
Honokiol Microemulsion Causes Stage-Dependent Toxicity Via Dual Roles in Oxidation-Reduction and Apoptosis through FoxO Signaling Pathway
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Zebrafish Husbandry
2.3. Honokiol Microemulsion Exposure
2.4. Determination of Oxidation Product Content and Antioxidant Enzyme Activity Levels
2.5. Analysis of Gene Expression
2.6. In Vivo Rapid Apoptosis Assay
2.7. PC12 Cell Culture and Treatment
2.8. MTT Assay
2.9. Statistical Analysis
3. Results
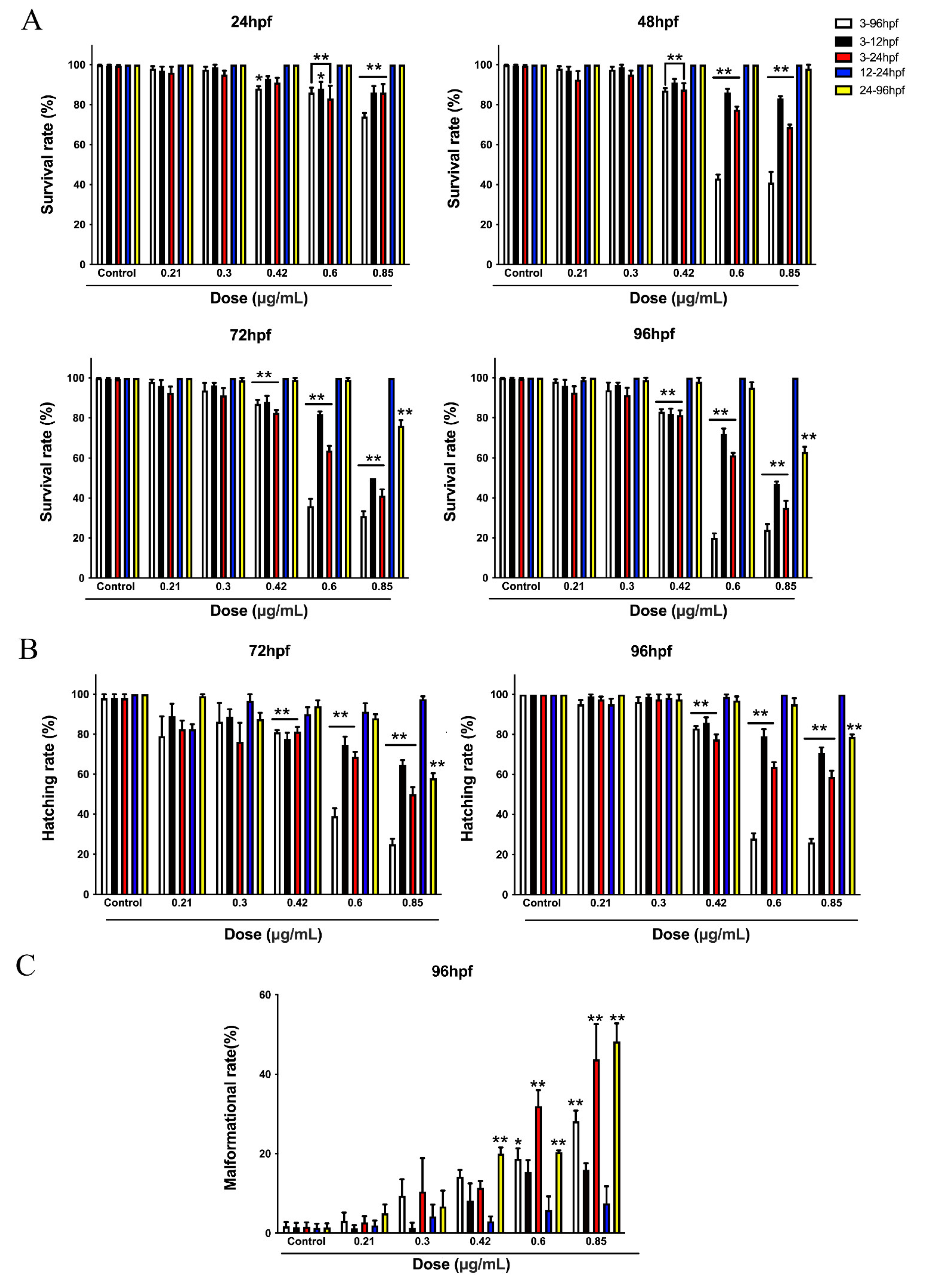
3.1. The Developmental Toxicity of the Honokiol Microemulsion on Zebrafish Embryos Was Stage-Dependent
3.2. Dual Roles of the Honokiol Microemulsion in Oxidative Stress in Zebrafish Embryos
3.3. The Honokiol Microemulsion Induced Apoptosis Mainly in the Zebrafish Brain
3.4. The Honokiol Microemulsion Influenced the mRNA Expression of Apoptosis-Related Genes and Regulatory Genes in Zebrafish Embryos
3.5. Dual Roles of the Honokiol Microemulsion in Oxidation-Reduction in PC12 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rauf, A.; Olatunde, A.; Imran, M.; Alhumaydhi, F.A.; Aljohani, A.S.M.; Khan, S.A.; Uddin, M.S.; Mitra, S.; Emran, T.B.; Khayrullin, M.; et al. Honokiol: A review of its pharmacological potential and therapeutic insights. Phytomedicine 2021, 90, 153647. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Singh, S.; Piazza, G.A.; Contreras, C.M.; Panyam, J.; Singh, A.P. Honokiol: A novel natural agent for cancer prevention and therapy. Curr. Mol. Med. 2012, 12, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Katiyar, S.K. Honokiol, an Active Compound of Magnolia Plant, Inhibits Growth, and Progression of Cancers of Different Organs. Anti-Inflamm. Nutraceuticals Chronic Dis. 2016, 928, 245–265. [Google Scholar] [CrossRef]
- Ong, C.P.; Lee, W.L.; Tang, Y.Q.; Yap, W.H. Honokiol: A Review of Its Anticancer Potential and Mechanisms. Cancers 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishitsuka, K.; Hideshima, T.; Hamasaki, M.; Raje, N.; Kumar, S.; Hideshima, H.; Shiraishi, N.; Yasui, H.; Roccaro, A.M.; Richardson, P.; et al. Honokiol overcomes conventional drug resistance in human multiple myeloma by induction of caspase-dependent and -independent apoptosis. Blood 2005, 106, 1794–1800. [Google Scholar] [CrossRef] [Green Version]
- Zang, H.; Qian, G.; Arbiser, J.; Owonikoko, T.K.; Ramalingam, S.S.; Fan, S.; Sun, S.Y. Overcoming acquired resistance of EGFR-mutant NSCLC cells to the third generation EGFR inhibitor, osimertinib, with the natural product honokiol. Mol. Oncol. 2020, 14, 882–895. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Cai, L.L.; Zhang, X.Y.; Deng, L.Y.; Zheng, H.; Deng, C.Y.; Wen, J.L.; Zhao, X.; Wei, Y.Q.; Chen, L.J. Improved solubility and pharmacokinetics of PEGylated liposomal honokiol and human plasma protein binding ability of honokiol. Int J Pharm 2011, 410, 169–174. [Google Scholar] [CrossRef]
- Gou, M.L.; Dai, M.; Li, X.Y.; Wang, X.H.; Gong, C.Y.; Xie, Y.; Wang, K.; Zhao, X.; Qian, Z.Y.; Wei, Y.Q. Preparation and characterization of honokiol nanoparticles. J. Mater. Sci. Mater. Med. 2008, 19, 2605–2608. [Google Scholar] [CrossRef]
- Gou, M.; Zheng, X.; Men, K.; Zhang, J.; Wang, B.; Lv, L.; Wang, X.; Zhao, Y.; Luo, F.; Chen, L.; et al. Self-assembled hydrophobic honokiol loaded MPEG-PCL diblock copolymer micelles. Pharm. Res. 2009, 26, 2164–2173. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, X.; Cui, W.; Zhang, X.; Li, N.; Chen, J.; Wong, A.W.; Roberts, A. Evaluation of short-term and subchronic toxicity of magnolia bark extract in rats. Regul. Toxicol. Pharmacol. 2007, 49, 160–171. [Google Scholar] [CrossRef]
- Li, N.; Song, Y.; Zhang, W.; Wang, W.; Chen, J.; Wong, A.W.; Roberts, A. Evaluation of the in vitro and in vivo genotoxicity of magnolia bark extract. Regul. Toxicol. Pharmacol. 2007, 49, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Sarrica, A.; Kirika, N.; Romeo, M.; Salmona, M.; Diomede, L. Safety and Toxicology of Magnolol and Honokiol. Planta Med. 2018, 84, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalman, D.S.; Feldman, S.; Feldman, R.; Schwartz, H.I.; Krieger, D.R.; Garrison, R. Effect of a proprietary Magnolia and Phellodendron extract on stress levels in healthy women: A pilot, double-blind, placebo-controlled clinical trial. Nutr. J. 2008, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert Garrison, W.C. Effect of a proprietary magnolia and phello- dendron extract on weight management a pilot, double-blind, placebo- controlled clinical trial. Altern. Ther. Health Med. 2006, 12, 50–54. [Google Scholar]
- Qin, X.; Yin, J.; Zhang, W.; Li, J.; Wen, J.; Chen, S. Acute and subchronic toxicities in dogs and genotoxicity of honokiol microemulsion. Regul. Toxicol. Pharmacol. 2018, 95, 362–370. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, J.; Zhang, W.; An, Q.; Wen, J.; Wang, A.; Jin, H.; Chen, S. Acute and sub-chronic toxicity studies of honokiol microemulsion. Regul. Toxicol. Pharmacol. 2015, 71, 428–436. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Q.; Li, W.; Li, H.; Bao, J.; Yang, C.; Wang, A.; Wei, J.; Chen, S.; Jin, H. Role of Nrf2 in the antioxidation and oxidative stress induced developmental toxicity of honokiol in zebrafish. Toxicol. Appl. Pharmacol. 2019, 373, 48–61. [Google Scholar] [CrossRef]
- Eliaz, I.; Weil, E. Intravenous Honokiol in Drug-Resistant Cancer: Two Case Reports. Integr. Cancer Ther. 2020, 19, 1534735420922615. [Google Scholar] [CrossRef]
- Mattsson, A.; Ulleras, E.; Patring, J.; Oskarsson, A. Albendazole causes stage-dependent developmental toxicity and is deactivated by a mammalian metabolization system in a modified zebrafish embryotoxicity test. Reprod. Toxicol. 2012, 34, 31–42. [Google Scholar] [CrossRef]
- Boix, N.; Teixido, E.; Vila-Cejudo, M.; Ortiz, P.; Ibanez, E.; Llobet, J.M.; Barenys, M. Triclabendazole sulfoxide causes stage-dependent embryolethality in zebrafish and mouse in vitro. PLoS ONE 2015, 10, e0121308. [Google Scholar] [CrossRef]
- Chandra, J.; Samali, A.; Orrenius, S. Triggering and modulation of apoptosis by oxidative stress. Free. Radic. Biol. Med. 2000, 29, 323–333. [Google Scholar] [CrossRef]
- Chen, B.; Cao, X.; Lu, H.; Wen, P.; Qi, X.; Chen, S.; Wu, L.; Li, C.; Xu, A.; Zhao, G. N-(3-oxo-acyl) homoserine lactone induced germ cell apoptosis and suppressed the over-activated RAS/MAPK tumorigenesis via mitochondrial-dependent ROS in C. elegans. Apoptosis 2018, 23, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Kim, M.O.; Choi, Y.H.; Hyun, J.W.; Chang, W.Y.; Kim, G.Y. Butein induces G(2)/M phase arrest and apoptosis in human hepatoma cancer cells through ROS generation. Cancer Lett. 2010, 288, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Klotz, L.-O.; Sánchez-Ramos, C.; Prieto-Arroyo, I.; Urbánek, P.; Steinbrenner, H.; Monsalve, M. Redox regulation of FoxO transcription factors. Redox Biol. 2015, 6, 51–72. [Google Scholar] [CrossRef] [Green Version]
- De Keizer, P.L.; Burgering, B.M.; Dansen, T.B. Forkhead Box O as a Sensor, Mediator, and Regulator of Redox Signaling. Antioxid. Redox Signal. 2011, 14, 1093–1106. [Google Scholar] [CrossRef]
- Facciol, A.; Bailleul, C.; Nguyen, S.; Chatterjee, D.; Gerlai, R. Developmental stage-dependent deficits induced by embryonic ethanol exposure in zebrafish: A neurochemical analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 99, 109859. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Tang, R.; Dodd, A.; Lai, D.; McNabb, W.C.; Love, D.R. Validation of zebrafish (Danio rerio) reference genes for quantitative real-time RT-PCR normalization. Acta Biochim. Biophys. Sin. 2007, 39, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Ustundag, U.V.; Unal, I.; Ates, P.S.; Alturfan, A.A.; Yigitbasi, T.; Emekli-Alturfan, E. Bisphenol A and di(2-ethylhexyl) phthalate exert divergent effects on apoptosis and the Wnt/beta-catenin pathway in zebrafish embryos: A possible mechanism of endocrine disrupting chemical action. Toxicol. Ind. Health 2017, 33, 901–910. [Google Scholar] [CrossRef]
- Tucker, B.; Lardelli, M. A Rapid Apoptosis Assay Measuring Relative Acridine Orange Fluorescence in Zebrafish Embryos. Zebrafish 2007, 4, 113–116. [Google Scholar] [CrossRef]
- Timothy, J.; Shafer, W.D.A. Transmitter ion channel and receptor properties of pheochromocytoma PC12 cells a model for neurotoxicological studies. NeuroToxicology 1991, 12, 473–492. [Google Scholar]
- Shen, J.L.; Man, K.M.; Huang, P.H.; Chen, W.C.; Chen, D.C.; Cheng, Y.W.; Liu, P.L.; Chou, M.C.; Chen, Y.H. Honokiol and magnolol as multifunctional antioxidative molecules for dermatologic disorders. Molecules 2010, 15, 6452–6465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, T.; Xu, W.; Liu, Y.; Qian, K.; Xiong, Y.; Chen, Y. Honokiol Alleviates Methionine-Choline Deficient Diet-Induced Hepatic Steatosis and Oxidative Stress in C57BL/6 Mice by Regulating CFLAR-JNK Pathway. Oxid. Med. Cell Longev. 2020, 2020, 2313641. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-H.; Chang, P.-C.; Chen, C.; Chan, M.-H. Protective and therapeutic activity of honokiol in reversing motor deficits and neuronal degeneration in the mouse model of Parkinson’s disease. Pharmacol. Rep. 2018, 70, 668–676. [Google Scholar] [CrossRef]
- Xia, S.; Lin, H.; Liu, H.; Lu, Z.; Wang, H.; Fan, S.; Li, N. Honokiol Attenuates Sepsis-Associated Acute Kidney Injury via the Inhibition of Oxidative Stress and Inflammation. Inflammation 2019, 42, 826–834. [Google Scholar] [CrossRef]
- Weng, T.I.; Wu, H.Y.; Kuo, C.W.; Liu, S.H. Honokiol rescues sepsis-associated acute lung injury and lethality via the inhibition of oxidative stress and inflammation. Intensive Care Med. 2011, 37, 533–541. [Google Scholar] [CrossRef]
- Ye, J.S.; Chen, L.; Lu, Y.Y.; Lei, S.Q.; Peng, M.; Xia, Z.Y. SIRT3 activator honokiol ameliorates surgery/anesthesia-induced cognitive decline in mice through anti-oxidative stress and anti-inflammatory in hippocampus. CNS Neurosci. Ther. 2019, 25, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Zhai, M.; Li, B.; Liu, Z.; Li, K.; Jiang, L.; Zhang, M.; Yi, W.; Yang, J.; Yi, D.; et al. Honokiol Ameliorates Myocardial Ischemia/Reperfusion Injury in Type 1 Diabetic Rats by Reducing Oxidative Stress and Apoptosis through Activating the SIRT1-Nrf2 Signaling Pathway. Oxid. Med. Cell Longev. 2018, 2018, 3159801. [Google Scholar] [CrossRef]
- Rauf, A.; Patel, S.; Imran, M.; Maalik, A.; Arshad, M.U.; Saeed, F.; Mabkhot, Y.N.; Al-Showiman, S.S.; Ahmad, N.; Elsharkawy, E. Honokiol: An anticancer lignan. Biomed Pharm. 2018, 107, 555–562. [Google Scholar] [CrossRef]
- Hellstrom, M.K.; Ramberg, P. The effect of a dentifrice containing Magnolia extract on established plaque and gingivitis in man: A six-month clinical study. Int. J. Dent. Hyg. 2014, 12, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ye, X.; Wang, L.; Peng, B.; Zhang, Y.; Bao, J.; Li, W.; Wei, J.; Wang, A.; Jin, H.; et al. Embryo-fetal development toxicity of honokiol microemulsion intravenously administered to pregnant rats. Regul. Toxicol. Pharmacol. 2016, 74, 117–122. [Google Scholar] [CrossRef] [PubMed]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Bambino, K.; Chu, J. Zebrafish in Toxicology and Environmental Health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.R.; Zhu, Y.X.; Duan, Q.Y.; Chen, Z.; Wu, F.G. Nanomaterials meet zebrafish: Toxicity evaluation and drug delivery applications. J. Control Release 2019, 311–312, 301–318. [Google Scholar] [CrossRef]
- Bhagat, J.; Zang, L.; Nishimura, N.; Shimada, Y. Zebrafish: An emerging model to study microplastic and nanoplastic toxicity. Sci. Total Environ. 2020, 728, 138707. [Google Scholar] [CrossRef]
- Felix, L.M.; Serafim, C.; Valentim, A.M.; Antunes, L.M.; Campos, S.; Matos, M.; Coimbra, A.M. Embryonic Stage-Dependent Teratogenicity of Ketamine in Zebrafish (Danio rerio). Chem. Res. Toxicol. 2016, 29, 1298–1309. [Google Scholar] [CrossRef]
- Peterson, S.M.; Zhang, J.; Weber, G.; Freeman, J.L. Global gene expression analysis reveals dynamic and developmental stage-dependent enrichment of lead-induced neurological gene alterations. Environ. Health Perspect. 2011, 119, 615–621. [Google Scholar] [CrossRef]
- Zhang, C.; Ojiaku, P.; Cole, G.J. Forebrain and hindbrain development in zebrafish is sensitive to ethanol exposure involving agrin, Fgf, and sonic hedgehog function. Birth Defects Res. A Clin. Mol. Teratol. 2013, 97, 8–27. [Google Scholar] [CrossRef] [Green Version]
- De Esch, C.; Slieker, R.; Wolterbeek, A.; Woutersen, R.; de Groot, D. Zebrafish as potential model for developmental neurotoxicity testing: A mini review. Neurotoxicol. Teratol. 2012, 34, 545–553. [Google Scholar] [CrossRef]
- Heyer, D.B.; Meredith, R.M. Environmental toxicology: Sensitive periods of development and neurodevelopmental disorders. Neurotoxicology 2017, 58, 23–41. [Google Scholar] [CrossRef] [PubMed]
- Domi, E.; Barbier, E.; Augier, E.; Augier, G.; Gehlert, D.; Barchiesi, R.; Thorsell, A.; Holm, L.; Heilig, M. Preclinical evaluation of the kappa-opioid receptor antagonist CERC-501 as a candidate therapeutic for alcohol use disorders. Neuropsychopharmacology 2018, 43, 1805–1812. [Google Scholar] [CrossRef] [PubMed]
- Eijkelenboom, A.; Burgering, B.M.T. FOXOs: Signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 2013, 14, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Furukawa-Hibi, Y.; Kobayashi, Y.; Chen, C.; Motoyama, N. FOXO transcription factors in cell-cycle regulation and the response to oxidative stress. Antioxid. Redox Signal. 2005, 7, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Manolopoulos, K.N.; Klotz, L.O.; Korsten, P.; Bornstein, S.R.; Barthel, A. Linking Alzheimer’s disease to insulin resistance: The FoxO response to oxidative stress. Mol. Psychiatry 2010, 15, 1046–1052. [Google Scholar] [CrossRef] [Green Version]
- Link, W. Introduction to FOXO Biology. Methods Mol. Biol. 2019, 1890, 1–9. [Google Scholar] [CrossRef]
- Ro, S.H.; Liu, D.; Yeo, H.; Paik, J.H. FoxOs in neural stem cell fate decision. Arch. Biochem. Biophys. 2013, 534, 55–63. [Google Scholar] [CrossRef]
- Maiese, K. FoxO Proteins in the Nervous System. Anal. Cell. Pathol. 2015, 2015, 569392. [Google Scholar] [CrossRef] [Green Version]
- Peng, K.; Li, Y.; Long, L.; Li, D.; Jia, Q.; Wang, Y.; Shen, Q.; Tang, Y.; Wen, L.; Kung, H.F.; et al. Knockdown of FoxO3a induces increased neuronal apoptosis during embryonic development in zebrafish. Neurosci. Lett. 2010, 484, 98–103. [Google Scholar] [CrossRef]
- Halasi, M.; Hitchinson, B.; Shah, B.N.; Varaljai, R.; Khan, I.; Benevolenskaya, E.V.; Gaponenko, V.; Arbiser, J.L.; Gartel, A.L. Honokiol is a FOXM1 antagonist. Cell Death Dis. 2018, 9, 84. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Li, W.; Li, J.; Li, S.; Kuang, L.; Pang, F.; Jiang, H.; Jin, H.; Bian, X. Honokiol Microemulsion Causes Stage-Dependent Toxicity Via Dual Roles in Oxidation-Reduction and Apoptosis through FoxO Signaling Pathway. Cells 2022, 11, 3562. https://doi.org/10.3390/cells11223562
Li H, Li W, Li J, Li S, Kuang L, Pang F, Jiang H, Jin H, Bian X. Honokiol Microemulsion Causes Stage-Dependent Toxicity Via Dual Roles in Oxidation-Reduction and Apoptosis through FoxO Signaling Pathway. Cells. 2022; 11(22):3562. https://doi.org/10.3390/cells11223562
Chicago/Turabian StyleLi, Hui, Wanfang Li, Jie Li, Sizheng Li, Lian Kuang, Fei Pang, Haiyan Jiang, Hongtao Jin, and Xiaolan Bian. 2022. "Honokiol Microemulsion Causes Stage-Dependent Toxicity Via Dual Roles in Oxidation-Reduction and Apoptosis through FoxO Signaling Pathway" Cells 11, no. 22: 3562. https://doi.org/10.3390/cells11223562