
Cold Shock Domain Protein DbpA Orchestrates Tubular Cell Damage and Interstitial Fibrosis in Inflammatory Kidney Disease
 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
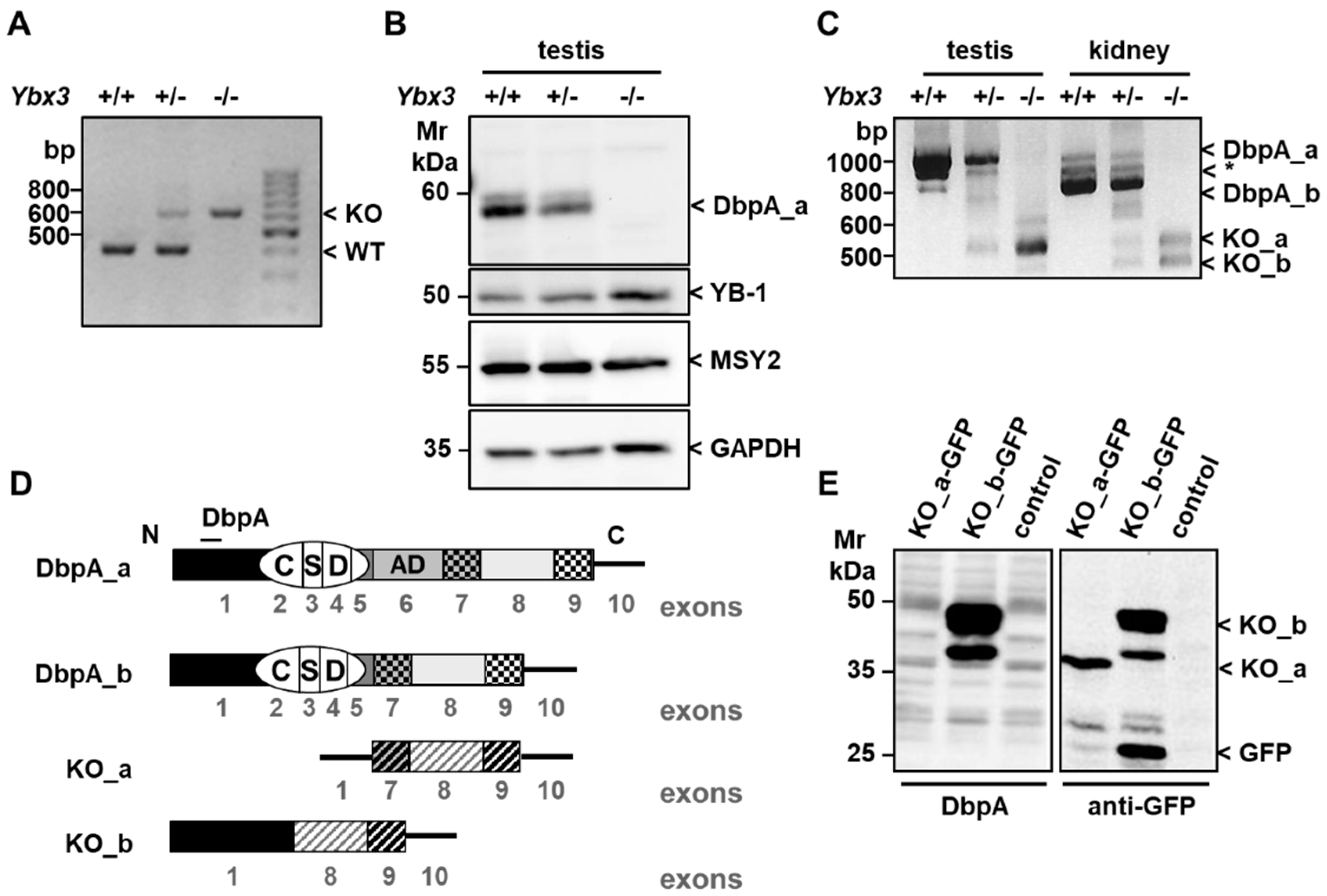
2.2. Genotyping Analyses
2.3. Animal Experiments
2.4. Isolation of Kidney Cells
2.5. Flow Cytometry
2.6. TaqMan Real-Time PCR Assay
2.7. Immunohistochemistry
2.8. Periodic Acid–Schiff (PAS) Staining
2.9. Sirius Red Staining
2.10. Western Blot Analysis
2.11. Automated Multidimension Microscopy
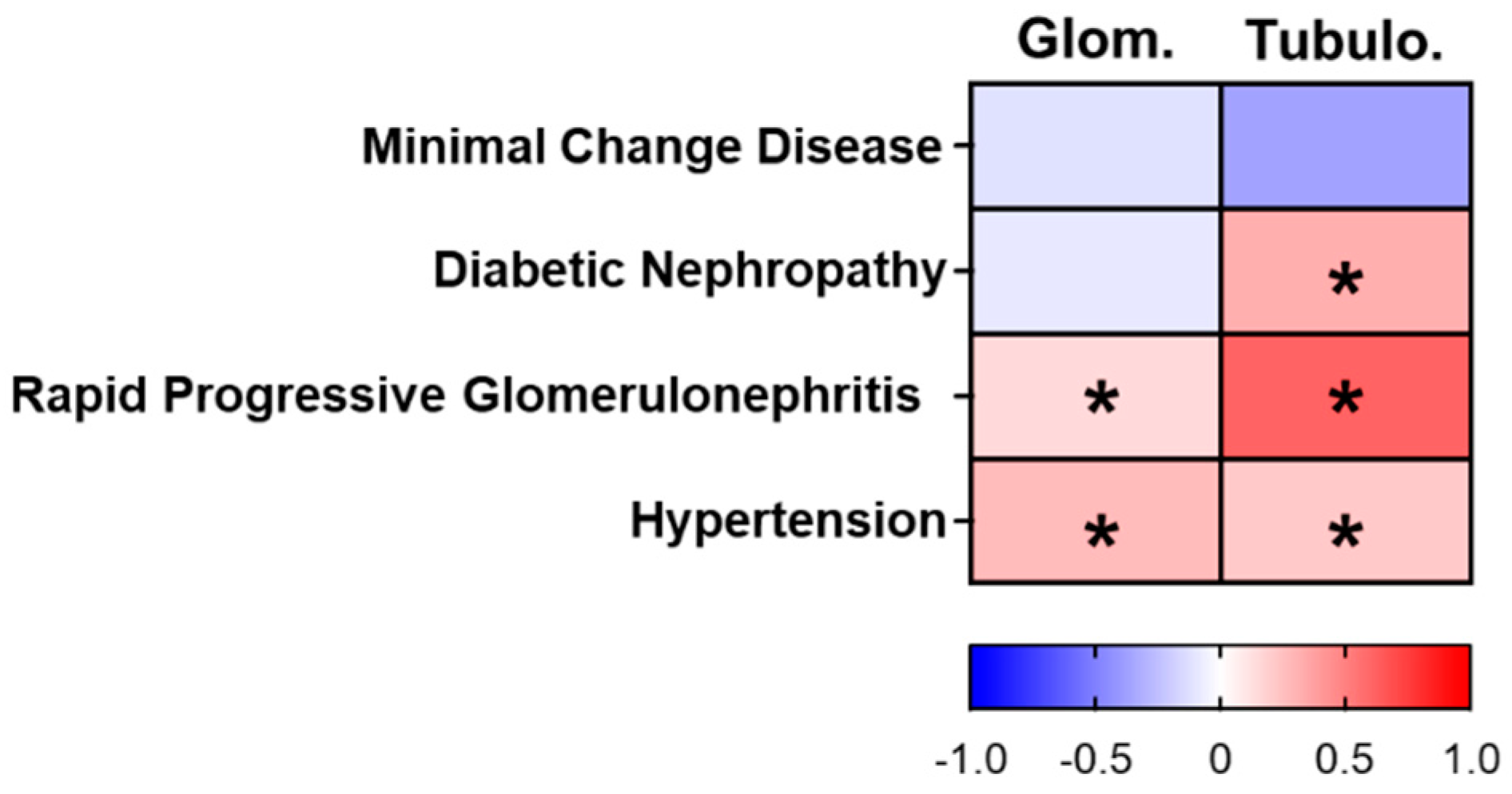
2.12. Human Microarray Analysis
2.13. Statistical Analyses
3. Results
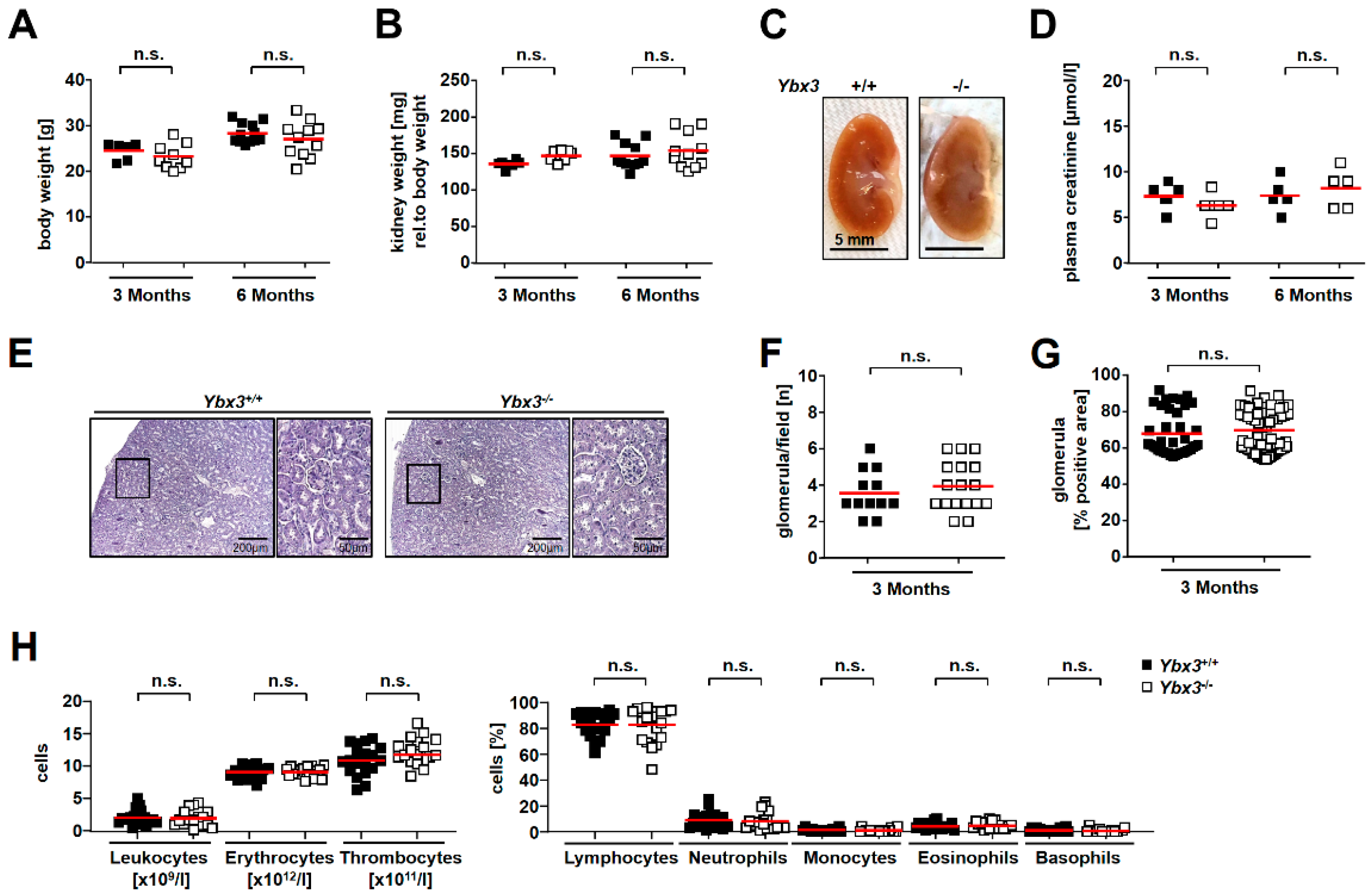
3.1. Verification and Characterization of the Ybx3 Knockout
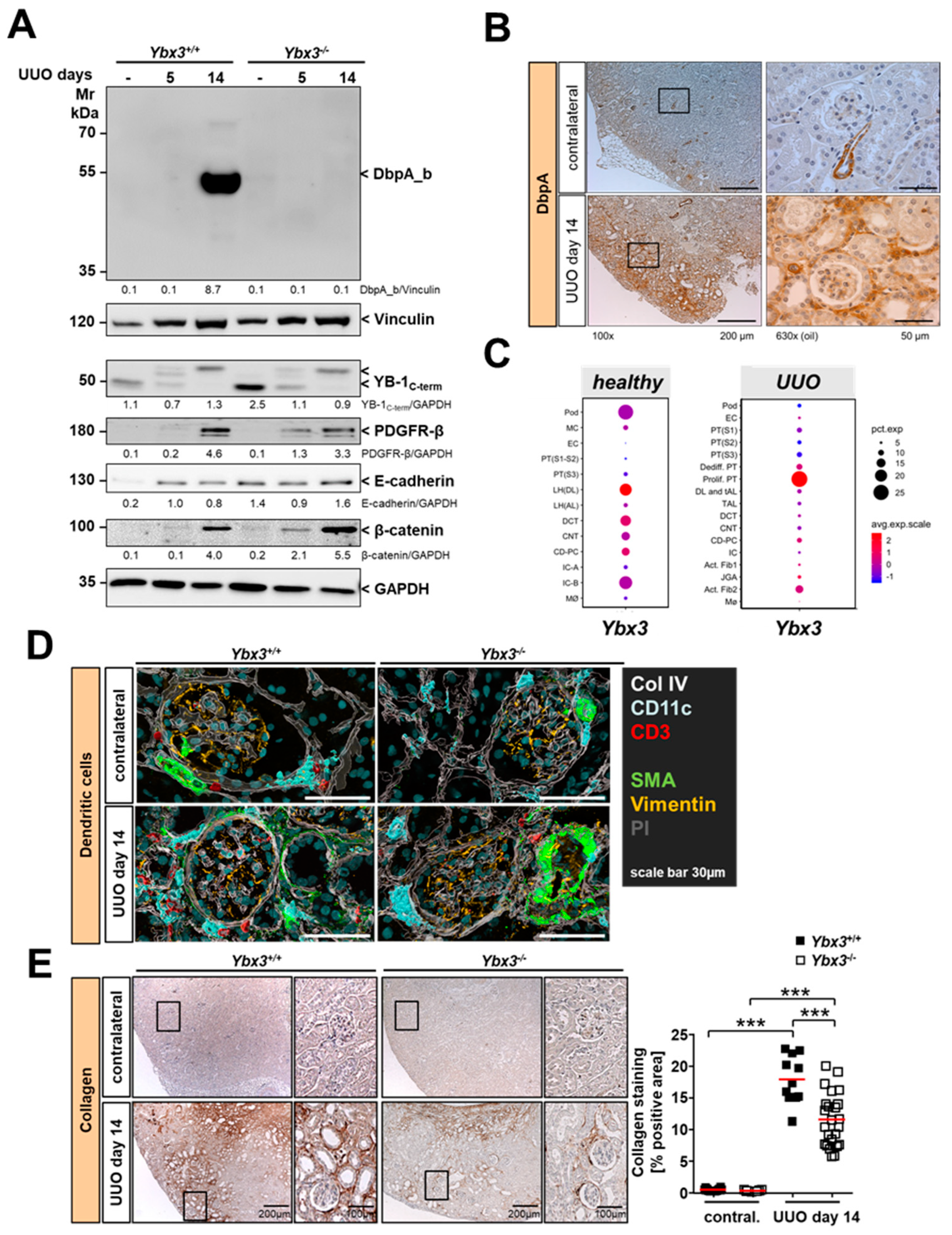
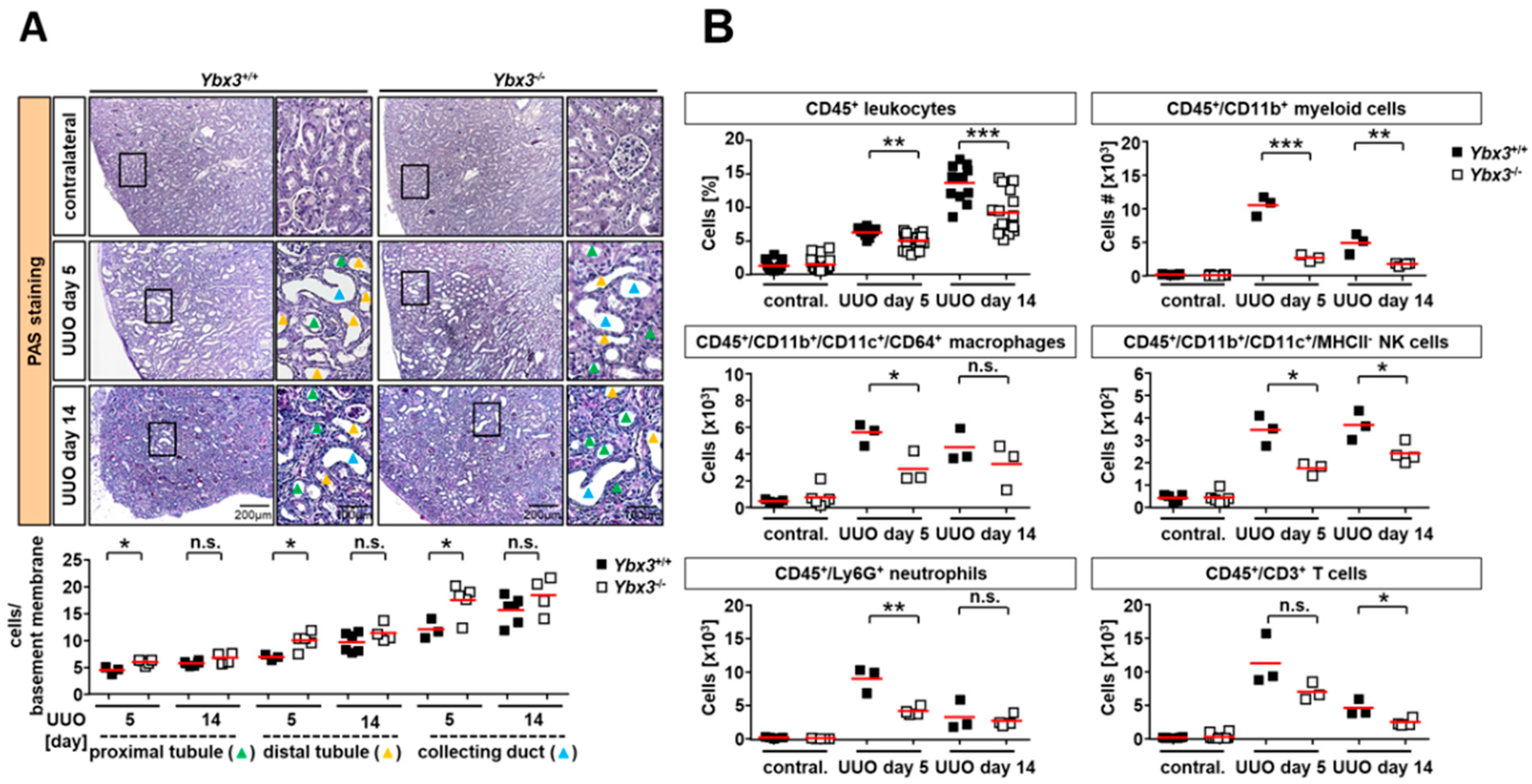
3.2. Immune Cell Infiltration Is Reduced in the Kidney of DbpA-Deficient UUO Animals
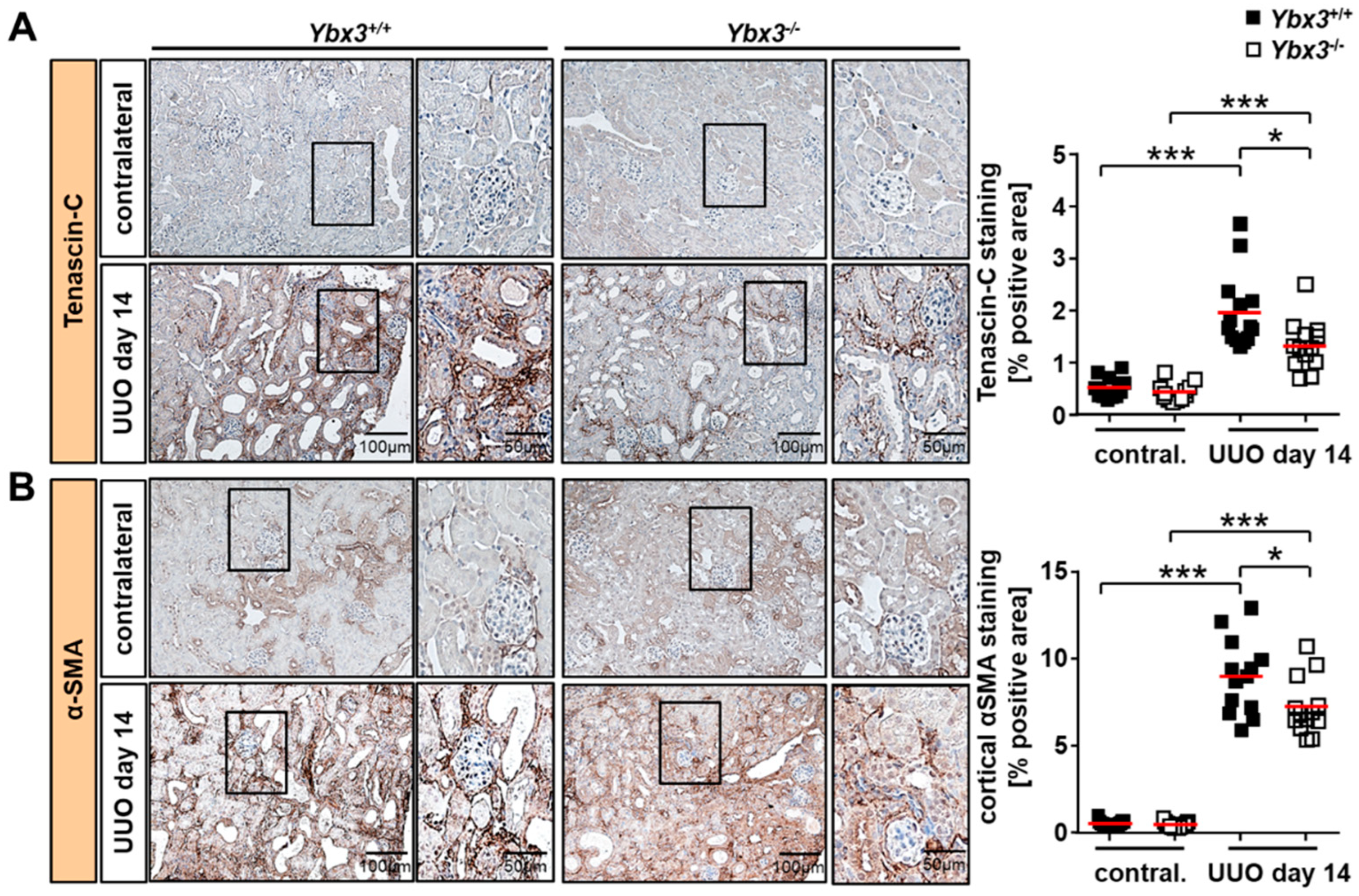
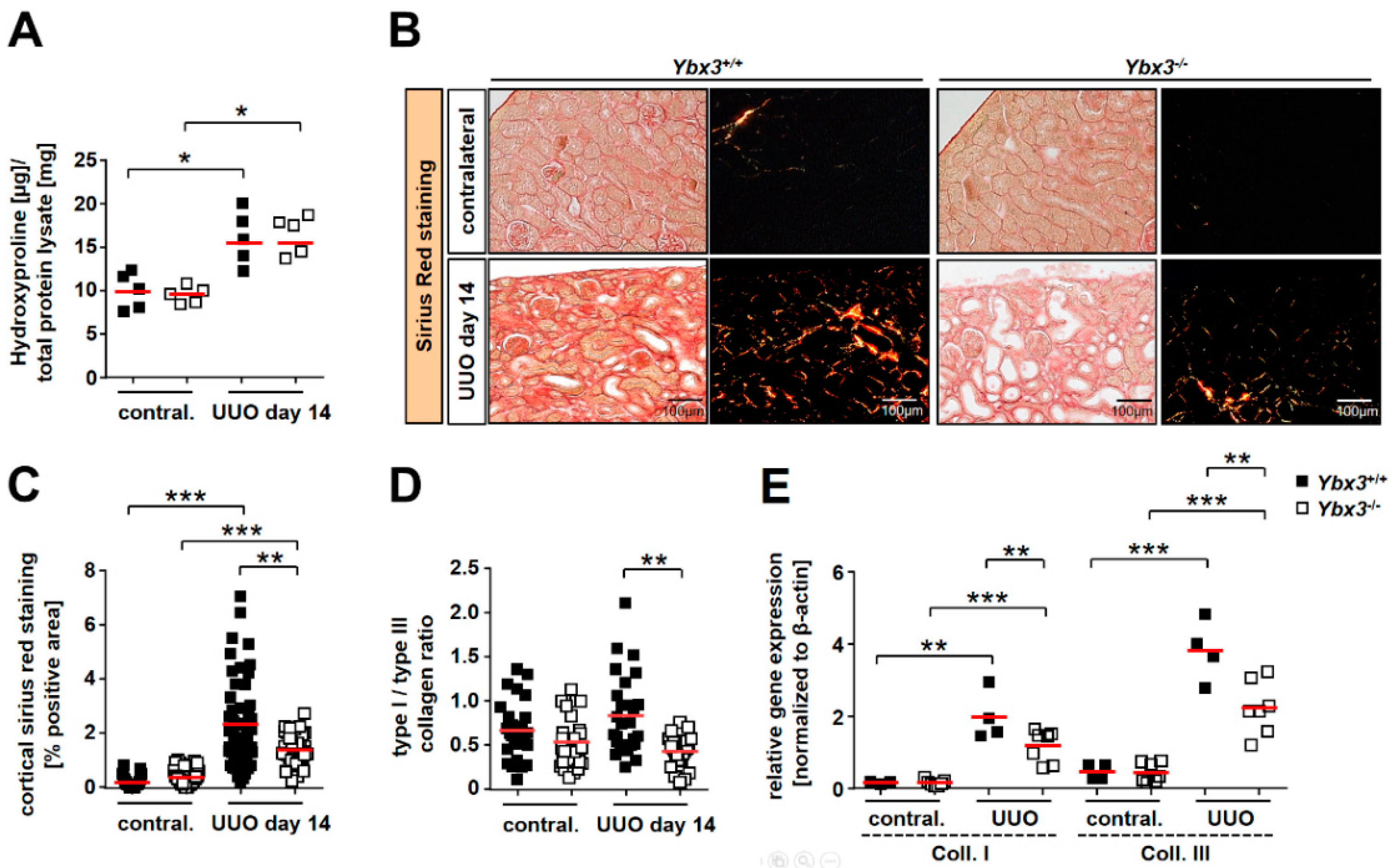
3.3. UUO-Induced Renal Interstitial Fibrosis Is Reduced in the Absence of Ybx3 In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clotet-Freixas, S.; Konvalinka, A. Too Little or Too Much? Extracellular Matrix Remodeling in Kidney Health and Disease. J. Am. Soc. Nephrol. 2021, 32, 1541–1543. [Google Scholar] [CrossRef]
- Djudjaj, S.; Boor, P. Cellular and molecular mechanisms of kidney fibrosis. Mol. Aspects Med. 2019, 65, 16–36. [Google Scholar] [CrossRef]
- Ruiz-Ortega, M.; Rayego-Mateos, S.; Lamas, S.; Ortiz, A.; Rodrigues-Diez, R.R. Targeting the progression of chronic kidney disease. Nat. Rev. Nephrol. 2020, 16, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, J.A.; Brandt, S.; Bernhardt, A.; Zhu, C.; Mertens, P.R. The role of cold shock domain proteins in inflammatory diseases. J. Mol. Med. 2014, 92, 207–216. [Google Scholar] [CrossRef]
- Lindquist, J.A.; Mertens, P.R. Cold shock proteins: From cellular mechanisms to pathophysiology and disease. Cell Commun. Signal. CCS 2018, 16, 63. [Google Scholar] [CrossRef]
- van Roeyen, C.R.; Eitner, F.; Martinkus, S.; Thieltges, S.R.; Ostendorf, T.; Bokemeyer, D.; Luscher, B.; Luscher-Firzlaff, J.M.; Floege, J.; Mertens, P.R. Y-box protein 1 mediates PDGF-B effects in mesangioproliferative glomerular disease. J. Am. Soc. Nephrol. 2005, 16, 2985–2996. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Sauter, E.; Schreiter, A.; van Roeyen, C.R.; Ostendorf, T.; Floege, J.; Gembardt, F.; Hugo, C.P.; Isermann, B.; Lindquist, J.A.; et al. Cold Shock Proteins Mediate GN with Mesangioproliferation. J. Am. Soc. Nephrol. 2016, 27, 3678–3689. [Google Scholar] [CrossRef] [PubMed]
- Brandt, S.; Mertens, P.R. A remedy for kidney disease successfully alters the cold shock protein response during inflammation. Kidney Int. 2016, 90, 1148–1150. [Google Scholar] [CrossRef]
- Wang, J.; Gibbert, L.; Djudjaj, S.; Alidousty, C.; Rauen, T.; Kunter, U.; Rembiak, A.; Enders, D.; Jankowski, V.; Braun, G.S.; et al. Therapeutic nuclear shuttling of YB-1 reduces renal damage and fibrosis. Kidney Int. 2016, 90, 1226–1237. [Google Scholar] [CrossRef]
- Sakura, H.; Maekawa, T.; Imamoto, F.; Yasuda, K.; Ishii, S. Two human genes isolated by a novel method encode DNA-binding proteins containing a common region of homology. Gene 1988, 73, 499–507. [Google Scholar]
- Balda, M.S.; Matter, K. The tight junction protein ZO-1 and an interacting transcription factor regulate ErbB-2 expression. EMBO J. 2000, 19, 2024–2033. [Google Scholar] [CrossRef] [PubMed]
- Kudo, S.; Mattei, M.G.; Fukuda, M. Characterization of the gene for dbpA, a family member of the nucleic-acid-binding proteins containing a cold-shock domain. Eur. J. Biochem. FEBS 1995, 231, 72–82. [Google Scholar] [CrossRef]
- Moorthamer, M.; Mecker, S.; Chaudhuri, B. DNA binding protein dbpA binds Cdk5 and inhibits its activity. FEBS Lett. 1999, 446, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Yasen, M.; Kajino, K.; Kano, S.; Tobita, H.; Yamamoto, J.; Uchiumi, T.; Kon, S.; Maeda, M.; Obulhasim, G.; Arii, S.; et al. The up-regulation of Y-box binding proteins (DNA binding protein A and Y-box binding protein-1) as prognostic markers of hepatocellular carcinoma. Clin. Cancer Res. 2005, 11, 7354–7361. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, Y.; Kajino, K.; Kano, S.; Tobita, H.; Hayashi, J.; Yasen, M.; Moriyama, M.; Arakawa, Y.; Hino, O. Transcription of dbpA, a Y box binding protein, is positively regulated by E2F1: Implications in hepatocarcinogenesis. Biochem. Biophys. Res. Commun. 2004, 322, 297–302. [Google Scholar] [CrossRef]
- Hayashi, J.; Kajino, K.; Umeda, T.; Takano, S.; Arakawa, Y.; Kudo, M.; Hino, O. Somatic mutation and SNP in the promoter of dbpA and human hepatocarcinogenesis. Int. J. Oncol. 2002, 21, 847–850. [Google Scholar] [CrossRef]
- Wang, G.R.; Zheng, Y.; Che, X.M.; Wang, X.Y.; Zhao, J.H.; Wu, K.J.; Zeng, J.; Pan, C.E.; He, D.L. Upregulation of human DNA binding protein A (dbpA) in gastric cancer cells. Acta Pharmacol. Sin. 2009, 30, 1436–1442. [Google Scholar] [CrossRef]
- Jayagopal, A.; Yang, J.L.; Haselton, F.R.; Chang, M.S. Tight junction-associated signaling pathways modulate cell proliferation in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 588–593. [Google Scholar] [CrossRef]
- Sears, D.; Luong, P.; Yuan, M.; Nteliopoulos, G.; Man, Y.K.; Melo, J.V.; Basu, S. Functional phosphoproteomic analysis reveals cold-shock domain protein A to be a Bcr-Abl effector-regulating proliferation and transformation in chronic myeloid leukemia. Cell Death Dis. 2010, 1, e93. [Google Scholar] [CrossRef]
- Sourisseau, T.; Georgiadis, A.; Tsapara, A.; Ali, R.R.; Pestell, R.; Matter, K.; Balda, M.S. Regulation of PCNA and cyclin D1 expression and epithelial morphogenesis by the ZO-1-regulated transcription factor ZONAB/DbpA. Mol. Cell. Biol. 2006, 26, 2387–2398. [Google Scholar] [CrossRef]
- Lima, W.R.; Parreira, K.S.; Devuyst, O.; Caplanusi, A.; N’Kuli, F.; Marien, B.; Van Der Smissen, P.; Alves, P.M.; Verroust, P.; Christensen, E.I.; et al. ZONAB promotes proliferation and represses differentiation of proximal tubule epithelial cells. J. Am. Soc. Nephrol. 2010, 21, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Balda, M.S.; Garrett, M.D.; Matter, K. The ZO-1-associated Y-box factor ZONAB regulates epithelial cell proliferation and cell density. J. Cell Biol. 2003, 160, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Aijaz, S.; Leefa Chong San, I.V.; Balda, M.S.; Matter, K. The Y-box factor ZONAB/DbpA associates with GEF-H1/Lfc and mediates Rho-stimulated transcription. EMBO Rep. 2009, 10, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Frankel, P.; Aronheim, A.; Kavanagh, E.; Balda, M.S.; Matter, K.; Bunney, T.D.; Marshall, C.J. RalA interacts with ZONAB in a cell density-dependent manner and regulates its transcriptional activity. EMBO J. 2005, 24, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.C.; Lau, J.S.; Rawlings, S.; Lee, A.S. Positive and negative regulation of the human thymidine kinase promoter mediated by CCAAT binding transcription factors NF-Y/CBF, dbpA, and CDP/cut. Cell Growth Differ. 1997, 8, 1329–1338. [Google Scholar]
- Chevalier, R.L.; Forbes, M.S.; Thornhill, B.A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int. 2009, 75, 1145–1152. [Google Scholar] [CrossRef]
- Ucero, A.C.; Benito-Martin, A.; Izquierdo, M.C.; Sanchez-Nino, M.D.; Sanz, A.B.; Ramos, A.M.; Berzal, S.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A. Unilateral ureteral obstruction: Beyond obstruction. Int. Urol. Nephrol. 2014, 46, 765–776. [Google Scholar] [CrossRef]
- Bernhardt, A.; Fehr, A.; Brandt, S.; Jerchel, S.; Ballhause, T.M.; Philipsen, L.; Stolze, S.; Geffers, R.; Weng, H.; Fischer, K.D.; et al. Inflammatory cell infiltration and resolution of kidney inflammation is orchestrated by the cold-shock protein Y-box binding protein-1. Kidney Int. 2017, 92, 1157–1177. [Google Scholar] [CrossRef]
- Brandt, S.; Ballhause, T.M.; Bernhardt, A.; Becker, A.; Salaru, D.; Le-Deffge, H.M.; Fehr, A.; Fu, Y.; Philipsen, L.; Djudjaj, S.; et al. Fibrosis and Immune Cell Infiltration Are Separate Events Regulated by Cell-Specific Receptor Notch3 Expression. J. Am. Soc. Nephrol. 2020, 31, 2589–2608. [Google Scholar] [CrossRef]
- Cohen, C.D.; Frach, K.; Schlondorff, D.; Kretzler, M. Quantitative gene expression analysis in renal biopsies: A novel protocol for a high-throughput multicenter application. Kidney Int. 2002, 61, 133–140. [Google Scholar] [CrossRef]
- Cohen, C.D.; Klingenhoff, A.; Boucherot, A.; Nitsche, A.; Henger, A.; Brunner, B.; Schmid, H.; Merkle, M.; Saleem, M.A.; Koller, K.P.; et al. Comparative promoter analysis allows de novo identification of specialized cell junction-associated proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 5682–5687. [Google Scholar] [CrossRef] [PubMed]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.H.; Books, J.T.; Ley, T.J. Cold shock domain family members YB-1 and MSY4 share essential functions during murine embryogenesis. Mol. Cell. Biol. 2006, 26, 8410–8417. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Samaras, P.; Frejno, M.; Gessulat, S.; Barnert, M.; Kienegger, H.; Krcmar, H.; Schlegl, J.; Ehrlich, H.C.; Aiche, S.; et al. ProteomicsDB. Nucleic Acids Res. 2018, 46, D1271–D1281. [Google Scholar] [CrossRef]
- Ito, K.; Tsutsumi, K.; Kuzumaki, T.; Gomez, P.F.; Otsu, K.; Ishikawa, K. A novel growth-inducible gene that encodes a protein with a conserved cold-shock domain. Nucleic Acids Res. 1994, 22, 2036–2041. [Google Scholar] [CrossRef]
- Sommer, M.; Eismann, U.; Deuther-Conrad, W.; Wendt, T.; Mohorn, T.; Funfstuck, R.; Stein, G. Time course of cytokine mRNA expression in kidneys of rats with unilateral ureteral obstruction. Nephron 2000, 84, 49–57. [Google Scholar] [CrossRef]
- Docherty, N.G.; Calvo, I.F.; Quinlan, M.R.; Perez-Barriocanal, F.; McGuire, B.B.; Fitzpatrick, J.M.; Watson, R.W. Increased E-cadherin expression in the ligated kidney following unilateral ureteric obstruction. Kidney Int. 2009, 75, 205–213. [Google Scholar] [CrossRef]
- Surendran, K.; Schiavi, S.; Hruska, K.A. Wnt-dependent beta-catenin signaling is activated after unilateral ureteral obstruction, and recombinant secreted frizzled-related protein 4 alters the progression of renal fibrosis. J. Am. Soc. Nephrol. 2005, 16, 2373–2384. [Google Scholar] [CrossRef]
- Bernhardt, A.; Haberer, S.; Xu, J.; Damerau, H.; Steffen, J.; Reichardt, C.; Wolters, K.; Steffen, H.; Isermann, B.; Borucki, K.; et al. High salt diet-induced proximal tubular phenotypic changes and sodium-glucose cotransporter-2 expression are coordinated by cold shock Y-box binding protein-1. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2021, 35, e21912. [Google Scholar] [CrossRef]
- Lyabin, D.N.; Smolin, E.A.; Budkina, K.S.; Eliseeva, I.A.; Ovchinnikov, L.P. Towards the mechanism(s) of YB-3 synthesis regulation by YB-1. RNA Biol. 2021, 18, 1630–1641. [Google Scholar] [CrossRef]
- Lyabin, D.N.; Eliseeva, I.A.; Smolin, E.A.; Doronin, A.N.; Budkina, K.S.; Kulakovskiy, I.V.; Ovchinnikov, L.P. YB-3 substitutes YB-1 in global mRNA binding. RNA Biol. 2020, 17, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Malone, A.F.; Donnelly, E.L.; Kirita, Y.; Uchimura, K.; Ramakrishnan, S.M.; Gaut, J.P.; Humphreys, B.D. Single-Cell Transcriptomics of a Human Kidney Allograft Biopsy Specimen Defines a Diverse Inflammatory Response. J. Am. Soc. Nephrol. 2018, 29, 2069–2080. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Kirita, Y.; Donnelly, E.L.; Humphreys, B.D. Advantages of Single-Nucleus over Single-Cell RNA Sequencing of Adult Kidney: Rare Cell Types and Novel Cell States Revealed in Fibrosis. J. Am. Soc. Nephrol. 2019, 30, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Bulow, R.D.; Boor, P. Extracellular Matrix in Kidney Fibrosis: More Than Just a Scaffold. J. Histochem. Cytochem. 2019, 67, 643–661. [Google Scholar] [CrossRef] [PubMed]
- Potter, J.A.; Price, G.W.; Cliff, C.L.; Green, C.R.; Squires, P.E.; Hills, C.E. Collagen I Modifies Connexin-43 Hemichannel Activity via Integrin alpha2beta1 Binding in TGFbeta1-Evoked Renal Tubular Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 3644. [Google Scholar] [CrossRef] [PubMed]
- Buchtler, S.; Grill, A.; Hofmarksrichter, S.; Stockert, P.; Schiechl-Brachner, G.; Rodriguez Gomez, M.; Neumayer, S.; Schmidbauer, K.; Talke, Y.; Klinkhammer, B.M.; et al. Cellular Origin and Functional Relevance of Collagen I Production in the Kidney. J. Am. Soc. Nephrol. 2018, 29, 1859–1873. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Tian, Y.; Zhou, L.; Zhou, D.; Tan, R.J.; Stolz, D.B.; Liu, Y. Tenascin-C Is a Major Component of the Fibrogenic Niche in Kidney Fibrosis. J. Am. Soc. Nephrol. 2017, 28, 785–801. [Google Scholar] [CrossRef]
- Snelgrove, S.L.; Kausman, J.Y.; Lo, C.; Lo, C.; Ooi, J.D.; Coates, P.T.; Hickey, M.J.; Holdsworth, S.R.; Kurts, C.; Engel, D.R.; et al. Renal dendritic cells adopt a pro-inflammatory phenotype in obstructive uropathy to activate T cells but do not directly contribute to fibrosis. Am. J. Pathol. 2012, 180, 91–103. [Google Scholar] [CrossRef]
- Stamatiades, E.G.; Tremblay, M.E.; Bohm, M.; Crozet, L.; Bisht, K.; Kao, D.; Coelho, C.; Fan, X.; Yewdell, W.T.; Davidson, A.; et al. Immune Monitoring of Trans-endothelial Transport by Kidney-Resident Macrophages. Cell 2016, 166, 991–1003. [Google Scholar] [CrossRef]
- Wyczanska, M.; Lange-Sperandio, B. DAMPs in Unilateral Ureteral Obstruction. Front. Immunol. 2020, 11, 581300. [Google Scholar] [CrossRef]
- van der Zande, H.J.P.; Nitsche, D.; Schlautmann, L.; Guigas, B.; Burgdorf, S. The Mannose Receptor: From Endocytic Receptor and Biomarker to Regulator of (Meta)Inflammation. Front. Immunol. 2021, 12, 765034. [Google Scholar] [CrossRef] [PubMed]
- Kuppe, C.; Ibrahim, M.M.; Kranz, J.; Zhang, X.; Ziegler, S.; Perales-Paton, J.; Jansen, J.; Reimer, K.C.; Smith, J.R.; Dobie, R.; et al. Decoding myofibroblast origins in human kidney fibrosis. Nature 2021, 589, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Dooley, S.; Said, H.M.; Gressner, A.M.; Floege, J.; En-Nia, A.; Mertens, P.R. Y-box protein-1 is the crucial mediator of antifibrotic interferon-gamma effects. J. Biol. Chem. 2006, 281, 1784–1795. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Zheng, M.; Luo, Q.; Jiang, D.; Zhang, H.; Chen, C. YB-1 as an Oncoprotein: Functions, Regulation, Post-Translational Modifications, and Targeted Therapy. Cells 2022, 11, 1217. [Google Scholar] [CrossRef] [PubMed]
- Arvaniti, E.; Moulos, P.; Vakrakou, A.; Chatziantoniou, C.; Chadjichristos, C.; Kavvadas, P.; Charonis, A.; Politis, P.K. Whole-transcriptome analysis of UUO mouse model of renal fibrosis reveals new molecular players in kidney diseases. Sci. Rep. 2016, 6, 26235. [Google Scholar] [CrossRef]
- Uchiumi, T.; Fotovati, A.; Sasaguri, T.; Shibahara, K.; Shimada, T.; Fukuda, T.; Nakamura, T.; Izumi, H.; Tsuzuki, T.; Kuwano, M.; et al. YB-1 is important for an early stage embryonic development: Neural tube formation and cell proliferation. J. Biol. Chem. 2006, 281, 40440–40449. [Google Scholar] [CrossRef]
- Lu, Z.H.; Books, J.T.; Ley, T.J. YB-1 is important for late-stage embryonic development, optimal cellular stress responses, and the prevention of premature senescence. Mol. Cell. Biol. 2005, 25, 4625–4637. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindquist, J.A.; Bernhardt, A.; Reichardt, C.; Sauter, E.; Brandt, S.; Rana, R.; Lindenmeyer, M.T.; Philipsen, L.; Isermann, B.; Zhu, C.; et al. Cold Shock Domain Protein DbpA Orchestrates Tubular Cell Damage and Interstitial Fibrosis in Inflammatory Kidney Disease. Cells 2023, 12, 1426. https://doi.org/10.3390/cells12101426
Lindquist JA, Bernhardt A, Reichardt C, Sauter E, Brandt S, Rana R, Lindenmeyer MT, Philipsen L, Isermann B, Zhu C, et al. Cold Shock Domain Protein DbpA Orchestrates Tubular Cell Damage and Interstitial Fibrosis in Inflammatory Kidney Disease. Cells. 2023; 12(10):1426. https://doi.org/10.3390/cells12101426
Chicago/Turabian StyleLindquist, Jonathan A., Anja Bernhardt, Charlotte Reichardt, Eva Sauter, Sabine Brandt, Rajiv Rana, Maja T. Lindenmeyer, Lars Philipsen, Berend Isermann, Cheng Zhu, and et al. 2023. "Cold Shock Domain Protein DbpA Orchestrates Tubular Cell Damage and Interstitial Fibrosis in Inflammatory Kidney Disease" Cells 12, no. 10: 1426. https://doi.org/10.3390/cells12101426