvine, D.J.; et al. Actin foci facilitate activation of the phospholipase C-gamma in primary T lymphocytes via the WASP pathway. Elife 2015, 4, e04953. [Google Scholar] [CrossRef]
- Calvez, R.; Lafouresse, F.; De Meester, J.; Galy, A.; Valitutti, S.; Dupre, L. The Wiskott-Aldrich syndrome protein permits assembly of a focused immunological synapse enabling sustained T-cell receptor signaling. Haematologica 2011, 96, 1415–1423. [Google Scholar] [CrossRef] [Green Version]
- Tamzalit, F.; Wang, M.S.; Jin, W.; Tello-Lafoz, M.; Boyko, V.; Heddleston, J.M.; Black, C.T.; Kam, L.C.; Huse, M. Interfacial actin protrusions mechanically enhance killing by cytotoxic T cells. Sci. Immunol. 2019, 4, eaav5445. [Google Scholar] [CrossRef]
- Gorgun, G.; Holderried, T.A.; Zahrieh, D.; Neuberg, D.; Gribben, J.G. Chronic lymphocytic leukemia cells induce changes in gene expression of CD4 and CD8 T cells. J. Clin. Investig. 2005, 115, 1797–1805. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, A.G.; Clear, A.J.; Fatah, R.; Gribben, J.G. Multiple inhibitory ligands induce impaired T-cell immunologic synapse function in chronic lymphocytic leukemia that can be blocked with lenalidomide: Establishing a reversible immune evasion mechanism in human cancer. Blood 2012, 120, 1412–1421. [Google Scholar] [CrossRef]
- Ramsay, A.G.; Johnson, A.J.; Lee, A.M.; Gorgun, G.; Le Dieu, R.; Blum, W.; Byrd, J.C.; Gribben, J.G. Chronic lymphocytic leukemia T cells show impaired immunological synapse formation that can be reversed with an immunomodulating drug. J. Clin. Investig. 2008, 118, 2427–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, A.G.; Evans, R.; Kiaii, S.; Svensson, L.; Hogg, N.; Gribben, J.G. Chronic lymphocytic leukemia cells induce defective LFA-1-directed T-cell motility by altering Rho GTPase signaling that is reversible with lenalidomide. Blood 2013, 121, 2704–2714. [Google Scholar] [CrossRef] [PubMed]
- Somersalo, K.; Anikeeva, N.; Sims, T.N.; Thomas, V.K.; Strong, R.K.; Spies, T.; Lebedeva, T.; Sykulev, Y.; Dustin, M.L. Cytotoxic T lymphocytes form an antigen-independent ring junction. J. Clin. Investig. 2004, 113, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgart, F.; Arnold, A.M.; Leskovar, K.; Staszek, K.; Folser, M.; Weghuber, J.; Stockinger, H.; Schutz, G.J. Varying label density allows artifact-free analysis of membrane-protein nanoclusters. Nat. Methods 2016, 13, 661–664. [Google Scholar] [CrossRef] [PubMed]
- Lub, M.; van Kooyk, Y.; van Vliet, S.J.; Figdor, C.G. Dual role of the actin cytoskeleton in regulating cell adhesion mediated by the integrin lymphocyte function-associated molecule-1. Mol. Biol. Cell. 1997, 8, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Campi, G.; Varma, R.; Dustin, M.L. Actin and agonist MHC-peptide complex-dependent T cell receptor microclusters as scaffolds for signaling. J. Exp. Med. 2005, 202, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Comrie, W.A.; Babich, A.; Burkhardt, J.K. F-actin flow drives affinity maturation and spatial organization of LFA-1 at the immunological synapse. J. Cell Biol. 2015, 208, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Stinchcombe, J.C.; Salio, M.; Cerundolo, V.; Pende, D.; Arico, M.; Griffiths, G.M. Centriole polarisation to the immunological synapse directs secretion from cytolytic cells of both the innate and adaptive immune systems. BMC Biol. 2011, 9, 45. [Google Scholar] [CrossRef]
- de la Roche, M.; Asano, Y.; Griffiths, G.M. Origins of the cytolytic synapse. Nat. Rev. Immunol. 2016, 16, 421–432. [Google Scholar] [CrossRef]
- Gross, C.C.; Brzostowski, J.A.; Liu, D.; Long, E.O. Tethering of intercellular adhesion molecule on target cells is required for LFA-1-dependent NK cell adhesion and granule polarization. J. Immunol. 2010, 185, 2918–2926. [Google Scholar] [CrossRef]
- Tabdanov, E.; Kam, L.C.; Gondarenko, S.; Liapis, A.; Kumari, S.; Dustin, M.L.; Iskratsch, T.; Sheetz, M.P. Micropatterning of TCR and LFA-1 ligands reveals complementary effects on cytoskeleton mechanics in T cells. Integr. Biol. 2015, 7, 1272–1284. [Google Scholar] [CrossRef]
- Houmadi, R.; Guipouy, D.; Rey-Barroso, J.; Vasconcelos, Z.; Cornet, J.; Manghi, M.; Destainville, N.; Valitutti, S.; Allart, S.; Dupre, L. The Wiskott-Aldrich Syndrome Protein Contributes to the Assembly of the LFA-1 Nanocluster Belt at the Lytic Synapse. Cell Rep. 2018, 22, 979–991. [Google Scholar] [CrossRef]
- Comrie, W.A.; Li, S.; Boyle, S.; Burkhardt, J.K. The dendritic cell cytoskeleton promotes T cell adhesion and activation by constraining ICAM-1 mobility. J. Cell Biol. 2015, 208, 457–473. [Google Scholar] [CrossRef] [Green Version]
- Abouzahr, S.; Bismuth, G.; Gaudin, C.; Caroll, O.; Van Endert, P.; Jalil, A.; Dausset, J.; Vergnon, I.; Richon, C.; Kauffmann, A.; et al. Identification of target actin content and polymerization status as a mechanism of tumor resistance after cytolytic T lymphocyte pressure. Proc. Natl. Acad. Sci. USA 2006, 103, 1428–1433. [Google Scholar] [CrossRef] [Green Version]
- Ghoshdastider, U.; Popp, D.; Burtnick, L.D.; Robinson, R.C. The expanding superfamily of gelsolin homology domain proteins. Cytoskeleton 2013, 70, 775–795. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.A.; Henkemeyer, M. The SH2/SH3 adaptor Grb4 transduces B-ephrin reverse signals. Nature 2001, 413, 174–179. [Google Scholar] [CrossRef]
- Abouzahr-Rifai, S.; Hasmim, M.; Boukerche, H.; Hamelin, J.; Janji, B.; Jalil, A.; Kieda, C.; Mami-Chouaib, F.; Bertoglio, J.; Chouaib, S. Resistance of tumor cells to cytolytic T lymphocytes involves Rho-GTPases and focal adhesion kinase activation. J. Biol. Chem. 2008, 283, 31665–31672. [Google Scholar] [CrossRef]
- Guan, J.L. Role of focal adhesion kinase in integrin signaling. Int. J. Biochem. Cell Biol. 1997, 29, 1085–1096. [Google Scholar] [CrossRef]
- Swaminathan, V.; Fischer, R.S.; Waterman, C.M. The FAK-Arp2/3 interaction promotes leading edge advance and haptosensing by coupling nascent adhesions to lamellipodia actin. Mol. Biol. Cell 2016, 27, 1085–1100. [Google Scholar] [CrossRef] [PubMed]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in cancer: Mechanistic findings and clinical applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef]
- Krzewski, K.; Coligan, J.E. Human NK cell lytic granules and regulation of their exocytosis. Front. Immunol. 2012, 3, 335. [Google Scholar] [CrossRef]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef]
- Barda-Saad, M.; Braiman, A.; Titerence, R.; Bunnell, S.C.; Barr, V.A.; Samelson, L.E. Dynamic molecular interactions linking the T cell antigen receptor to the actin cytoskeleton. Nat. Immunol. 2005, 6, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Martín, N.; Fernández-Arenas, E.; Cemerski, S.; Delgado, P.; Turner, M.; Heuser, J.; Irvine, D.J.; Huang, B.; Bustelo, X.R.; Shaw, A.; et al. T cell receptor internalization from the immunological synapse is mediated by TC21 and RhoG GTPase-dependent phagocytosis. Immunity 2011, 35, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Vardhana, S.; Cammer, M.; Curado, S.; Santos, L.; Sheetz, M.; Dustin, M. T Lymphocyte Myosin IIA is Required for Maturation of the Immunological Synapse. Front. Immunol. 2012, 3, 230. [Google Scholar] [CrossRef]
- Ilani, T.; Vasiliver-Shamis, G.; Vardhana, S.; Bretscher, A.; Dustin, M.L. T cell antigen receptor signaling and immunological synapse stability require myosin IIA. Nat. Immunol. 2009, 10, 531. [Google Scholar] [CrossRef]
- Hammer, J.A.; Burkhardt, J.K. Controversy and consensus regarding myosin II function at the immunological synapse. Curr. Opin. Immunol. 2013, 25, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Fooksman, D.R.; Vardhana, S.; Vasiliver-Shamis, G.; Liese, J.; Blair, D.A.; Waite, J.; Sacristán, C.; Victora, G.D.; Zanin-Zhorov, A.; Dustin, M.L. Functional anatomy of T cell activation and synapse formation. Annu. Rev. Immunol. 2010, 28, 79–105. [Google Scholar] [CrossRef]
- Wurzer, H.; Hoffmann, C.; Al Absi, A.; Thomas, C. Do tumor cells escape from natural killer cell cytotoxicity by mimicking dendritic cells? Oncotarget 2019, 10, 2419–2420. [Google Scholar] [CrossRef]
- Husson, J.; Chemin, K.; Bohineust, A.; Hivroz, C.; Henry, N. Force generation upon T cell receptor engagement. PLoS ONE 2011, 6, e19680. [Google Scholar] [CrossRef]
- Comrie, W.A.; Burkhardt, J.K. Action and Traction: Cytoskeletal Control of Receptor Triggering at the Immunological Synapse. Front. Immunol. 2016, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Basu, R.; Huse, M. Mechanical Communication at the Immunological Synapse. Trends Cell Biol. 2017, 27, 241–254. [Google Scholar] [CrossRef]
- Roy, N.H.; Burkhardt, J.K. The Actin Cytoskeleton: A Mechanical Intermediate for Signal Integration at the Immunological Synapse. Front. Cell Dev. Biol. 2018, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.T.; Takeuchi, K.; Sun, Z.Y.; Touma, M.; Castro, C.E.; Fahmy, A.; Lang, M.J.; Wagner, G.; Reinherz, E.L. The alphabeta T cell receptor is an anisotropic mechanosensor. J. Biol. Chem. 2009, 284, 31028–31037. [Google Scholar] [CrossRef]
- Brazin, K.N.; Mallis, R.J.; Das, D.K.; Feng, Y.; Hwang, W.; Wang, J.H.; Wagner, G.; Lang, M.J.; Reinherz, E.L. Structural Features of the alphabetaTCR Mechanotransduction Apparatus That Promote pMHC Discrimination. Front. Immunol. 2015, 6, 441. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, B.H.; Louban, I.; Djandji, D.; Wabnitz, G.H.; Deeg, J.; Bulbuc, N.; Samstag, Y.; Gunzer, M.; Spatz, J.P.; Hammerling, G.J. Immune synapse formation determines interaction forces between T cells and antigen-presenting cells measured by atomic force microscopy. Proc. Natl. Acad. Sci. USA 2009, 106, 17852–17857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, T.S.; Mortellaro, A.; Lim, C.T.; Hammerling, G.J.; Ricciardi-Castagnoli, P. Mechanical interactions between dendritic cells and T cells correlate with T cell responsiveness. J. Immunol. 2011, 187, 258–265. [Google Scholar] [CrossRef]
- Hivroz, C.; Saitakis, M. Biophysical Aspects of T Lymphocyte Activation at the Immune Synapse. Front. Immunol. 2016, 7, 46. [Google Scholar] [CrossRef]
- Kong, F.; Garcia, A.J.; Mould, A.P.; Humphries, M.J.; Zhu, C. Demonstration of catch bonds between an integrin and its ligand. J. Cell Biol. 2009, 185, 1275–1284. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Lou, J.; Zhu, C. Forcing switch from short- to intermediate- and long-lived states of the alphaA domain generates LFA-1/ICAM-1 catch bonds. J. Biol. Chem. 2010, 285, 35967–35978. [Google Scholar] [CrossRef]
- Basu, R.; Whitlock, B.M.; Husson, J.; Le Floc’h, A.; Jin, W.; Oyler-Yaniv, A.; Dotiwala, F.; Giannone, G.; Hivroz, C.; Biais, N.; et al. Cytotoxic T Cells Use Mechanical Force to Potentiate Target Cell Killing. Cell 2016, 165, 100–110. [Google Scholar] [CrossRef]
- Mattila, P.K.; Batista, F.D.; Treanor, B. Dynamics of the actin cytoskeleton mediates receptor cross talk: An emerging concept in tuning receptor signaling. J. Cell Biol. 2016, 212, 267–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, A.; Hordijk, P.L. Cell-stiffness-induced mechanosignaling—A key driver of leukocyte transendothelial migration. J. Cell Sci. 2015, 128, 2221–2230. [Google Scholar] [CrossRef] [PubMed]
- Bufi, N.; Saitakis, M.; Dogniaux, S.; Buschinger, O.; Bohineust, A.; Richert, A.; Maurin, M.; Hivroz, C.; Asnacios, A. Human Primary Immune Cells Exhibit Distinct Mechanical Properties that Are Modified by Inflammation. Biophys. J. 2015, 108, 2181–2190. [Google Scholar] [CrossRef] [Green Version]
- Bouma, G.; Mendoza-Naranjo, A.; Blundell, M.P.; de Falco, E.; Parsley, K.L.; Burns, S.O.; Thrasher, A.J. Cytoskeletal remodeling mediated by WASp in dendritic cells is necessary for normal immune synapse formation and T-cell priming. Blood 2011, 118, 2492–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Alwan, M.M.; Rowden, G.; Lee, T.D.; West, K.A. The dendritic cell cytoskeleton is critical for the formation of the immunological synapse. J. Immunol. 2001, 166, 1452–1456. [Google Scholar] [CrossRef]
- O’Connor, R.S.; Hao, X.; Shen, K.; Bashour, K.; Akimova, T.; Hancock, W.W.; Kam, L.C.; Milone, M.C. Substrate rigidity regulates human T cell activation and proliferation. J. Immunol. 2012, 189, 1330–1339. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.R.; Poenie, M. Dynamic polarization of the microtubule cytoskeleton during CTL-mediated killing. Immunity 2002, 16, 111–121. [Google Scholar] [CrossRef]
- Mallik, R.; Gross, S.P. Molecular motors: Strategies to get along. Curr. Biol. 2004, 14, R971–R982. [Google Scholar] [CrossRef]
- Ritter, A.T.; Asano, Y.; Stinchcombe, J.C.; Dieckmann, N.M.; Chen, B.C.; Gawden-Bone, C.; van Engelenburg, S.; Legant, W.; Gao, L.; Davidson, M.W.; et al. Actin depletion initiates events leading to granule secretion at the immunological synapse. Immunity 2015, 42, 864–876. [Google Scholar] [CrossRef]
- Trapani, J.A. Target cell apoptosis induced by cytotoxic T cells and natural killer cells involves synergy between the pore-forming protein, perforin, and the serine protease, granzyme B. Aust. N. Z. J. Med. 1995, 25, 793–799. [Google Scholar] [CrossRef]
- Bots, M.; Medema, J.P. Granzymes at a glance. J. Cell Sci. 2006, 119, 5011–5014. [Google Scholar] [CrossRef] [Green Version]
- Gomez, T.S.; Kumar, K.; Medeiros, R.B.; Shimizu, Y.; Leibson, P.J.; Billadeau, D.D. Formins regulate the actin-related protein 2/3 complex-independent polarization of the centrosome to the immunological synapse. Immunity 2007, 26, 177–190. [Google Scholar] [CrossRef]
- Andres-Delgado, L.; Anton, O.M.; Alonso, M.A. Centrosome polarization in T cells: A task for formins. Front. Immunol. 2013, 4, 191. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.T.; Carisey, A.F.; Orange, J.S. Measurement of Lytic Granule Convergence After Formation of an NK Cell Immunological Synapse. Methods Mol. Biol. 2017, 1584, 497–515. [Google Scholar] [Green Version]
- Wen, P.J.; Grenklo, S.; Arpino, G.; Tan, X.; Liao, H.S.; Heureaux, J.; Peng, S.Y.; Chiang, H.C.; Hamid, E.; Zhao, W.D.; et al. Actin dynamics provides membrane tension to merge fusing vesicles into the plasma membrane. Nat. Commun. 2016, 7, 12604. [Google Scholar] [CrossRef] [Green Version]
- Sanborn, K.B.; Rak, G.D.; Maru, S.Y.; Demers, K.; Difeo, A.; Martignetti, J.A.; Betts, M.R.; Favier, R.; Banerjee, P.P.; Orange, J.S. Myosin IIA Associates with NK Cell Lytic Granules to Enable Their Interaction with F-Actin and Function at the Immunological Synapse. J. Immunol. 2009, 182, 6969. [Google Scholar] [CrossRef]
- Andzelm, M.M.; Chen, X.; Krzewski, K.; Orange, J.S.; Strominger, J.L. Myosin IIA is required for cytolytic granule exocytosis in human NK cells. J. Exp. Med. 2007, 204, 2285–2291. [Google Scholar] [CrossRef] [Green Version]
- Krzewski, K.; Chen, X.; Orange, J.S.; Strominger, J.L. Formation of a WIP-, WASp-, actin-, and myosin IIA-containing multiprotein complex in activated NK cells and its alteration by KIR inhibitory signaling. J. Cell Biol. 2006, 173, 121–132. [Google Scholar] [CrossRef]
- Iizuka, Y.; Cichocki, F.; Sieben, A.; Sforza, F.; Karim, R.; Coughlin, K.; Isaksson Vogel, R.; Gavioli, R.; McCullar, V.; Lenvik, T.; et al. UNC-45A Is a Nonmuscle Myosin IIA Chaperone Required for NK Cell Cytotoxicity via Control of Lytic Granule Secretion. J. Immunol. 2015, 195, 4760. [Google Scholar] [CrossRef]
- Ritter, A.T.; Kapnick, S.M.; Murugesan, S.; Schwartzberg, P.L.; Griffiths, G.M.; Lippincott-Schwartz, J. Cortical actin recovery at the immunological synapse leads to termination of lytic granule secretion in cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. USA 2017, 114, E6585–E6594. [Google Scholar] [CrossRef] [Green Version]
- Stanton, R.J.; Prod’homme, V.; Purbhoo, M.A.; Moore, M.; Aicheler, R.J.; Heinzmann, M.; Bailer, S.M.; Haas, J.; Antrobus, R.; Weekes, M.P.; et al. HCMV pUL135 remodels the actin cytoskeleton to impair immune recognition of infected cells. Cell Host Microbe 2014, 16, 201–214. [Google Scholar] [CrossRef]
- Marques, C.A.; Hahnel, P.S.; Wolfel, C.; Thaler, S.; Huber, C.; Theobald, M.; Schuler, M. An immune escape screen reveals Cdc42 as regulator of cancer susceptibility to lymphocyte-mediated tumor suppression. Blood 2008, 111, 1413–1419. [Google Scholar] [CrossRef] [PubMed]
- Haynes, J.; Srivastava, J.; Madson, N.; Wittmann, T.; Barber, D.L. Dynamic actin remodeling during epithelial-mesenchymal transition depends on increased moesin expression. Mol. Biol Cell 2011, 22, 4750–4764. [Google Scholar] [CrossRef] [PubMed]
- Dave, B.; Mittal, V.; Tan, N.M.; Chang, J.C. Epithelial-mesenchymal transition, cancer stem cells and treatment resistance. Breast Cancer Res. 2012, 14, 202. [Google Scholar] [CrossRef]
- Heerboth, S.; Housman, G.; Leary, M.; Longacre, M.; Byler, S.; Lapinska, K.; Willbanks, A.; Sarkar, S. EMT and tumor metastasis. Clin. Transl. Med. 2015, 4, 6. [Google Scholar] [CrossRef]
- Kim, S.; Koh, J.; Kim, M.Y.; Kwon, D.; Go, H.; Kim, Y.A.; Jeon, Y.K.; Chung, D.H. PD-L1 expression is associated with epithelial-to-mesenchymal transition in adenocarcinoma of the lung. Hum. Pathol. 2016, 58, 7–14. [Google Scholar] [CrossRef]
- Zheng, X.; Carstens, J.L.; Kim, J.; Scheible, M.; Kaye, J.; Sugimoto, H.; Wu, C.C.; LeBleu, V.S.; Kalluri, R. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature 2015, 527, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Terry, S.; Buart, S.; Tan, T.Z.; Gros, G.; Noman, M.Z.; Lorens, J.B.; Mami-Chouaib, F.; Thiery, J.P.; Chouaib, S. Acquisition of tumor cell phenotypic diversity along the EMT spectrum under hypoxic pressure: Consequences on susceptibility to cell-mediated cytotoxicity. Oncoimmunology 2017, 6, e1271858. [Google Scholar] [CrossRef] [Green Version]
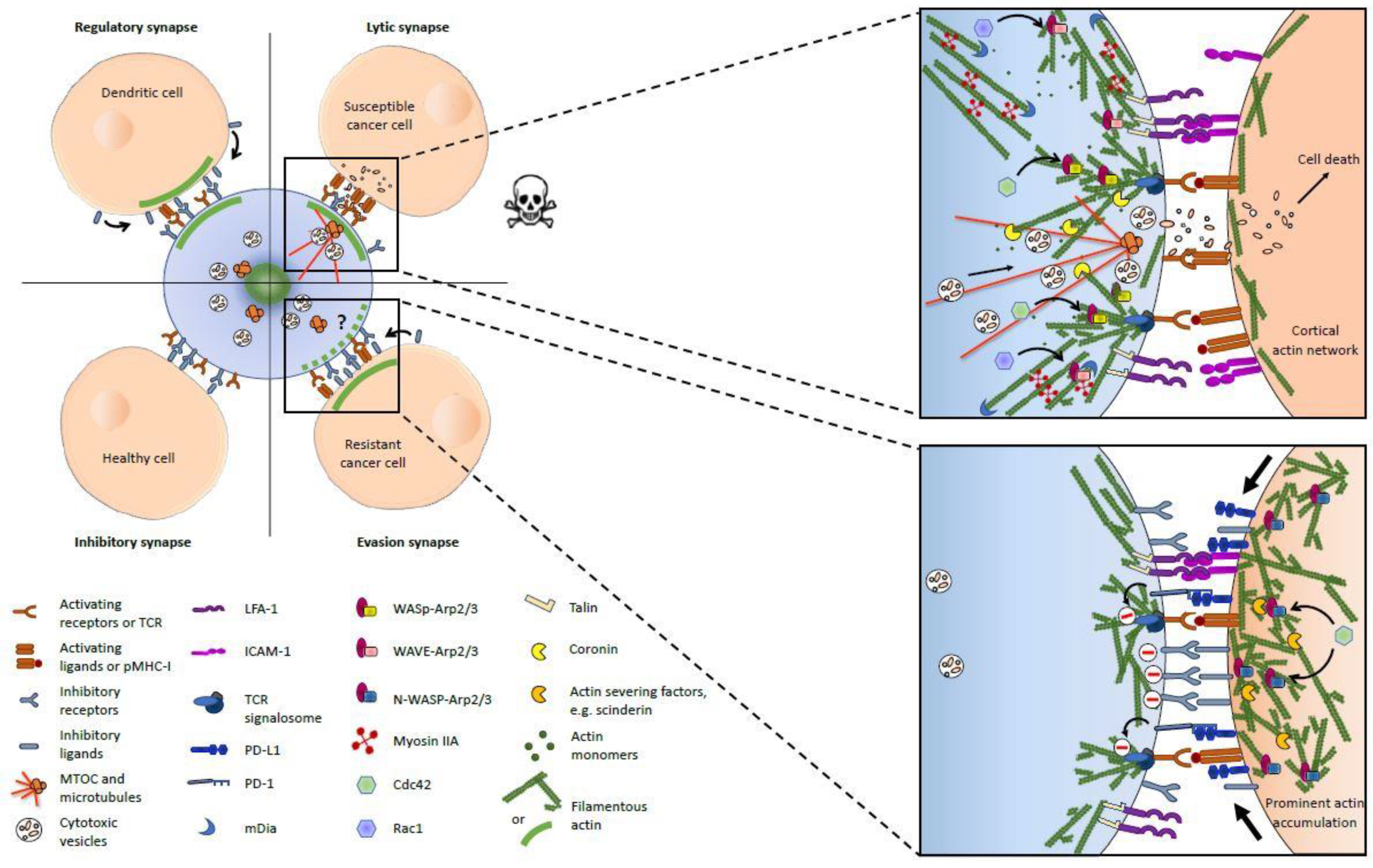
Figure 1.
Actin cytoskeleton at the immunological synapses between a cytotoxic lymphocyte (blue) and its target cells. The left panel provides a general overview of the four types of immunological synapses (ISs) discussed in the article and shows the presence or absence of F-actin accumulation (large green line) on both sides of the IS (inspired from [45]). The specific actin filaments (AF) organization and some key upstream regulators, as well as the functions of AFs in intercellular signaling are depicted in the right panels for the lytic (upper panel) and evasion (lower panel) synapses. The reader should pay attention that the provided information sometimes originates exclusively from studies based on either natural killer (NK) cells or cytotoxic T lymphocytes (CTLs) and is invited to refer to the main text for more details.
Figure 1.
Actin cytoskeleton at the immunological synapses between a cytotoxic lymphocyte (blue) and its target cells. The left panel provides a general overview of the four types of immunological synapses (ISs) discussed in the article and shows the presence or absence of F-actin accumulation (large green line) on both sides of the IS (inspired from [45]). The specific actin filaments (AF) organization and some key upstream regulators, as well as the functions of AFs in intercellular signaling are depicted in the right panels for the lytic (upper panel) and evasion (lower panel) synapses. The reader should pay attention that the provided information sometimes originates exclusively from studies based on either natural killer (NK) cells or cytotoxic T lymphocytes (CTLs) and is invited to refer to the main text for more details.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wurzer, H.; Hoffmann, C.; Al Absi, A.; Thomas, C. Actin Cytoskeleton Straddling the Immunological Synapse between Cytotoxic Lymphocytes and Cancer Cells. Cells 2019, 8, 463. https://doi.org/10.3390/cells8050463
AMA Style
Wurzer H, Hoffmann C, Al Absi A, Thomas C. Actin Cytoskeleton Straddling the Immunological Synapse between Cytotoxic Lymphocytes and Cancer Cells. Cells. 2019; 8(5):463. https://doi.org/10.3390/cells8050463
Chicago/Turabian StyleWurzer, Hannah, Céline Hoffmann, Antoun Al Absi, and Clément Thomas. 2019. "Actin Cytoskeleton Straddling the Immunological Synapse between Cytotoxic Lymphocytes and Cancer Cells" Cells 8, no. 5: 463. https://doi.org/10.3390/cells8050463
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.