
Identification of the Telomere elongation Mutation in Drosophila
by
 , , , , , and
, , , , , and
Hemakumar M. Reddy
1,2,
Thomas A. Randall
3,
Francesca Cipressa
4,
Antonella Porrazzo
5,6 ,
,
Giovanni Cenci
5,7,*,
Radmila Capkova Frydrychova
1,8,9,* and
James M. Mason
1 1
Laboratory of Molecular Genetics, National Institute of Environmental Health Sciences, Research Triangle Park, NC 27713, USA
2
Nabsys 2.0, LLC, 60 Clifford Street, Providence, RI 02903, USA
3
Integrative Bioinformatics, National Institute of Environmental Health Sciences, Research Triangle Park, NC 27713, USA
4
Department of Ecological and Biological Sciences, Università degli Studi della Tuscia, 01100 Viterbo, Italy
5
Dipartimento di Biologia e Biotecnologie “C. Darwin”, Sapienza Università di Roma, 00185 Rome, Italy
6
Unit of Molecular Genetics of Complex Phenotypes, Bambino Gesù Children’s Hospital, IRCSS, 00146 Rome, Italy
7
Fondazione Cenci Bolognetti/Istituto Pasteur Italia, 00185 Rome, Italy
8
Institute of Entomology, Biology Centre AS CR, v.v.i., 370 05 Ceske Budejovice, Czech Republic
9
Faculty of Science, University of South Bohemia, 370 05 Ceske Budejovice, Czech Republic
*
Authors to whom correspondence should be addressed.
Cells 2022, 11(21), 3484; https://doi.org/10.3390/cells11213484
Submission received: 7 September 2022
/
Revised: 31 October 2022
/
Accepted: 1 November 2022
/
Published: 3 November 2022
(This article belongs to the Special Issue Heterochromatin Is No Longer the Dark Side of the Genome: A Themed Issue in Honor of Prof. Sergio Pimpinelli)
Abstract
:Telomeres in Drosophila melanogaster, which have inspired a large part of Sergio Pimpinelli work, are similar to those of other eukaryotes in terms of their function. Yet, their length maintenance relies on the transposition of the specialized retrotransposons Het-A, TART, and TAHRE, rather than on the activity of the enzyme telomerase as it occurs in most other eukaryotic organisms. The length of the telomeres in Drosophila thus depends on the number of copies of these transposable elements. Our previous work has led to the isolation of a dominant mutation, Tel1, that caused a several-fold elongation of telomeres. In this study, we molecularly identified the Tel1 mutation by a combination of transposon-induced, site-specific recombination and next-generation sequencing. Recombination located Tel1 to a 15 kb region in 92A. Comparison of the DNA sequence in this region with the Drosophila Genetic Reference Panel of wild-type genomic sequences delimited Tel1 to a 3 bp deletion inside intron 8 of Ino80. Furthermore, CRISPR/Cas9-induced deletions surrounding the same region exhibited the Tel1 telomere phenotype, confirming a strict requirement of this intron 8 gene sequence for a proper regulation of Drosophila telomere length.
1. Introduction
Telomeres in all eukaryotes are functionally similar, although structural differences between some species exist [1]. Linear chromosome ends are not replicated completely, and telomeres must compensate this end replication problem by adding new sequences at the chromosome end. The majority of eukaryotes use a specialized reverse transcriptase, telomerase, which adds a short, tandemly repeated DNA sequence to chromosome ends for telomere elongation [2,3]. Insects in the order Diptera lack both telomerase and the short terminal repeats found in other organisms. In particular, the telomeres of Drosophila melanogaster contain three families of non-long terminal repeat (LTR) retrotransposons, HeT-A, TART, and TAHRE (jointly termed HTT), which transpose specifically to chromosome ends and attach using their 3’ oligo(A) tails [4,5]. Among these three families of elements, HeT-A is most abundant, comprising as much as 80–90% of the total number of elements [4,5]. Telomeric chromatin consists of the HTT elements and different proteins that are bound to them [6]. The rate of transposition of these HTT elements may depend on an equilibrium between the level of their expression and the chromatin-bound proteins [5,7,8].
Telomere elongation may also be accomplished by a recombination-based mechanism, including terminal gene conversion using a neighboring telomere as a template [9,10,11]. This mechanism has been also been observed in yeast and in certain human cancers and immortalized mammalian cells, in which overall telomere length increases in the absence of telomerase activity. This recombination-mediated telomere elongation mechanism is called Alternative Lengthening of Telomeres (ALT) [12,13,14] and tends to be most prevalent in tumors of mesenchymal origins [15]. A recent study showed that ALT may also be found in normal mammalian somatic cells [16].
The genes involved in telomere elongation and the mechanisms of elongation are not well studied in Drosophila. Two independent studies on Drosophila identified the dominant factors Tel [17] and E(tc) [18], which developed telomeres several-fold longer than controls, to the extent that these differences can be observed microscopically in polytene chromosomes. Mutations in Su(var)205, which encodes the HP1a protein, and deficiencies for components of the Ku70-Ku80 complex are also dominant telomere elongation mutations. While Su(var)205 mutants seem to increase both HTT transposition frequencies and terminal gene conversion, and Ku deficiencies increase gene conversion, the specific mechanisms by which telomere elongation occurs in these mutants are not understood [4,5]. Increased expression of Het-A transcripts and elongated telomeres were also found as a consequence of loss of the Drosophila hnRNPA1 homolog, Hrb87F [19], which plays several roles in different processes such as gene expression, organization of the nuclear matrix, and heterochromatin formation. However, whether these effects on telomere elongation are indirect or due to a specific function at chromosome ends is still unclear. Interestingly, the involvement of hnRNPA1 also in telomere regulation of higher eukaryotes [20,21] indicates that it could serve an evolutionarily conserved role at chromosome ends.
Early efforts to map Tel [17] allowed meiotic recombination between the Tel1-bearing chromosome and a multiply marked chromosome; the results located Tel1 at 69 on the genetic map, which translates roughly to 92 on the cytogenetic map. Meiotic mapping of E(tc) [18] indicated that this gene is in the same vicinity. In the present study, we took advantage of the observation that there is no meiotic recombination in Drosophila males. Thus, site-specific genetic recombination induced by double-strand breaks that result from the excision of DNA transposons can be identified [22]. We used both P elements and Minos transposons to induce recombination, which allowed the localization of Tel1 to a 15 kb region in the middle of the right arm of chromosome 3 (3R) at 92A. Whole-genome sequencing resulted in the identification of many single-nucleotide polymorphisms (SNPs) and small insertion/deletion polymorphisms (indels) in the Tel1-bearing genome relative to the reference sequence. Comparison of the Tel1 genomic sequence with a collection of inbred lines of the Drosophila Genetic Reference Panel (DGRP) [23] eliminated all of these SNPs and most of the indels, and mapped Tel1 to a 3 bp deletion (TGT) at 3R:19,366,069-71 in the middle of intron 8 of Ino80. Finally, CRISPR/Cas9-induced deletions that removed TGT and surrounding regions exhibited the Tel1 telomere phenotype, confirming that the middle region of Ino80 intron is indeed required for a proper regulation of telomere length.
2. Materials and Methods
2.1. Mapping by Site-Specific Recombination
Transposon-induced male recombination was performed as per the mating scheme reported earlier [22,24]. The chromosome carrying the Tel1 mutation was marked with two mutations with eye color phenotypes, st and ca. An st Tel ca chromosome was made heterozygous in males with a P element and the Δ2-3 transposase. Recombinant chromosomes bearing either st or ca were collected and put into stocks. Stock generation zero occurs at the time the stock was established, two generations after the homozygous recombinants were obtained. These stocks were maintained for a further 12 generations, flies from homozygous recombinants at generation zero, six, nine, and twelve were collected and frozen for DNA isolation in order to assay the HeT-A copy number over time.
Minos element-induced male recombination mapping is the same as the P element-induced male recombination procedure, except that the P[hsILMiT] transposase was used in place of Δ2-3. The P[hsILMiT]2.4 transposase is under the control of a heat shock promoter; therefore, the larvae generated by the cross of heterozygous st Tel ca/Minos males, were exposed to heat shock at 37 °C in a water bath for 1 hr daily from day two to day six post egg laying [25].
2.2. Genome Sequencing, Mapping to Reference, and De Novo Assembly
DNA was isolated from approximately 30–40 adult flies by standard procedures of lysis, phenol:chloroform extraction, and ethanol precipitation. The DNA pellet was resuspended in TE buffer (10 mM Tris, 0.1 mM EDTA, pH 7.8). DNA quality and concentration were estimated using a Qubit dsDNA BR Assay Kit and measured by Qubit 2.0 Fluorometer (Life Technologies), as per the manufacturer’s protocol. Five micrograms of DNA were taken for library preparation.
All the Illumina genome sequence generated for this project can be found in BioProject Accession PRJNA255315 at NCBI. Genome sequencing was done on an Illumina GA IIx sequencer following standard protocols by the NIH Intramural Sequencing Center. For a detailed step-wise protocol for library preparation and genome sequencing, see Supplementary Information. For Tel, 66,654,840 paired reads of 101 bp length were obtained, and for y w, 76,757,736 reads were obtained, representing a genome coverage of 48X and 55X, respectively. Genomic reads for each strain were mapped to the D. melanogaster reference by two methods. First, all the reads were imported into CLC Genomics Workbench 4.8 and mapped using the parameters Min distance = 150, Max distance = 10,000 to the GenBank-annotated chr3R of dm3 (Release 5 from ftp.ncbi.nih.gov). Second, the same raw reads were mapped to the same reference sequence with bwa 0.6.0 [26] using default settings.
The genomic reads were also assembled de novo by two methods: First, in CLC Genomics Workbench 4.8 using parameters of Min distance = 150, Max distance = 2000, which generated the highest N50 for Tel (28.8 kb); Then, in ABySS 1.2.3 [27], a range of kmers from 25–65 was tested using the Tel sequence, with a kmer setting of 45 generating the highest N50 (45.3 kb). This setting was subsequently used for both genomic assemblies; all other ABySS settings were default.
2.3. Variant Detection
SNPs and indels (referred to as deletion–insertion polymorphisms, DIPs, by CLC Genomics Workbench) were identified from mapped reads in comparison to the reference genome by two methods: First, using CLC Genomics Workbench 4.8 with the SNP and DIP Detection tools at default settings. Separately, the bwa assemblies were imported into CLC as bam files, and both SNP and indel detection were performed on these assemblies, as above. As this software is not trained for detecting large indels (>5 bp), we scanned a large mapped region of 79 kb (chr3R: 19,325,278–19,404,278) manually and identified additional indels, which had not been detected by the CLC Work bench software (QIAGENE, DK-8000 Aarhus C, Denmark). To complement this SNP/indel analysis by CLC, the same assemblies (CLC and bwa) were also analyzed for SNPs and indels using the pileup program of SAMtools 1.6 [28].
Separately, the contigs spanning the 79 kb region of interest were extracted from each of the de novo assemblies and were aligned to the corresponding reference region with MAFFT 6.849 [29] and manually inspected for indels.
2.4. Comparison to DGRP Data
Files containing the SNPs identified in 162 DGRP [23] lines on chr3R were downloaded (Freeze 1, August 2010 release; http://www.hgsc.bcm.tmc.edu/content/drosophila-genetic-reference-panel (accessed on 3 May 2011)), and the chromosomal coordinates of those 159 strains of normal telomere length were compared with the SNPs identified in the Tel genome assembly (SNPs identified by either CLC or pileup). Any SNP identified in the Tel genome that was also identified in the SNP collection from DGRP was ruled out as possibly causing the Tel phenotype.
As there were no indel data for DGRP lines in Freeze 1, each indel found in the Tel genome assembly, as described above, was compared to the DGRP data. For a subset of eight DGRP lines, the Illumina fastq sequence was downloaded from SRA (SRP000694, Lines 40, 85, 177, 321, 352, 405, 426, 802), imported into CLC Genomics Workbench, and assembled to the chr3R reference, as above, and indel detection was performed. Any indel identified in the Tel genome and also found in one or more of these DGRP lines was ruled out as potentially causative. For the indels discovered by manual inspection of both the Tel assembly to reference and the Tel de novo assemblies, a separate local de novo assembly strategy was used for comparison to a subset of the DGRP population. Using 200–300 bp of reference sequence around a candidate indel as bait, BLAT [30] was used to identify individual reads covering this region from the fasta sequence of a given DGRP line. These reads were then extracted, assembled, and compared to both the Tel and reference sequences. Any manually identified indel also found in one or more of these DGRP lines was ruled out as a candidate mutation.
2.5. Real-Time PCR
DNA was isolated from 20–30 flies by using DNeasy Blood & Tissue Kit (Qiagen) columns, as per the manufacturer’s protocol. For large numbers of samples, DNA was isolated from 10 flies of each line using Agencourt DNAdvance Genomic DNA Isolation Kit (Beckman Coulter), as per the manufacturer’s protocol. DNA isolation steps were handled by Biomek 4000 Liquid Handling System (Beckman Coulter robotic system). DNA was eluted in 50 ul water. The DNA concentration was estimated by using NanoDrop 2000 (Thermo Fisher Scientific, CA, USA) and diluted to a concentration of 10 ng/µL, using sterile water.
Primers used for real-time PCR are:
RpS17-F: 5′AAGCGCATCTGCGAGGAG3′,
RpS17-R: 5′CCTCCTCCTGCAACTTGATG3′,
HeT-9D4GAG-ORF-F: 5′TTGTCTTCTCCTCCGTCCACC3′,
HeT-9D4GAG-ORF-R: 5′GAGCTGAGATTTTTCTCTATGCTACTG3′.
Predicted sizes of amplicons are 195 bp for RpS17 and 152 bp for the HeT-9 D4 GAG-ORF. GenBank accession number for HeT-A element 9D4 is X68130 and for RpS17 is M22142 [31,32]. An aliquot of 20 ng of each DNA sample was taken for quantitative PCR using 50 nM of each primer and 5 µL of 2X Power SYBR green PCR Master Mix (Applied Biosystems) in a 10 µL reaction volume. These samples were amplified under the following conditions: 95 °C for 10 min (polymerase activation), followed by 40 cycles containing denaturation at 95 °C for 15 s, and annealing/extension at 60 °C for 1 min. Real-time PCR was run using ABI Prism 7900 HT Sequence detection system (Applied Biosystems).
Competitive threshold (Ct) values for each sample were collected for HeT-A primers (9D4 element GAG ORF) and for control Rps17 (ribosomal protein17) primers. Delta Ct values for each sample were calculated by normalizing HeT-A Ct values to control Ct values and graphed using Microsoft Excel. Each DNA sample was run in triplicate to estimate average Ct values.
2.6. Generation of Crispr/Cas 9-Induced Deletions
To create CRISPR/Cas 9-induced deletions that include the TGT sequence (3R:19,366,069-71) of the Ino80 intron-8 region, we introduced two sgRNAs, each containing a protospacer sequence, into the pCFD4 U6-1 U6-3 tandem gRNAs vector. PAM sequences were selected by using CRISPR Optimal Target Finder online tool (http://targetfinder.flycrispr.neuro.brown.edu/ (accessed on 10 February 2020). Guide sequences were cloned into pCDF4 using a ligation-independent homology-directed cloning strategy by following CRISPR Fly Design protocol (https://www.crisprflydesign.org/grna-expression-vectors/ (accessed on 14 February 2020)). The following primers were designed:
G1 F:
5’-TATATAGGAAAGATATCCGGGTGAACTTCGGGGAAAGAGGGCAAACGAAG
TTTTAGAGCTAGAAATAGCAAG-3’ containing a 5’ U6-1 promoter complementary region followed by a G-N 19/20 sgRNA sequence (in bold) and a 3’ gRNA core complementary sequence.
G2-R:
5’-ATTTTAACTTGCTATTTCTAGCTCTAAAACTCCCTTTGAGGCCTTTCAACGAC
GTTAAATTGAAAATAGGTC-3’ with a 5’ gRNA core, N19/20 sgRNA sequence (in bold), and 3’ U6-3 promoter reverse and complementary sequences.
Oligos were employed to amplify the pCDF4 template following the program: initial denaturation at 98 °C for 30 s; 35 cycles of the denaturation step at 98 °C for 10 s, annealing at 66 °C for 15 s, extension at 72 °C for 30 s; final extension at 72 °C for 5 min. After purification with Nucleospin gel and PCR Clean Up Kit (Macherey-Nagel), the PCR product was combined with linear pCFD4 plasmid, previously digested with BbsI enzyme and gel-purified, in a cloning reaction with Neb Builder HiFi DNA kit. A final pCDF4 vector containing both guides was sent to BestGene Inc (Chino Hills, CA) and injected into y v flies. Transgenic v+ flies bearing the two sgRNAs (y v; gRNAs v+ Ch. II) were selected and then crossed with y w; nos Cas9/Cyo flies, which express Cas 9 only in the germline. The resulting gRNAs/nos Cas9 embryos were injected with the ssDNA template (BiofabResearch, Rome, IT) containing the TGT-specific deletion flanked by homology arms of the corresponding Ino80 intron-8 region to allow Homologous Recombination-mediated repair. The ssDNA sequence is indicated below:
5′GGCATTTGGTGCTTGGTAGCTTGGTAGAATATTGGGGAAAGAGGGCAAACGAAAGGCCGATTAAATACCAATATGAGTTTTTGGTCAAATTATTCCGGTATGCGCCAAATACATGAACGGCAAATACCTTTGTTCTTGTTGAAAGGCCTCAAAGGGAAGGAGACGAGAATAAGGGGCCACACTCCTTCCAATGGTGTTTGAGAA 3′.
To recover lines bearing mutant third chromosomes, single adults resulting from the ssDNA injection (designed as A, B, C, D and F) were crossed with MKRS/TM6B flies and balanced over TM6c by crossing the TM6B progeny with the Apxa/TM6C strain. For each ssDNA-injected fly, we established at least five independent single-chromatid stocks (indicated as A1-5, B1-5, C1-5, etc…). Total DNA from each homozygous line was extracted, PCR-amplified, and then sequenced.
2.7. Chromosome Cytology and Immunostaining
Polytene chromosomes for anti-Hoap immunostaining were prepared as previously described [33] and incubated with rabbit anti-HOAP (1:100). Slides were mounted in Vectashield medium H-1200 with DAPI to stain DNA, and salivary gland preparations were analyzed using a Zeiss Axioplan epifluorescence microscope (CarlZeiss, Oberkochen, Germany), equipped with a cooled CCD (charge-coupled device camera; Photometrics, Woburn, MA, USA). Greyscale digital images were acquired as separate files, which were converted to .psd format, pseudocolored, and merged.
3. Results
3.1. Transposase-Induced Male Recombination Mapping
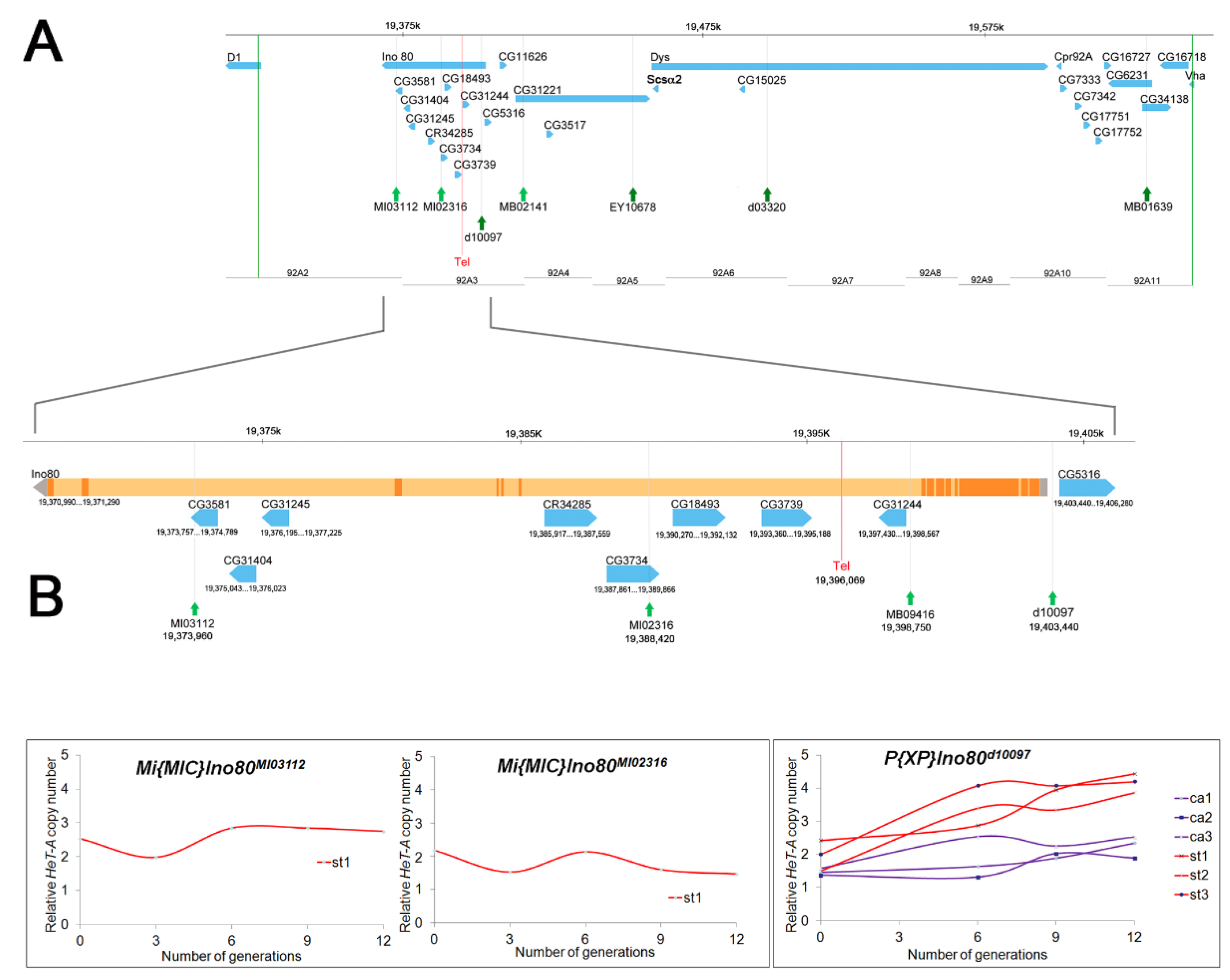
As there is no meiotic recombination in Drosophila males, it is possible to identify site-specific recombination events generated by transposable elements that transpose by a cut-and-paste mechanism and leave a double-strand DNA break in their wake [22,24]. In general, our procedure was similar to previous work [22,24]; in particular, it involved generating heterozygous st Tel1 ca/transposon males, inducing transposition with an exogenous transposase, and recovering recombinant st or ca chromosomes to be tested for the Tel phenotype. Initial efforts to map the Tel1 mutation by male recombination were limited by the paucity of useful transposon insertions in the surrounding chromosomal region and continued as new insertion chromosomes became available. The assay used to identify Tel1 on the recombinant chromosomes changed over time. The assay used in early recombination experiments (Table 1, Round 1) used a cytogenetic analysis of heterozygous chromosomes exposed to Tel1 for two years [17]. Later, relative telomere length was estimated by measuring the relative copy number of the open reading frame (ORF) of HeT-A at zero, six, nine, and twelve generations after a recombinant stock was established [31,32]. The initial round of mapping, using seven P element insertions lying in the 91–93 cytogenetic region (Table 1), showed that Tel1 mapped between the P[PZ]Dl05151 and P[SUPor-P]CG16718KG06218 transposon insertion sites (hereafter, we refer to the transposon insertions simply with their allele designation; full names are listed in Table 1). The physical location is 3R: 19,326,218 to 3R 19,641,774, a region of 316 kb. This region showed a surprising paucity of P element insertions.
As new P element insertions became available, we used three transposons lying within this 316 kb region: P[XP]Ino80d10097, P[EPgy2]CG31221EY10678, and P[XP]Dysd03320 (Table 1, Round 2; Figure 1A). All three st-bearing recombinant chromosomes generated using P element insertion d10097 showed telomere elongation from generation 0 to 12, whereas the three ca-bearing recombinant chromosomes from the same P element did not show significant telomere elongation (Figure 1B). Thus, Tel1 lies to the left of this P element insertion (3R:19,403,413). Similar results were obtained for recombinants from P element insertions EY10678 and d03320 (see Figure S1), both of which are to the right of d10097 (Figure 1A). These results mapped Tel to 3R: 19,326,218 to 19,403,413, a region of ~77 kb.
Minos elements were also used to induce recombination (Table 1), although Minos elements had not previously been shown to induce recombination in males. Two Minos insertions, MB02141 and MB0163, lying to the right of d10097 in the 316 kb region showed similar results (see Figure S2), indicating that they are situated to the right of Tel1, as expected. Two Minos insertions in the 77 kb region were selected for further mapping studies (Table 1, Round 3; Figure 1). Even after heat shock, these Minos elements generated only a few recombinant males. We obtained only one st-bearing recombinant chromosome from each of these Minos transposons (Table 1). The st recombinants for MI03112 and MI02316 showed no evidence of telomere elongation from generation zero through twelve (Figure 1B). This result eliminated the region to the left of MI02316 as containing Tel1 and mapped Tel1 to a ~15 kb region (3R: 19,388,379 to 3R:19,403,413).
3.2. Telomere Length in Transposon Insertion Lines
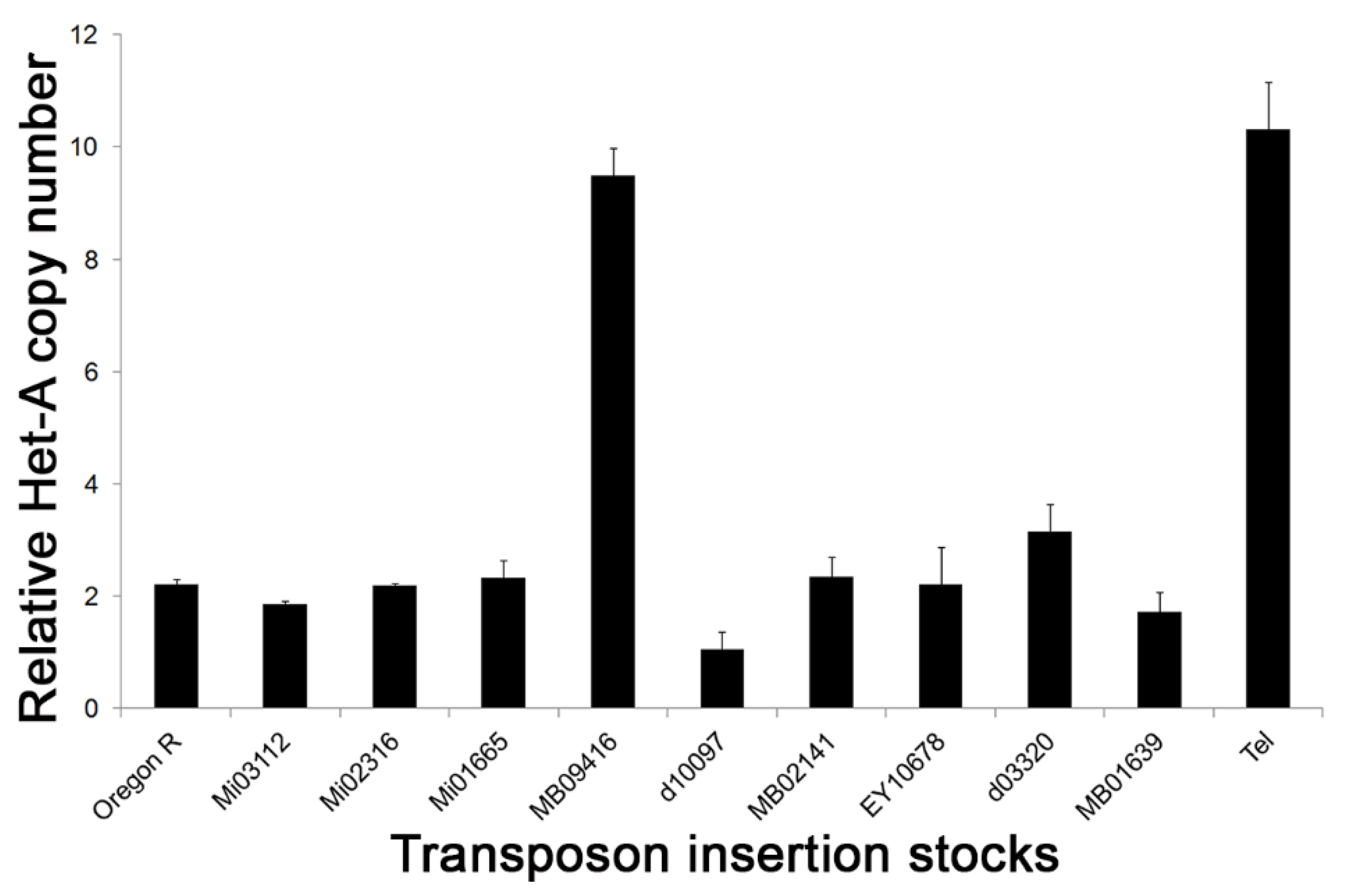
We measured the relative HeT-A copy number in the transposon lines used to induce site-specific recombination. Q-PCR analysis showed that all of the lines, except one bearing MB09416, had telomeres comparable in length to the Oregon-R control (Figure 2). The relative HeT-A copy number in the MB09416 insertion stock was highly elevated and similar to that of Tel1. The MB09416 Minos element is inserted at 3R: 19,398,726, which is in intron 8 of Ino80 and within the 15 kb Tel1 region identified above. UCSC genome browser maps indicate that the insertion site lies in a not well-conserved sequence (Figure S3). As it is likely that the high HeT-A copy number in this line might interfere with the ability to observe an increase in telomere length, this transposon was not used in the mapping of Tel1. It is also possible that the genome of the MB09416 line carries a genetic factor, either at the insertion site or elsewhere, and that it has a phenotype similar to that of Tel1 and might therefore confound the analysis.
3.3. Effect of Tel Copy Number on Telomere Length
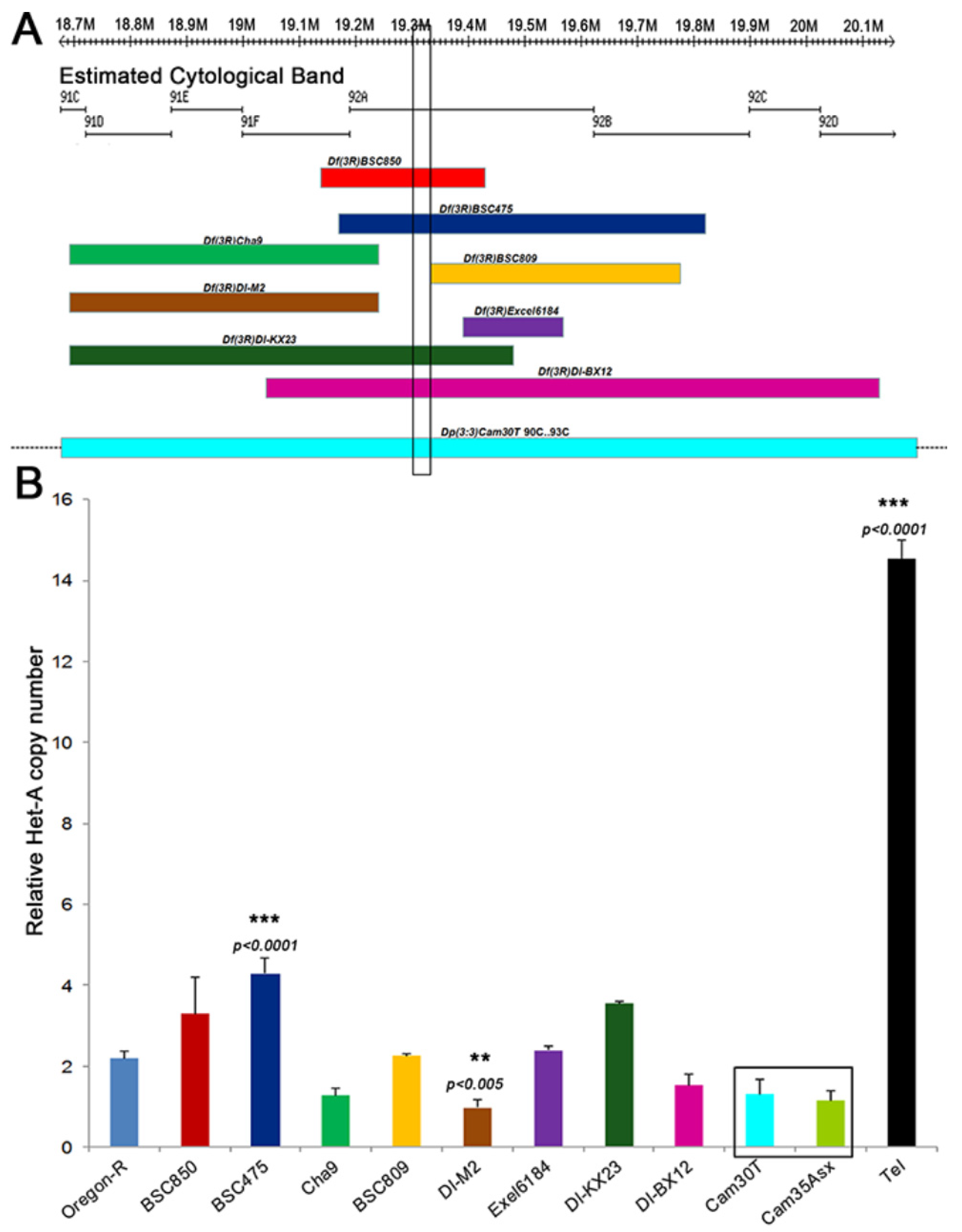
Different deficiencies and duplications spanning region 92A3 (Figure 3A), where Tel was mapped, were analyzed for an effect on telomere length by measuring the relative HeT-A copy number by Q-PCR in the respective stocks. With the exception of Df(3)BSC475 and Df(3R)DI-M2, which induced as yet unclear moderate increase and decrease in Het-A genomic copies (p < 0.001), respectively, none of the deficiency and duplication stocks showed a substantial accumulation of telomeric HeT-A copy number (Figure 3B). Thus, it appears that neither a 50% increase nor a 50% decrease in the copy number of the region around Tel had an effect on telomere length.
3.4. SNP and Indel Identification
The genomes of the three strains Tel1, y1 w1, and E(tc) were sequenced using the Illumina GAIIx platform. As Tel1 appeared in a natural strain (caught in Endine Gaiano, near Bergamo, Italy) that had not been outbred to any laboratory flies [17], there was no useful wild-type control. E(tc), however, appeared in a y1 w1 laboratory stock [18]; therefore, we used a y1 w1 stock as a wild-type control. Upon sequencing, the E(tc) genome appeared to be highly heterozygous. We therefore assumed the stock was contaminated and did not pursue it further. The other two stocks were sequenced to 48X and 55X coverage, respectively. Both genomes were assembled to reference using CLC Genomics Workbench and bwa. In addition, two de novo assemblies, using CLC Genomics Workbench and ABySS, were generated. Concurrently with the Minos transposon recombination mapping, a 79 kb genomic region of 3R: 19,325,278 to 19,404,278, roughly the region between inserts 05151 and d10097, extended slightly on either end, was analyzed for SNP and indel variations with the above assemblies. Within this region, there were 626 SNPs and 88 indels identified on the Tel1 chromosome compared to the reference using the CLC Genomics SNP and DIP Detection analysis (Table 2). A similar number of variations (586 SNPs and 80 indels) were also found in the y1 w1 control strain. After eliminating common variations between Tel1 and y1 w1 in this region, we are left with 332 SNPs and 53 indels that appear to be unique to the Tel1 genome in this 79 kb region.
Current variant calling tools are only proficient at defining small indels (1–5 bp) [34,35]. To detect larger indels, we scanned Tel1 genomic assemblies manually and detected 13 polymorphisms of 5 bp or larger (Table 2). These large indels, present in the Tel1 genome but not in y1 w1, were analyzed by PCR with primers flanking these indels and by Sanger sequencing of PCR products (Supplementary Information, see Figure S4). A limitation of the assembly to reference strategy for variant identification is that potential novel insertions that are not present in the reference sequence are not detected by this approach. To search for such variations, we aligned the de novo assemblies of the Tel1 genome to the reference. Manually scanning this alignment identified an additional 14 insertions not found by the above methods (Table 2).
3.5. Comparison of Variations to DGRP Data
To differentiate natural polymorphisms among these remaining SNP and indel variations found in the Tel1 genome, we compared them with the genomes available from the DGRP [23], a collection of wild-caught, inbred Drosophila strains whose genomes have been sequenced. Our hypothesis is that, if any variant found in the Tel1 genome is also found in a DGRP line with normal-length telomeres, that variant can be ruled out as causing the Tel phenotype. As a first step, all DGRP lines were tested for the relative HeT-A copy number as a proxy for telomere length. The HeT-A copy number data for these strains fit a log-normal distribution, with three outliers that had copy numbers higher than the Tel1 strain (Figure 4). These three lines, RAL-161, -703, and -882, have been excluded from the following discussion and are described elsewhere [32].
As the remaining DGRP lines have what we consider to be normal telomere length, close to that found in the Oregon-R control, the SNPs identified from Freeze1 (August 2010) of the DGRP lines [23] were compared to the SNPs in the Tel1 genome identified by the CLC SNP detection software. All the SNPs found in the 79 kb region around Tel1 were also found in the DGRP lines (Table 2). Thus, all the SNPs found in the Tel1 genome are natural polymorphisms with little expected effect on telomere elongation. No indel data were available for DGRP lines in Freeze1. We therefore identified indels in a selection of eight DGRP lines for the 79 kb region of interest. All except two indels found on the Tel1 chromosome, a deletion of C at 3R:19,352,437, and a deletion of TGT at 3R:19,396,067-69 were also found in one or more lines from the DGRP collection (Table 2). The deletion of C is located in a large intergenic region, 11 kb to the right of Dl and more than 16 kb to the left of CG43203, while the deletion of TGT is located in intron 8 of Ino80. The latter is the only indel-specific to the Tel1 genome in the 15 kb region of interest and therefore was identified as the Tel1 mutation.
3.6. Comparison to modENCODE Data
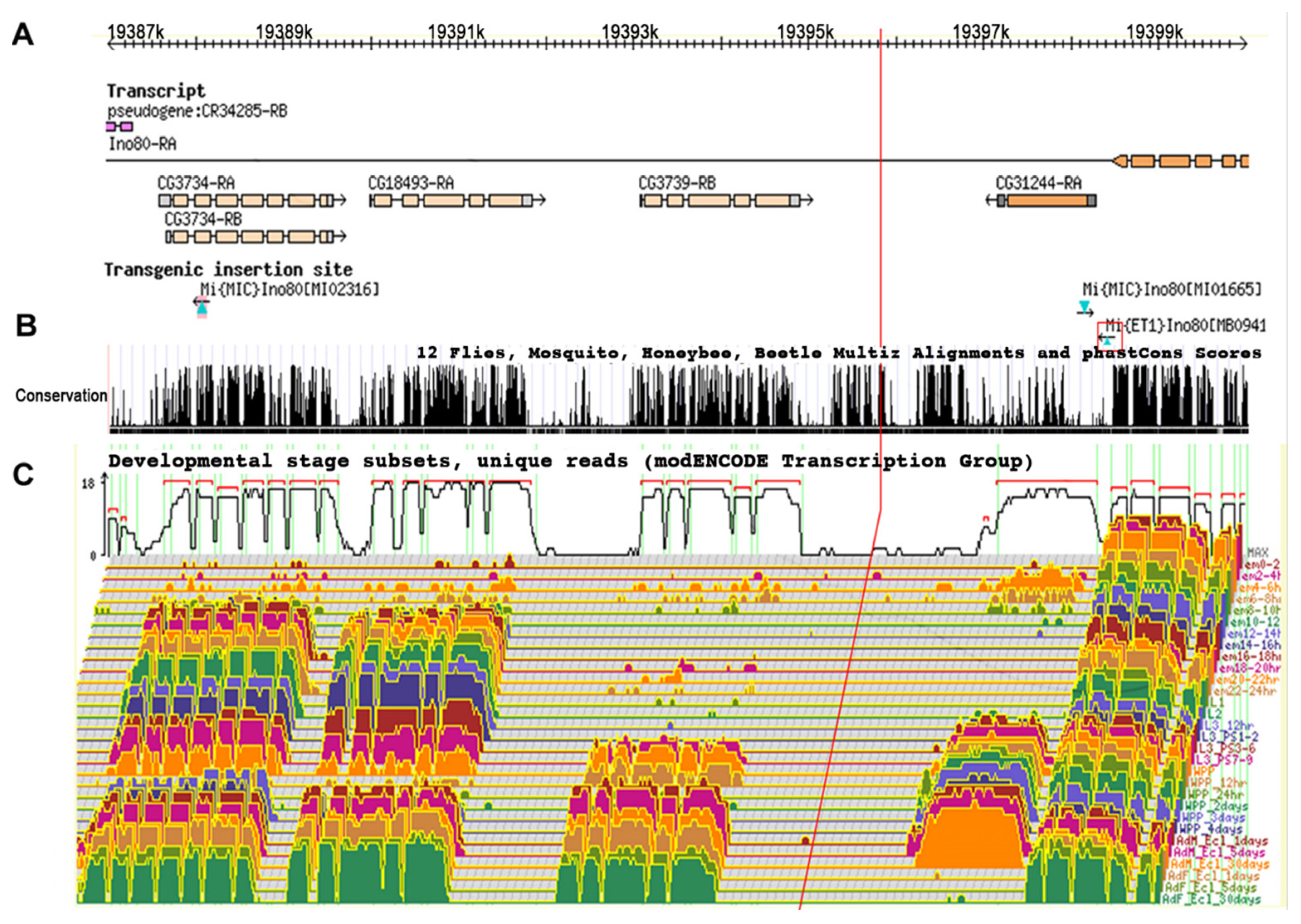
RNA sequence coverage for the 15 kb Tel region was analyzed by comparison with the modENCODE database, including stage and tissue-specific transcript expression levels. This analysis shows that the candidate Tel1 mutation was not included in a transcript at any stage in any tissue (Figure 5) and suggests that Tel1 could be acting to alter the expression of other transcripts near or within the Ino80 locus. The UCSC Genome browser map for this 15 kb region was examined for sequence conservation among Drosophila species and other insect species. This analysis shows that, even though the candidate Tel1 mutation is noncoding, it is in a well-conserved region, similar in the level of conservation to neighboring coding regions (see Figure S3).
3.7. Transcript Analysis
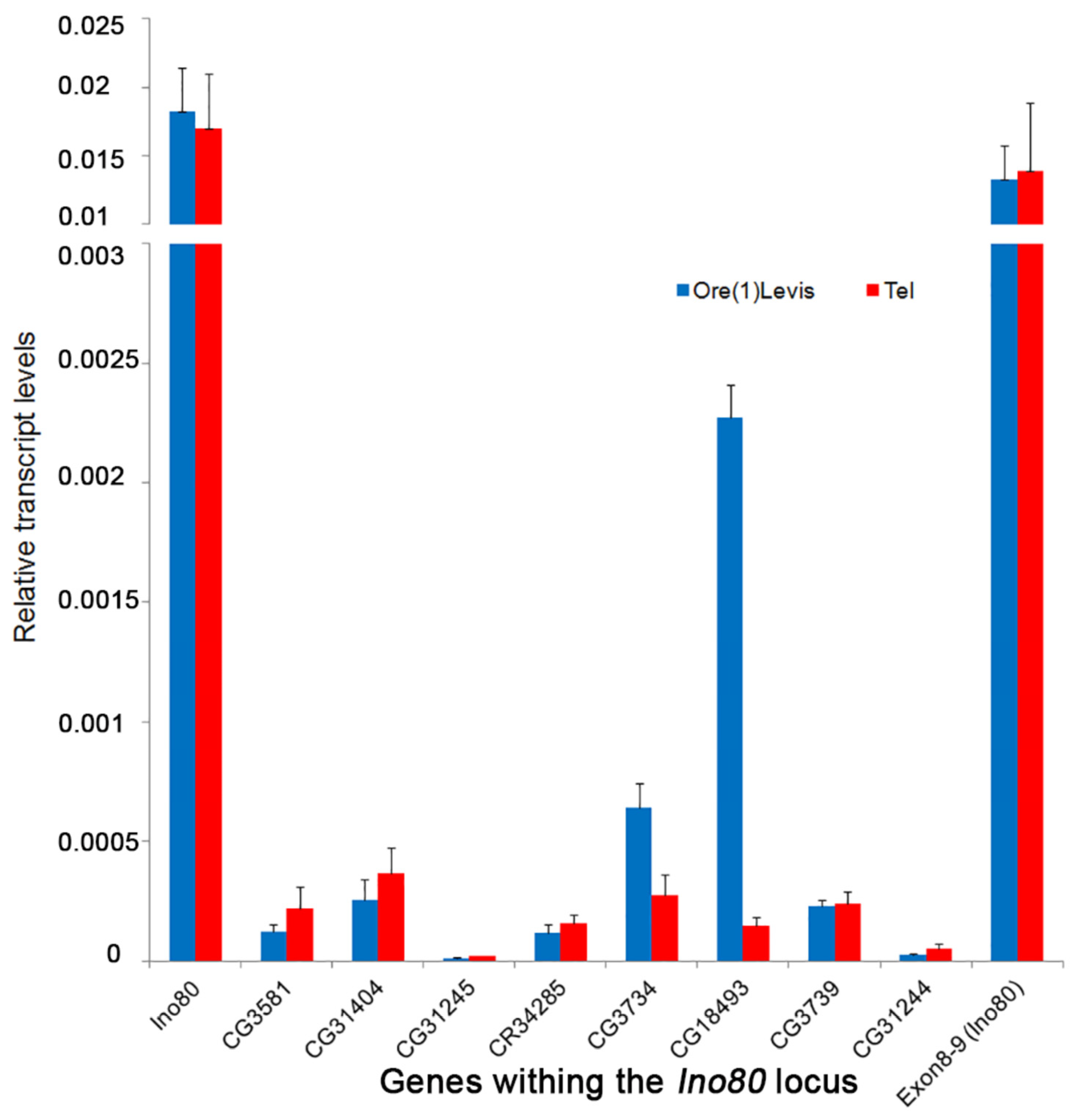
The transcript levels from the ovaries of Tel1 and Oregon-R were analyzed for nine genes in the vicinity of the Tel1 mutation (TGT deletion), which include Ino80 and those found within its introns. Quantitative PCR with cDNA from these two lines showed that there is no significant difference (p > 0.05) in the expression levels between the two strains for most of these genes (Figure 6). The transcript level of Ino80 around the exon 8–9 junction that spans the intron 8, where Tel1 mutation is located, also showed an intact transcript with normal expression similar to other parts of the transcript, indicating that the Tel1 mutation does not interfere with local splicing (Figure 6). CG18493, however, showed a 15-fold lower expression in Tel1 compared to the control (p = 0.0006), and CG3734 showed a slight reduction in expression in Tel1 ovaries (p = 0.0103). Given that a 50% reduction in the Tel+ copy number appears to have no effect on telomere length (Figure 3), that after a Bonferroni correction the small decrease seen for CG3734 expression (Figure 6) may not be considered significant, and the expression of CG18493 was not statistically reduced in CRISPR/Cas9-induced Tel alleles described below, it seems likely that expression of both CG18493 and CG3734 genes is not relevant for the Tel phenotype.
3.8. Generation of CRISPR/Cas 9-Induced Tel1 Deletion Alleles
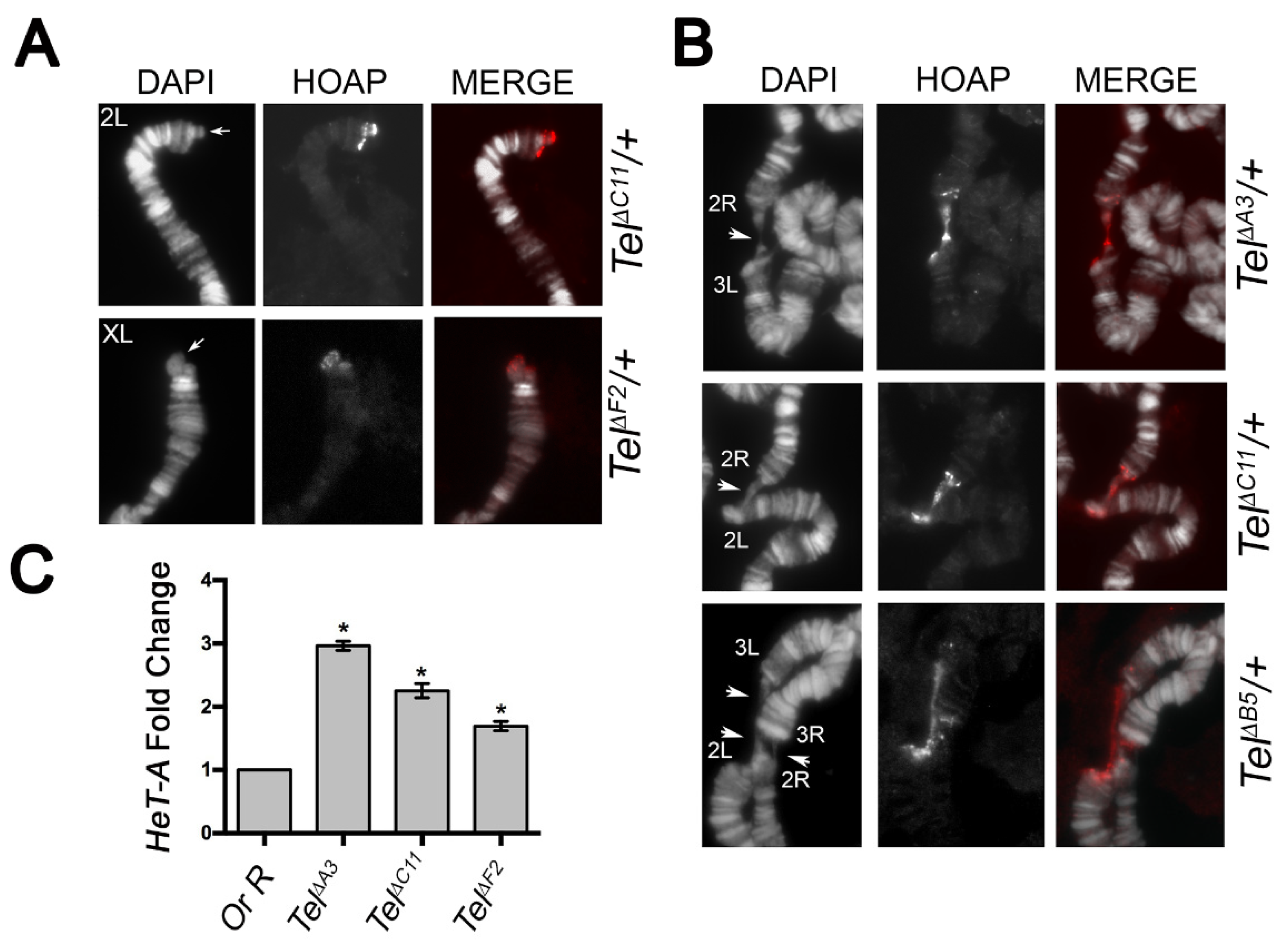
To ask whether a deletion of the TGT sequence in the Ino80 intron 8 yields a Tel phenotype, we sought to use CRISPR/Cas9-mediated homology repair to induce the 3 nt deletion, as well as other small deletions encompassing TGT, and check whether these deletions elicited elongated and/or fused telomeres. By using a 205bp ssDNA donor as the template bearing the TGT deletion (see Methods), we obtained 22 potential viable deletions that were established as independent stocks by September 2020. Sequence analysis revealed that these lines identified 10 distinctive small deletions of different sizes ranging from 6 to 222 bps, but none of them identified the 3 nt TGT deletion only (Figure S5). We also noticed that while seven deletions (namely ΔA3, ΔA4, ΔB5, ΔC11, ΔD5, ΔF2, and ΔF5) uncovered the TGT sequence, the ΔC6, ΔC10, and ΔG3 deletions removed small sequences of 12, 11, and 22 nt, respectively, adjacent to the expected 5′ break site, without including TGT (Supplementary Information, Figure S5). Moreover, whereas the 3′ junction sequences of all deletions and the 5′ junction sequences of ΔC4, ΔF5, and ΔG3 were “clean” breaks, the 5′ breakpoint regions of the remaining deletions contained either putative microhomology sequences (ΔA4, ΔC10, and ΔF2) or stretches of AATATTGG repeats that, in the case of ΔA3, ΔA4, and ΔD5, replaced the TGT-containing region (Supplementary Information, Figure S5). We then asked whether any of these deficiency-bearing lines exhibited a Tel phenotype. Three months after the establishment of stocks bearing the deletions (December 2020), by immunostaining for the telomeric specific marker HOAP [36], we found that polytene chromosomes from heterozygotes for each deletion elicited neither elongated telomeres nor telomere associations, which are normally diagnostic of the Tel1 phenotype [17]. However, when we repeated the cytological characterization after 6 months (March, 2021), we observed that, in the same heterozygotes, telomeres were longer than the wild-type and underwent fusions as expected for Tel1 mutants (Figure 7). Finally, our Q-PCR analysis revealed that these deletion-bearing lines also show a statistically significant increase in the HeT-A copy number after 6 months, thus confirming that these lines represent bona fide new CRISPR/Cas 9-induced Tel alleles (Figure 7C). Interestingly, the finding that deletions which do not enclose TGT also display elongated and fused telomeres indicates that a proper Tel function requires intact regions within the Ino80 intron beyond TGT. Thus, although our CRISPR/Cas 9 approach failed to induce a specific 3nt TGT deletion, it allowed the identification of additional sequences in the Ino80 intron 8 that are required for the regulation of telomere length.
4. Discussion
4.1. Mechanism of Telomere Elongation
The stability of telomeres in Drosophila depends on terminin and non-terminin telomeric proteins [9,37]. The terminin proteins Moi, Ver, HipHop, and HOAP are found only at telomeres, whereas non-terminin proteins HP1, the ATM, and ATR kinases, and the proteins of MRN complex also have biological roles apart from their involvement in telomere maintenance and structure [9,37]. Mutations in any of the genes encoding these proteins cause telomere fusions and abnormal cell divisions. However, only mutations in the HP1-encoding gene Su(var)205 [38,39], Hrb87F [19], and the Ku70/Ku80 complex are associated with telomere elongation. The exact mechanism and the genes involved in telomere length homeostasis in Drosophila are largely unknown. There are reports of RNAi control over HeT-A, TART, and TAHRE transcript levels (reviewed in [5]), but the exact mechanism of its involvement in telomere length homeostasis is not well-characterized. Two dominant mutations, Tel and E(tc), showed telomere elongation [17,18]. However, whereas the E(tc) mutation is associated with elevated rates of gene conversion in telomeric regions, the Tel mutation was previously associated with both transposition of the telomeric retrotransposons and gene conversion [40] but not with the transcription of telomeric retrotransposons [41]. The data thus suggest the involvement of Tel in a recombination pathway. Interestingly, the original Tel1 mutation and our CRISPR/Cas 9-induced Tel deletion alleles have no known phenotype other than telomere elongation and end-to-end attachment in polytene chromosomes [17,42] (this work). Thus, the understanding of the molecular mechanisms that underpin the Tel phenotype and the identification of a candidate Tel gene will provide us fundamental insights into how Drosophila regulate telomere length independently of telomere capping and maintenance. Moreover, the molecular characterization of Tel could also contribute to the understanding of recombination-mediated telomere maintenance mechanisms, such as the ALT pathway found in some human cancer cells. Although the ALT pathway is a favorite mechanism of telomere maintenance for some human cancers, the molecular details remain still unknown [12,43].
4.2. Mapping Tel Using Transposon-Induced Male Recombination
The Tel1 mutation in Drosophila was previously localized by meiotic recombination to 69 on the genetic map [17]. Meiotic recombination in Drosophila occurs only in females, but it is possible to induce site-specific recombination in males using transposons and use this for mapping [22,44]. A collection of more than 15,000 publicly available P element insertions means that, in many regions, a resolution of 5–10 kb is possible for P element-induced recombination [45,46].
One major drawback of the P element, however, is its strong bias for insertion into some genes (hot spots) and against insertion into others (cold spots). The region around Tel is a cold spot for P element insertions. Minos, a member of the Tc1/Mariner family of transposable elements, is active in diverse organisms and cultured cells; it produces stable integrants in the germ line of several insect species, in the mouse, and in human cells [25]. To expand the usefulness of transposon mapping in Drosophila, collections of other transposable elements with different insertional specificities, such as Minos [47,48], have been introduced [46]. Minos elements were found to exhibit a generally uniform distribution in the genome [49]. We used available Minos elements to refine our mapping of the Tel1 mutation and show for the first time that these transposons can induce recombination events useful for this purpose. This approach localized Tel1 to a region of 15 kb.
4.3. Genome Sequencing and DGRP Resources for Tel1 Mapping
The molecular lesion associated with Tel1 was identified by deep sequencing of the Tel1 genome and analyzing this sequence for novel SNP and indel variants not found in the DGRP lines [23]. After comparing the variants in the genome bearing Tel1 with DGRP polymorphisms, we ruled out all SNPs and all but one indel in intron 8 of Ino80 as candidates for Tel1. Thus, the combination of formal genetics and next-generation sequencing resulted in the identification of the molecular defect in Tel as a 3 bp deletion (TGT) at 3R:19,396,067-69. To our knowledge, this is the first study using the DGRP collection to map a Mendelian trait in D. melanogaster. The Tel1 mutant used in this study was caught near Endine Gaiano, Bergamo, in northern Italy, likely prior to 1946 [17]. It is of interest that the natural genetic diversity captured by DGRP in a Raleigh, North Carolina, population was of sufficient diversity to identify all of the SNPs and all but two indels within our 79 kb region defined by the transposon mapping. This suggests that the DGRP is an important general resource for genetic mapping of genes in Drosophila melanogaster, even from strains not closely related to the standard reference isolates. CRISPR/Cas9-induced deletions in the intronic region encompassing TGT also resulted in a Tel-specific phenotype, confirming that intron 8 of Ino80 contains the Tel locus. Telomere elongation was evident 6 months, but not 3 months, after the induction of deletions. Consistent with our previous characterization [17], we speculate that this difference in telomere length depends on the progressive accumulation of HTT genomic copies through recombination and/or gene conversion events over time. The Drosophila Ino80 is the ortholog of the human INO80 ATPase, a member of the SNF2 family of ATPases that functions as an integral component of a multisubunit ATP-dependent chromatin remodeling complex [50]. In flies, Ino80 is involved in the regulation of homeotic gene expression and regression of ecdysone-dependent transcription [51,52]. However, our genetic and transcript analyses exclude the possibility that Tel1 is an Ino80 mutant allele that specifically affects telomere length maintenance. The reason why the TGT deletion and all additional deletions surrounding TGT recovered by CRISPR/Cas9 yielded elongated telomeres still remains unclear. We can speculate that Tel specifies a regulatory element in the intron 8 of Ino80 required for telomere homeostasis. One possibility is that it may encode a still unannotated ncRNA (i.e., microRNA), which could influence the expression of genes required for the maintenance of chromosome end length. Moreover, this sequence may act as a binding platform for trans-acting factors, which are required for the activity of genes that prevent recombination events at telomeres. Although these genes are not known, we can rule out that the genes located in the intron 8 are potential candidates, as their expression is not affected by Tel1 mutations. Alternatively, we can envisage that the TGT-containing sequence could regulate appropriate transcript maturation (i.e., working as a splicing enhancer) and that its loss could affect normal splicing of adjacent exons and generate an aberrant gain of function Ino80 isoform with a dominant effect. Whatever the hypotheses, further genetic and transcriptomic studies are necessary to reveal the entire sequence that identifies Tel and the genes regulated by Tel.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cells11213484/s1, Figure S1: Telomere length in recombinants from transposons used for Tel1 mapping; Figure S2: FlyBase browser map of the 77kb Tel1 region with indels and Minos insertion sites; Figure S3: Sequence conservation among insect species at Tel1 and MB09416 insertion loci. Figure S4: PCR with flanking primers to large indels found in Tel genome. Figure S5: Schematic map of CRISPR/Cas 9-induced deletions.
Author Contributions
Conceptualization, H.M.R., R.C.F. and J.M.M.; methodology, H.M.R., T.A.R., F.C., A.P.; software, H.M.R., T.A.R., R.C.F.; validation, H.M.R., T.A.R., G.C., R.C.F. and J.M.M.; formal analysis, H.M.R., F.C., A.P.; investigation, H.M.R., T.A.R., F.C., A.P.; resources, J.M.M.; data curation, H.M.R., R.C.F. and J.M.M.; writing—original draft preparation, G.C., R.C.F. and J.M.M.; writing—review and editing, H.M.R., G.C., R.C.F. and J.M.M.; supervision, G.C., R.C.F. and J.M.M.; funding acquisition, G.C., R.C.F. and J.M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Institute Pasteur of Paris (PTR-24-2017) and the Institute Pasteur of Rome (A. Tramontano 2018) to G.C.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All Illumina genome sequence generated for this project can be found in BioProject Accession PRJNA255315 at NCBI. All Supplementary Materials are available in the Supplementary Information. The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
Transposon insertion stocks, transposase stocks and DGRP stocks received from the Bloomington Drosophila Stock Center at Indiana University were greatly appreciated. We thank Frank Day for support with computational infrastructure, David Fargo for preliminary work on this project and Sailesh Surapureddi for help with real-time PCR.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Frydrychova, R.C.; Mason, J.M. Telomeres: Their Structure and Maintenance. In The Mechaisms of DNA Replication; Stuart, D., Ed.; InTech, Open Access Publisher: Rijeka, Croatia, 2013; pp. 423–443. [Google Scholar]
- Lansdorp, P. Telomere Length Regulation. Front. Oncol. 2022, 12, 943622. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, A.; de Lange, T. Removal of shelterin reveals the telomere end-protection problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capkova Frydrychova, R.; Biessmann, H.; Mason, J.M. Regulation of telomere length in Drosophila. Cytogenet. Genome Res. 2008, 122, 356–364. [Google Scholar] [CrossRef] [Green Version]
- Cacchione, S.; Cenci, G.; Raffa, G.D. Silence at the End: How Drosophila Regulates Expression and Transposition of Telomeric Retroelements. J. Mol. Biol. 2020, 432, 4305–4321. [Google Scholar] [CrossRef] [PubMed]
- Andreyeva, E.N.; Belyaeva, E.S.; Semeshin, V.F.; Pokholkova, G.V.; Zhimulev, I.F. Three distinct chromatin domains in telomere ends of polytene chromosomes in Drosophila melanogaster Tel mutants. J. Cell Sci. 2005, 118, 5465–5477. [Google Scholar] [CrossRef] [Green Version]
- Silva-Sousa, R.; Varela, M.D.; Casacuberta, E. The Putzig partners DREF, TRF2 and KEN are involved in the regulation of the Drosophila telomere retrotransposons, HeT-A and TART. Mob. DNA 2013, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Takacs, S.; Biessmann, H.; Reddy, H.M.; Mason, J.M.; Torok, T. Protein interactions on telomeric retrotransposons in Drosophila. Int. J. Biol. Sci. 2012, 8, 1055–1061. [Google Scholar] [CrossRef]
- Mason, J.M.; Frydrychova, R.C.; Biessmann, H. Drosophila telomeres: An exception providing new insights. Bioessays 2008, 30, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Roth, C.W.; Kobeski, F.; Walter, M.F.; Biessmann, H. Chromosome end elongation by recombination in the mosquito Anopheles gambiae. Mol. Cell Biol. 1997, 17, 5176–5183. [Google Scholar] [CrossRef] [Green Version]
- Kahn, T.; Savitsky, M.; Georgiev, P. Attachment of HeT-A sequences to chromosomal termini in Drosophila melanogaster may occur by different mechanisms. Mol. Cell Biol. 2000, 20, 7634–7642. [Google Scholar] [CrossRef]
- Apte, M.S.; Cooper, J.P. Life and cancer without telomerase: ALT and other strategies for making sure ends (don’t) meet. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.A.; Reddel, R.R. Telomere maintenance and cancer—Look, no telomerase. Nat. Rev. Cancer 2002, 2, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Sobinoff, A.P.; Pickett, H.A. Alternative Lengthening of Telomeres: DNA Repair Pathways Converge. Trends Genet. 2017, 33, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Hoang, S.M.; O’Sullivan, R.J. Alternative Lengthening of Telomeres: Building Bridges To Connect Chromosome Ends. Trends Cancer 2020, 6, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Neumann, A.A.; Watson, C.M.; Noble, J.R.; Pickett, H.A.; Tam, P.P.; Reddel, R.R. Alternative lengthening of telomeres in normal mammalian somatic cells. Genes Dev. 2013, 27, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Siriaco, G.M.; Cenci, G.; Haoudi, A.; Champion, L.E.; Zhou, C.; Gatti, M.; Mason, J.M. Telomere elongation (Tel), a new mutation in Drosophila melanogaster that produces long telomeres. Genetics 2002, 160, 235–245. [Google Scholar] [CrossRef]
- Melnikova, L.; Georgiev, P. Enhancer of terminal gene conversion, a new mutation in Drosophila melanogaster that induces telomere elongation by gene conversion. Genetics 2002, 162, 1301–1312. [Google Scholar] [CrossRef]
- Singh, A.K.; Lakhotia, S.C. The hnRNP A1 homolog Hrb87F/Hrp36 is important for telomere maintenance in Drosophila melanogaster. Chromosoma 2016, 125, 373–388. [Google Scholar] [CrossRef]
- LaBranche, H.; Dupuis, S.; Ben-David, Y.; Bani, M.R.; Wellinger, R.J.; Chabot, B. Telomere elongation by hnRNP A1 and a derivative that interacts with telomeric repeats and telomerase. Nat. Genet. 1998, 19, 199–202. [Google Scholar] [CrossRef]
- Zhang, Q.S.; Manche, L.; Xu, R.M.; Krainer, A.R. hnRNP A1 associates with telomere ends and stimulates telomerase activity. RNA 2006, 12, 1116–1128. [Google Scholar] [CrossRef]
- Chen, B.; Chu, T.; Harms, E.; Gergen, J.P.; Strickland, S. Mapping of Drosophila mutations using site-specific male recombination. Genetics 1998, 149, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Mackay, T.F.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The Drosophila melanogaster Genetic Reference Panel. Nature 2012, 482, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Zhai, R.G.; Hiesinger, P.R.; Koh, T.W.; Verstreken, P.; Schulze, K.L.; Cao, Y.; Jafar-Nejad, H.; Norga, K.K.; Pan, H.; Bayat, V.; et al. Mapping Drosophila mutations with molecularly defined P element insertions. Proc. Natl. Acad. Sci. USA 2003, 100, 10860–10865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metaxakis, A.; Oehler, S.; Klinakis, A.; Savakis, C. Minos as a genetic and genomic tool in Drosophila melanogaster. Genetics 2005, 171, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Neron, B.; Menager, H.; Maufrais, C.; Joly, N.; Maupetit, J.; Letort, S.; Carrere, S.; Tuffery, P.; Letondal, C. Mobyle: A new full web bioinformatics framework. Bioinformatics 2009, 25, 3005–3011. [Google Scholar] [CrossRef] [Green Version]
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Torok, T.; Benitez, C.; Takacs, S.; Biessmann, H. The protein encoded by the gene proliferation disrupter (prod) is associated with the telomeric retrotransposon array in Drosophila melanogaster. Chromosoma 2007, 116, 185–195. [Google Scholar] [CrossRef]
- Wei, K.H.; Reddy, H.M.; Rathnam, C.; Lee, J.; Lin, D.; Ji, S.; Mason, J.M.; Clark, A.G.; Barbash, D.A. A Pooled Sequencing Approach Identifies a Candidate Meiotic Driver in Drosophila. Genetics 2017, 206, 451–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipressa, F.; Morciano, P.; Bosso, G.; Mannini, L.; Galati, A.; Raffa, G.D.; Cacchione, S.; Musio, A.; Cenci, G. A role for Separase in telomere protection. Nat. Commun. 2016, 7, 10405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawitz, P.; Rodelsperger, C.; Jager, M.; Jostins, L.; Bauer, S.; Robinson, P.N. Microindel detection in short-read sequence data. Bioinformatics 2010, 26, 722–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rawe, J.; Jiang, T.; Sun, G.; Wu, Y.; Wang, W.; Hu, J.; Bodily, P.; Tian, L.; Hakonarson, H.; Johnson, W.E.; et al. Low concordance of multiple variant-calling pipelines: Practical implications for exome and genome sequencing. Genome Med. 2013, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cenci, G.; Siriaco, G.; Raffa, G.D.; Kellum, R.; Gatti, M. The Drosophila HOAP protein is required for telomere capping. Nat. Cell Biol. 2003, 5, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Raffa, G.D.; Cenci, G.; Ciapponi, L.; Gatti, M. Organization and maintenance of Drosophila telomeres: The roles of terminin and non-terminin proteins. Tsitologiia 2013, 55, 204–208. [Google Scholar] [PubMed]
- Perrini, B.; Piacentini, L.; Fanti, L.; Altieri, F.; Chichiarelli, S.; Berloco, M.; Turano, C.; Ferraro, A.; Pimpinelli, S. HP1 controls telomere capping, telomere elongation, and telomere silencing by two different mechanisms in Drosophila. Mol. Cell 2004, 15, 467–476. [Google Scholar] [CrossRef]
- Savitsky, M.; Kravchuk, O.; Melnikova, L.; Georgiev, P. Heterochromatin protein 1 is involved in control of telomere elongation in Drosophila melanogaster. Mol. Cell Biol. 2002, 22, 3204–3218. [Google Scholar] [CrossRef] [Green Version]
- Proskuryakov, K.A.; Melnikova, L.S. Functional separation of genetic factors telomere elongation (Tel) and enhancer of terminal gene conversion (E(tc)) involved in telomere elongation in Drosophila melanogaster. Dokl. Biochem. Biophys. 2008, 421, 199–203. [Google Scholar] [CrossRef]
- Pineyro, D.; Lopez-Panades, E.; Lucena-Perez, M.; Casacuberta, E. Transcriptional analysis of the HeT-A retrotransposon in mutant and wild type stocks reveals high sequence variability at Drosophila telomeres and other unusual features. BMC Genom. 2011, 12, 573. [Google Scholar] [CrossRef]
- Walter, M.F.; Biessmann, M.R.; Benitez, C.; Torok, T.; Mason, J.M.; Biessmann, H. Effects of telomere length in Drosophila melanogaster on life span, fecundity, and fertility. Chromosoma 2007, 116, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Nabetani, A.; Ishikawa, F. Alternative lengthening of telomeres pathway: Recombination-mediated telomere maintenance mechanism in human cells. J. Biochem. 2011, 149, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryder, E.; Russell, S. Transposable elements as tools for genomics and genetics in Drosophila. Brief. Funct. Genom. Proteom. 2003, 2, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Spradling, A.C.; Stern, D.M.; Kiss, I.; Roote, J.; Laverty, T.; Rubin, G.M. Gene disruptions using P transposable elements: An integral component of the Drosophila genome project. Proc. Natl. Acad. Sci. USA 1995, 92, 10824–10830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellen, H.J.; Levis, R.W.; Liao, G.; He, Y.; Carlson, J.W.; Tsang, G.; Evans-Holm, M.; Hiesinger, P.R.; Schulze, K.L.; Rubin, G.M.; et al. The BDGP gene disruption project: Single transposon insertions associated with 40% of Drosophila genes. Genetics 2004, 167, 761–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, G.; Savakis, C. Minos, a new transposable element from Drosophila hydei, is a member of the Tc1-like family of transposons. Nucleic Acids Res. 1991, 19, 6646. [Google Scholar] [CrossRef]
- Loukeris, T.G.; Arca, B.; Livadaras, I.; Dialektaki, G.; Savakis, C. Introduction of the transposable element Minos into the germ line of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1995, 92, 9485–9489. [Google Scholar] [CrossRef] [Green Version]
- Venken, K.J.; Schulze, K.L.; Haelterman, N.A.; Pan, H.; He, Y.; Evans-Holm, M.; Carlson, J.W.; Levis, R.W.; Spradling, A.C.; Hoskins, R.A.; et al. MiMIC: A highly versatile transposon insertion resource for engineering Drosophila melanogaster genes. Nat. Methods 2011, 8, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, R.; Prakash, T.; Dash, D.; Brahmachari, V. In silico characterization of the INO80 subfamily of SWI2/SNF2 chromatin remodeling proteins. Biochem. Biophys. Res. Commun. 2004, 320, 197–204. [Google Scholar] [CrossRef]
- Neuman, S.D.; Ihry, R.J.; Gruetzmacher, K.M.; Bashirullah, A. INO80-dependent regression of ecdysone-induced transcriptional responses regulates developmental timing in Drosophila. Dev. Biol. 2014, 387, 229–239. [Google Scholar] [CrossRef]
- Bhatia, S.; Pawar, H.; Dasari, V.; Mishra, R.K.; Chandrashekaran, S.; Brahmachari, V. Chromatin remodeling protein INO80 has a role in regulation of homeotic gene expression in Drosophila. Genes Cells 2010, 15, 725–735. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Localization of Tel1 by site-specific recombination. (A) The upper chromosome map shows the candidate genes between two P element insertion sites, 05151 and 05113 (vertical green lines). This region was identified as containing Tel1 based on Table 1, round 1. Positions of transposons used for further mapping are indicated by green arrows. The Tel1 mutation is boxed in red. The lower chromosome map shows expansion of the 92A3 region. (B) Graphs showing the change in relative HeT-A copy number (telomere length) in recombinants of Tel1/MI02316, Tel1/MI03112, and Tel1/d10097 over 12 generations. The st recombinants are shown as red lines; ca recombinants as purple lines. These data delimit Tel1 to a 15 kb between inserts MI02316 and d10097 (shown as red rectangles in (A)).
Figure 1.
Localization of Tel1 by site-specific recombination. (A) The upper chromosome map shows the candidate genes between two P element insertion sites, 05151 and 05113 (vertical green lines). This region was identified as containing Tel1 based on Table 1, round 1. Positions of transposons used for further mapping are indicated by green arrows. The Tel1 mutation is boxed in red. The lower chromosome map shows expansion of the 92A3 region. (B) Graphs showing the change in relative HeT-A copy number (telomere length) in recombinants of Tel1/MI02316, Tel1/MI03112, and Tel1/d10097 over 12 generations. The st recombinants are shown as red lines; ca recombinants as purple lines. These data delimit Tel1 to a 15 kb between inserts MI02316 and d10097 (shown as red rectangles in (A)).

Figure 2.
Telomere length in transposon insertion stocks. Q-PCR analysis of HeT-A copy number in different transposon insertion stocks used for mapping Tel1 mutation. Error bars represent standard deviation measured from the triplicate Q-PCR results. MB09416 was not used for subsequent site-specific recombination mapping.
Figure 2.
Telomere length in transposon insertion stocks. Q-PCR analysis of HeT-A copy number in different transposon insertion stocks used for mapping Tel1 mutation. Error bars represent standard deviation measured from the triplicate Q-PCR results. MB09416 was not used for subsequent site-specific recombination mapping.

Figure 3.
Dosage effects of Tel on HeT-A copy number. (A) The cytogenetic and physical map of genomic region 91C-91D is displayed, highlighting the 15 kb Tel1 region as a vertical box. The lower part of the panel shows the extent of chromosome deficiencies. The bottom rectangle, Dp(3:3)cam30T, is a duplication for this region. Dotted lines beyond this rectangle show that the duplication extends beyond the represented region. (B) Shows the relative HeT-A copy number in stocks of the aberrations shown in A. The highlighted box represents a duplication that includes mapped region (Dp(3;3)cam30T covers 90C-93C) and another duplication of a neighboring region of the genome (Dp(3;3)cam35 covers 67C5-69A5) as a control. The mean from three replicates is represented here and error bars represent standard deviation. ** p < 0.005; *** p < 0.0001, one-way ANOVA with Sidak correction.
Figure 3.
Dosage effects of Tel on HeT-A copy number. (A) The cytogenetic and physical map of genomic region 91C-91D is displayed, highlighting the 15 kb Tel1 region as a vertical box. The lower part of the panel shows the extent of chromosome deficiencies. The bottom rectangle, Dp(3:3)cam30T, is a duplication for this region. Dotted lines beyond this rectangle show that the duplication extends beyond the represented region. (B) Shows the relative HeT-A copy number in stocks of the aberrations shown in A. The highlighted box represents a duplication that includes mapped region (Dp(3;3)cam30T covers 90C-93C) and another duplication of a neighboring region of the genome (Dp(3;3)cam35 covers 67C5-69A5) as a control. The mean from three replicates is represented here and error bars represent standard deviation. ** p < 0.005; *** p < 0.0001, one-way ANOVA with Sidak correction.

Figure 4.
Telomere lengths in DGRP lines. A bar graph shows the log-normal distribution of telomere lengths among the 162 DGRP lines measured. The blue arrow shows the position of the Oregon-R control, and the red arrow shows the position of Tel1. Three lines have Het-A copy numbers that exceed three standard deviations from the mean. These are RAL-161, -703, and -882. The red curve indicates the expected distribution.
Figure 4.
Telomere lengths in DGRP lines. A bar graph shows the log-normal distribution of telomere lengths among the 162 DGRP lines measured. The blue arrow shows the position of the Oregon-R control, and the red arrow shows the position of Tel1. Three lines have Het-A copy numbers that exceed three standard deviations from the mean. These are RAL-161, -703, and -882. The red curve indicates the expected distribution.

Figure 5.
Genetic activity and conservation of the 15 kb Tel1 region. (A) The genes found in this region are aligned with molecular coordinates. Minos insertions used for mapping are shown in cyan triangles. Minos insertion MB09416, which showed telomere elongation, is highlighted in the red square. (B) The University of California Santa Cruz genome browser map highlights sequence conservation in this region among different insect species. (C) A developmental transcriptome analysis for the same region as determined by the modENCODE project is also shown. The red vertical line spanning all three panels indicates the position of the TGT deletion.
Figure 5.
Genetic activity and conservation of the 15 kb Tel1 region. (A) The genes found in this region are aligned with molecular coordinates. Minos insertions used for mapping are shown in cyan triangles. Minos insertion MB09416, which showed telomere elongation, is highlighted in the red square. (B) The University of California Santa Cruz genome browser map highlights sequence conservation in this region among different insect species. (C) A developmental transcriptome analysis for the same region as determined by the modENCODE project is also shown. The red vertical line spanning all three panels indicates the position of the TGT deletion.

Figure 6.
Transcript analysis in the genes near Tel1. The histogram represents the relative transcript levels from Tel1 mutant (red) and Oregon-R (blue) adult ovaries. The genes analyzed are found near Tel1 (TGT mutation), which include Ino80 and the genes within its introns. The last bar represents the expression of Ino80 gene spanning exons 8–9, around the position of the TGT deletion in intron 8. The expression levels of CG31244 and CG31245 are very low, similar to the modENCODE data.
Figure 6.
Transcript analysis in the genes near Tel1. The histogram represents the relative transcript levels from Tel1 mutant (red) and Oregon-R (blue) adult ovaries. The genes analyzed are found near Tel1 (TGT mutation), which include Ino80 and the genes within its introns. The last bar represents the expression of Ino80 gene spanning exons 8–9, around the position of the TGT deletion in intron 8. The expression levels of CG31244 and CG31245 are very low, similar to the modENCODE data.

Figure 7.
Telomere elongation in the CRISPR/Cas 9-induced Tel deletion alleles. (A) anti-HOAP immunostaining on 2L and XL chromosome tips from Tel∆F2/+ and Tel∆C11/+ hemyzygotes, respectively, showing protruding telomeric DNA only on the Tel mutant chromosomes (arrows). Note that, as expected, Tel mutant elongated telomeres do not impair HOAP localization. (B) Examples of telomere fusions involving either two or four chromosome tips (arrowheads) from different Tel∆ hemizygotes. (C) qPCR analysis on third instar larval DNA from two representative Tel∆ deletion alleles showing a robust increase in Het-A copy number compared to control (OR-R). Note that Tel∆B5 bears a deletion that uncovers TGT, while Tel∆C10 does not include TGT. * (p < 0.05).
Figure 7.
Telomere elongation in the CRISPR/Cas 9-induced Tel deletion alleles. (A) anti-HOAP immunostaining on 2L and XL chromosome tips from Tel∆F2/+ and Tel∆C11/+ hemyzygotes, respectively, showing protruding telomeric DNA only on the Tel mutant chromosomes (arrows). Note that, as expected, Tel mutant elongated telomeres do not impair HOAP localization. (B) Examples of telomere fusions involving either two or four chromosome tips (arrowheads) from different Tel∆ hemizygotes. (C) qPCR analysis on third instar larval DNA from two representative Tel∆ deletion alleles showing a robust increase in Het-A copy number compared to control (OR-R). Note that Tel∆B5 bears a deletion that uncovers TGT, while Tel∆C10 does not include TGT. * (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Recombinants obtained from each transposon used for site-specific recombination.
| Round a | Transposon Insertion | Cytology b | Coordinate c | No. Recombinants | Tel1 Position d |
|---|---|---|---|---|---|
| 1 | P{PZ}sqz02102 | 91F4 | 19,165,876 | 3 | Right |
| 1 | P{lacW}vibj5A6 | 91F12 | 19,226,365 | 6 | Right |
| 1 | P{PZ}l(3)0582005820 | 91F12 | 19,229,496 | 3 | Right |
| 1 | P{PZ}Dl05151 | 92A2 | 19,326,218 | 2 | Right |
| 3 | Mi{MIC}Ino80MI03112 | 92A3 | 19,373,898 | 1 | Right |
| 3 | Mi{MIC}Ino80MI02316 | 92A3 | 19,388,379 | 1 | Right |
| 2 | P{XP}Ino80d10097 | 92A3 | 19,403,413 | 6 | Left |
| 2 | Mi{ET1}CG31221MB02141 | 92A3 | 19,414,147 | 5 | Left |
| 2 | P{EPgy2}CG31221EY10678 | 92A5 | 19,453,650 | 4 | Left |
| 2 | P{XP}Dysd03320 | 92A6 | 19,498,929 | 3 | Left |
| 2 | MI{ET1}CG6231MB01639 | 92A11 | 19,629,652 | 6 | Left |
| 1 | P{SUPor-P}CG16718KG06218 | 92A11 | 19,641,774 | 3 | Left |
| 1 | P{PZ}Vha1305113 | 92A11 | 19,644,018–196,44,026 | 3 | Left |
| 1 | P{hsneo}l(3)neo501 | 92B3 | 19,836,871 | 3 | Left |
a Transposons were used to induce recombination as they became available. Succeeding rounds used slightly different procedures, as described in the text. b Estimated cytological band as reported in FlyBase. c Nucleotide position in the genomic sequence of chromosome arm 3R. d The position of Tel1 relative to the insertion site.
Table 2.
SNPs and indels found in the Tel1 genome relative to the standard reference sequence of chromosome arm 3R between coordinates 19,325,278 and 19,404,278.
Table 2.
SNPs and indels found in the Tel1 genome relative to the standard reference sequence of chromosome arm 3R between coordinates 19,325,278 and 19,404,278.
| Polymorphisms | SNPs | Indels |
|---|---|---|
| Identified by CLC-Genomics | 626 | 88 |
| Identified by manual comparison | - | 13 |
| Identified by de novo assembly | - | 14 |
| Total | 626 | 115 |
| Not in DGRP a | 0 | 2 |
a There were 159 DGRP lines used in the SNP comparison (Freeze1 data) and eight used in the indel comparison.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Reddy, H.M.; Randall, T.A.; Cipressa, F.; Porrazzo, A.; Cenci, G.; Frydrychova, R.C.; Mason, J.M. Identification of the Telomere elongation Mutation in Drosophila. Cells 2022, 11, 3484. https://doi.org/10.3390/cells11213484
AMA Style
Reddy HM, Randall TA, Cipressa F, Porrazzo A, Cenci G, Frydrychova RC, Mason JM. Identification of the Telomere elongation Mutation in Drosophila. Cells. 2022; 11(21):3484. https://doi.org/10.3390/cells11213484
Chicago/Turabian StyleReddy, Hemakumar M., Thomas A. Randall, Francesca Cipressa, Antonella Porrazzo, Giovanni Cenci, Radmila Capkova Frydrychova, and James M. Mason. 2022. "Identification of the Telomere elongation Mutation in Drosophila" Cells 11, no. 21: 3484. https://doi.org/10.3390/cells11213484
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.