
Heat Shock Protein 27 Is Involved in the Bioactive Glass Induced Osteogenic Response of Human Mesenchymal Stem Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Expansion of hASCs and hBMSCs
2.2. Characterization of Immunophenotype of the Cells
2.3. 3-06 Bioactive Glass Manufacturing and Extract Preparation
2.4. Cell Seeding and Differentiation Culture
2.5. Western Blot and Immunodetection
2.6. Cell Proliferation and Quantitative Analysis of Alkaline Phosphatase Activity
2.7. Alizarin Red Staining of Matrix Mineralization
2.8. Immunocytochemical Staining
2.9. Fluorescence Imaging and Image-Based Analysis of Stain Intensity
2.10. Statistical Analysis
3. Results
3.1. Characterization of the Mesenchymal Origin of the CELLS
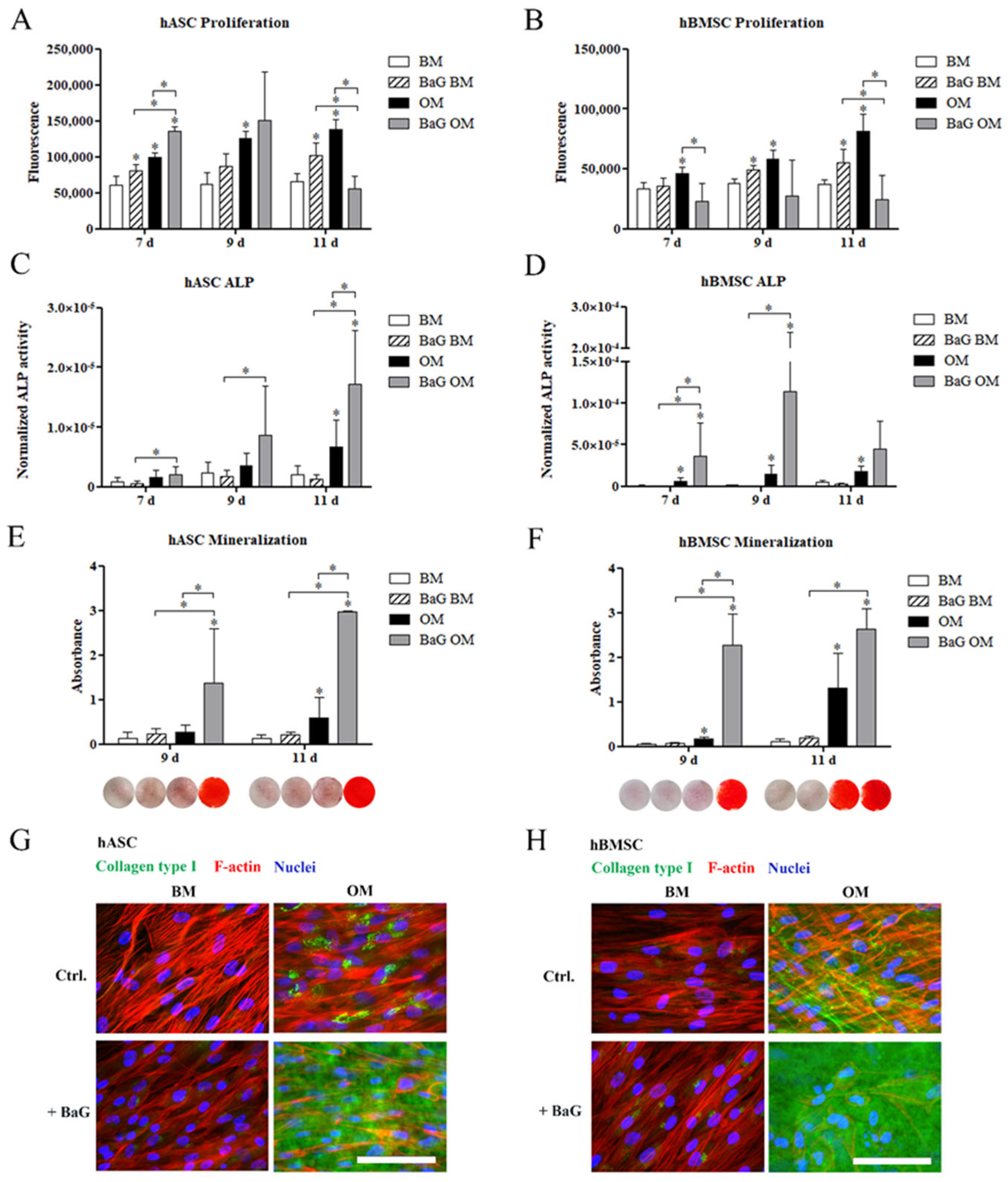
3.2. Proliferation and Osteogenic Differentiation
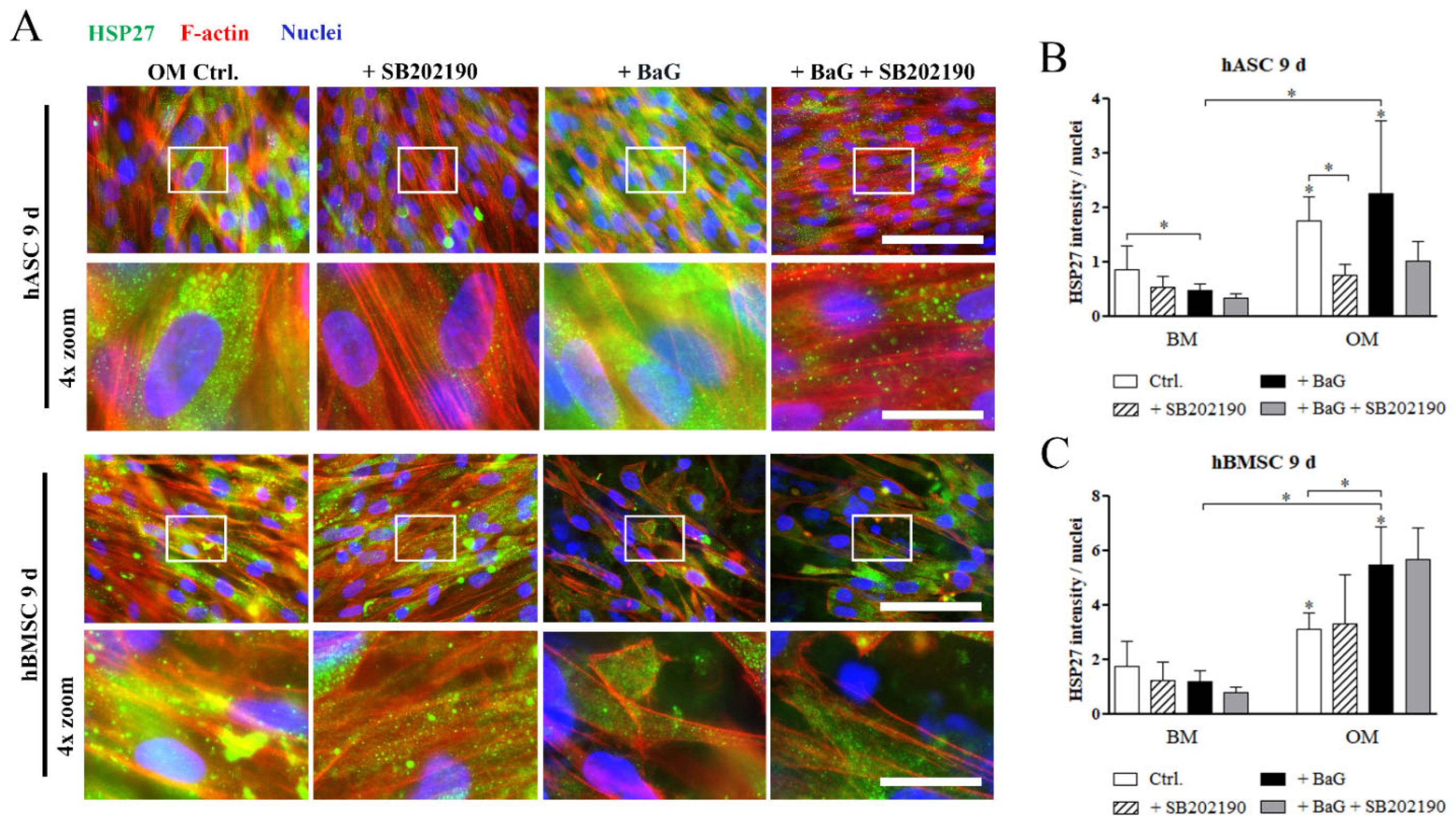
3.3. Activation of the p38/MK2/HSP27 Pathway
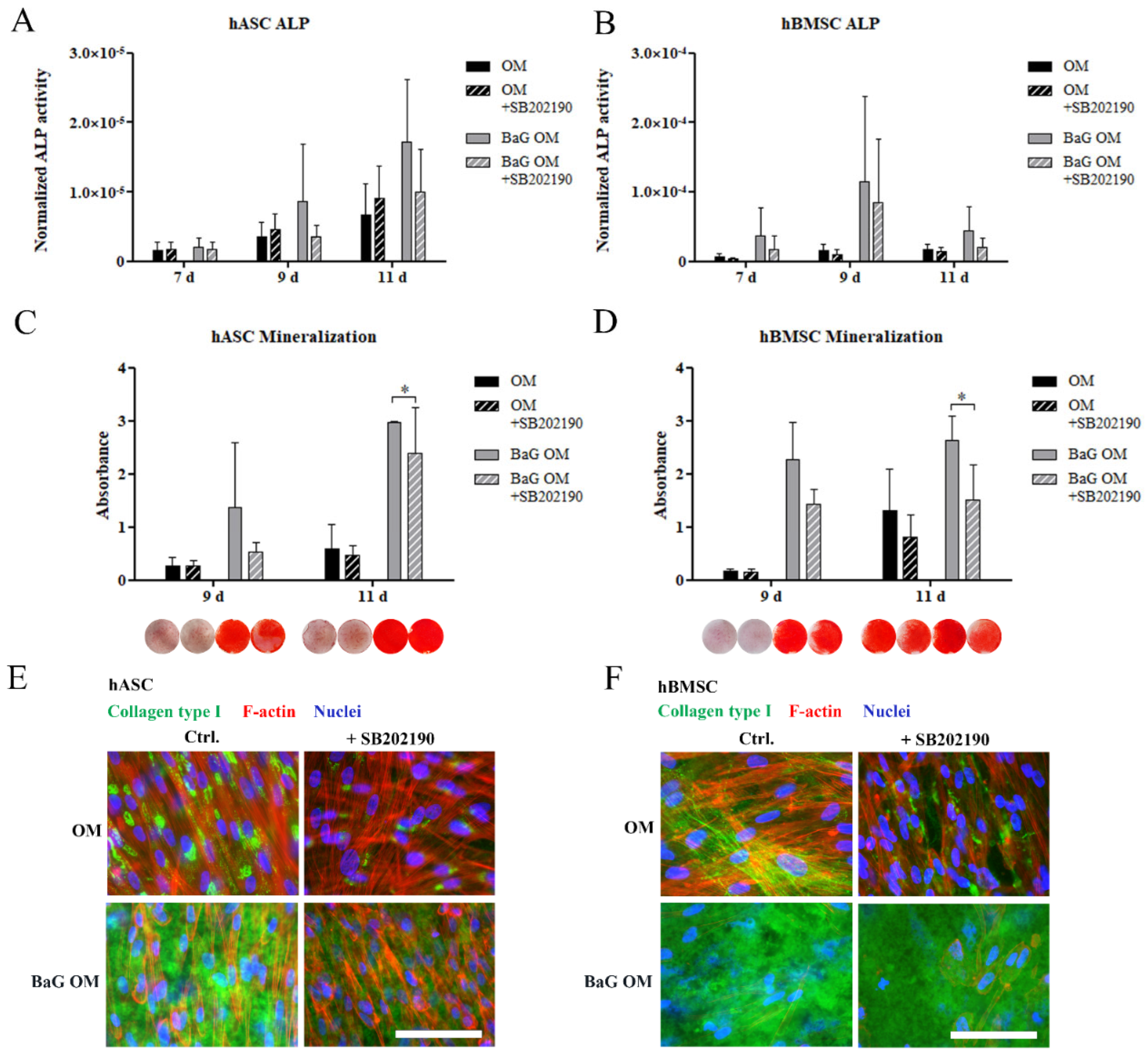
3.4. Inhibition of HSP27 Phosphorylation
3.5. HSP27 and pHSP27 Localization
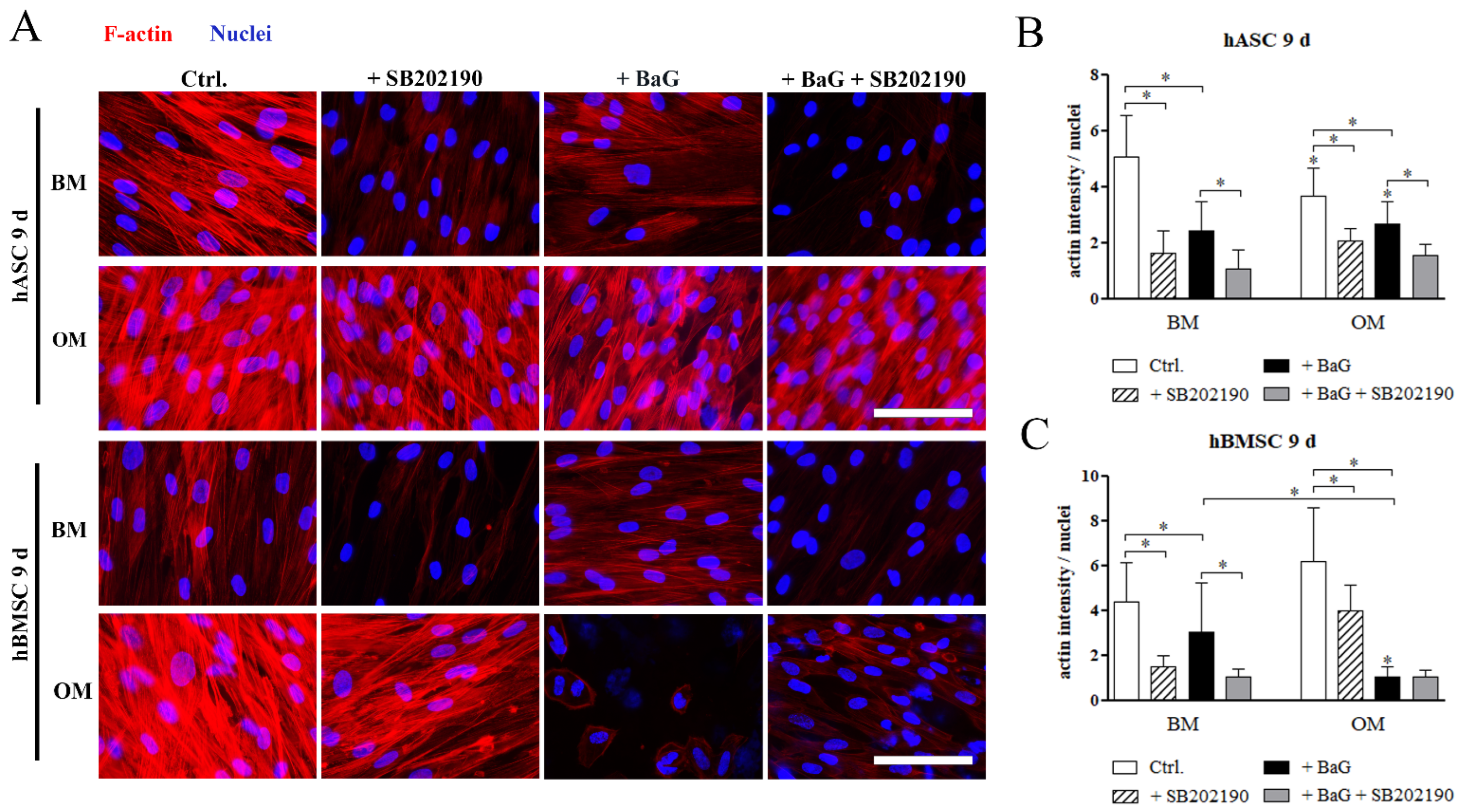
3.6. The Cell Morphology and F-Actin Intensity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knight, M.N.; Hankenson, K.D. Mesenchymal Stem Cells in Bone Regeneration. Adv. Wound Care 2013, 2, 306–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage Potential of Adult Human Mesenchymal Stem Cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I.; Bruder, S.P. Mesenchymal Stem Cells: Building Blocks for Molecular Medicine in the 21st Century. Trends Mol. Med. 2001, 7, 259–264. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Kluter, H.; Bieback, K. Comparative Analysis of Mesenchymal Stem Cells from Bone Marrow, Umbilical Cord Blood, or Adipose Tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Varma, M.J.; Breuls, R.G.; Schouten, T.E.; Jurgens, W.J.; Bontkes, H.J.; Schuurhuis, G.J.; van Ham, S.M.; van Milligen, F.J. Phenotypical and Functional Characterization of Freshly Isolated Adipose Tissue-Derived Stem Cells. Stem Cells Dev. 2007, 16, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Rahaman, M.N.; Day, D.E.; Bal, B.S.; Fu, Q.; Jung, S.B.; Bonewald, L.F.; Tomsia, A.P. Bioactive Glass in Tissue Engineering. Acta Biomater. 2011, 7, 2355–2373. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.R. Review of Bioactive Glass: From Hench to Hybrids. Acta Biomater. 2013, 9, 4457–4486. [Google Scholar] [CrossRef]
- Hench, L.L.; Jones, J.R. Bioactive Glasses: Frontiers and Challenges. Front. Bioeng. Biotechnol. 2015, 3, 194. [Google Scholar] [CrossRef] [Green Version]
- Bosch-Rué, E.; Diez-Tercero, L.; Giordano-Kelhoffer, B.; Delgado, L.M.; Bosch, B.M.; Hoyos-Nogués, M.; Mateos-Timoneda, M.A.; Tran, P.A.; Gil, F.J.; Perez, R.A. Biological Roles and Delivery Strategies for Ions to Promote Osteogenic Induction. Front. Cell Dev. Biol. 2021, 8, 614545. [Google Scholar] [CrossRef]
- Wang, X.; Molino, B.Z.; Pitkänen, S.; Ojansivu, M.; Xu, C.; Hannula, M.; Hyttinen, J.; Miettinen, S.; Hupa, L.; Wallace, G. 3D Scaffolds of Polycaprolactone/Copper-Doped Bioactive Glass: Architecture Engineering with Additive Manufacturing and Cellular Assessments in a Coculture of Bone Marrow Stem Cells and Endothelial Cells. ACS Biomater. Sci. Eng. 2019, 5, 4496–4510. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, C.; Chang, J. Bioceramics to Regulate Stem Cells and Their Microenvironment for Tissue Regeneration. Materials Today 2019, 24, 41–56. [Google Scholar] [CrossRef]
- Sun, J.-Y.; Yang, Y.-S.; Zhong, J.; Greenspan, D.C. The Effect of the Ionic Products of Bioglass® Dissolution on Human Osteoblasts Growth Cycle in Vitro. J. Tissue Eng. Regen Med. 2007, 1, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Vuornos, K.; Ojansivu, M.; Koivisto, J.T.; Hakkanen, H.; Belay, B.; Montonen, T.; Huhtala, H.; Kaariainen, M.; Hupa, L.; Kellomaki, M.; et al. Bioactive Glass Ions Induce Efficient Osteogenic Differentiation of Human Adipose Stem Cells Encapsulated in Gellan Gum and Collagen Type I Hydrogels. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 99, 905–918. [Google Scholar] [CrossRef]
- Ojansivu, M.; Vanhatupa, S.; Bjorkvik, L.; Hakkanen, H.; Kellomaki, M.; Autio, R.; Ihalainen, J.A.; Hupa, L.; Miettinen, S. Bioactive Glass Ions as Strong Enhancers of Osteogenic Differentiation in Human Adipose Stem Cells. Acta Biomater. 2015, 21, 190–203. [Google Scholar] [CrossRef]
- Gong, W.; Huang, Z.; Dong, Y.; Gan, Y.; Li, S.; Gao, X.; Chen, X. Ionic Extraction of a Novel Nano-Sized Bioactive Glass Enhances Differentiation and Mineralization of Human Dental Pulp Cells. J. Endod. 2014, 40, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Xynos, I.D.; Edgar, A.J.; Buttery, L.D.K.; Hench, L.L.; Polak, J.M. Gene-Expression Profiling of Human Osteoblasts Following Treatment with the Ionic Products of Bioglass® 45S5 Dissolution. J. Biomed Mater. Res. 2001, 55, 151–157. [Google Scholar] [CrossRef]
- Hoffman, L.; Jensen, C.C.; Yoshigi, M.; Beckerle, M. Mechanical Signals Activate P38 MAPK Pathway-Dependent Reinforcement of Actin via Mechanosensitive HspB1. Mol. Biol. Cell 2017, 28, 2661–2675. [Google Scholar] [CrossRef]
- Rodriguez-Carballo, E.; Gamez, B.; Ventura, F. P38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Sonowal, H.; Kumar, A.; Bhattacharyya, J.; Gogoi, P.K.; Jaganathan, B.G. Inhibition of Actin Polymerization Decreases Osteogeneic Differentiation of Mesenchymal Stem Cells through P38 MAPK Pathway. J. Biomed Sci. 2013, 20, 71. [Google Scholar] [CrossRef] [Green Version]
- Canovas, B.; Nebreda, A.R. Diversity and Versatility of P38 Kinase Signalling in Health and Disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 346–366. [Google Scholar] [CrossRef]
- Gong, W.; Dong, Y.; Wang, S.; Gao, X.; Chen, X. A Novel Nano-Sized Bioactive Glass Stimulates Osteogenesis via the MAPK Pathway. RSC Adv. 2017, 7, 13760–13767. [Google Scholar] [CrossRef] [Green Version]
- Xynos, I.D.; Hukkanen, M.V.J.; Batten, J.J.; Buttery, L.D.; Hench, L.L.; Polak, J.M. Bioglass ®45S5 Stimulates Osteoblast Turnover and Enhances Bone Formation In Vitro: Implications and Applications for Bone Tissue Engineering. Calcif Tissue Int. 2000, 67, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Ambriz, X.; de Lanerolle, P.; Ambrosio, J.R. The Mechanobiology of the Actin Cytoskeleton in Stem Cells during Differentiation and Interaction with Biomaterials. Stem Cells Int. 2018, 2018, 2891957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilian, K.A.; Bugarija, B.; Lahn, B.T.; Mrksich, M. Geometric Cues for Directing the Differentiation of Mesenchymal Stem Cells. Proc. Natl. Acad. Sci. USA 2010, 107, 4872–4877. [Google Scholar] [CrossRef] [Green Version]
- Yourek, G.; Hussain, M.A.; Mao, J.J. Cytoskeletal Changes of Mesenchymal Stem Cells During Differentiation. ASAIO J. 2007, 53, 219. [Google Scholar] [CrossRef] [Green Version]
- Hyväri, L.; Ojansivu, M.; Juntunen, M.; Kartasalo, K.; Miettinen, S.; Vanhatupa, S. Focal Adhesion Kinase and ROCK Signaling Are Switch-Like Regulators of Human Adipose Stem Cell Differentiation towards Osteogenic and Adipogenic Lineages. Stem Cells Int. 2018, 2018, 2190657. [Google Scholar] [CrossRef]
- Hyväri, L.; Vanhatupa, S.; Halonen, H.T.; Kääriäinen, M.; Miettinen, S. Myocardin-Related Transcription Factor A (MRTF-A) Regulates the Balance between Adipogenesis and Osteogenesis of Human Adipose Stem Cells. Stem Cells Int. 2020, 2020, 8853541. [Google Scholar] [CrossRef]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell Shape, Cytoskeletal Tension, and RhoA Regulate Stem Cell Lineage Commitment. Dev. Cell 2004, 6, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.P.; Mearow, K.M. Cell Stress Promotes the Association of Phosphorylated HspB1 with F-Actin. PLoS ONE 2013, 8, e68978. [Google Scholar] [CrossRef] [Green Version]
- Hirano, S.; Shelden, E.A.; Gilmont, R.R. HSP27 Regulates Fibroblast Adhesion, Motility, and Matrix Contraction. Cell Stress Chaperones 2004, 9, 29–37. [Google Scholar] [CrossRef]
- Arrigo, A.-P. Mammalian HspB1 (Hsp27) Is a Molecular Sensor Linked to the Physiology and Environment of the Cell. Cell Stress Chaperones 2017, 22, 517–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakoori, A.R.; Oberdorf, A.M.; Owen, T.A.; Weber, L.A.; Hickey, E.; Stein, J.L.; Lian, J.B.; Stein, G.S. Expression of Heat Shock Genes during Differentiation of Mammalian Osteoblasts and Promyelocytic Leukemia Cells. J. Cell Biochem. 1992, 48, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Hronik-Tupaj, M.; Rice, W.L.; Cronin-Golomb, M.; Kaplan, D.L.; Georgakoudi, I. Osteoblastic Differentiation and Stress Response of Human Mesenchymal Stem Cells Exposed to Alternating Current Electric Fields. Biomed Eng. Online 2011, 10, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, R.; Barbato, E.; Paganelli, C.; Muzio, L. lo Immunolocalization of Heat Shock Protein 27 in Developing Jaw Bones and Tooth Germs of Human Fetuses. Calcif Tissue Int. 2004, 75, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Tiffee, J.C.; Griffin, J.P.; Cooper, L.F. Immunolocalization of Stress Proteins and Extracellular Matrix Proteins in the Rat Tibia. Tissue Cell 2000, 32, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Large Potentials of Small Heat Shock Proteins. Physiol. Rev. 2011, 91, 1123–1159. [Google Scholar] [CrossRef] [Green Version]
- Kainuma, S.; Tokuda, H.; Yamamoto, N.; Kuroyanagi, G.; Fujita, K.; Kawabata, T.; Sakai, G.; Matsushima-Nishiwaki, R.; Kozawa, O.; Otsuka, T. Heat Shock Protein 27 (HSPB1) Suppresses the PDGF-BB-Induced Migration of Osteoblasts. Int. J. Mol. Med. 2017, 40, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Lindroos, B.; Boucher, S.; Chase, L.; Kuokkanen, H.; Huhtala, H.; Haataja, R.; Vemuri, M.; Suuronen, R.; Miettinen, S. Serum-Free, Xeno-Free Culture Media Maintain the Proliferation Rate and Multipotentiality of Adipose Stem Cells in Vitro. Cytotherapy 2009, 11, 958–972. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tirkkonen, L.; Haimi, S.; Huttunen, S.; Wolff, J.; Pirhonen, E.; Sandor, G.K.; Miettinen, S. Osteogenic Medium Is Superior to Growth Factors in Differentiation of Human Adipose Stem Cells towards Bone-Forming Cells in 3D Culture. Eur. Cell Mater. 2013, 25, 144–158. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Galateanu, B.; Dinescu, S.; Cimpean, A.; Dinischiotu, A.; Costache, M. Modulation of Adipogenic Conditions for Prospective Use of HADSCs in Adipose Tissue Engineering. Int. J. Mol. Sci. 2012, 13, 15881–15900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grau-Vorster, M.; Laitinen, A.; Nystedt, J.; Vives, J. HLA-DR Expression in Clinical-Grade Bone Marrow-Derived Multipotent Mesenchymal Stromal Cells: A Two-Site Study. Stem Cell Res. Ther. 2019, 10, 164. [Google Scholar] [CrossRef] [Green Version]
- Ojansivu, M.; Wang, X.; Hyväri, L.; Kellomäki, M.; Hupa, L.; Vanhatupa, S.; Miettinen, S. Bioactive Glass Induced Osteogenic Differentiation of Human Adipose Stem Cells Is Dependent on Cell Attachment Mechanism and Mitogen-Activated Protein Kinases. Eur. Cell Mater. 2018, 35, 54–72. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Zhang, Z.; Huang, H.; Shen, J.; Zhang, S.; Jiang, Y.; Luo, L.; Yin, Z. HSP27 Regulates IL-1 Stimulated IKK Activation through Interacting with TRAF6 and Affecting Its Ubiquitination. Cell Signal 2009, 21, 143–150. [Google Scholar] [CrossRef]
- Huang, C.; Dai, J.; Zhang, X.A. Environmental Physical Cues Determine the Lineage Specification of Mesenchymal Stem Cells. Biochim. Biophys. Acta 2015, 1850, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Moritz, N.; Vedel, E.; Hupa, L.; Aro, H.T. Mechanical Verification of Soft-Tissue Attachment on Bioactive Glasses and Titanium Implants. Acta Biomater. 2008, 4, 1118–1122. [Google Scholar] [CrossRef]
- Hoppe, A.; Guldal, N.S.; Boccaccini, A.R. A Review of the Biological Response to Ionic Dissolution Products from Bioactive Glasses and Glass-Ceramics. Biomaterials 2011, 32, 2757–2774. [Google Scholar] [CrossRef]
- Lakhkar, N.J.; Lee, I.-H.; Kim, H.-W.; Salih, V.; Wall, I.B.; Knowles, J.C. Bone Formation Controlled by Biologically Relevant Inorganic Ions: Role and Controlled Delivery from Phosphate-Based Glasses. Adv. Drug Deliv. Rev. 2013, 65, 405–420. [Google Scholar] [CrossRef]
- Ying, X.; Cheng, S.; Wang, W.; Lin, Z.; Chen, Q.; Zhang, W.; Kou, D.; Shen, Y.; Cheng, X.; Rompis, F.A.; et al. Effect of Boron on Osteogenic Differentiation of Human Bone Marrow Stromal Cells. Biol. Trace Elem Res. 2011, 144, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Miron, R.; Sculean, A.; Kaskel, S.; Doert, T.; Schulze, R.; Zhang, Y. Proliferation, Differentiation and Gene Expression of Osteoblasts in Boron-Containing Associated with Dexamethasone Deliver from Mesoporous Bioactive Glass Scaffolds. Biomaterials 2011, 32, 7068–7078. [Google Scholar] [CrossRef] [PubMed]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-Derived and Bone Marrow Mesenchymal Stem Cells: A Donor-Matched Comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef]
- Lynch, M.P.; Capparelli, C.; Stein, J.L.; Stein, G.S.; Lian, J.B. Apoptosis during Bone-like Tissue Development in Vitro. J. Cell Biochem. 1998, 68, 31–49. [Google Scholar] [CrossRef]
- Ojansivu, M.; Mishra, A.; Vanhatupa, S.; Juntunen, M.; Larionova, A.; Massera, J.; Miettinen, S. The Effect of S53P4-Based Borosilicate Glasses and Glass Dissolution Products on the Osteogenic Commitment of Human Adipose Stem Cells. PLoS ONE 2018, 13, e0202740. [Google Scholar] [CrossRef] [Green Version]
- Amirrah, I.N.; Lokanathan, Y.; Zulkiflee, I.; Wee, M.F.M.R.; Motta, A.; Fauzi, M.B. A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold. Biomedicines 2022, 10, 2307. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.B.; Stein, G.S. Development of the Osteoblast Phenotype: Molecular Mechanisms Mediating Osteoblast Growth and Differentiation. Iowa Orthop. J. 1995, 15, 118–140. [Google Scholar] [PubMed]
- Oosterlaken, B.M.; Vena, M.P.; de With, G. In Vitro Mineralization of Collagen. Advanced Materials 2021, 33, 2004418. [Google Scholar] [CrossRef]
- Jäger, I.; Fratzl, P. Mineralized Collagen Fibrils: A Mechanical Model with a Staggered Arrangement of Mineral Particles. Biophys. J. 2000, 79, 1737–1746. [Google Scholar] [CrossRef] [Green Version]
- Tomoaia, G.; Pasca, R.-D. On the Collagen Mineralization. A Review. Clujul Med. 2015, 88, 15–22. [Google Scholar] [CrossRef]
- Tsigkou, O.; Jones, J.R.; Polak, J.M.; Stevens, M.M. Differentiation of Fetal Osteoblasts and Formation of Mineralized Bone Nodules by 45S5 Bioglass® Conditioned Medium in the Absence of Osteogenic Supplements. Biomaterials 2009, 30, 3542–3550. [Google Scholar] [CrossRef]
- Fan, T.; Qu, R.; Jiang, X.; Yang, Y.; Sun, B.; Huang, X.; Zhou, Z.; Ouyang, J.; Zhong, S.; Dai, J. Spatial Organization and Crosstalk of Vimentin and Actin Stress Fibers Regulate the Osteogenic Differentiation of Human Adipose-Derived Stem Cells. The FASEB Journal 2021, 35, e21175. [Google Scholar] [CrossRef] [PubMed]
- Jonak, C.; Mildner, M.; Klosner, G.; Paulitschke, V.; Kunstfeld, R.; Pehamberger, H.; Tschachler, E.; Trautinger, F. The Hsp27kD Heat Shock Protein and P38-MAPK Signaling Are Required for Regular Epidermal Differentiation. J. Dermatol Sci. 2011, 61, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Wettstein, G.; Bellaye, P.S.; Micheau, O.; Bonniaud, P. Small Heat Shock Proteins and the Cytoskeleton: An Essential Interplay for Cell Integrity? Int. J. Biochem. Cell Biol. 2012, 44, 1680–1686. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Adachi, S.; Matsushima-Nishiwaki, R.; Minamitani, C.; Natsume, H.; Katagiri, Y.; Hirose, Y.; Mizutani, J.; Tokuda, H.; Kozawa, O.; et al. Regulation by Heat Shock Protein 27 of Osteocalcin Synthesis in Osteoblasts. Endocrinology 2011, 152, 1872–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doshi, B.M.; Hightower, L.E.; Lee, J. The Role of Hsp27 and Actin in the Regulation of Movement in Human Cancer Cells Responding to Heat Shock. Cell Stress Chaperones 2009, 14, 445–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graceffa, P. Hsp27-Actin Interaction. Biochem. Res. Int. 2011, 2011, 901572. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.P.; Gonzalez, M.; Rios, S.; Cambiazo, V. Cytoskeletal Organization of Human Mesenchymal Stem Cells (MSC) Changes during Their Osteogenic Differentiation. J. Cell Biochem. 2004, 93, 721–731. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Type | Antibody | Host Species | Dilution | Incubation |
|---|---|---|---|---|
| Primary | anti-β-actin sc-47778 † | mouse | 1:2000 | RT, 2 h |
| Primary | anti-HSP27 (D6W5V) #95357S ‡ | rabbit | 1:1000 | +4 °C, overnight |
| Primary | anti-p-HSP27(S78) #2405S ‡ | rabbit | 1:1000 | +4 °C, overnight |
| Primary | anti-MAPKAPK2 #12155T ‡ | rabbit | 1:1000 | +4 °C, overnight |
| Primary | anti-p-MAPKAPK2 (T334) #3007T ‡ | rabbit | 1:1000 | +4 °C, overnight |
| Primary | anti-p38α sc-728 † | rabbit | 1:100 | +4 °C, overnight |
| Primary | anti-p-p38 MAPK(T180/Y182) #4511S ‡ | rabbit | 1:1000 | +4 °C, overnight |
| Secondary | anti-mouse IgG-HRP (sc-2005) † | goat | 1:2000 | RT, 1h |
| Secondary | anti-rabbit IgG-HRP #7074S ‡ | goat | 1:2000 | RT, 1h |
| Antibody Type | Antibody | Host Species | Dilution | Incubation |
|---|---|---|---|---|
| Primary | anti- collagen type I (ab90395) † | mouse | 1:2000 | +4 °C, overnight |
| Primary | anti-pHSP27 (S78) (ab32501) † | rabbit | 1:500 | +4 °C, overnight |
| Primary | anti-HSP27 (D6W5V) #95357S ‡ | rabbit | 1:500 | +4 °C, overnight |
| Secondary | anti-mouse IgG Alexa fluor 488 (A11029) § | goat | 1:500 | +4 °C, 45 min |
| Secondary | anti-rabbit IgG Alexa fluor 488 (A21206) § | donkey | 1:500 | +4 °C, 45 min |
| - | Phalloidin-TRITC § | - | 1:500 | +4 °C, 45 min |
| - | DAPI § | - | 1:2000 | RT, 5 min |
| Antigen | Surface Protein | Surface Marker Expression hASC (%) | Surface Marker Expression hBMSC (%) | Fluorophore | Manufacturer |
|---|---|---|---|---|---|
| CD11a | Integrin alpha L (Lymphocyte function-associated antigen 1) | 1.5 ± 0.7 | 0.7 ± 0.4 | APC | R&D Systems Inc. Minneapolis. MN. USA |
| CD14 | Lipopolysaccharide receptor | 1.2 ± 0.9 | 5.4 ± 1.8 | PECy7 | BD Biosciences. Franklin Lakes. NJ. USA |
| CD19 | B lymphocyte-lineage differentiation antigen | 1.1 ± 0.9 | 4.3 ± 2.4 | PECy7 | BD Biosciences |
| CD34 | Hematopoietic progenitor cell antigen 1 | 22.2 ± 23.4 | 2.5 ± 0.5 | APC | Immunotools GmbH. Friesoythe. Germany |
| CD45 | RO isoform of leucocyte common antigen | 1.8 ± 0.3 | 7.1 ± 1.4 | APC | BD Biosciences |
| CD73 | Ecto-5′-nucleotidase | 91.1 ± 3.7 | 93.4 ± 3.7 | PE | BD Biosciences |
| CD90 | Thy-1 (T cell surface glycoproteins) | 98.8 ± 1.0 | 88.6 ± 6.5 | APC | BD Biosciences |
| CD105 | SH-2. Endoglin | 94.7 ± 6.9 | 92.9 ± 6.1 | PE | R&D Systems Inc. |
| HLA-DR | Major histocompatibility class II antigen (MHC-II) | 1.3 ± 1.1 | 85.7 ± 8.3 | PE | Immunotools GmbH |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyväri, L.; Vanhatupa, S.; Ojansivu, M.; Kelloniemi, M.; Pakarinen, T.-K.; Hupa, L.; Miettinen, S. Heat Shock Protein 27 Is Involved in the Bioactive Glass Induced Osteogenic Response of Human Mesenchymal Stem Cells. Cells 2023, 12, 224. https://doi.org/10.3390/cells12020224
Hyväri L, Vanhatupa S, Ojansivu M, Kelloniemi M, Pakarinen T-K, Hupa L, Miettinen S. Heat Shock Protein 27 Is Involved in the Bioactive Glass Induced Osteogenic Response of Human Mesenchymal Stem Cells. Cells. 2023; 12(2):224. https://doi.org/10.3390/cells12020224
Chicago/Turabian StyleHyväri, Laura, Sari Vanhatupa, Miina Ojansivu, Minna Kelloniemi, Toni-Karri Pakarinen, Leena Hupa, and Susanna Miettinen. 2023. "Heat Shock Protein 27 Is Involved in the Bioactive Glass Induced Osteogenic Response of Human Mesenchymal Stem Cells" Cells 12, no. 2: 224. https://doi.org/10.3390/cells12020224