
Deep Insight of the Pathophysiology of Gestational Diabetes Mellitus
 , , ,
, , , 

Abstract
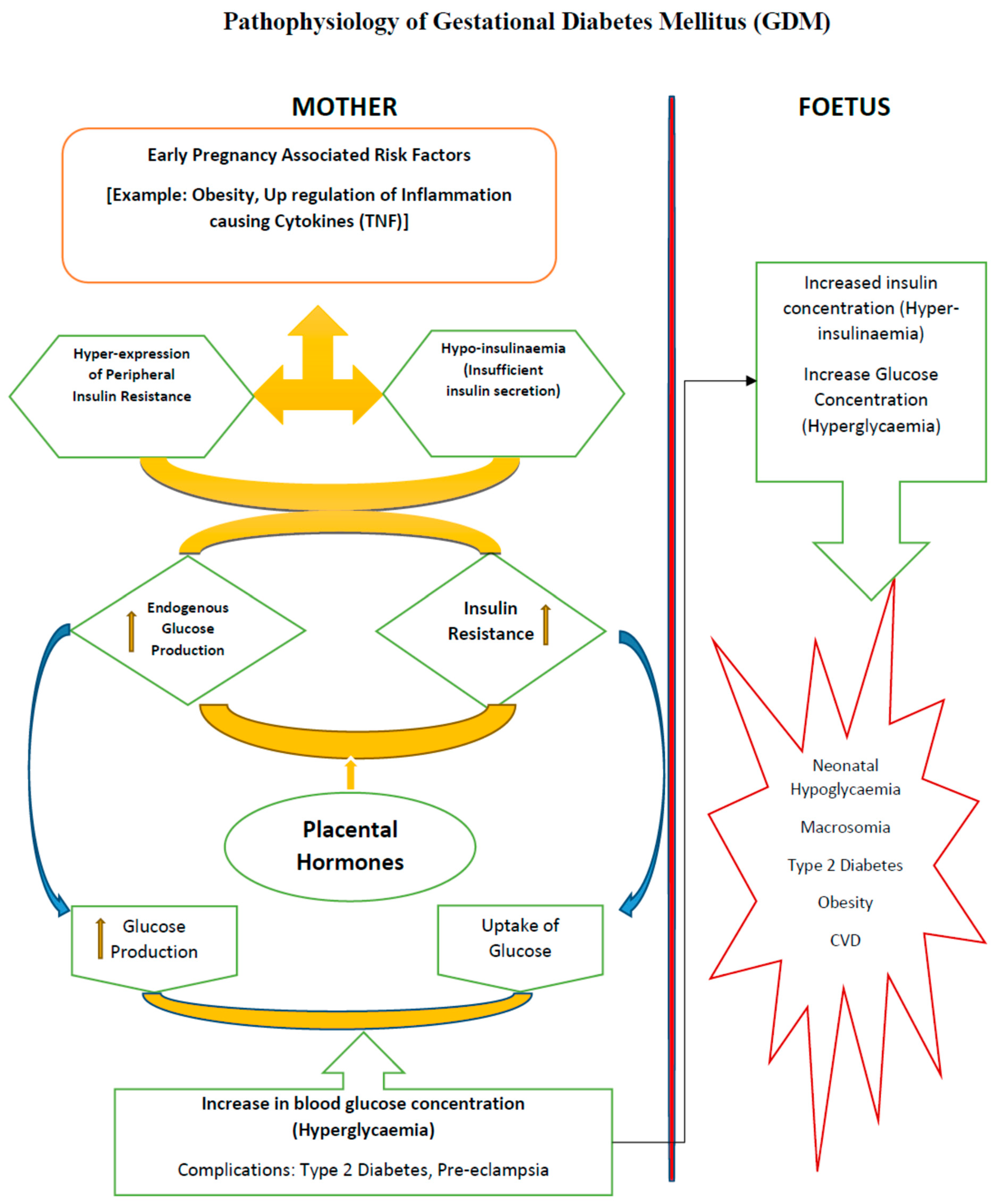
:1. Introduction
2. Obesity, a Major Predisposing Factor for GDM
3. Insulin Resistance
4. Adipokines and Hormones in the Pathophysiology of GDM
5. Foetal Derangements in GDM
6. Pathophysiology and Pharmacological Interventions in GDM
7. Vascular Endothelial Growth Factor—A Silent Player in GDM
8. Predictive Biomarkers for the Early Diagnosis of GDM
9. Role of microRNA in GDM
10. In Vitro and In Vivo Models for GDM Study
11. Future Perspectives
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kapur, A. Links between maternal health and NCDs. Best Pract. Res. Clin. Obstet. Gynaecol. 2015, 29, 32–42. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes. Diabetes Care 2018, 41, S13–S27. [CrossRef]
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; IDF: Brussels, Belgium, 2017. [Google Scholar]
- Goyal, A.; Gupta, Y.; Singla, R.; Kalra, S.; Tandon, N. American Diabetes Association “Standards of Medical Care—2020 for Gestational Diabetes Mellitus”: A Critical Appraisal. Diabetes Ther. 2020, 1, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Feig, D.S.; Moses, R.G. Metformin Therapy during Pregnancy Good for the goose and good for the gosling too? Diabetes Care 2011, 34, 2329–2330. [Google Scholar] [CrossRef] [PubMed]
- Camelo Castillo, W.; Boggess, K.; Stürmer, T.; Brookhart, M.A.; Benjamin, D.K.; Jonsson Funk, M. Association of Adverse Pregnancy Outcomes with Glyburide vs. Insulin in Women with Gestational Diabetes. JAMA Pediatr. 2015, 169, 452–458. [Google Scholar] [CrossRef]
- Trout, K.K.; Compher, C.; Durnwald, C.P. Protein vs. Carbohydrate in Gestational Diabetes. Diabetes 2019, 68, 753-P. [Google Scholar] [CrossRef]
- Dabelea, D.; Snell-Bergeon, J.K.; Hartsfield, C.L.; Bischoff, K.J.; Hamman, R.F.; McDuffie, R.S. Increasing prevalence of gestational diabetes mellitus (GDM) over time and by birth cohort: Kaiser Permanente of Colorado GDM screening program. Diabetes Care 2005, 28, 579–584. [Google Scholar] [CrossRef]
- Moses, R.G.; Morris, G.J.; Petocz, P.; Sangil, F.; Garg, D. The impact of potential new diagnostic criteria on the prevalence of gestational diabetes mellitus in Australia. Med. J. Aust. 2011, 194, 338–340. [Google Scholar] [CrossRef]
- HAPO Study Cooperative Research Group. Hyperglycemia and adverse pregnancy outcomes. N. Engl. J. Med. 2008, 358, 1991–2002. [Google Scholar] [CrossRef]
- Solomon, C.G.; Willett, W.C.; Carey, V.J.; Rich-Edwards, J.; Hunter, D.J.; Colditz, G.A.; Stampfer, M.J.; Speizer, F.E.; Spiegelman, D.; Manson, J.E. A prospective study of pregravid determinants of gestational diabetes mellitus. J. Am. Med. Assoc. 1997, 278, 1078–1083. [Google Scholar] [CrossRef]
- Chiefari, E.; Arcidiacono, B.; Foti, D.; Brunetti, A. Gestational diabetes mellitus: An updated overview. J. Endocrinol. Investig. 2017, 40, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Riskin-Mashiah, S.; Damti, A.; Younes, G.; Auslender, R. First trimester fasting hyperglycemia as a predictor for the development of gestational diabetes mellitus. Eur. J. Obstet. Gynecol. Reprod. Biol. 2010, 152, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Maged, A.M.; Moety, G.A.F.; Mostafa, W.A.; Hamed, D.A. Comparative study between different biomarkers for early prediction of gestational diabetes mellitus. J. Matern. Fetal Neonatal Med. 2014, 27, 1108–1112. [Google Scholar] [CrossRef] [PubMed]
- Teh, W.T.; Teede, H.J.; Paul, E.; Harrison, C.L.; Wallace, E.M.; Allan, C. Risk factors for gestational diabetes mellitus: Implications for the application of screening guidelines. Aust. N. Z. J. Obstet. Gynaecol. 2011, 51, 26–30. [Google Scholar] [CrossRef]
- Ben-Haroush, A.; Yogev, Y.; Hod, M. Epidemiology of gestational diabetes mellitus and its association with Type 2 diabetes. Diabet. Med. 2004, 21, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Permana, P.A.; Menge, C.; Reaven, P.D. Macrophage-secreted factors induce adipocyte inflammation and insulin resistance. Biochem. Biophys. Res. Commun. 2006, 341, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Kralisch, S.; Bluher, M.; Paschke, R.; Stumvoll, M.; Fasshauer, M. Adipokines and adipocyte targets in the future management of obesity and the metabolic syndrome. Mini-Rev. Med. Chem. 2007, 7, 39–45. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Wolf, M.; Sauk, J.; Shah, A.; VossenSmirnakis, K.; Jimenez-Kimble, R.; Ecker, J.L.; Thadhani, R. Inflammation and glucose intolerance: A prospective study of gestational diabetes mellitus. Diabetes Care 2004, 27, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Qiu, C.; Sorensen, T.K.; Luthy, D.A.; Williams, M.A. A prospective study of maternal serum C-reactive protein (CRP) concentrations and risk of gestational diabetes mellitus. Paediatr. Perinat. Epidemiol. 2004, 18, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Catalano, P.M.; Huston, L.; Amini, S.B.; Kalhan, S.C. Longitudinal changes in glucose metabo-lism during pregnancy in obese women with normal glucose tolerance and gestational diabetes mellitus. Am. J. Obstet. Gynecol. 1999, 180, 903–916. [Google Scholar] [CrossRef]
- Buchanan, T.A. Pancreatic B cell defects in gestational diabetes: Implications for the pathogenesis and prevention of type 2 diabetes. J. Clin. Endocrinol. Metab. 2001, 86, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.E.; Ishizuka, T.; Shao, J.; Huston, L.; Highman, T.; Catalano, P. Impaired glucose transport and insulin receptor tyrosine phosphorylation in skeletal muscle from obese women with gestational diabetes. Diabetes 1999, 48, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Catalano, P.M.; Tyzbir, E.D.; Roman, N.M.; Amini, S.B.; Sims, E.A. Longitudinal changes in insulin release and in-sulin resistance in nonobese pregnant women. Am. J. Obs. Gynecol. 1991, 165, 1667–1672. [Google Scholar] [CrossRef]
- Catalano, P.M. Obesity, insulin resistance, and pregnancy outcome. Reproduction 2010, 140, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Ategbo, J.M.; Grissa, O.; Yessoufou, A.; Hichami, A.; Dramane, K.L.; Moutairou, K.; Miled, A.; Grissa, A.; Jerbi, M.; Tabka, Z.; et al. Modulation of adipokines and cytokines in gestational diabetes and macrosomia. J. Clin. Endocrinol. Metab. 2006, 91, 4137–4143. [Google Scholar] [CrossRef]
- Kirwan, J.P.; Hauguel-De Mouzon, S.; Lepercq, J.; Challier, J.C.; HustonPresley, L.; Friedman, J.E.; Kalhan, S.C.; Catalano, P.M. TNF-alpha is a predictor of insulin resistance in human pregnancy. Diabetes 2002, 51, 2207–2213. [Google Scholar] [CrossRef]
- Berggren, E.K.; Roeder, H.A.; Boggess, K.A.; Moss, K.; Offenbacher, S.; Campbell, E.; Grotegut, C.A. First-trimester maternal serum C-reactive protein as a predictor of third-trimester impaired glucose tolerance. Reprod. Sci. 2015, 22, 90–93. [Google Scholar] [CrossRef]
- Georgiou, H.M.; Lappas, M.; Georgiou, G.M.; Marita, A.; Bryant, V.J.; Hiscock, R.; Permezel, M.; Khalil, Z.; Rice, G. Screening for biomarkers predictive of gestational diabetes mellitus. Acta Diabetol. 2008, 45, 157–165. [Google Scholar] [CrossRef]
- Van Assche, F.A.; Aerts, L.; De Prins, F. A morphological study of the endocrine pancreas in human pregnancy. Br. J. Obstet. Gynaecol. 1978, 85, 818–820. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, H.D.; Catalano, P.; Zhang, C.; Desoye, G.; Mathiesen, E.R.; Damm, P. Gestational diabetes mellitus. Nat. Rev. Dis. Primers 2019, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Chandran, M.; Phillips, S.A.; Ciaraldi, T.; Henry, R.R. Adiponectin: More than just another fat cell hormone? Diabetes Care 2003, 26, 2442–2450. [Google Scholar] [CrossRef]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K.; et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. 1999. Biochem. Biophys. Res. Commun. 1999, 257, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Spranger, J.; Kroke, A.; Mohlig, M.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Adiponectin and protection against type 2 diabetes mellitus. Lancet 2003, 361, 226–228. [Google Scholar] [CrossRef]
- Nakashima, R.; Kamei, N.; Yamane, K.; Nakanishi, S.; Nakashima, A.; Kohno, N. Decreased total and high molecular weight adiponectin are independent risk factors for the development of type 2 diabetes in Japanese-Americans. J. Clin. Endocrinol. Metab. 2006, 91, 3873–3877. [Google Scholar] [CrossRef]
- Doruk, M.; Ugur, M.; Oruc, A.S.; Demirel, N.; Yildiz, Y. Serum adiponectin in gestational diabetes and its relation to pregnancy outcome. J. Obstet. Gynaecol. 2014, 34, 471–475. [Google Scholar] [CrossRef]
- Pala, H.G.; Ozalp, Y.; Yener, A.S.; Gerceklioglu, G.; Uysal, S.; Onvural, A. Adiponectin levels in gestational diabetes mellitus and in pregnant women without glucose intolerance. Adv. Clin. Exp. Med. 2015, 24, 85–92. [Google Scholar] [CrossRef]
- Tsai, P.J.; Yu, C.H.; Hsu, S.P.; Lee, Y.H.; Huang, I.T.; Ho, S.C.; Chu, C.H. Maternal plasma adiponectin concentrations at 24 to 31 weeks of gestation: Negative association with gestational diabetes mellitus. Nutrition 2005, 21, 1095–1099. [Google Scholar] [CrossRef]
- Soheilykhah, S.; Mohammadi, M.; Mojibian, M.; Rahimi-Saghand, S.; Rashidi, M.; Hadinedoushan, H.; Afkhami-Ardekani, M. Maternal serum adiponectin concentration in gestational diabetes. Gynecol. Endocrinol. 2009, 25, 593–596. [Google Scholar] [CrossRef]
- Wauters, M.; Considine, R.V.; Van Gaal, L.F. Human leptin: From an adipocyte hormone to an endocrine mediator. Eur. J. Endocrinol. 2000, 143, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Fasshauer, M.; Bluher, M.; Stumvoll, M. Adipokines in gestational diabetes. Lancet Diabetes Endocrinol. 2014, 2, 488–499. [Google Scholar] [CrossRef]
- Miehle, K.; Stepan, H.; Fasshauer, M. Leptin, adiponectin and other adipokines in gestational diabetes mellitus and pre-eclampsia. Clin. Endocrinol. 2012, 76, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Williams, M.A.; Vadachkoria, S.; Frederick, I.O.; Luthy, D.A. Increased maternal plasma leptin in early pregnancy and risk of gestational diabetes mellitus. Obstet. Gynecol. 2004, 103, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Adeghate, E. Visfatin: Structure, function and relation to diabetes mellitus and other dysfunctions. Curr. Med. Chem. 2008, 15, 1851–1862. [Google Scholar] [CrossRef]
- Mazaki-Tovi, S.; Romero, R.; Kusanovic, J.P.; Vaisbuch, E.; Erez, O.; Than, N.G.; Chaiworapongsa, T.; Nhan-Chang, C.L.; Pacora, P.; Gotsch, F.; et al. Maternal visfatin concentration in normal pregnancy. J. Perinat. Med. 2009, 37, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.F.; Rezende, J.C.; Vaikousi, E.; Akolekar, R.; Nicolaides, K.H. Maternal serum visfatin at 11–13 weeks of gestation in gestational diabetes mellitus. Clin. Chem. 2011, 57, 609–613. [Google Scholar] [CrossRef]
- Smirnakis, K.V.; Plati, A.; Wolf, M.; Thadhani, R.; Ecker, J.L. Predicting gestational diabetes: Choosing the optimal early serum marker. Am. J. Obstet. Gynecol. 2007, 196, e1–e7. [Google Scholar] [CrossRef]
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The Role of Placental Hormones in Mediating Maternal Adapta-tions to Support Pregnancy and Lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef]
- Valencia-Ortega, J.; González-Reynoso, R.; Ramos-Martínez, E.G.; Ferreira-Hermosillo, A.; Peña-Cano, M.I.; Morales-Ávila, E.; Saucedo, R. New Insights into Adipokines in Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2022, 23, 6279. [Google Scholar] [CrossRef]
- Bao, W.; Michels, K.B.; Tobias, D.K.; Li, S.; Chavarro, J.E.; Gaskins, A.J.; Vaag, A.A.; Hu, F.B.; Zhang, C. Parental smoking during pregnancy and the risk of gestational diabetes in the daughter. Int. J. Epidemiol. 2016, 45, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Pettitt, D.J.; Knowler, W.C.; Baird, H.R.; Bennett, P.H. Gestational diabetes: Infant and maternal complications of pregnancy in relation to third-trimester glucose tolerance in the Pima Indians. Diabetes Care 1980, 3, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Jensen, D.M.; Damm, P.; Sørensen, B.; Mølsted-Pedersen, L.; Westergaard, J.G.; Korsholm, L.; Ovesen, P.; Beck-Nielsen, H. Proposed diagnostic thresholds for gestational diabetes mellitus according to a 75-g oral glucose tolerance test. Maternal and perinatal outcomes in 3260 Danish women. Diabet Med. 2003, 20, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Huang, L.; Wang, M.; Chen, D.; Chen, Z.; Jiang, S.W. A Critical Review of Proteomic Studies in Gestational Diabetes Mellitus. J. Diabetes Res. 2020, 6450352. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A. From the Triumvirate to the Ominous Octet: A New Paradigm for the Treatment of Type 2 Diabetes Mellitus. Diabetes 2009, 58, 773–795. [Google Scholar] [CrossRef]
- Prentki, M.; Nolan, C.J. Islet beta cell failure in type 2 diabetes. J. Clin. Investig. 2006, 116, 1802–1812. [Google Scholar] [CrossRef]
- Ashcroft, F.M.; Rohm, M.; Clark, A.; Brereton, M.F. Is Type 2 Diabetes a Glycogen Storage Disease of Pancreatic β Cells? Cell Metab. 2017, 26, 17–23. [Google Scholar] [CrossRef]
- Delghingaro-Augusto, V.; Nolan, C.J.; Gupta, D.; Jetton, T.L.; Latour, M.G.; Peshavaria, M.; Madiraju, S.R.M.; Joly, E.; Peyot, M.L.; Prentki, M.; et al. Islet beta cell failure in the 60% pancreatectomised obese hyperlipidaemic Zucker fatty rat: Severe dysfunction with altered glycerolipid metabolism without steatosis or a falling beta cell mass. Diabetologia 2009, 52, 1122–1132. [Google Scholar] [CrossRef]
- Simmons, R.A.; Templeton, L.J.; Gertz, S.J. Intrauterine growth retardation leads to the development of type 2 diabetes in the rat. Diabetes 2001, 50, 2279–2286. [Google Scholar] [CrossRef]
- Auffret, J.; Freemark, M.; Carré, N.; Mathieu, Y.; Tourrel-Cuzin, C.; Lombès, M.; Movassat, J.; Binart, N. Defective prolactin signaling impairs pancreatic β-cell development during the perinatal period. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1309–E1318. [Google Scholar] [CrossRef] [Green Version]
- Rahier, J.; Guiot, Y.; Goebbels, R.M.; Sempoux, C.; Henquin, J.C. Pancreatic beta-cell mass in European subjects with type 2 diabetes. Diabetes Obes. Metab. 2008, 10, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Song, Y.; Chen, D.B.; Zheng, J. Protein phosphatase 3 differentially modulates vascular endothelial growth factor- and fibroblast growth factor 2-stimulated cell proliferation and signaling in ovine fetoplacental artery endothelial cells. Biol. Reprod. 2008, 79, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Leach, L.; Taylor, A.; Sciota, F. Vascular dysfunction in the diabetic placenta: Causes and consequences. J. Anat. 2009, 215, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Lang, I.; Pabst, M.A.; Hiden, U.; Blaschitz, A.; Dohr, G.; Hahn, T.; Desoye, G. Heterogeneity of microvascular endothelial cells iso-lated from human term placenta and macrovascu- lar umbilical vein endothelial cells. Eur. J. Cell Biol. 2003, 82, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Dye, J.; Lawrence, L.; Linge, C.; Leach, L.; Firth, J.; Clark, P. Distinct patterns of microvascular endothelial cell morphology are determined by extracellular matrix composition. Endothelium 2004, 11, 151–167. [Google Scholar] [CrossRef]
- San Martin, R.; Sobrevia, L. Gestational diabetes and the adenosine/L-arginine/nitric oxide (ALANO) pathway in human umbilical vein endothelium. Placenta 2006, 27, 1–10. [Google Scholar] [CrossRef]
- Escudero, C.; Puebla, C.; Westermeier, F.; Sobrevia, L. Potential cell signalling mechanisms involved in differential placental angiogenesis in mild and severe preeclampsia. Curr. Vasc. Pharm. 2009, 7, 475–485. [Google Scholar] [CrossRef]
- Baumgartner-Parzer, S.M.; Wagner, L.; Pettermann, M.; Grillari, J.; Gessl, A.; Waldhäusl, W. High-glucose-triggered apoptosis in cultured endothelial cells. Diabetes 1995, 44, 1323–1327. [Google Scholar] [CrossRef]
- Sultan, S.A.; Liu, W.; Peng, Y.; Roberts, W.; Whitelaw, D.; Graham, A.M. The Role of Maternal Gestational Diabetes in Inducing Fetal Endothelial Dysfunction. J. Cell. Physiol. 2015, 230, 2695–2705. [Google Scholar] [CrossRef]
- Grissa, O.; Yessoufou, A.; Mrisak, I.; Hichami, A.; Amoussou-Guenou, D.; Grissa, A.; Djrolo, F.; Moutairou, K.; Miled, A.; Khairi, H.; et al. Growth factor concentrations and their placental mRNA expression are modulated in gestational diabetes mellitus: Possible interactions with macrosomia. BMC Pregnancy Childbirth 2010, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Perez, F.; Hiden, U.; Gauster, M.; Lang, I.; Konya, V.; Heinemann, A.; Lögl, J.; Saffery, R.; Desoye, G.; Cvitic, S. Post-transcriptional down regulation of ICAM-1 in feto-placental endothelium in GDM. Cell Adh. Migr. 2016, 10, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Marcantoni, E.; Dovizio, M.; O’Gaora, P.; Di Francesco, L.; Bendaya, I.; Schiavone, S.; Trenti, A.; Guillem-Llobat, P.; Zambon, A.; Nardelli, G.B.; et al. Dysregulation of gene expression in human fetal endothelial cells from gestational diabetes in response to TGF-β1. Prostaglandins Other Lipid Mediat. 2015, 120, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.; Rohrbach, A.; Borns, K.; Hillemanns, P.; Feng, L.; Hubel, C.A.; von Versen-Höynck, F. Vitamin D rescues dysfunction of fetal endothelial colony forming cells from individuals with gestational diabetes. Placenta 2015, 36, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Ozmen, A.; Unek, G.; Kipmen-Korgun, D.; Korgun, E.T. The PI3K/Akt and MAPK-ERK1/2 pathways are altered in STZ induced diabetic rat placentas. Histol. Histopathol. 2014, 29, 743–756. [Google Scholar]
- Madazli, R.; Tuten, A.; Calay, Z.; Uzun, H.; Uludag, S.; Ocak, V. The incidence of placental abnormalities, maternal and cord plasma malondialdehyde and vascular endothelial growth factor levels in women with gestational diabetes mellitus and nondiabetic controls. Gynecol. Obstet. Investig. 2008, 65, 227–232. [Google Scholar] [CrossRef]
- Jirkovska, M.; Kubinova, L.; Janacek, J.; Moravcova, M.; Krejci, V.; Karen, P. Topological properties and spatial organization of villous capillaries in normal and diabetic placentas. J. Vasc. Res. 2002, 39, 268–278. [Google Scholar] [CrossRef]
- Meng, Q.; Shao, L.; Luo, X.; Mu, Y.; Xu, W.; Gao, L.; Xu, H.; Cui, Y. Expressions of VEGF-A and VEGFR-2 in placentae from GDM pregnancies. Reprod. Biol. Endocrinol. 2016, 14, 61. [Google Scholar] [CrossRef]
- Leach, L.; Gray, C.; Staton, S.; Babawale, M.O.; Gruchy, A.; Foster, C.; Mayhew, T.M.; James, D.K. Vascular endothelial cadherin and beta-catenin in human fetoplacental vessels of pregnancies complicated by Type 1 diabetes: Associations with angiogenesis and perturbed barrier function. Diabetologia 2004, 47, 695–709. [Google Scholar] [CrossRef]
- Dubova, E.A.; Pavlov, K.A.; Esayan, R.M.; Degtyareva, E.I.; Shestakova, M.V.; Shchegolev, A.I.; Sukhikh, G.T. Vascular endothelial growth factor and its receptors in the placenta of women with type 1 diabetes mellitus. Bull. Exp. Biol. Med. 2012, 152, 367–370. [Google Scholar] [CrossRef]
- Daskalakis, G.; Marinopoulos, S.; Krielesi, V.; Papapanagiotou, A.; Papantoniou, N.; Mesogitis, S.; Antsaklis, A. Placental pathology in women with gestational diabetes. Acta Obstet. Gynecol. Scand. 2008, 87, 403–407. [Google Scholar] [CrossRef]
- Aitken, N.; Smith, S.; Schwarz, C.; Morin, P.A. Single nucleotide polymorphism (SNP) discovery in mammals: A targeted-gene approach. Mol. Ecol. 2004, 13, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Hong, S.H. Association between the polymorphisms of the vascular endothelial growth factor gene and metabolic syndrome. Biomed. Rep. 2015, 3, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Yang, F.; Urban, T.J.; Li, L.; Chalasani, N.; Flockhart, D.A.; Liu, W. Impact of the Interaction between 3’-UTR SNPs and microRNA on the expression of human xenobiotic metabolism enzyme and transporter genes. Front. Genet. 2012, 3, 248. [Google Scholar] [CrossRef] [PubMed]
- Haas, U.; Sczakiel, G.; Laufer, S.D. MicroRNA-mediated regulation of gene expression is affected by disease-associated SNPs within the 3’-UTR via altered RNA structure. RNA Biol. 2012, 9, 924–937. [Google Scholar] [CrossRef]
- Sung, K.C.; Reaven, G.M.; Kim, S.H. Utility of homeostasis model assessment of beta-cell function in predicting diabetes in 12,924 healthy Koreans. Diabetes Care 2010, 33, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Su, M.T.; Lin, S.H.; Lee, I.W.; Chen, Y.C.; Kuo, P.L. Association of polymorphisms/haplotypes of the genes encoding vascular endothelial growth factor and its KDR receptor with recurrent pregnancy loss. Hum. Reprod. 2011, 26, 758–764. [Google Scholar] [CrossRef]
- Bao, W.; Tobias, D.K.; Olsen, S.F.; Zhang, C. Pre-pregnancy fried food consumption and the risk of gestational diabetes mellitus: A prospective cohort study. Diabetologia 2014, 57, 2485–2491. [Google Scholar] [CrossRef]
- Zhou, J.; Ni, X.; Huang, X.; Yao, J.; He, Q.; Wang, K.; Duan, T. Potential Role of Hyperglycemia in Fetoplacental Endothelial Dysfunction in Gestational Diabetes Mellitus. Cell. Physiol. Biochem. 2016, 39, 1317–1328. [Google Scholar] [CrossRef]
- Huhn, E.A.; Rossi, S.W.; Hoesli, I.; Göbl, C.S. Controversies in Screening and Diagnostic Criteria for Gestational Diabetes in Early and Late Pregnancy. Front. Endocrinol. 2018, 9, 696. [Google Scholar] [CrossRef]
- Shiefa, S.; Amargandhi, M.; Bhupendra, J.; Moulali, S.; Kristine, T. First Trimester Maternal Serum Screening Using Biochemical Markers PAPP-A and Free beta-hCG for Down Syndrome, Patau Syndrome and Edward Syndrome. Indian J. Clin. Biochem. 2013, 28, 3–12. [Google Scholar] [CrossRef]
- Husslein, H.; Lausegger, F.; Leipold, H.; Worda, C. Association between pregnancy-associated plasma protein-A and gestational diabetes requiring insulin treatment at 11–14 weeks of gestation. J. Matern. Fetal Neonatal Med. 2012, 25, 2230–2233. [Google Scholar] [CrossRef] [PubMed]
- Quattrocchi, T.; Baviera, G.; Pochiero, T.; Basile, F.; Rizzo, L.; Santamaria, A.; Corrado, F.; D’Anna, R. Maternal serum PAPP-A as an early marker of obstetric complications? Fetal Diagn. Ther. 2015, 37, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Sirikunalai, P.; Wanapirak, C.; Sirichotiyakul, S.; Tongprasert, F.; Srisupundit, K.; Luewan, S.; Traisrisilp, K.; Tongsong, T. Associations between maternal serum free beta human chorionic gonadotropin (β-hCG) levels and adverse pregnancy outcomes. J. Obstet. Gynaecol. 2016, 36, 178–182. [Google Scholar] [CrossRef]
- Sweeting, A.N.; Wong, J.; Appelblom, H.; Ross, G.P.; Kouru, H.; Williams, P.F.; Sairanen, M.; Hyett, J.A. A first trimester prediction model for gestational diabetes utilizing aneuploidy and pre-eclampsia screening markers. J. Matern. Fetal Neonatal Med. 2018, 31, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Menegazzo, L.; Ciciliot, S.; Poncina, N.; Mazzucato, M.; Persano, M.; Bonora, B.; Albiero, M.; Vigili de Kreutzenberg, S.; Avogaro, A.; Fadini, G.P. NETosis is induced by high glucose and associated with type 2 diabetes. Acta Diabetol. 2015, 52, 497–503. [Google Scholar] [CrossRef]
- Moreli, J.B.; Morceli, G.; De Luca, A.K.; Magalhaes, C.G.; Costa, R.A.; Damasceno, D.C.; Rudge, M.V.; Calderon, I.M. Influence of maternal hyperglycemia on IL-10 and TNF-alpha production: The relationship with perinatal outcomes. J. Clin. Immunol. 2012, 32, 604–610. [Google Scholar] [CrossRef]
- Gupta, A.K.; Hasler, P.; Holzgreve, W.; Hahn, S. Neutrophil NETs: A novel contributor to preeclampsia-associated placental hypoxia? Semin. Immunopathol. 2007, 29, 163–167. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, Y.; Zhong, L.; Ye, D.; Zhang, J.; Tu, Y.; Bornstein, S.R.; Zhou, Z.; Lam, K.S.; Xu, A. Increased neutrophil elastase and proteinase 3 and augmented NETosis are closely associated with beta-cell autoimmunity in patients with type 1 diabetes. Diabetes 2014, 63, 4239–4248. [Google Scholar] [CrossRef]
- Liu, Z.; Jiang, Y.; Liu, X.; Yang, M.; Chen, C.; Zhao, B.; Huang, H.; Luo, Q. MiRNAs in Gestational Diabetes Mellitus: Potential Mechanisms and Clinical Applications. J. Diabetes Res. 2021, 2021, 4632745. [Google Scholar] [CrossRef]
- Kokkinopoulou, I.; Maratou, E.; Mitrou, P.; Boutati, E.; Sideris, D.C.; Fragoulis, E.G.; Christodoulou, M.I. Decreased expression of microRNAs targeting type-2 diabetes susceptibility genes in peripheral blood of patients and predisposed individuals. Endocrine 2019, 66, 226–239. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, T.; Shi, Z.; Ding, H.; Ling, X. MicroRNA-518d regulates PPARα protein expression in the placentas of females with gestational diabetes mellitus. Mol. Med. Rep. 2014, 9, 2085–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mac-Marcjanek, K.; Zieleniak, A.; Zurawska-Klis, M.; Cypryk, K.; Wozniak, L.; Wojcik, M. Expression Profile of Diabetes-Related Genes Associated with Leukocyte Sirtuin 1 Overexpression in Gestational Diabetes. Int. J. Mol. Sci. 2018, 19, 3826. [Google Scholar] [CrossRef] [PubMed]
- Tagoma, A.; Alnek, K.; Kirss, A.; Uibo, R.; Haller-Kikkatalo, K. MicroRNA profiling of second trimester maternal plasma shows upregulation of miR-195-5p in patients with gestational diabetes. Gene 2018, 672, 137–142. [Google Scholar] [CrossRef]
- Shi, Z.; Zhao, C.; Guo, X.; Ding, H.; Cui, Y.; Shen, R.; Liu, J. Differential expression of microRNAs in omental adipose tissue from gestational diabetes mellitus subjects reveals miR-222 as a regulator of ERα expression in estrogen-induced insulin resistance. Endocrinology 2014, 155, 1982–1990. [Google Scholar] [CrossRef]
- Cao, J.L.; Zhang, L.; Li, J.; Tian, S.; Lv, X.D.; Wang, X.Q.; Su, X.; Li, Y.; Hu, Y.; Ma, X.; et al. Up-regulation of miR-98 and unraveling regulatory mechanisms in gestational diabetes mellitus. Sci. Rep. 2016, 6, 32268. [Google Scholar] [CrossRef] [PubMed]
- Stirm, L.; Huypens, P.; Sass, S.; Batra, R.; Fritsche, L.; Brucker, S.; Abele, H.; Hennige, A.M.; Theis, F.; Beckers, J.; et al. Maternal whole blood cell miRNA-340 is elevated in gestational diabetes and inversely regulated by glucose and insulin. Sci. Rep. 2018, 8, 1. [Google Scholar]
- Wang, Y.C.; Li, Y.; Wang, X.Y.; Zhang, D.; Zhang, H.; Wu, Q.; He, Y.Q.; Wang, J.Y.; Zhang, L.; Xia, H.; et al. Circulating miR-130b mediates metabolic crosstalk between fat and muscle in overweight/obesity. Diabetologia 2013, 56, 2275–2285. [Google Scholar] [CrossRef]
- Tryggestad, J.B.; Vishwanath, A.; Jiang, S.N.; Mallappa, A.; Teague, A.M.; Takahashi, Y.; Thompson, D.M.; Chernausek, S.D. Influence of gestational diabetes mellitus on human umbilical vein endothelial cell miRNA. Clin. Sci. 2016, 130, 1955–1967. [Google Scholar] [CrossRef]
- Collares, C.V.; Evangelista, A.F.; Xavier, D.J.; Rassi, D.M.; Arns, T.; Foss-Freitas, M.C.; Foss, M.C.; Puthier, D.; Sakamoto-Hojo, E.T.; Passos, G.A.; et al. Identifying common and specific microRNAs expressed in peripheral blood mononuclear cell of type 1, type 2, and gestational diabetes mellitus patients. BMC Res. Notes 2013, 26, 491. [Google Scholar] [CrossRef]
- Juchnicka, I.; Kuźmicki, M.; Niemira, M.; Bielska, A.; Sidorkiewicz, I.; Zbucka-Krętowska, M.; Krętowski, A.J.; Szamatowicz, J. miRNAs as Predictive Factors in Early Diagnosis of Gestational Diabetes Mellitus. Front. Endocrinol. 2022, 13, 839344. [Google Scholar] [CrossRef]
- Wander, P.L.; Boyko, E.J.; Hevner, K.; Parikh, V.J.; Tadesse, M.G.; Sorensen, T.K.; Williams, M.A.; Enquobahrie, D.A. Circulating early- and mid-pregnancy microRNAs and risk of gestational diabetes. Diabetes Res. Clin. Pract. 2017, 132, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.Y.; Tian, S.; Cao, J.L.; Wang, X.Q.; Ma, X.; Xia, H.F. Down-Regulated miR-21 in Gestational Diabetes Mellitus Placenta Induces PPAR-α to Inhibit Cell Proliferation and Infiltration. Diabetes Metab. Syndr. Obes. 2020, 13, 3009–3034. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.L.; Chen, T.; Xiong, J.L.; Wu, D.; Xi, Q.Y.; Luo, J.Y.; Sun, J.J.; Zhang, Y.L. miR-146b Inhibits Glucose Consumption by Targeting IRS1 Gene in Porcine Primary Adipocytes. Int. J. Mol. Sci. 2018, 19, 783. [Google Scholar] [CrossRef]
- Cao, X.; Lu, B.; Gu, Y.; Li, X.; Guo, D.; Xia, F. miR-210-3p Impairs Pancreatic β-Cell Function by Targeting Dtx1 in Gestational Diabetes Mellitus. J. Environ. Pathol. Toxicol. Oncol. 2022, 41, 11–23. [Google Scholar] [CrossRef]
- Sadeghzadeh, S.; Ashkezari, M.D.; Seifati, S.M.; Mehrjardi, M.Y.V.; Tezerjani, M.D.; Sadeghzadeh, S.; Ladan, S.A.B. Circulating miR-15a and miR-222 as Potential Biomarkers of Type 2 Diabetes. Diabetes Metab. Syndr. Obes. 2020, 13, 3461–3469. [Google Scholar] [CrossRef]
- Lilao-Garzón, J.; Valverde-Tercedor, C.; Muñoz-Descalzo, S.; Brito-Casillas, Y.; Wägner, A.M. In Vivo and In Vitro Models of Diabetes: A Focus on Pregnancy. Adv. Exp. Med. Biol. 2021, 1307, 553–576. [Google Scholar] [PubMed]
- Asfari, M.; Janjic, D.; Meda, P.; Li, G.; Halban, P.A.; Wollheim, C.B. Establishment of 2-mercaptoethanol-dependent differentiated insulin-secreting cell lines. Endocrinology 1992, 130, 167–178. [Google Scholar] [CrossRef]
- Narushima, M.; Kobayashi, N.; Okitsu, T.; Tanaka, Y.; Li, S.A.; Chen, Y.; Miki, A.; Tanaka, K.; Nakaji, S.; Takei, K.; et al. A human beta-cell line for transplantation therapy to control type 1 diabetes. Nat. Biotechnol. 2005, 23, 1274–1282. [Google Scholar] [CrossRef]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 2013, 5, 178ra140. [Google Scholar] [CrossRef]
- Chaiworapongsa, T.; Chaemsaithong, P.; Korzeniewski, S.J.; Yeo, L.; Romero, R. Pre-eclampsia part 2: Prediction, prevention and management. Nat. Rev. Nephrol. 2014, 10, 531–540. [Google Scholar] [CrossRef]
- Houghton, A.M.; Rzymkiewicz, D.M.; Ji, H.; Gregory, A.D.; Egea, E.E.; Metz, H.E.; Stolz, D.B.; Land, S.R.; Marconcini, L.A.; Kliment, C.R.; et al. Neutrophil elastase-mediated degradation of IRS-1 accelerates lung tumor growth. Nat. Med. 2010, 16, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Borissoff, J.I.; Joosen, I.A.; Versteylen, M.O.; Brill, A.; Fuchs, T.A.; Savchenko, A.S.; Gallant, M.; Martinod, K.; Ten Cate, H.; Hofstra, L.; et al. Elevated levels of circulating DNA and chromatin are independently associated with severe coronary atherosclerosis and a prothrombotic state. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2032–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasek, R.C.; Gannon, M. Advancements and challenges in generating accurate animal models of gestational diabetes mellitus. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1327–E1338. [Google Scholar] [CrossRef] [PubMed]
- Feige, J.N.; Lagouge, M.; Auwerx, J. Dietary manipulation of mouse metabolism. Curr. Protoc. Mol. Biol. 2008, 84, 29B.5.1–29B.5.12. [Google Scholar] [CrossRef] [PubMed]
- Martínez, N.; Capobianco, E.; White, V.; Pustovrh, M.C.; Higa, R.; Jawerbaum, A. Peroxisome proliferator-activated receptor alpha activation regulates lipid metabolism in the feto-placental unit from diabetic rats. Reproduction 2008, 136, 95–103. [Google Scholar] [CrossRef]
- Damasceno, D.C.; Netto, A.O.; Iessi, I.L.; Gallego, F.Q.; Corvino, S.B.; Dallaqua, B.; Sinzato, Y.K.; Bueno, A.; Calderon, I.M.; Rudge, M.V. Streptozotocin-induced diabetes models: Pathophysiological mechanisms and fetal outcomes. Biomed. Res. Int. 2014, 2014, 819065. [Google Scholar] [CrossRef]
- Chang, A.S.; Dale, A.N.; Moley, K.H. Maternal diabetes adversely affects Preovulatory oocyte maturation, development, and granulosa cell apoptosis. Endocrinology 2005, 146, 2445–2453. [Google Scholar] [CrossRef]
- Yamashita, H.; Shao, J.; Ishizuka, T.; Klepcyk, P.J.; Muhlenkamp, P.; Qiao, L.; Hoggard, N.; Friedman, J.E. Leptin administration prevents spontaneous gestational diabetes in heterozygous Lepr(db/+) mice: Effects on placental leptin and fetal growth. Endocrinology 2001, 142, 2888–2897. [Google Scholar] [CrossRef]
- Nteeba, J.; Kubota, K.; Wang, W.; Zhu, H.; Vivian, J.; Dai, G.; Soares, M. Pancreatic prolactin receptor signaling regulates maternal glucose homeostasis. J. Endocrinol. 2019, 241, 71–83. [Google Scholar] [CrossRef]
- Baeyens, L.; Hindi, S.; Sorenson, R.L.; German, M.S. Beta-Cell adaptation in pregnancy. Diabetes Obes. Metab. 2016, 18, 63–70. [Google Scholar] [CrossRef]
- Ge, Z.J.; Liang, Q.X.; Hou, Y.; Han, Z.M.; Schatten, H.; Sun, Q.Y.; Zhang, C.L. Maternal obesity and diabetes may cause DNA methylation alteration in the spermatozoa of offspring in mice. Reprod. Biol. Endocrinol. 2014, 12, 29. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.; Eklund, E.; Bernabini, D.; Paladino, M.; Righetti, F.; Monti, G.; Lambert-Messerlian, G.A. A First-Trimester Biomarker Panel for Predicting the Development of Gestational Diabetes. Reprod. Sci. 2017, 24, 954–959. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| miRNA Isoform | Functional Role | References |
|---|---|---|
| miR-222 | The downregulated miR-222 and upregulated CXCR4 promotes insulin sensitivity and inhibits apoptosis in GDM. | Shi et al., 2014 [105] |
| miR-98 | Upregulation of miR-98 in the placental tissues of human GDM is linked to the global DNA methylation via targeting MECP2. | Cao et al., 2016 [106] |
| miR-518d | Increased expression of miR-518d is correlated with decreased protein expression of peroxisome proliferator-activated receptor-α (PPARα), a nuclear hormone receptor controlling glucose homeostasis. | Zhao et al., 2021 [100] |
| miR-340 | Insulin increased and glucose reduced miR-340 expression in GDM-affected women. | Stirm et al., 2018 [107] |
| miR-130b | This miR reflects the degree of obesity and serves as a potential biomarker for the diagnosis of GDM. | Wang et al., 2013 [108] |
| miR-148a | Regulates AMPKα1 activity (AMPK activity is significantly reduced in adipose tissue and the skeletal muscle of GDM-affected women). | Tryggestad et al., 2016 [109] |
| miR-144 | Upregulated in GDM-affected pregnant women. | Collares et al., 2013 [110]; Juchnicka et al., 2022 [111] |
| miR-517-5p | Expressed specifically in placenta; potential role in GDM pathogenesis. | Wander et al., 2017 [112] |
| miR-21-5p | Placental miR-21-5p expression inhibits cell growth and infiltration by upregulating PPAR-α. It is downregulated in GDM, thereby affecting the placental function. | Guan et al., 2020 [113] |
| miR-146b-5p | Has a pivotal role in pregnancy and pregnancy-related complications; associated only with GDM-affected patients carrying male foetuses | Zhu et al., 2018 [114] |
| miR-210-3p | Impairs pancreatic β-cell function by targeting Dtx1 in GDM | Cao et al., 2022 [115] |
| miR-222-3p | Upregulated expression of miR-222-3p is a potential circulating biomarker for the pathogenesis of GDM | Sadeghzadeh et al., 2020 [116] |
| Experimental Model | Species | Model Type | Experimental Features | References |
|---|---|---|---|---|
| Streptozotocin-Induced Diabetes Model. | Rat | In vivo | To study pathophysiology of GDM | Damasceno et al., 2014 [127] |
| C57BL/6 | Mouse | In vivo | Pregestational DM defined by non-fasting BG > 13.3 mmol/L | Chang et al., 2005 [128] |
| db/db | Mouse | In vivo | Obese, insulin resistant model with elevated glucose level; target gene is ObR gene. | Yamashita et al., 2001 [129] |
| PrlR± | Mouse | In vivo | Target gene is PrlR gene. Decreased β-cells, decreased tolerance to glucose; repressed insulin secretion; impaired glucose homeostasis. | Nteeba et al., 2019 [130] |
| HTR2b−/− | Mouse | In vivo | Target Gene is 5-HTR2b; decreased β-cell proliferation; high glucose intolerance. | Baeyens et al., 2016 [131] |
| CD1 | Mouse | In vivo | Pregestational DM defined by non-fasting BG > 17.0 mmol/L | Ge et al., 2014 [132] |
| Murine β-cell lines | Rat/Mouse | In vitro | Easy to culture; good option to test drugs and study cell physiology | Lilao-Garzón et al., 2021 [117] |
| Human pancreatic islets | Human | In vitro | Maintain the islet structure and all cell types; used to study the biology of the human pancreas. | Lilao-Garzón et al., 2021 [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.K.; Singh, S.; Singh, H.; Mahajan, D.; Kolli, P.; Mandadapu, G.; Kumar, B.; Kumar, D.; Kumar, S.; Jena, M.K. Deep Insight of the Pathophysiology of Gestational Diabetes Mellitus. Cells 2022, 11, 2672. https://doi.org/10.3390/cells11172672
Sharma AK, Singh S, Singh H, Mahajan D, Kolli P, Mandadapu G, Kumar B, Kumar D, Kumar S, Jena MK. Deep Insight of the Pathophysiology of Gestational Diabetes Mellitus. Cells. 2022; 11(17):2672. https://doi.org/10.3390/cells11172672
Chicago/Turabian StyleSharma, Amarish Kumar, Sanjeev Singh, Himanshu Singh, Deviyani Mahajan, Prachetha Kolli, Gowtham Mandadapu, Bimlesh Kumar, Dharmendra Kumar, Sudarshan Kumar, and Manoj Kumar Jena. 2022. "Deep Insight of the Pathophysiology of Gestational Diabetes Mellitus" Cells 11, no. 17: 2672. https://doi.org/10.3390/cells11172672