

The Potential Role of Cytotoxic Immune Effectors in Induction, Progression and Pathogenesis of Amyotrophic Lateral Sclerosis (ALS)
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. ALS Patients and Healthy Individuals’ Information
2.1.1. Cell Lines, Reagents, and Antibodies
2.1.2. Isolation of Human PBMCs, NK Cells, T Cells, and Monocytes
2.1.3. Enzyme-Linked Immunosorbent Assays (ELISAs), Enzyme-Linked Immunospot (ELISpot), and Multiplex Cytokine Arrays
2.1.4. Surface Staining
2.1.5. 51Cr Release Cytotoxicity Assay
2.1.6. NAC Preparation and Infusion
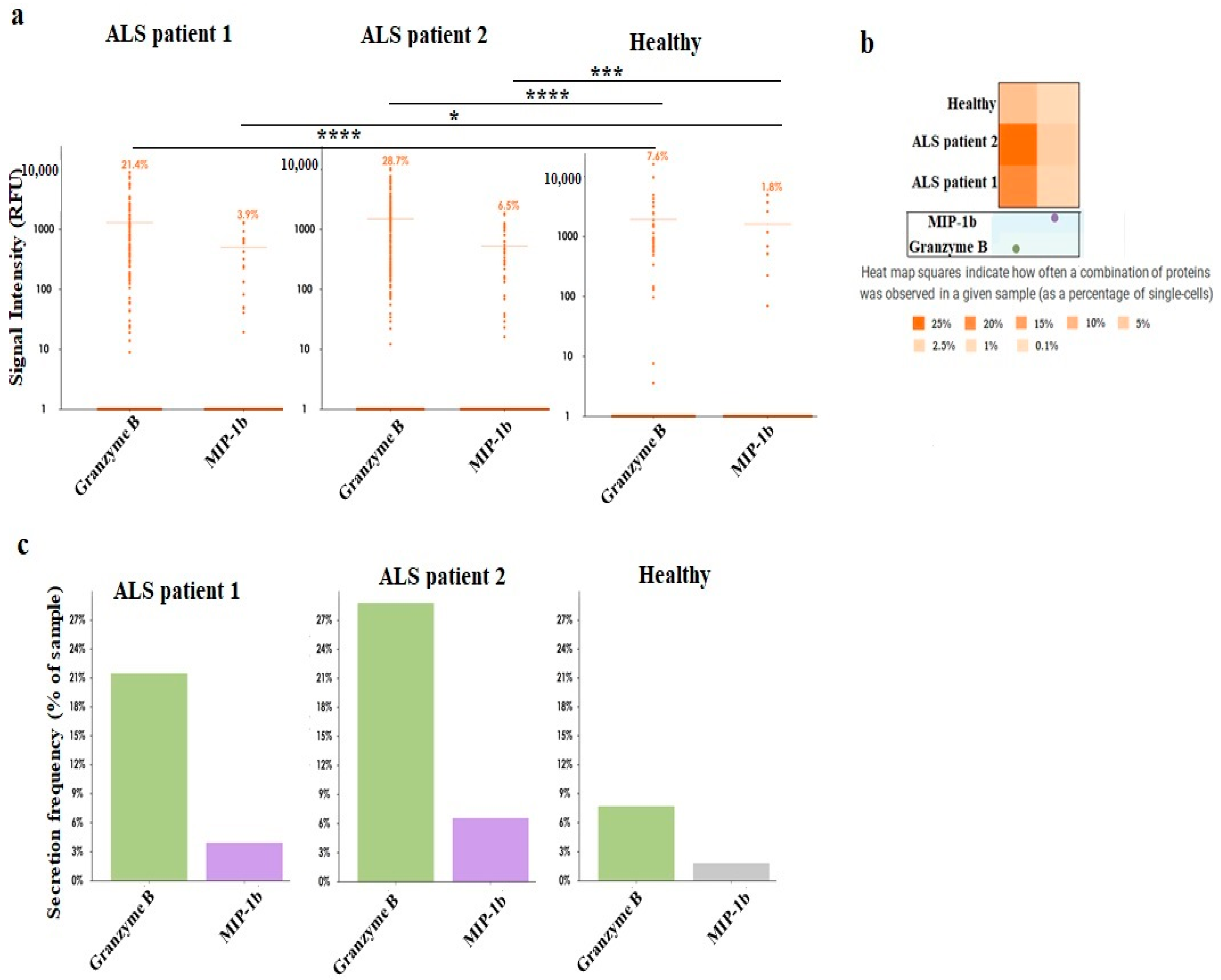
2.1.7. Single-Cell Protein Analysis and Polyfunctionality of NK and CD8+ T Cells
2.1.8. Sonication of Probiotic Bacteria AJ2
2.1.9. Generation of Osteoclasts and Osteoclasts-Induced NK Cell Expansion
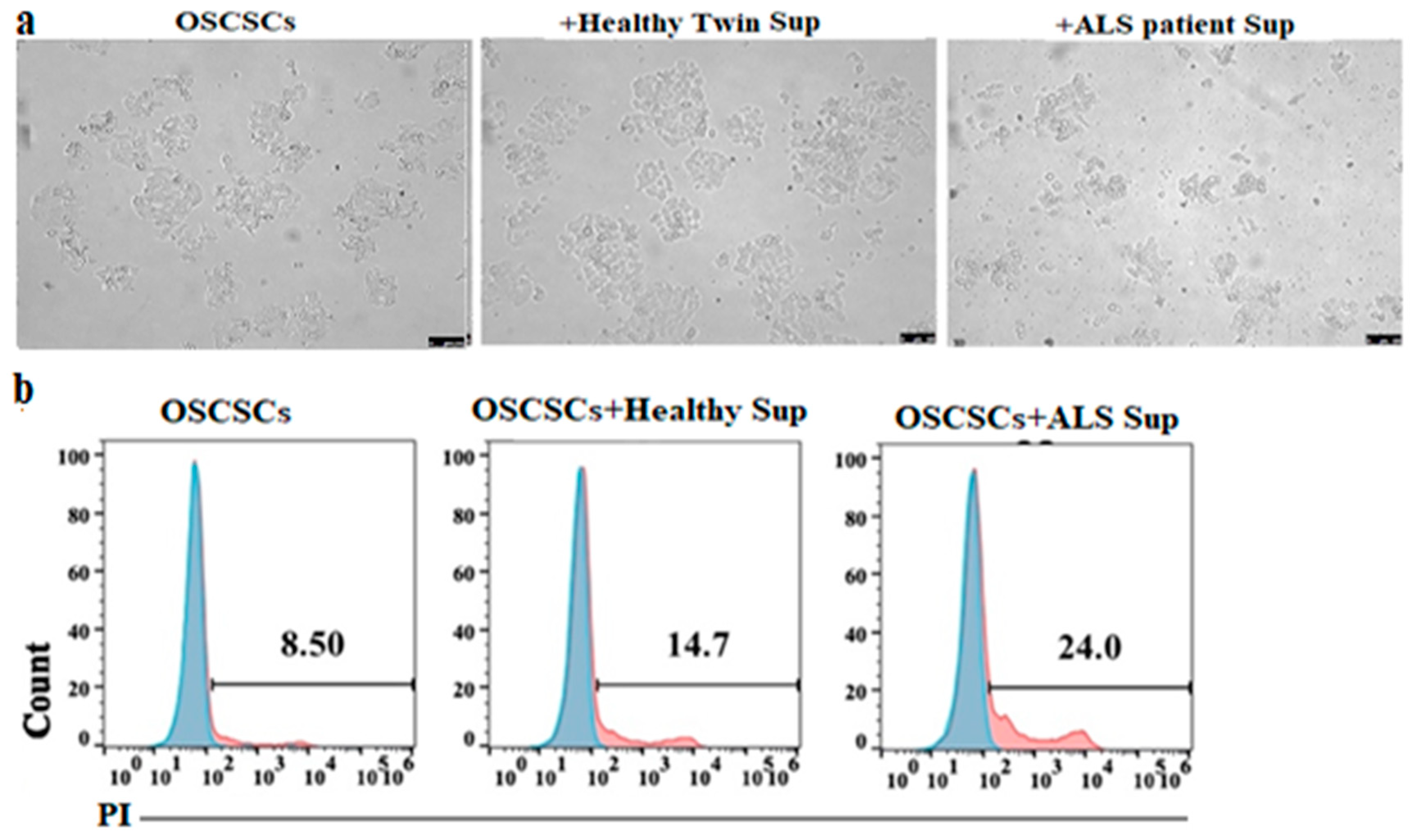
2.1.10. Oral Squamous Carcinoma Stem Cells (OSCSCs) Treatment with CD8+ T Cells Supernatant
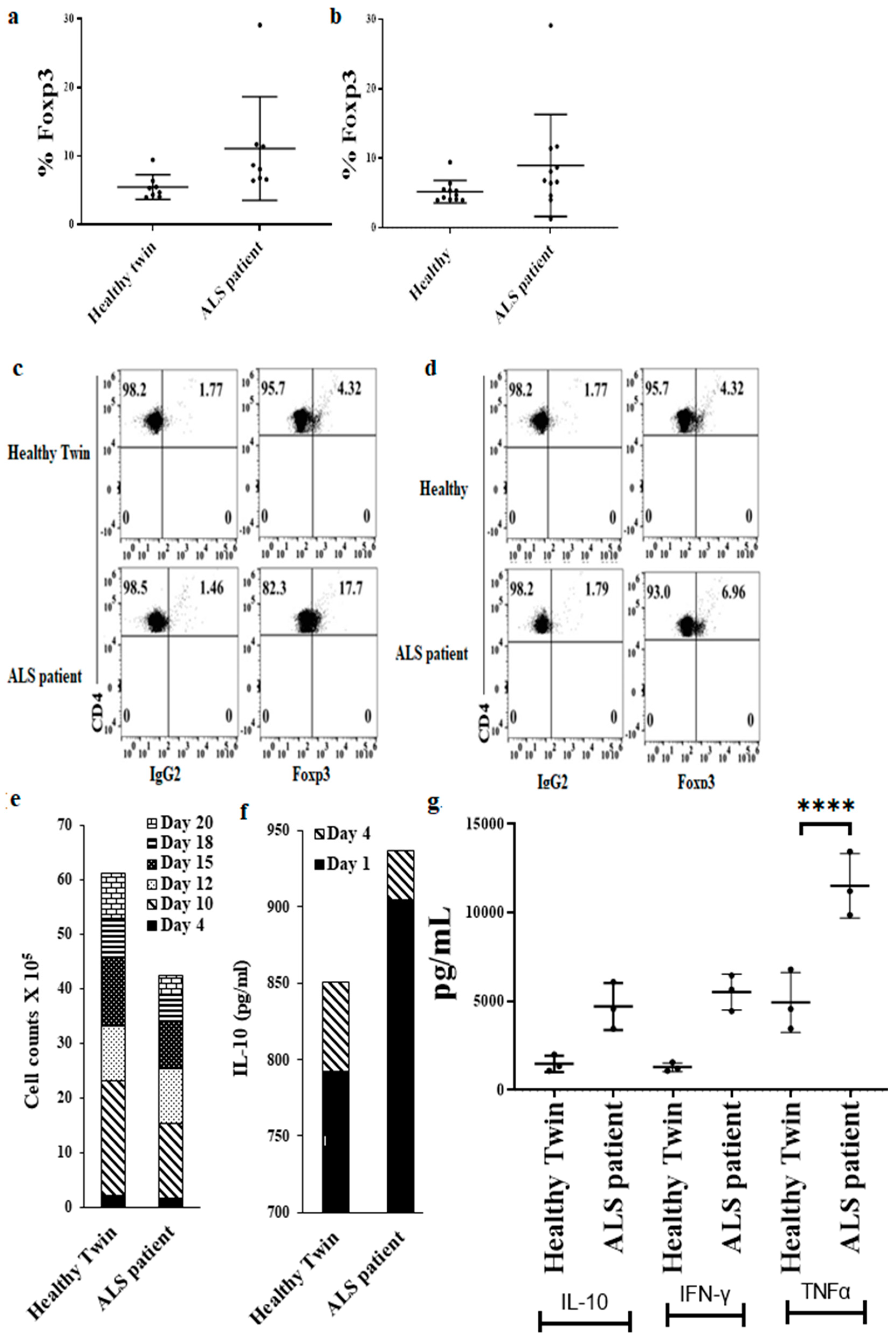
2.1.11. CD4+ T Cells Differentiation to Treg Cells
2.1.12. Statistical Analysis
3. Results
3.1. Genetic Mutational Differences between ALS Patient and Healthy Twin
3.2. Increased NK and B Cell Percentages in the Peripheral Blood of ALS Patients in Comparison to Healthy Individuals
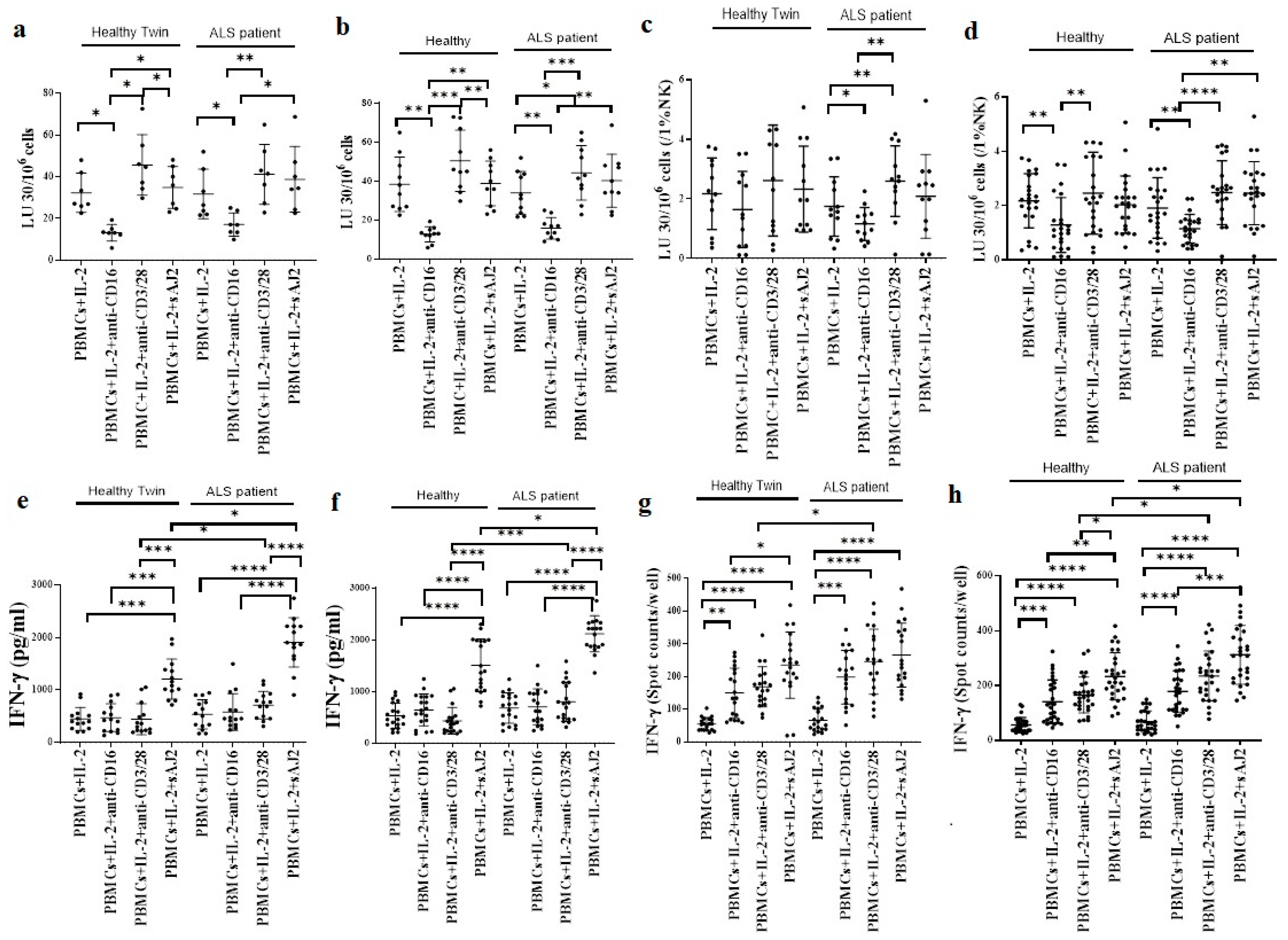
3.3. Similar Levels of Cytotoxicity but Significantly Increased IFN-γ Secretion in ALS Patients’ PBMCs when compared to Healthy Individuals
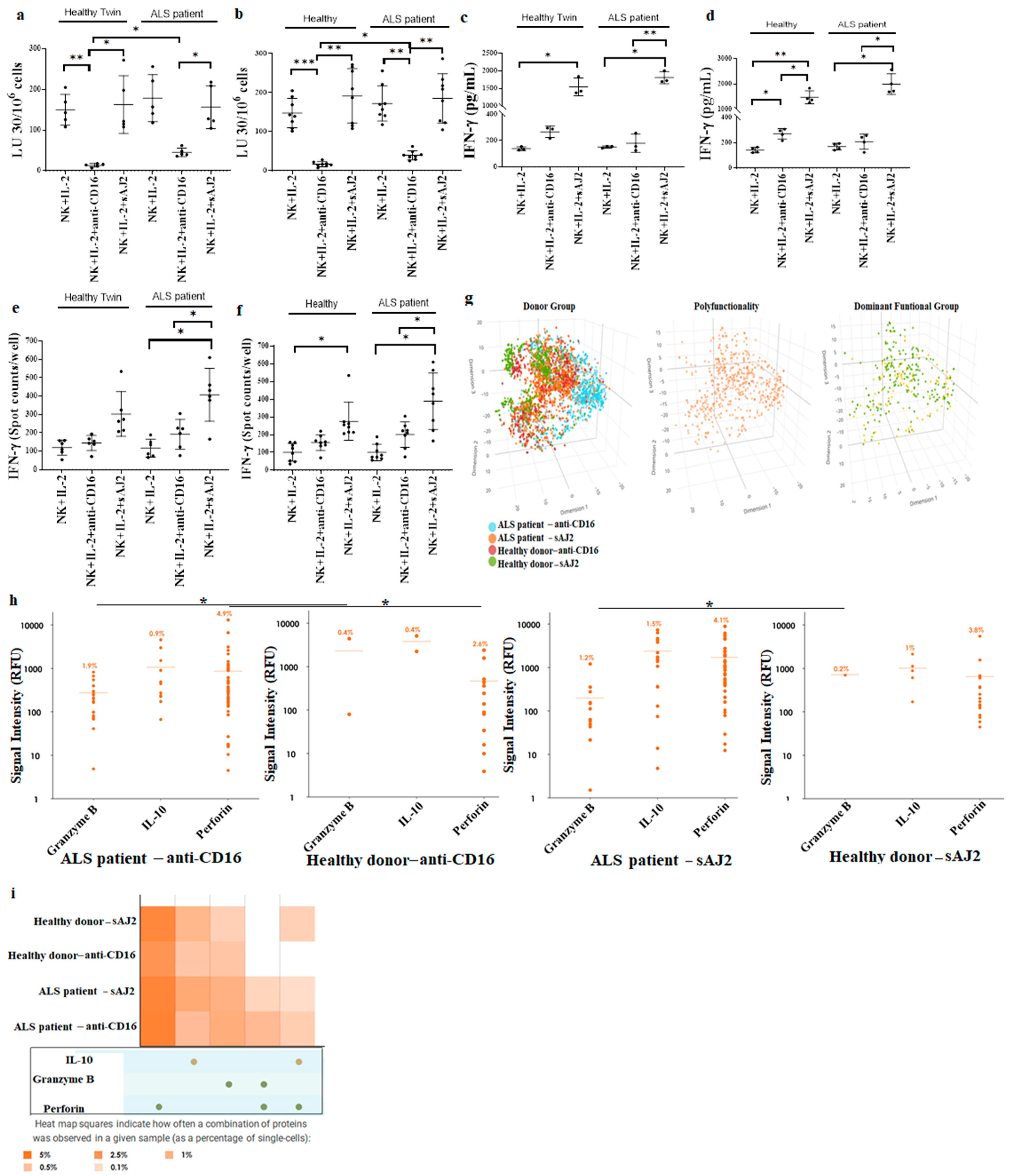
3.4. Increased Cell-Mediated Cytotoxicity Was Seen in ALS Patients’ NK Cells in Comparison to Healthy Individuals’ NK Cells
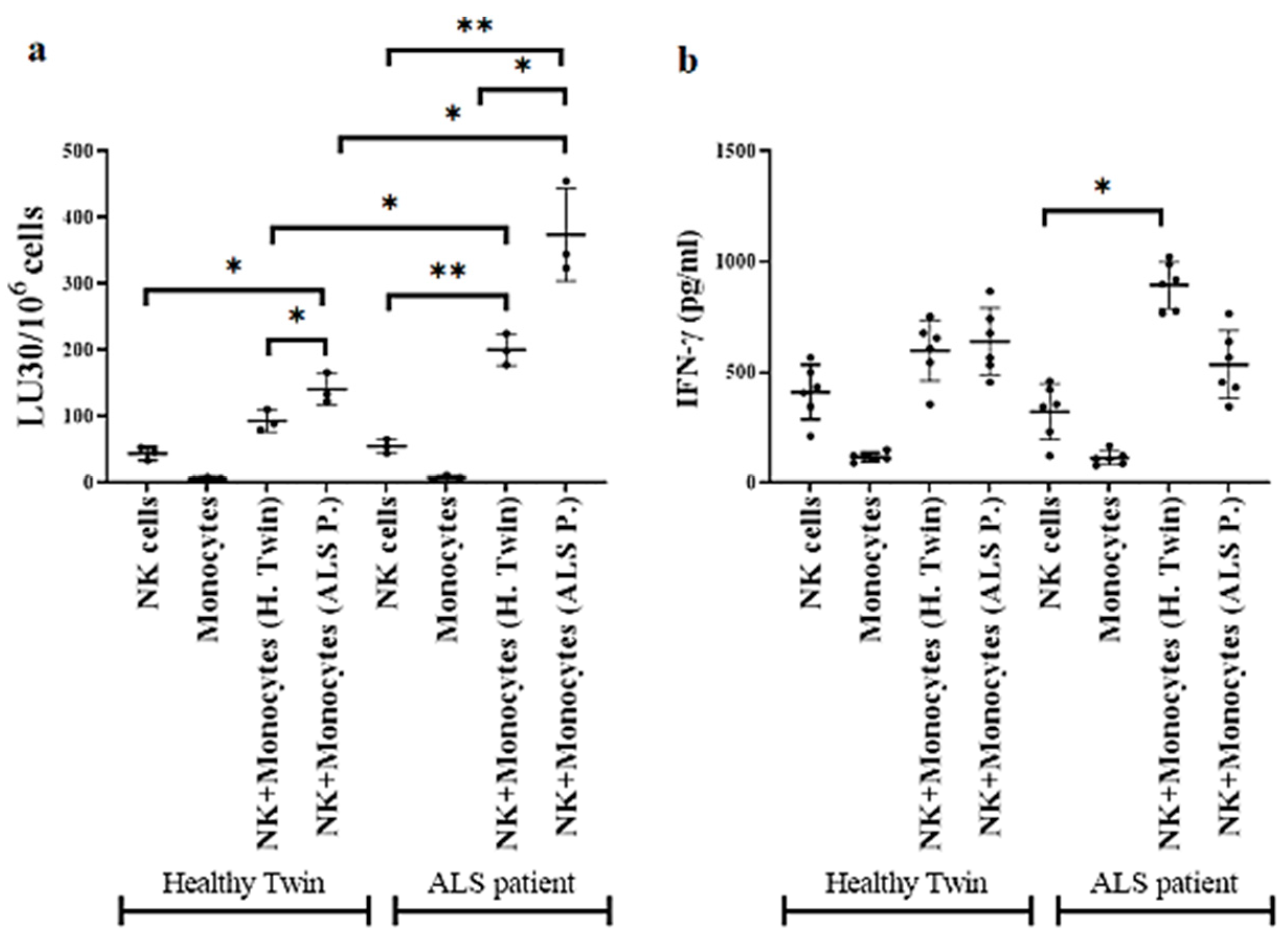
3.5. Monocytes Induced Increased Cell-Mediated Cytotoxicity and Secretion of IFN-γ by NK Cells
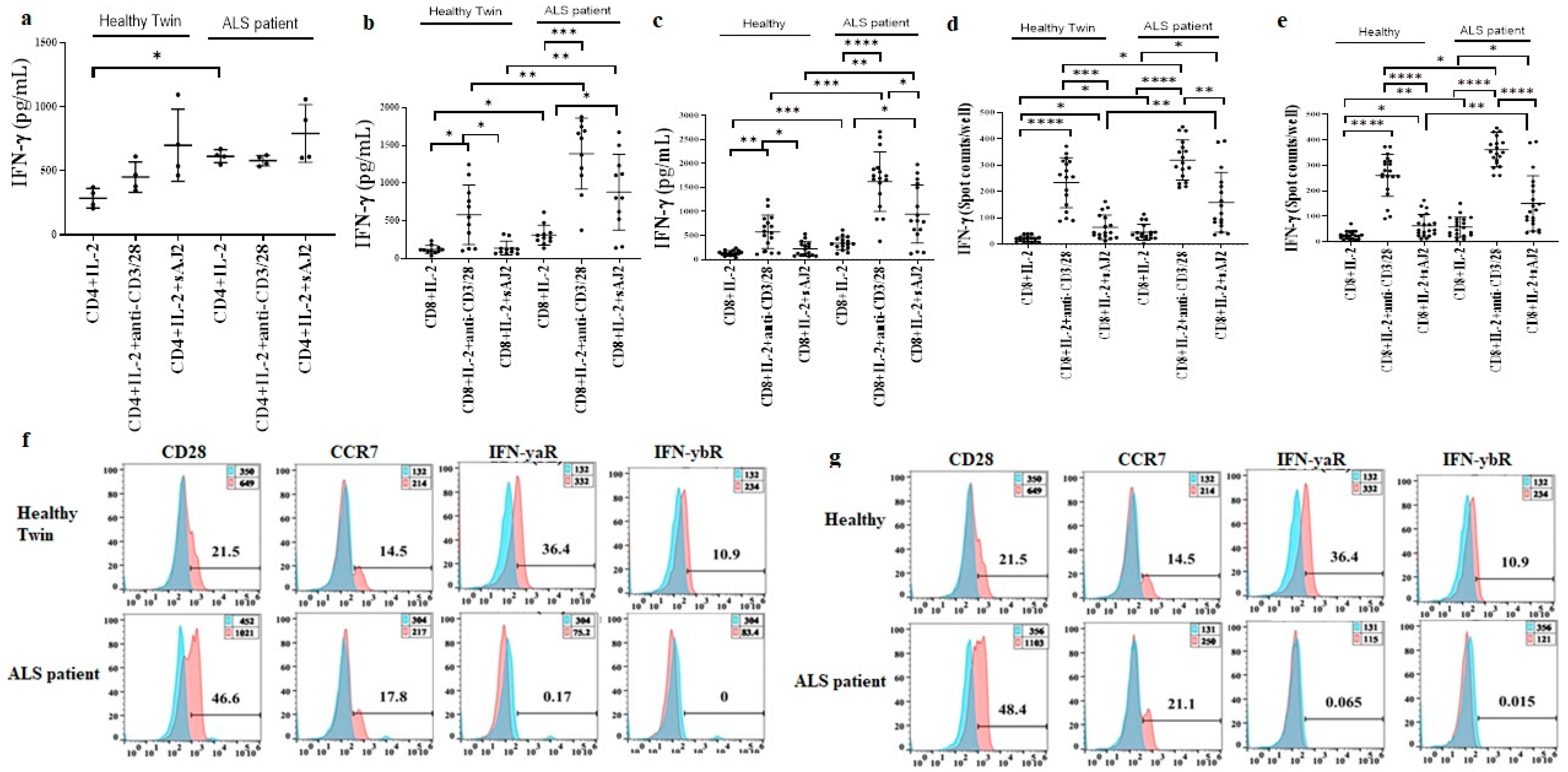
3.6. Increased Effector Memory and Secretion of IFN-γ by ALS Patients’ CD8+ T Cells in Comparison to Healthy Individuals’ CD8+ T Cells
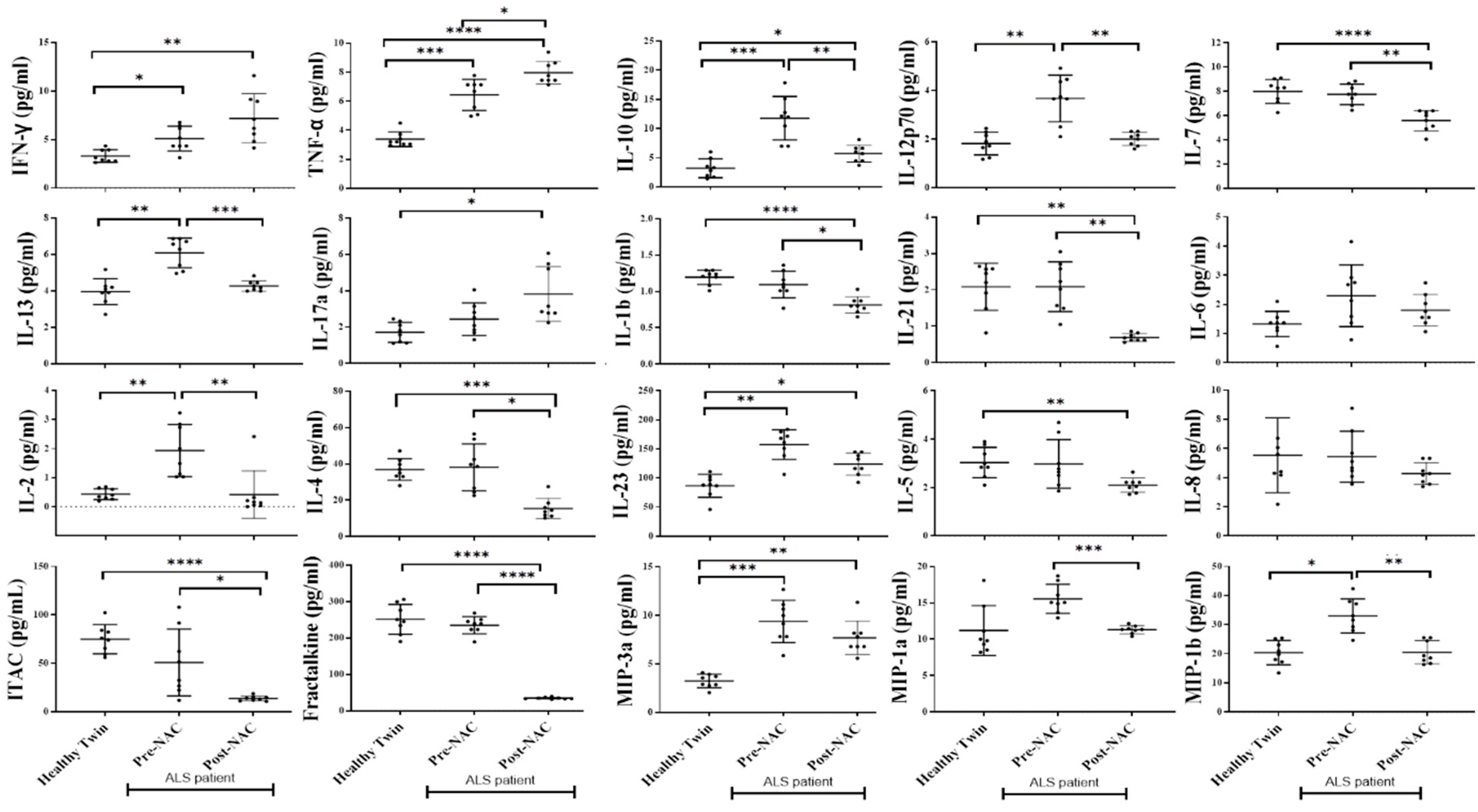
3.7. Increased Inflammatory Cytokines in the Serum of ALS Patients in Comparison to Healthy Individuals
3.8. Supernatants Obtained from ALS Patients’ CD8+ T Cells Induced Higher Cell Death and Differentiation of Epithlial Tumor
3.9. Increased Regulatory CD4+ T Cell Subsets in ALS Patients’ PBMCs in Comparison to Healthy Individuals’ PBMCs
3.10. Weekly NAC Injections in ALS Patients Decreased Inflammatory Cytokines in Peripheral Blood except for IFN-γ, TNF-α, IL-17a, and GMCSF
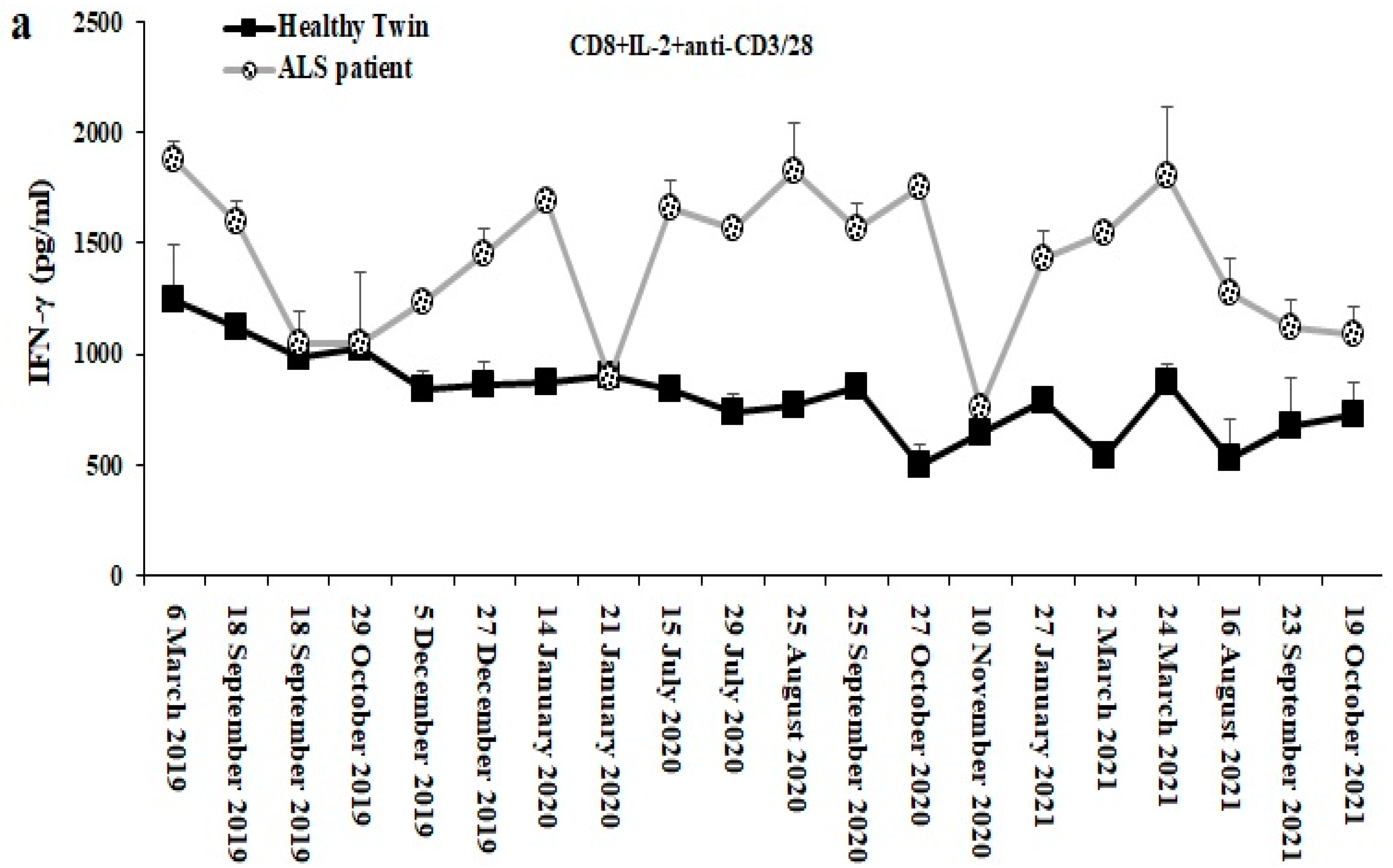
3.11. Longitudinal Analysis of CD8+ T Cell Mediated IFN g Secretion from ALS Patient as Compared to Those of the Healthy Identical Twin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; Corr, E.M.; Logroscino, G.; Robberecht, W.; Shaw, P.J.; Simmons, Z.; van den Berg, L.H. Amyotrophic lateral sclerosis. Nat. Rev. Dis. Primers 2017, 3, 17085. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.H.; Al-Chalabi, A. Amyotrophic Lateral Sclerosis. N. Engl. J. Med. 2017, 377, 162–172. [Google Scholar] [CrossRef] [Green Version]
- van Es, M.A.; Hardiman, O.; Chio, A.; Al-Chalabi, A.; Pasterkamp, R.J.; Veldink, J.H.; van den Berg, L.H. Amyotrophic lateral sclerosis. Lancet 2017, 390, 2084–2098. [Google Scholar] [CrossRef]
- Masrori, P.; Van Damme, P. Amyotrophic lateral sclerosis: A clinical review. Eur. J. Neurol. 2020, 27, 1918–1929. [Google Scholar] [CrossRef]
- Fecto, F.; Yan, J.; Vemula, S.P.; Liu, E.; Yang, Y.; Chen, W.; Zheng, J.G.; Shi, Y.; Siddique, N.; Arrat, H.; et al. SQSTM1 mutations in familial and sporadic amyotrophic lateral sclerosis. Arch. Neurol. 2011, 68, 1440–1446. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Bharathi, V.; Sivalingam, V.; Girdhar, A.; Patel, B.K. Molecular Mechanisms of TDP-43 Misfolding and Pathology in Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2019, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Suk, T.R.; Rousseaux, M.W.C. The role of TDP-43 mislocalization in amyotrophic lateral sclerosis. Mol. Neurodegener. 2020, 15, 45. [Google Scholar] [CrossRef] [PubMed]
- Murdock, B.J.; Bender, D.E.; Segal, B.M.; Feldman, E.L. The dual roles of immunity in ALS: Injury overrides protection. Neurobiol. Dis. 2015, 77, 1–12. [Google Scholar] [CrossRef]
- Zhao, W.; Beers, D.R.; Appel, S.H. Immune-mediated mechanisms in the pathoprogression of amyotrophic lateral sclerosis. J. Neuroimmune Pharmacol. 2013, 8, 888–899. [Google Scholar] [CrossRef] [Green Version]
- Beers, D.R.; Henkel, J.S.; Zhao, W.; Wang, J.; Appel, S.H. CD4+ T cells support glial neuroprotection, slow disease progression, and modify glial morphology in an animal model of inherited ALS. Proc. Natl. Acad. Sci. USA 2008, 105, 15558–15563. [Google Scholar] [CrossRef]
- Butovsky, O.; Siddiqui, S.; Gabriely, G.; Lanser, A.J.; Dake, B.; Murugaiyan, G.; Doykan, C.E.; Wu, P.M.; Gali, R.R.; Iyer, L.K.; et al. Modulating inflammatory monocytes with a unique microRNA gene signature ameliorates murine ALS. J. Clin. Investig. 2012, 122, 3063–3087. [Google Scholar] [CrossRef] [Green Version]
- Coque, E.; Salsac, C.; Espinosa-Carrasco, G.; Varga, B.; Degauque, N.; Cadoux, M.; Crabé, R.; Virenque, A.; Soulard, C.; Fierle, J.K.; et al. Cytotoxic CD8(+) T lymphocytes expressing ALS-causing SOD1 mutant selectively trigger death of spinal motoneurons. Proc. Natl. Acad. Sci. USA 2019, 116, 2312–2317. [Google Scholar] [CrossRef] [Green Version]
- Murdock, B.J.; Zhou, T.; Kashlan, S.R.; Little, R.J.; Goutman, S.A.; Feldman, E.L. Correlation of Peripheral Immunity with Rapid Amyotrophic Lateral Sclerosis Progression. JAMA Neurol. 2017, 74, 1446–1454. [Google Scholar] [CrossRef]
- Murdock, B.J.; Famie, J.P.; Piecuch, C.E.; Raue, K.D.; Mendelson, F.E.; Pieroni, C.H.; Iniguez, S.D.; Zhao, L.; Goutman, S.A.; Feldman, E.L. NK cells associate with ALS in a sex- and age-dependent manner. JCI Insight 2021, 6, e147129. [Google Scholar] [CrossRef]
- Joyce, P.I.; McGoldrick, P.; Saccon, R.A.; Weber, W.; Fratta, P.; West, S.J.; Zhu, N.; Carter, S.; Phatak, V.; Stewart, M.; et al. A novel SOD1-ALS mutation separates central and peripheral effects of mutant SOD1 toxicity. Hum. Mol. Genet. 2015, 24, 1883–1897. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.; Gautier, O.; Tassoni-Tsuchida, E.; Ma, X.R.; Gitler, A.D. ALS Genetics: Gains, Losses, and Implications for Future Therapies. Neuron 2020, 108, 822–842. [Google Scholar] [CrossRef]
- Schweikl, H.; Hartmann, A.; Hiller, K.A.; Spagnuolo, G.; Bolay, C.; Brockhoff, G.; Schmalz, G. Inhibition of TEGDMA and HEMA-induced genotoxicity and cell cycle arrest by N-acetylcysteine. Dent. Mater. Off. Publ. Acad. Dent. Mater. 2007, 23, 688–695. [Google Scholar] [CrossRef]
- Mokhtari, V.; Afsharian, P.; Shahhoseini, M.; Kalantar, S.M.; Moini, A. A Review on Various Uses of N-Acetyl Cysteine. Cell J. 2017, 19, 11–17. [Google Scholar]
- Parasassi, T.; Brunelli, R.; Krasnowska, E.K.; Lundeberg, T.; Pittaluga, E.; Romano, M.C. Into the redox control: N-acetyl-cysteine pleiotropic effects from the laboratory to clinical applications. Acupunct. Relat. Ther. 2014, 2, 2–13. [Google Scholar] [CrossRef]
- Beg, A.A.; Baltimore, D. An essential role for NF-kappaB in preventing TNF-alpha-induced cell death. Science 1996, 274, 782–784. [Google Scholar] [CrossRef]
- Van Antwerp, D.J.; Martin, S.J.; Kafri, T.; Green, D.R.; Verma, I.M. Suppression of TNF-alpha-induced apoptosis by NF-kappaB. Science 1996, 274, 787–789. [Google Scholar] [CrossRef]
- Tseng, H.C.; Arasteh, A.; Paranjpe, A.; Teruel, A.; Yang, W.; Behel, A.; Alva, J.A.; Walter, G.; Head, C.; Ishikawa, T.O.; et al. Increased lysis of stem cells but not their differentiated cells by natural killer cells; de-differentiation or reprogramming activates NK cells. PLoS ONE 2010, 5, e11590. [Google Scholar] [CrossRef] [Green Version]
- Tseng, H.C.; Bui, V.; Man, Y.G.; Cacalano, N.; Jewett, A. Induction of Split Anergy Conditions Natural Killer Cells to Promote Differentiation of Stem Cells through Cell-Cell Contact and Secreted Factors. Front. Immunol. 2014, 5, 269. [Google Scholar] [CrossRef] [Green Version]
- Tseng, H.C.; Inagaki, A.; Bui, V.T.; Cacalano, N.; Kasahara, N.; Man, Y.G.; Jewett, A. Differential Targeting of Stem Cells and Differentiated Glioblastomas by NK Cells. J. Cancer 2015, 6, 866–876. [Google Scholar] [CrossRef] [Green Version]
- Bui, V.T.; Tseng, H.-C.; Maung, P.O.; Kozlowska, A.; Mann, K.; Topchyan, P.; Jewett, A. Augmented IFN-γ and TNF-α Induced by Probiotic Bacteria in NK Cells Mediate Differentiation of Stem-Like Tumors Leading to Inhibition of Tumor Growth and Reduction in Inflammatory Cytokine Release; Regulation by IL-10. Front. Immunol. 2015, 6, 576. [Google Scholar] [CrossRef] [Green Version]
- Jewett, A.; Bonavida, B. Target-induced inactivation and cell death by apoptosis in a subset of human NK cells. J. Immunol. 1996, 156, 907–915. [Google Scholar]
- Jewett, A.; Cavalcanti, M.; Bonavida, B. Pivotal role of endogenous TNF-alpha in the induction of functional inactivation and apoptosis in NK cells. J. Immunol. 1997, 159, 4815–4822. [Google Scholar]
- Jewett, A.; Bonavida, B. Interferon-alpha activates cytotoxic function but inhibits interleukin-2-mediated proliferation and tumor necrosis factor-alpha secretion by immature human natural killer cells. J. Clin. Immunol. 1995, 15, 35–44. [Google Scholar] [CrossRef]
- Jewett, A.; Wang, M.Y.; Teruel, A.; Poupak, Z.; Bostanian, Z.; Park, N.H. Cytokine dependent inverse regulation of CD54 (ICAM1) and major histocompatibility complex class I antigens by nuclear factor kappaB in HEp2 tumor cell line: Effect on the function of natural killer cells. Hum. Immunol. 2003, 64, 505–520. [Google Scholar] [CrossRef]
- Parisi, G.; Saco, J.D.; Salazar, F.B.; Tsoi, J.; Krystofinski, P.; Puig-Saus, C.; Zhang, R.; Zhou, J.; Cheung-Lau, G.C.; Garcia, A.J.; et al. Persistence of adoptively transferred T cells with a kinetically engineered IL-2 receptor agonist. Nat. Commun. 2020, 11, 660. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, M.L.; Nixon, M.J.; Gonzalez-Ericsson, P.I.; Bergman, R.E.; Pilkinton, M.A.; McDonnell, W.J.; Sanchez, V.; Opalenik, S.R.; Loi, S.; Zhou, J.; et al. Changes in Peripheral and Local Tumor Immunity after Neoadjuvant Chemotherapy Reshape Clinical Outcomes in Patients with Breast Cancer. Clin. Cancer Res. 2020, 26, 5668–5681. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, J.; Ghinnagow, R.; Seki, T.; Iketani, S.; Soulard, D.; Paczkowski, P.; Tsuji, Y.; MacKay, S.; Cruz, L.J.; et al. Targeted Co-delivery of Tumor Antigen and alpha-Galactosylceramide to CD141(+) Dendritic Cells Induces a Potent Tumor Antigen-Specific Human CD8(+) T Cell Response in Human Immune System Mice. Front. Immunol. 2020, 11, 2043. [Google Scholar] [CrossRef]
- Ma, C.; Cheung, A.F.; Chodon, T.; Koya, R.C.; Wu, Z.; Ng, C.; Avramis, E.; Cochran, A.J.; Witte, O.N.; Baltimore, D.; et al. Multifunctional T-cell analyses to study response and progression in adoptive cell transfer immunotherapy. Cancer Discov. 2013, 3, 418–429. [Google Scholar] [CrossRef]
- Rossi, J.; Paczkowski, P.; Shen, Y.W.; Morse, K.; Flynn, B.; Kaiser, A.; Ng, C.; Gallatin, K.; Cain, T.; Fan, R.; et al. Preinfusion polyfunctional anti-CD19 chimeric antigen receptor T cells are associated with clinical outcomes in NHL. Blood 2018, 132, 804–814. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Xue, Q.; Eisele, M.R.; Sulistijo, E.S.; Brower, K.; Han, L.; Amir el, A.D.; Pe’er, D.; Miller-Jensen, K.; Fan, R. Highly multiplexed profiling of single-cell effector functions reveals deep functional heterogeneity in response to pathogenic ligands. Proc. Natl. Acad. Sci. USA 2015, 112, E607–E615. [Google Scholar] [CrossRef] [Green Version]
- Iacoangeli, A.; Initiative, A.D.N.; Al Khleifat, A.; Jones, A.R.; Sproviero, W.; Shatunov, A.; Opie-Martin, S.; Morrison, K.E.; Shaw, P.; Shaw, C.E.; et al. C9orf72 intermediate expansions of 24–30 repeats are associated with ALS. Acta Neuropathol. Commun. 2019, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Cook, J.; Park, S.H.; Topchyan, P.; Kozlowska, A.; Ohanian, N.; Fang, C.; Nishimura, I.; Jewett, A. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front. Immunol. 2017, 8, 297. [Google Scholar] [CrossRef] [Green Version]
- Jewett, A.; Kos, J.; Kaur, K.; Turnsek, T.L.; Breznik, B.; Senjor, E.; Wong, P.; Nguyen, K.Y.; Ko, M.W. Multiple Defects of Natural Killer Cells in Cancer Patients: Anarchy, Dysregulated Systemic Immunity, and Immunosuppression in Metastatic Cancer. Crit. Rev. Immunol. 2020, 40, 93–133. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Arasteh, A.; Kaur, K.; Kozlowska, A.; Topchyan, P.; Jewett, A. Differential Cytotoxicity but Augmented IFN-gamma Secretion by NK Cells after Interaction with Monocytes from Humans, and Those from Wild Type and Myeloid-Specific COX-2 Knockout Mice. Front. Immunol. 2015, 6, 259. [Google Scholar] [CrossRef]
- Cacalano, N.A.; Le, D.; Paranjpe, A.; Wang, M.Y.; Fernandez, A.; Evazyan, T.; Park, N.H.; Jewett, A. Regulation of IGFBP6 gene and protein is mediated by the inverse expression and function of c-jun N-terminal kinase (JNK) and NFkappaB in a model of oral tumor cells. Apoptosis 2008, 13, 1439–1449. [Google Scholar] [CrossRef]
- Fu, X.Y. STAT3 in immune responses and inflammatory bowel diseases. Cell Res. 2006, 16, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Jewett, A.; Cacalano, N.A.; Teruel, A.; Romero, M.; Rashedi, M.; Wang, M.; Nakamura, H. Inhibition of nuclear factor kappa B (NFkappaB) activity in oral tumor cells prevents depletion of NK cells and increases their functional activation. Cancer Immunol. Immunother. 2006, 55, 1052–1063. [Google Scholar] [CrossRef]
- Kozlowska, A.K.; Topchyan, P.; Kaur, K.; Tseng, H.C.; Teruel, A.; Hiraga, T.; Jewett, A. Differentiation by NK cells is a prerequisite for effective targeting of cancer stem cells/poorly differentiated tumors by chemopreventive and chemotherapeutic drugs. J. Cancer 2017, 8, 537–554. [Google Scholar] [CrossRef]
- Dobbs, K.; Tabellini, G.; Calzoni, E.; Patrizi, O.; Martinez, P.; Giliani, S.C.; Moratto, D.; Al-Herz, W.; Cancrini, C.; Cowan, M.; et al. Natural Killer Cells from Patients with Recombinase-Activating Gene and Non-Homologous End Joining Gene Defects Comprise a Higher Frequency of CD56(bright) NKG2A(+++) Cells, and Yet Display Increased Degranulation and Higher Perforin Content. Front. Immunol. 2017, 8, 798. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Ko, M.W.; Ohanian, N.; Cook, J.; Jewett, A. Osteoclast-expanded super-charged NK-cells preferentially select and expand CD8+ T cells. Sci. Rep. 2020, 10, 20363. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, Y.; Feng, Y.; Jin, C.; Yang, Q.; Qiu, H.; Xie, H.; Xie, S.; Zhou, Y.; Huang, J. Expression of TLR2, TLR3, TLR4, and TLR7 on pulmonary lymphocytes of Schistosoma japonicum-infected C57BL/6 mice. Innate Immun. 2019, 25, 224–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewett, A.; Bonavida, B. Pentoxifylline suppresses interleukin-2-mediated activation of immature human natural killer cells by inhibiting endogenous tumor necrosis factor-alpha secretion. J. Clin. Immunol. 1994, 14, 31–38. [Google Scholar] [CrossRef]
- Cox, M.A.; Kahan, S.M.; Zajac, A.J. Anti-viral CD8 T cells and the cytokines that they love. Virology 2013, 435, 157–169. [Google Scholar] [CrossRef]
- Ivashkiv, L.B. IFNγ: Signalling, epigenetics and roles in immunity, metabolism, disease and cancer immunotherapy. Nat. Rev. Immunol. 2018, 18, 545–558. [Google Scholar] [CrossRef]
- Frank, K.; Paust, S. Dynamic Natural Killer Cell and T Cell Responses to Influenza Infection. Front. Cell. Infect. Microbiol. 2020, 10, 425. [Google Scholar] [CrossRef]
- Miller, C.H.T.; Maher, S.G.; Young, H.A. Clinical Use of Interferon-gamma. Ann. N. Y. Acad. Sci. 2009, 1182, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Whole Genome Sequencing Analyses | ||
|---|---|---|
| ALS Patient | Healthy | Shared Gene Mutations |
| TARDBP | C9orf72 | ALS2 |
| ERBB4 | HNRNPA1 | NEK1 |
| PRF1 | PRPH2 | |
| ANG | FIG4 | |
| SPG11 | ELP3 | |
| ATXN21 | SIGMAR1 | |
| FUS | SETX | |
| OPTN | ||
| ATXN2 | ||
| TRPM7 | ||
| PFN1 | ||
| SARM1 | ||
| TAF15 | ||
| UNC13A | ||
| NEFH | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, K.; Chen, P.-C.; Ko, M.-W.; Mei, A.; Chovatiya, N.; Huerta-Yepez, S.; Ni, W.; Mackay, S.; Zhou, J.; Maharaj, D.; et al. The Potential Role of Cytotoxic Immune Effectors in Induction, Progression and Pathogenesis of Amyotrophic Lateral Sclerosis (ALS). Cells 2022, 11, 3431. https://doi.org/10.3390/cells11213431
Kaur K, Chen P-C, Ko M-W, Mei A, Chovatiya N, Huerta-Yepez S, Ni W, Mackay S, Zhou J, Maharaj D, et al. The Potential Role of Cytotoxic Immune Effectors in Induction, Progression and Pathogenesis of Amyotrophic Lateral Sclerosis (ALS). Cells. 2022; 11(21):3431. https://doi.org/10.3390/cells11213431
Chicago/Turabian StyleKaur, Kawaljit, Po-Chun Chen, Meng-Wei Ko, Ao Mei, Nishant Chovatiya, Sara Huerta-Yepez, Weiming Ni, Sean Mackay, Jing Zhou, Dipanarine Maharaj, and et al. 2022. "The Potential Role of Cytotoxic Immune Effectors in Induction, Progression and Pathogenesis of Amyotrophic Lateral Sclerosis (ALS)" Cells 11, no. 21: 3431. https://doi.org/10.3390/cells11213431