
Intracellular Mono-ADP-Ribosylation in Signaling and Disease


Abstract
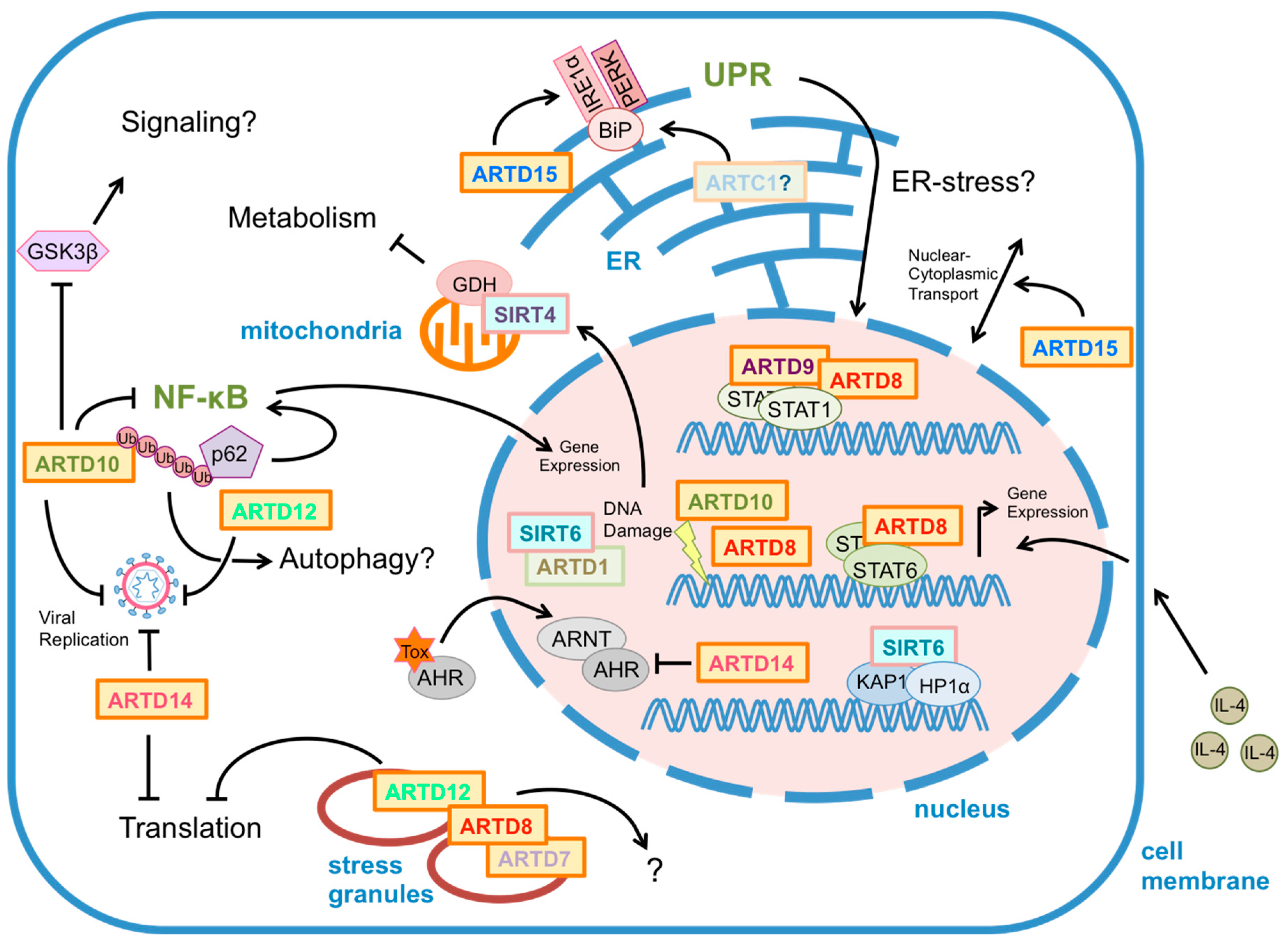
:1. Introduction

{kind=link}
| Protein | Enzymatic activity | Cellular process/ pathway | Disease-related function |
|---|---|---|---|
| ARTD3 | MAR | DNA repair [11] | Not reported |
| ARTD4 (PARP4, vPARP) | MAR | Vault particles [12] | Multidrug resistance? [13,14,15,16] Innate immunity? [13] |
| ARTD7 (PARP15, BAL3) | MAR | Component of stress granules [17] | Not reported |
| ARTD8 (PARP14, BAL2, CoaSt6) | MAR | IL-4/STAT6 signaling [18,19,20] JNK signaling [21] TH cell differentiation [22,23,24] Component of stress granules [17] | Promotes cell survival in B cells/multiple myeloma and lymphomagenesis [21,25,26] Allergic airway disease [22] |
| ARTD9 (PARP9) | inactive | IFNγ/STAT1 signaling [27,28,29] | Risk factor in DLBCL [27,28] Cell survival and chemoresistance [27,28,29] |
| ARTD10 (PARP10) | MAR | NF-κB signaling [30] Translation [31] Autophagy? [32] GSK3β signaling? [33] | Regulation of apoptosis [34] Inflammation? [30] IFN-stimulated gene, inhibition of viral replication [31] |
| ARTD12 (PARP12) | MAR | Translation [31] NF-κB signaling? [35] Autophagy? [35] Component of stress granules [17] | IFN-stimulated gene, inhibition of viral replication [31] |
| ARTD13 (PARP13, ZAP, ZC3HAV1) | inactive | mRNA regulation and miRNA silencing [36] Component of stress granules [17] | Viral defense [36] |
| ARTD14 (PARP7, TiPARP) | MAR | AHR signaling [37,38,39,40] Translation [31] | TCCD-induced hepatotoxicity [40] Inhibition of viral replication [31,41] |
| ARTD15 (PARP16) | MAR | Unfolded protein response [42] | Not reported |
| ARTD16 (PARP8) | MAR | Unclear | Not reported |
| ARTD17 (PARP6) | MAR | Regulates cell cycle progression [43] | Inhibits cell proliferation, survival benefit in colorectal cancer [43] |
| SIRT4 | MAR | Glutamine metabolism [44,45] | Tumor-suppressive [45,46] |
| SIRT6 | MAR | DNA repair [47,48,49] Retrotransposon silencing [50] | Tumor suppressive [50,51,52,53,54,55,56,57] and oncogenic functions [58,59] |
| MACROD1 (LRP16) | Hydrolase | ERα signaling [60,61] AR signaling [62] | Promotes cell proliferation [60,62] Metastasis, invasion and survival in gastric and colorectal cancer [63,64]. |
| MACROD2 (C20ORF133) | Hydrolase | Unclear | Tamoxifen-resistance in breast cancer cell lines [65] |
| TARG1 (C6ORF130, OARD1) | Hydrolase | DNA repair? [66] | Neurodegeneration [66] |
2. Regulation of Signaling Pathways and Gene Expression by MARylation
2.1. ARTD8 Regulates IL4/STAT6 Signaling
2.2. ARTD10 Modulates GSK3β Kinase Activity and NF-κB Signaling

2.3. ARTD14 Regulates AHR Signaling
2.4. MARylation in the Unfolded Protein Response
2.5. MARylation in Response to Genotoxic Stress
3. MARylation in Disease
3.1. Cancer and Metabolism
3.2. Innate Immunity and Inflammation
3.2.1. Mono-ARTDs in the Defense against Viral Infections
3.2.2. Bacterial mono-ADP-ribosyltransferases
3.3.3. ARTD8 Modulates TH-Cell Differentiation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Feijs, K.L.; Verheugd, P.; Luscher, B. Expanding functions of intracellular resident mono-ADP-ribosylation in cell physiology. Febs J. 2013, 280, 3519–3529. [Google Scholar] [CrossRef]
- Feldman, J.L.; Dittenhafer-Reed, K.E.; Denu, J.M. Sirtuin catalysis and regulation. J. Biol. Chem. 2012, 287, 42419–42427. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.A.; Kraus, W.L. New insights into the molecular and cellular functions of poly(ADP-ribose) and PARPS. Nat. Rev. Mol. Cell. Biol. 2012, 13, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Glowacki, G.; Braren, R.; Firner, K.; Nissen, M.; Kuhl, M.; Reche, P.; Bazan, F.; Cetkovic-Cvrlje, M.; Leiter, E.; Haag, F.; et al. The family of toxin-related ecto-ADP-ribosyltransferases in humans and the mouse. Protein Sci. 2002, 11, 1657–1670. [Google Scholar] [CrossRef] [PubMed]
- Seman, M.; Adriouch, S.; Haag, F.; Koch-Nolte, F. Ecto-ADP-ribosyltransferases (arts): Emerging actors in cell communication and signaling. Curr Med. Chem. 2004, 11, 857–872. [Google Scholar] [CrossRef] [PubMed]
- Kleine, H.; Poreba, E.; Lesniewicz, K.; Hassa, P.O.; Hottiger, M.O.; Litchfield, D.W.; Shilton, B.H.; Luscher, B. Substrate-assisted catalysis by parp10 limits its activity to mono-ADP-ribosylation. Mol. Cell 2008, 32, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Hottiger, M.O.; Hassa, P.O.; Luscher, B.; Schuler, H.; Koch-Nolte, F. Toward a unified nomenclature for mammalian ADP-ribosyltransferases. Trends Biochem. Sci. 2010, 35, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Karlberg, T.; Klepsch, M.; Thorsell, A.G.; Andersson, C.D.; Linusson, A.; Schuler, H. Structural basis for lack of ADP-ribosyltransferase activity in poly(ADP-ribose) polymerase-13/zinc finger antiviral protein. J. Biol. Chem. 2015, 290, 7336–7344. [Google Scholar] [CrossRef] [PubMed]
- Loseva, O.; Jemth, A.S.; Bryant, H.E.; Schuler, H.; Lehtio, L.; Karlberg, T.; Helleday, T. Parp-3 is a mono-ADP-ribosylase that activates PARP-1 in the absence of DNA. J. Biol. Chem. 2010, 285, 8054–8060. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Matic, I.; Uchima, L.; Rood, J.; Zaja, R.; Hay, R.T.; Ahel, I.; Chang, P. Family-wide analysis of poly(ADP-ribose) polymerase activity. Nat. Commun. 2014, 5, 4426. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Robert, I.; Reina-San-Martin, B.; Schreiber, V.; Dantzer, F. Poly(ADP-ribose) polymerases in double-strand break repair: Focus on PARP1, PARP2 and PARP3. Exp. Cell. Res. 2014, 329, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Kickhoefer, V.A.; Siva, A.C.; Kedersha, N.L.; Inman, E.M.; Ruland, C.; Streuli, M.; Rome, L.H. The 193-kd vault protein, vparp, is a novel poly(ADP-ribose) polymerase. J. Cell. Biol. 1999, 146, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Berger, W.; Steiner, E.; Grusch, M.; Elbling, L.; Micksche, M. Vaults and the major vault protein: Novel roles in signal pathway regulation and immunity. Cell. Mol. Life Sci. 2009, 66, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Mossink, M.H.; van Zon, A.; Scheper, R.J.; Sonneveld, P.; Wiemer, E.A. Vaults: A ribonucleoprotein particle involved in drug resistance? Oncogene 2003, 22, 7458–7467. [Google Scholar] [CrossRef] [PubMed]
- Siva, A.C.; Raval-Fernandes, S.; Stephen, A.G.; LaFemina, M.J.; Scheper, R.J.; Kickhoefer, V.A.; Rome, L.H. Up-regulation of vaults may be necessary but not sufficient for multidrug resistance. Int. J. Cancer 2001, 92, 195–202. [Google Scholar] [CrossRef]
- Schroeijers, A.B.; Siva, A.C.; Scheffer, G.L.; de Jong, M.C.; Bolick, S.C.; Dukers, D.F.; Slootstra, J.W.; Meloen, R.H.; Wiemer, E.; Kickhoefer, V.A.; et al. The mr 193,000 vault protein is up-regulated in multidrug-resistant cancer cell lines. Cancer Res. 2000, 60, 1104–1110. [Google Scholar] [PubMed]
- Leung, A.K.; Vyas, S.; Rood, J.E.; Bhutkar, A.; Sharp, P.A.; Chang, P. Poly(ADP-ribose) regulates stress responses and microrna activity in the cytoplasm. Mol. Cell 2011, 42, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Goenka, S.; Boothby, M. Selective potentiation of Stat-dependent gene expression by collaborator of Stat6 (CoaSt6), a transcriptional cofactor. Proc. Natl. Acad. Sci. USA 2006, 103, 4210–4215. [Google Scholar] [CrossRef] [PubMed]
- Goenka, S.; Cho, S.H.; Boothby, M. Collaborator of stat6 (CoaSt6)-associated poly(ADP-ribose) polymerase activity modulates Stat6-dependent gene transcription. J. Biol. Chem. 2007, 282, 18732–18739. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, P.; Riley, J.P.; Patel, R.; Li, F.; Voss, L.; Goenka, S. Parp-14 functions as a transcriptional switch for Stat6-dependent gene activation. J. Biol. Chem. 2011, 286, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Barbarulo, A.; Iansante, V.; Chaidos, A.; Naresh, K.; Rahemtulla, A.; Franzoso, G.; Karadimitris, A.; Haskard, D.O.; Papa, S.; Bubici, C. Poly(ADP-ribose) polymerase family member 14 (parp14) is a novel effector of the Jnk2-dependent pro-survival signal in multiple myeloma. Oncogene 2013, 32, 4231–4242. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, P.; Hollenbeck, A.; Riley, J.P.; Li, F.; Patel, R.J.; Akhtar, N.; Goenka, S. Poly (ADP-ribose) polymerase 14 and its enzyme activity regulates T(H)2 differentiation and allergic airway disease. J. Allergy. Clin. Immunol. 2013, 131, 521–531 e12. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Raybuck, A.; Wei, M.; Erickson, J.; Nam, K.T.; Cox, R.G.; Trochtenberg, A.; Thomas, J.W.; Williams, J.; Boothby, M. B cell-intrinsic and -extrinsic regulation of antibody responses by parp14, an intracellular (ADP-ribosyl)transferase. J. Immunol. 2013, 191, 3169–3178. [Google Scholar] [CrossRef] [PubMed]
- Riley, J.P.; Kulkarni, A.; Mehrotra, P.; Koh, B.; Perumal, N.B.; Kaplan, M.H.; Goenka, S. Parp-14 binds specific DNA sequences to promote TH2 cell gene expression. PLoS One 2013, 8, e83127. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Goenka, S.; Henttinen, T.; Gudapati, P.; Reinikainen, A.; Eischen, C.M.; Lahesmaa, R.; Boothby, M. PARP-14, a member of the b aggressive lymphoma family, transduces survival signals in primary b cells. Blood 2009, 113, 2416–2425. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Ahn, A.K.; Bhargava, P.; Lee, C.H.; Eischen, C.M.; McGuinness, O.; Boothby, M. Glycolytic rate and lymphomagenesis depend on PARP14, an adp ribosyltransferase of the B aggressive lymphoma (Bal) family. Proc. Natl. Acad. Sci. USA 2011, 108, 15972–15977. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, R.C.; Yakushijin, Y.; Kharbanda, S.; Salgia, R.; Fletcher, J.A.; Shipp, M.A. Bal is a novel risk-related gene in diffuse large B-cell lymphomas that enhances cellular migration. Blood 2000, 96, 4328–4334. [Google Scholar] [PubMed]
- Camicia, R.; Bachmann, S.B.; Winkler, H.C.; Beer, M.; Tinguely, M.; Haralambieva, E.; Hassa, P.O. Bal1/ARTD9 represses the anti-proliferative and pro-apoptotic ifngamma-STAT1-IRF1-p53 axis in diffuse large B-cell lymphoma. J. Cell. Sci. 2013, 126, 1969–1980. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, S.B.; Frommel, S.C.; Camicia, R.; Winkler, H.C.; Santoro, R.; Hassa, P.O. DTX3l and ARTD9 inhibit IRF1 expression and mediate in cooperation with ARTD8 survival and proliferation of metastatic prostate cancer cells. Mol. Cancer 2014, 13, 1–24. [Google Scholar]
- Verheugd, P.; Forst, A.H.; Milke, L.; Herzog, N.; Feijs, K.L.; Kremmer, E.; Kleine, H.; Luscher, B. Regulation of NF-kappab signalling by the mono-ADP-ribosyltransferase artd10. Nat. Commun. 2013, 4, 1683. [Google Scholar] [CrossRef] [PubMed]
- Atasheva, S.; Frolova, E.I.; Frolov, I. Interferon-stimulated poly(ADP-ribose) polymerases are potent inhibitors of cellular translation and virus replication. J. Virol. 2014, 88, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Kleine, H.; Herrmann, A.; Lamark, T.; Forst, A.H.; Verheugd, P.; Luscher-Firzlaff, J.; Lippok, B.; Feijs, K.L.; Herzog, N.; Kremmer, E.; et al. Dynamic subcellular localization of the mono-ADP-ribosyltransferase ARTD10 and interaction with the ubiquitin receptor p62. Cell. Commun. Signal. 2012, 10, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feijs, K.L.; Kleine, H.; Braczynski, A.; Forst, A.H.; Herzog, N.; Verheugd, P.; Linzen, U.; Kremmer, E.; Luscher, B. ARTD10 substrate identification on protein microarrays: Regulation of gsk3beta by mono-ADP-ribosylation. Cell. Commun. Signal. 2013, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, N.; Hartkamp, J.D.; Verheugd, P.; Treude, F.; Forst, A.H.; Feijs, K.L.; Lippok, B.E.; Kremmer, E.; Kleine, H.; Luscher, B. Caspase-dependent cleavage of the mono-ADP-ribosyltransferase artd10 interferes with its pro-apoptotic function. Febs J. 2013, 280, 1330–1343. [Google Scholar] [CrossRef] [PubMed]
- Welsby, I.; Hutin, D.; Gueydan, C.; Kruys, V.; Rongvaux, A.; Leo, O. Parp12, an interferon-stimulated gene involved in the control of protein translation and inflammation. J. Biol. Chem. 2014, 289, 26642–26657. [Google Scholar] [CrossRef] [PubMed]
- Bock, F.J.; Todorova, T.T.; Chang, P. Rna regulation by poly(ADP-ribose) polymerases. Mol. Cell 2015, 58, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Baldwin, K.T.; Renzelli, A.J.; McDaniel, A.; Dong, L. Tcdd-inducible poly(ADP-ribose) polymerase: A novel response to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Biochem. Biophys. Res. Commun. 2001, 289, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Induction and superinduction of 2,3,7,8-tetrachlorodibenzo-rho-dioxin-inducible poly(adp-ribose) polymerase: Role of the aryl hydrocarbon receptor/aryl hydrocarbon receptor nuclear translocator transcription activation domains and a labile transcription repressor. Arch. Biochem. Biophys. 2002, 404, 309–316. [Google Scholar] [PubMed]
- MacPherson, L.; Tamblyn, L.; Rajendra, S.; Bralha, F.; McPherson, J.P.; Matthews, J. 2,3,7,8-Tetrachlorodibenzo-p-dioxin poly(ADP-ribose) polymerase (TIPARP, ARTD14) is a mono-ADP-ribosyltransferase and repressor of aryl hydrocarbon receptor transactivation. Nucleic Acids Res. 2013, 41, 1604–1621. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Bott, D.; Gomez, A.; Tamblyn, L.; Rasheed, A.; MacPherson, L.; Sugamori, K.S.; Cho, T.; Yang, Y.; Grant, D.M.; et al. Loss of the mono-adp-ribosyltransferase, tiparp, increases sensitivity to dioxin-induced steatohepatitis and lethality. J. Biol. Chem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Atasheva, S.; Akhrymuk, M.; Frolova, E.I.; Frolov, I. New parp gene with an anti-alphavirus function. J. Virol. 2012, 86, 8147–8160. [Google Scholar] [CrossRef] [PubMed]
- Jwa, M.; Chang, P. Parp16 is a tail-anchored endoplasmic reticulum protein required for the perk- and ire1alpha-mediated unfolded protein response. Nat. Cell. Biol. 2012, 14, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Tuncel, H.; Tanaka, S.; Oka, S.; Nakai, S.; Fukutomi, R.; Okamoto, M.; Ota, T.; Kaneko, H.; Tatsuka, M.; Shimamoto, F. PARP6, a mono(ADP-ribosyl) transferase and a negative regulator of cell proliferation, is involved in colorectal cancer development. Int J. Oncol. 2012, 41, 2079–2086. [Google Scholar] [PubMed]
- Haigis, M.C.; Mostoslavsky, R.; Haigis, K.M.; Fahie, K.; Christodoulou, D.C.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.D.; Karow, M.; Blander, G.; et al. SIRT4 inhibits glutamate dehydrogenase and opposes the effects of calorie restriction in pancreatic beta cells. Cell 2006, 126, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.M.; Xiao, C.; Finley, L.W.; Lahusen, T.; Souza, A.L.; Pierce, K.; Li, Y.H.; Wang, X.; Laurent, G.; German, N.J.; et al. SIRT4 has tumor-suppressive activity and regulates the cellular metabolic response to DNA damage by inhibiting mitochondrial glutamine metabolism. Cancer Cell. 2013, 23, 450–463. [Google Scholar] [CrossRef] [PubMed]
- Csibi, A.; Fendt, S.M.; Li, C.; Poulogiannis, G.; Choo, A.Y.; Chapski, D.J.; Jeong, S.M.; Dempsey, J.M.; Parkhitko, A.; Morrison, T.; et al. The mtorc1 pathway stimulates glutamine metabolism and cell proliferation by repressing SIRT4. Cell 2013, 153, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Hine, C.; Tian, X.; Van Meter, M.; Au, M.; Vaidya, A.; Seluanov, A.; Gorbunova, V. SIRT6 promotes DNA repair under stress by activating parp1. Science 2011, 332, 1443–1446. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Tian, X.; Van Meter, M.; Ke, Z.; Gorbunova, V.; Seluanov, A. Sirtuin 6 (SIRT6) rescues the decline of homologous recombination repair during replicative senescence. Proc. Natl. Acad. Sci. USA 2012, 109, 11800–11805. [Google Scholar] [CrossRef] [PubMed]
- McCord, R.A.; Michishita, E.; Hong, T.; Berber, E.; Boxer, L.D.; Kusumoto, R.; Guan, S.; Shi, X.; Gozani, O.; Burlingame, A.L.; et al. SIRT6 stabilizes DNA-dependent protein kinase at chromatin for DNA double-strand break repair. Aging (Albany NY) 2009, 1, 109–121. [Google Scholar] [PubMed]
- Van Meter, M.; Kashyap, M.; Rezazadeh, S.; Geneva, A.J.; Morello, T.D.; Seluanov, A.; Gorbunova, V. Sirt6 represses line1 retrotransposons by ribosylating kap1 but this repression fails with stress and age. Nat. Commun. 2014, 5, 5011. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yin, X.J.; Xu, C.J.; Ning, Y.X.; Chen, M.; Zhang, H.; Chen, S.F.; Yao, L.Q. The histone deacetylase SIRT6 inhibits ovarian cancer cell proliferation via down-regulation of notch 3 expression. Eur Rev. Med. Pharmacol. Sci. 2015, 19, 818–824. [Google Scholar] [PubMed]
- Fukuda, T.; Wada-Hiraike, O.; Oda, K.; Tanikawa, M.; Makii, C.; Inaba, K.; Miyasaka, A.; Miyamoto, Y.; Yano, T.; Maeda, D.; et al. Putative tumor suppression function of SIRT6 in endometrial cancer. FEBS Lett. 2015, 589, 2274–2281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Qin, C.Y. Sirt6 suppresses hepatocellular carcinoma cell growth via inhibiting the extracellular signalregulated kinase signaling pathway. Mol. Med. Rep. 2014, 9, 882–888. [Google Scholar] [PubMed]
- Marquardt, J.U.; Fischer, K.; Baus, K.; Kashyap, A.; Ma, S.; Krupp, M.; Linke, M.; Teufel, A.; Zechner, U.; Strand, D.; et al. Sirtuin-6-dependent genetic and epigenetic alterations are associated with poor clinical outcome in hepatocellular carcinoma patients. Hepatology 2013, 58, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, C.; Zwaans, B.M.; Silberman, D.M.; Gymrek, M.; Goren, A.; Zhong, L.; Ram, O.; Truelove, J.; Guimaraes, A.R.; Toiber, D.; et al. The histone deacetylase SIRT6 is a tumor suppressor that controls cancer metabolism. Cell 2012, 151, 1185–1199. [Google Scholar] [CrossRef] [PubMed]
- Van Meter, M.; Mao, Z.; Gorbunova, V.; Seluanov, A. Sirt6 overexpression induces massive apoptosis in cancer cells but not in normal cells. Cell. Cycle 2011, 10, 3153–3158. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Ji, Y.; Bakiri, L.; Qiu, Z.; Cen, J.; Chen, X.; Chen, L.; Scheuch, H.; Zheng, H.; Qin, L.; et al. Liver cancer initiation is controlled by AP-1 through SIRT6-dependent inhibition of survivin. Nat. Cell. Biol. 2012, 14, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, Q.R.; Wang, B.; Shao, J.; Zhang, T.; Liu, T.; Huang, G.; Xia, W. Inhibition of SIRT6 in prostate cancer reduces cell viability and increases sensitivity to chemotherapeutics. Protein Cell. 2013, 4, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Ming, M.; Han, W.; Zhao, B.; Sundaresan, N.R.; Deng, C.X.; Gupta, M.P.; He, Y.Y. SIRT6 promotes COX-2 expression and acts as an oncogene in skin cancer. Cancer Res. 2014, 74, 5925–5933. [Google Scholar] [CrossRef] [PubMed]
- Han, W.D.; Mu, Y.M.; Lu, X.C.; Xu, Z.M.; Li, X.J.; Yu, L.; Song, H.J.; Li, M.; Lu, J.M.; Zhao, Y.L.; et al. Up-regulation of LRP16 mrna by 17beta-estradiol through activation of estrogen receptor alpha (eralpha), but not erbeta, and promotion of human breast cancer MCF-7 cell proliferation: A preliminary report. Endocr. Relat. Cancer 2003, 10, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Han, W.D.; Zhao, Y.L.; Meng, Y.G.; Zang, L.; Wu, Z.Q.; Li, Q.; Si, Y.L.; Huang, K.; Ba, J.M.; Morinaga, H.; et al. Estrogenically regulated LRP16 interacts with estrogen receptor alpha and enhances the receptor’s transcriptional activity. Endocr. Relat. Cancer 2007, 14, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, Y.L.; Wu, Z.Q.; Si, Y.L.; Meng, Y.G.; Fu, X.B.; Mu, Y.M.; Han, W.D. The single-macro domain protein LRP16 is an essential cofactor of androgen receptor. Endocr. Relat. Cancer 2009, 16, 139–153. [Google Scholar] [CrossRef]
- Li, Y.Z.; Zhao, P.; Han, W.D. Clinicopathological significance of LRP16 protein in 336 gastric carcinoma patients. World J. Gastroenterol. 2009, 15, 4833–4837. [Google Scholar] [CrossRef] [PubMed]
- Xi, H.Q.; Zhao, P.; Han, W.D. Clinicopathological significance and prognostic value of LRP16 expression in colorectal carcinoma. World J. Gastroenterol. 2010, 16, 1644–1648. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, M.; Cidado, J.; Croessmann, S.; Cravero, K.; Cimino-Mathews, A.; Wong, H.Y.; Scharpf, R.; Zabransky, D.J.; Abukhdeir, A.M.; Garay, J.P.; et al. Macrod2 overexpression mediates estrogen independent growth and tamoxifen resistance in breast cancers. Proc. Natl. Acad. Sci. USA 2014, 111, 17606–17611. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, R.; Morra, R.; Appel, C.D.; Tallis, M.; Chioza, B.; Jankevicius, G.; Simpson, M.A.; Matic, I.; Ozkan, E.; Golia, B.; et al. Deficiency of terminal ADP-ribose protein glycohydrolase TARG1/C6orf130 in neurodegenerative disease. Embo J. 2013, 32, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Liszt, G.; Ford, E.; Kurtev, M.; Guarente, L. Mouse sir2 homolog sirt6 is a nuclear ADP-ribosyltransferase. J. Biol. Chem. 2005, 280, 21313–21320. [Google Scholar] [CrossRef] [PubMed]
- Haigis, M.C.; Sinclair, D.A. Mammalian sirtuins: Biological insights and disease relevance. Annu Rev. Pathol 2010, 5, 253–295. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Guarente, L. NAD+ and sirtuins in aging and disease. Trends Cell. Biol. 2014, 24, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Frye, R.A. Characterization of five human cdnas with homology to the yeast SIR2 gene: SIR2-like proteins (sirtuins) metabolize nad and may have protein adp-ribosyltransferase activity. Biochem. Biophys Res. Commun. 1999, 260, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Tanny, J.C.; Dowd, G.J.; Huang, J.; Hilz, H.; Moazed, D. An enzymatic activity in the yeast SIR2 protein that is essential for gene silencing. Cell 1999, 99, 735–745. [Google Scholar] [CrossRef]
- Daniels, C.M.; Ong, S.E.; Leung, A.K. The promise of proteomics for the study of ADP-ribosylation. Mol. Cell 2015, 58, 911–924. [Google Scholar] [CrossRef] [PubMed]
- Haag, F.; Buck, F. Identification and analysis of ADP-ribosylated proteins. Curr. Top. Microbiol. Immunol. 2015, 384, 33–50. [Google Scholar] [PubMed]
- Rosenthal, F.; Hottiger, M.O. Identification of ADP-ribosylated peptides and adp-ribose acceptor sites. Front. Biosci. (Landmark Ed.) 2014, 19, 1041–1056. [Google Scholar] [CrossRef] [PubMed]
- Otto, H.; Reche, P.A.; Bazan, F.; Dittmar, K.; Haag, F.; Koch-Nolte, F. In silico characterization of the family of PARP-like poly(ADP-ribosyl)transferases (parts). BMC Genomics 2005, 6, 139. [Google Scholar] [CrossRef] [PubMed]
- Flick, F.; Luscher, B. Regulation of sirtuin function by posttranslational modifications. Front. Pharmacol. 2012, 3, 29. [Google Scholar] [CrossRef] [PubMed]
- Bai, P. Biology of poly(ADP-ribose) polymerases: The factotums of cell maintenance. Mol. Cell 2015, 58, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Feijs, K.L.; Forst, A.H.; Verheugd, P.; Luscher, B. Macrodomain-containing proteins: Regulating new intracellular functions of mono(ADP-ribosyl)ation. Nat. Rev. Mol. Cell. Biol. 2013, 14, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Forst, A.H.; Karlberg, T.; Herzog, N.; Thorsell, A.G.; Gross, A.; Feijs, K.L.; Verheugd, P.; Kursula, P.; Nijmeijer, B.; Kremmer, E.; et al. Recognition of mono-ADP-ribosylated ARTD10 substrates by ARTD8 macrodomains. Structure 2013, 21, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Jankevicius, G.; Hassler, M.; Golia, B.; Rybin, V.; Zacharias, M.; Timinszky, G.; Ladurner, A.G. A family of macrodomain proteins reverses cellular mono-ADP-ribosylation. Nat. Struct. Mol. Biol. 2013, 20, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, F.; Feijs, K.L.; Frugier, E.; Bonalli, M.; Forst, A.H.; Imhof, R.; Winkler, H.C.; Fischer, D.; Caflisch, A.; Hassa, P.O.; et al. Macrodomain-containing proteins are new mono-ADP-ribosylhydrolases. Nat. Struct. Mol. Biol. 2013, 20, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, R.C.; Takeyama, K.; He, C.; Kreinbrink, K.; Shipp, M.A. B-aggressive lymphoma family proteins have unique domains that modulate transcription and exhibit poly(ADP-ribose) polymerase activity. J. Biol. Chem. 2005, 280, 33756–33765. [Google Scholar] [CrossRef] [PubMed]
- Walford, H.H.; Doherty, T.A. STAT6 and lung inflammation. JAKSTAT 2013, 2, e25301. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Schreek, S.; Cerni, C.; Schamberger, C.; Lesniewicz, K.; Poreba, E.; Vervoorts, J.; Walsemann, G.; Grotzinger, J.; Kremmer, E.; et al. PARP-10, a novel myc-interacting protein with poly(ADP-ribose) polymerase activity, inhibits transformation. Oncogene 2005, 24, 1982–1993. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, M.; Feijs, K.L.; Luscher, B. Function and regulation of the mono-ADP-ribosyltransferase artd10. Curr. Top. Microbiol. Immunol. 2015, 384, 167–188. [Google Scholar] [PubMed]
- Wu, D.; Pan, W. Gsk3: A multifaceted kinase in WNT signaling. Trends Biochem. Sci. 2010, 35, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Michalek, S.M.; Jope, R.S. Innate and adaptive immune responses regulated by glycogen synthase kinase-3 (GSK3). Trends Immunol. 2010, 31, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J. Ubiquitination in signaling to and activation of ikk. Immunol Rev. 2012, 246, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-kappab signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Gautheron, J.; Courtois, G. “Without ub i am nothing”: Nemo as a multifunctional player in ubiquitin-mediated control of NF-kappab activation. Cell. Mol. Life Sci. 2010, 67, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Gardner, B.M.; Pincus, D.; Gotthardt, K.; Gallagher, C.M.; Walter, P. Endoplasmic reticulum stress sensing in the unfolded protein response. Cold Spring Harb. Perspect Biol. 2013, 5, a013169. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ozcan, U. Unfolded protein response signaling and metabolic diseases. J. Biol. Chem. 2014, 289, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell. Biol. 2000, 2, 326–332. [Google Scholar] [PubMed]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell. Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, S.; Micaroni, M.; Di Tullio, G.; Buccione, R.; Di Girolamo, M. PARP16/ARTD15 is a novel endoplasmic-reticulum-associated mono-ADP-ribosyltransferase that interacts with, and modifies karyopherin-SS1. PLoS ONE 2012, 7, e37352. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.E.; Petrova, K.; Tomba, G.; Vendruscolo, M.; Ron, D. ADP ribosylation adapts an ER chaperone response to short-term fluctuations in unfolded protein load. J. Cell. Biol. 2012, 198, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, G.; Di Paola, S.; Stilla, A.; Giannotta, M.; Ruggiero, C.; Menzel, S.; Koch-Nolte, F.; Sallese, M.; Di Girolamo, M. Artc1-mediated ADP-ribosylation of GRP78/BiP: A new player in endoplasmic-reticulum stress responses. Cell. Mol. Life Sci. 2015, 72, 1209–1225. [Google Scholar] [CrossRef] [PubMed]
- Nicolae, C.M.; Aho, E.R.; Vlahos, A.H.; Choe, K.N.; De, S.; Karras, G.I.; Moldovan, G.L. The ADP-ribosyltransferase PARP10/ARTD10 interacts with proliferating cell nuclear antigen (PCNA) and is required for DNA damage tolerance. J. Biol. Chem. 2014, 289, 13627–13637. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Nicolae, C.M.; Aho, E.R.; Choe, K.N.; Constantin, D.; Hu, H.J.; Lee, D.; Myung, K.; Moldovan, G.L. A novel role for the mono-ADP-ribosyltransferase PARP14/artd8 in promoting homologous recombination and protecting against replication stress. Nucleic Acids Res. 2015, 43, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Beauharnois, J.M.; Bolivar, B.E.; Welch, J.T. Sirtuin 6: A review of biological effects and potential therapeutic properties. Mol. Biosyst. 2013, 9, 1789–1806. [Google Scholar] [CrossRef] [PubMed]
- Kugel, S.; Mostoslavsky, R. Chromatin and beyond: The multitasking roles for SIRT6. Trends Biochem. Sci. 2014, 39, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Van Meter, M.; Mao, Z.; Gorbunova, V.; Seluanov, A. Repairing split ends: SIRT6, mono-ADP ribosylation and DNA repair. Aging (Albany NY) 2011, 3, 829–835. [Google Scholar] [PubMed]
- Mostoslavsky, R.; Chua, K.F.; Lombard, D.B.; Pang, W.W.; Fischer, M.R.; Gellon, L.; Liu, P.; Mostoslavsky, G.; Franco, S.; Murphy, M.M.; et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell 2006, 124, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Chang, P. New parp targets for cancer therapy. Nat. Rev. Cancer 2014, 14, 502–509. [Google Scholar] [CrossRef]
- Dang, C.V. Links between metabolism and cancer. Genes Dev. 2012, 26, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Thompson, C.B. Tumor suppressors and cell metabolism: A recipe for cancer growth. Genes Dev. 2009, 23, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Chiarugi, A.; Dolle, C.; Felici, R.; Ziegler, M. The NAD metabolome—A key determinant of cancer cell biology. Nat. Rev. Cancer 2012, 12, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Pollak, N.; Dolle, C.; Ziegler, M. The power to reduce: Pyridine nucleotides—Small molecules with a multitude of functions. Biochem J. 2007, 402, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, E.S.; Fabrizio, G.; Di Girolamo, M. A role of intracellular mono-ADP-ribosylation in cancer biology. Febs J. 2013, 280, 3551–3562. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.Y.; Chou, H.T.; Lee, S.C. CDK-dependent activation of poly(ADP-ribose) polymerase member 10 (parp10). J. Biol. Chem. 2006, 281, 15201–15207. [Google Scholar] [CrossRef] [PubMed]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-kappab as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Raval-Fernandes, S.; Kickhoefer, V.A.; Kitchen, C.; Rome, L.H. Increased susceptibility of vault poly(ADP-ribose) polymerase-deficient mice to carcinogen-induced tumorigenesis. Cancer Res. 2005, 65, 8846–8852. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Cui, F.; Yu, F.; Lu, H.; Zhang, M.; Tang, H.; Peng, Z. Sirtuin-4 (sirt4) is downregulated and associated with some clinicopathological features in gastric adenocarcinoma. Biomed. Pharmacother. 2015, 72, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Miyo, M.; Yamamoto, H.; Konno, M.; Colvin, H.; Nishida, N.; Koseki, J.; Kawamoto, K.; Ogawa, H.; Hamabe, A.; Uemura, M.; et al. Tumour-suppressive function of SIRT4 in human colorectal cancer. Br. J. Cancer 2015, 113, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Blaveri, E.; Simko, J.P.; Korkola, J.E.; Brewer, J.L.; Baehner, F.; Mehta, K.; Devries, S.; Koppie, T.; Pejavar, S.; Carroll, P.; et al. Bladder cancer outcome and subtype classification by gene expression. Clin. Cancer Res. 2005, 11, 4044–4055. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.L.; Han, W.D.; Li, Q.; Mu, Y.M.; Lu, X.C.; Yu, L.; Song, H.J.; Li, X.; Lu, J.M.; Pan, C.Y. Mechanism of transcriptional regulation of LRP16 gene expression by 17-beta estradiol in mcf-7 human breast cancer cells. J. Mol. Endocrinol. 2005, 34, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Shang, Y. Estrogen and cancer. Annu Rev. Physiol. 2013, 75, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.G.; Han, W.D.; Zhao, Y.L.; Huang, K.; Si, Y.L.; Wu, Z.Q.; Mu, Y.M. Induction of the LRP16 gene by estrogen promotes the invasive growth of ishikawa human endometrial cancer cells through the downregulation of E-cadherin. Cell. Res. 2007, 17, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.A.; Beare, D.; Gunasekaran, P.; Leung, K.; Bindal, N.; Boutselakis, H.; Ding, M.; Bamford, S.; Cole, C.; Ward, S.; et al. Cosmic: Exploring the world’s knowledge of somatic mutations in human cancer. Nucleic Acids Res. 2015, 43, D805–811. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Trocoli, A.; Djavaheri-Mergny, M. The complex interplay between autophagy and NF-kappab signaling pathways in cancer cells. Am. J. Cancer Res. 2011, 1, 629–649. [Google Scholar] [PubMed]
- Daugherty, M.D.; Young, J.M.; Kerns, J.A.; Malik, H.S. Rapid evolution of PARP genes suggests a broad role for ADP-ribosylation in host-virus conflicts. PLoS Genet. 2014, 10, e1004403. [Google Scholar] [CrossRef] [PubMed]
- Bick, M.J.; Carroll, J.W.; Gao, G.; Goff, S.P.; Rice, C.M.; MacDonald, M.R. Expression of the zinc-finger antiviral protein inhibits alphavirus replication. J. Virol. 2003, 77, 11555–11562. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Guo, X.; Goff, S.P. Inhibition of retroviral RNA production by ZAP, a CCCH-type zinc finger protein. Science 2002, 297, 1703–1706. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Nie, H.; Cai, D.; Zhang, J.; Liu, H.; Yan, R.; Cuconati, A.; Block, T.M.; Guo, J.T.; Guo, H. Inhibition of hepatitis B virus replication by the host zinc finger antiviral protein. PLoS Pathog 2013, 9, e1003494. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Moller, P.; Bick, M.J.; Wurr, S.; Becker, S.; Gunther, S.; Kummerer, B.M. Inhibition of filovirus replication by the zinc finger antiviral protein. J. Virol. 2007, 81, 2391–2400. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, G.; Lv, F.; Wang, X.; Ji, X.; Xu, Y.; Sun, J.; Wu, L.; Zheng, Y.T.; Gao, G. Zinc-finger antiviral protein inhibits HIV-1 infection by selectively targeting multiply spliced viral mrnas for degradation. Proc. Natl. Acad. Sci. USA 2011, 108, 15834–15839. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, Y.; Zhang, K.; Wang, X.; Sun, J.; Gao, G.; Liu, Y. Structure of N-terminal domain of ZAP indicates how a zinc-finger protein recognizes complex rna. Nat. Struct. Mol. Biol. 2012, 19, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Carroll, J.W.; Macdonald, M.R.; Goff, S.P.; Gao, G. The zinc finger antiviral protein directly binds to specific viral mrnas through the CCCH zinc finger motifs. J. Virol. 2004, 78, 12781–12787. [Google Scholar] [CrossRef]
- Jeong, M.S.; Kim, E.J.; Jang, S.B. Expression and RNA-binding of human zinc-finger antiviral protein. Biochem. Biophys. Res. Commun. 2010, 396, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ma, J.; Sun, J.; Gao, G. The zinc-finger antiviral protein recruits the RNA processing exosome to degrade the target mrna. Proc. Natl. Acad. Sci. USA 2007, 104, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Glasker, S.; Toller, M.; Kummerer, B.M. The alternate triad motif of the poly(ADP-ribose) polymerase-like domain of the human zinc finger antiviral protein is essential for its antiviral activity. J. Gen. Virol. 2014, 95, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Buchan, J.R.; Parker, R. Eukaryotic stress granules: The INS and outs of translation. Mol. Cell 2009, 36, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.; Todorova, T.; Ando, Y.; Chang, P. Poly(ADP-ribose) regulates post-transcriptional gene regulation in the cytoplasm. RNA Biol. 2012, 9, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Beckham, C.J.; Parker, R. P bodies, stress granules, and viral life cycles. Cell Host Microbe 2008, 3, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Onomoto, K.; Yoneyama, M.; Fung, G.; Kato, H.; Fujita, T. Antiviral innate immunity and stress granule responses. Trends Immunol. 2014, 35, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Egloff, M.P.; Malet, H.; Putics, A.; Heinonen, M.; Dutartre, H.; Frangeul, A.; Gruez, A.; Campanacci, V.; Cambillau, C.; Ziebuhr, J.; et al. Structural and functional basis for ADP-ribose and poly(ADP-ribose) binding by viral macro domains. J. Virol. 2006, 80, 8493–8502. [Google Scholar] [CrossRef] [PubMed]
- Neuvonen, M.; Ahola, T. Differential activities of cellular and viral Macro domain proteins in binding of ADP-ribose metabolites. J. Mol. Biol. 2009, 385, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Saikatendu, K.S.; Joseph, J.S.; Subramanian, V.; Clayton, T.; Griffith, M.; Moy, K.; Velasquez, J.; Neuman, B.W.; Buchmeier, M.J.; Stevens, R.C.; et al. Structural basis of severe acute respiratory syndrome coronavirus ADP-ribose-1′′-phosphate dephosphorylation by a conserved domain of NSP3. Structure 2005, 13, 1665–1675. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, K.K.; Cervantes-Barragan, L.; Ludewig, B.; Thiel, V. Mouse hepatitis virus liver pathology is dependent on ADP-ribose-1′′-phosphatase, a viral function conserved in the alpha-like supergroup. J. Virol. 2008, 82, 12325–12334. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Griffin, D.E. The NSP3 macro domain is important for sindbis virus replication in neurons and neurovirulence in mice. Virology 2009, 388, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Kuri, T.; Eriksson, K.K.; Putics, A.; Zust, R.; Snijder, E.J.; Davidson, A.D.; Siddell, S.G.; Thiel, V.; Ziebuhr, J.; Weber, F. The ADP-ribose-1′′-monophosphatase domains of severe acute respiratory syndrome coronavirus and human coronavirus 229e mediate resistance to antiviral interferon responses. J. Gen. Virol. 2011, 92, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Rack, J.G.; Morra, R.; Barkauskaite, E.; Kraehenbuehl, R.; Ariza, A.; Qu, Y.; Ortmayer, M.; Leidecker, O.; Cameron, D.R.; Matic, I.; et al. Identification of a class of protein ADP-ribosylating sirtuins in microbial pathogens. Mol. Cell 2015, 59, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Holbourn, K.P.; Shone, C.C.; Acharya, K.R. A family of killer toxins. Exploring the mechanism of ADP-ribosylating toxins. FEBS J. 2006, 273, 4579–4593. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Barbieri, J.T. Molecular mechanisms of the cytotoxicity of ADP-ribosylating toxins. Annu. Rev. Microbiol. 2008, 62, 271–288. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bütepage, M.; Eckei, L.; Verheugd, P.; Lüscher, B. Intracellular Mono-ADP-Ribosylation in Signaling and Disease. Cells 2015, 4, 569-595. https://doi.org/10.3390/cells4040569
Bütepage M, Eckei L, Verheugd P, Lüscher B. Intracellular Mono-ADP-Ribosylation in Signaling and Disease. Cells. 2015; 4(4):569-595. https://doi.org/10.3390/cells4040569
Chicago/Turabian StyleBütepage, Mareike, Laura Eckei, Patricia Verheugd, and Bernhard Lüscher. 2015. "Intracellular Mono-ADP-Ribosylation in Signaling and Disease" Cells 4, no. 4: 569-595. https://doi.org/10.3390/cells4040569