
The Impact of General Anesthesia on Redox Stability and Epigenetic Inflammation Pathways: Crosstalk on Perioperative Antioxidant Therapy


 ,
,  ,
,
Abstract
:1. Introduction
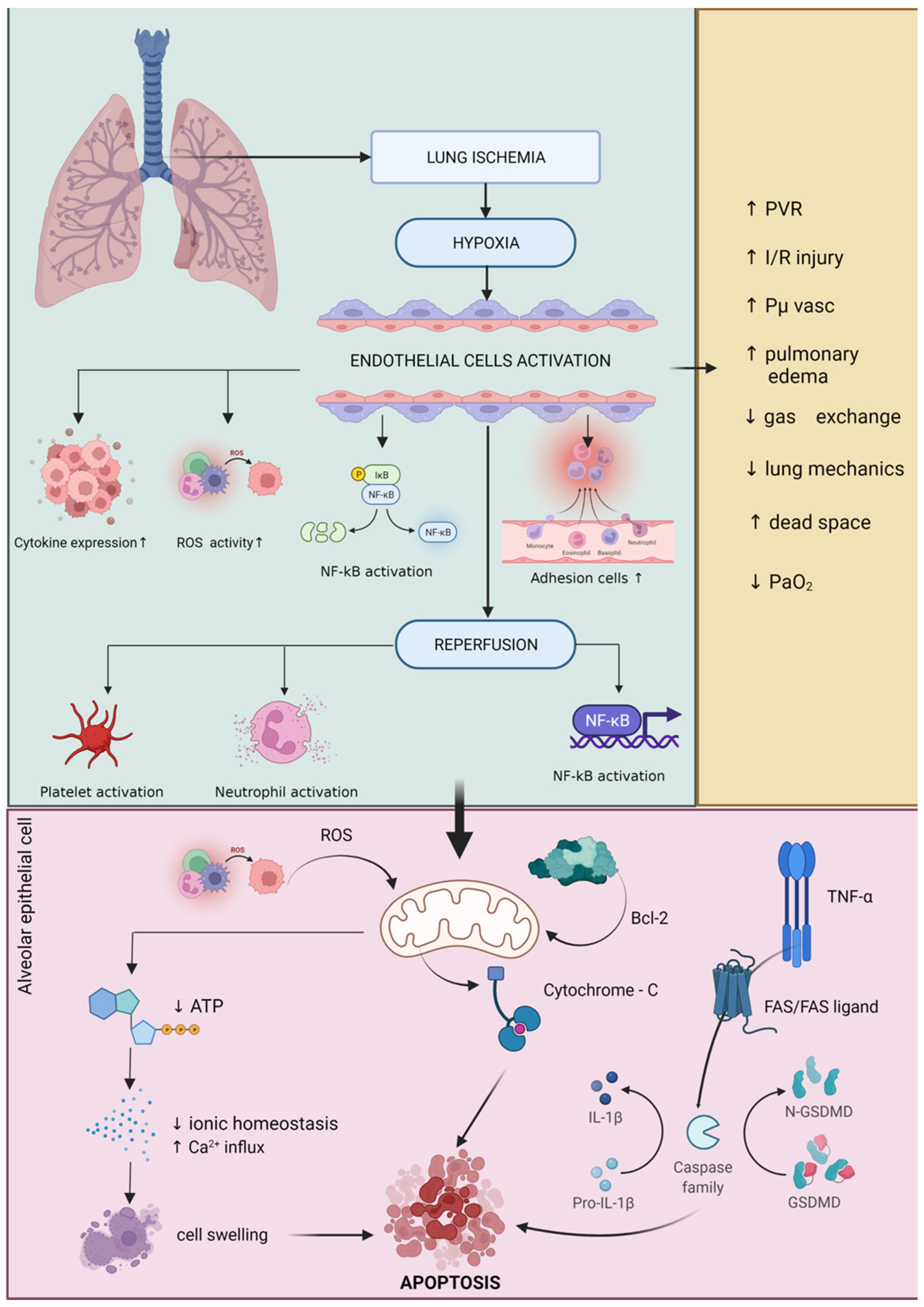
2. Redox Disturbance and Inflammation during General Anesthesia and Surgery Procedures
3. Implications of Genetic and Epigenetic Expression on Inflammation and Oxidative Pathways
4. Perioperative Antioxidant Therapy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kirov, K.; Motamed, C.; Ndoko, S.K.; Dhonneur, G. TOF Count at Corrugator Supercilii Reflects Abdominal Muscles Relaxation Better than at Adductor Pollicis. Br. J. Anaesth. 2007, 98, 611–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honing, M.; Martini, C.; van Velzen, M.; Niesters, M.; Dahan, A.; Boon, M. Cholinergic Chemotransmission and Anesthetic Drug Effects at the Carotid Bodies. Molecules 2020, 25, 5974. [Google Scholar] [CrossRef] [PubMed]
- Duţu, M.; Ivaşcu, R.; Tudorache, O.; Morlova, D.; Stanca, A.; Negoiţă, S.; Corneci, D. Neuromuscular monitoring: An update. Rom. J. Anaesth. Intensive Care 2018, 25, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funcke, S.; Saugel, B.; Koch, C.; Schulte, D.; Zajonz, T.; Sander, M.; Gratarola, A.; Ball, L.; Pelosi, P.; Spadaro, S.; et al. Individualized, Perioperative, Hemodynamic Goal-Directed Therapy in Major Abdominal Surgery (IPEGASUS Trial): Study Protocol for a Randomized Controlled Trial. Trials 2018, 19, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedreag, O.H.; Rogobete, A.F.; Sarandan, M.; Cradigati, A.C.; Papurica, M.; Dumbuleu, M.C.; Chira, A.M.; Rosu, O.M.; Sandesc, D. Oxidative Stress in Severe Pulmonary Trauma in Critical Ill Patients. Antioxidant Therapy in Patients with Multiple Trauma—A Review. Anestezjol. Intensywna Ter. 2015, 47, 351–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsing, C.H.; Wang, J.J. Clinical implication of perioperative inflammatory cytokine alteration. Acta Anaesthesiol. Taiwan 2015, 53, 23–28. [Google Scholar] [CrossRef]
- Boehm, O.; Baumgarten, G.; Hoeft, A. Epidemiology of the High-Risk Population: Perioperative Risk and Mortality after Surgery. Curr. Opin. Crit. Care 2015, 21, 322–327. [Google Scholar] [CrossRef]
- Riedel, B.; Browne, K.; Silbert, B. Cerebral Protection: Inflammation, Endothelial Dysfunction, and Postoperative Cognitive Dysfunction. Curr. Opin. Anaesthesiol. 2014, 27, 89–97. [Google Scholar] [CrossRef]
- Romagnoli, S.; Ricci, Z. Postoperative Acute Kidney Injury. Minerva Anestesiol. 2014, 81, 684–696. [Google Scholar]
- Snyder, G.L.; Greenberg, S. Effect of Anaesthetic Technique and Other Perioperative Factors on Cancer Recurrence. Br. J. Anaesth. 2010, 105, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Busti, A.J.; Hooper, J.S.; Amaya, C.J.; Kazi, S. Effects of Perioperative Antiinflammatory and Immunomodulating Therapy on Surgical Wound Healing. Pharmacotherapy 2005, 25, 1566–1591. [Google Scholar] [CrossRef] [PubMed]
- Papurica, M.; Rogobete, A.F.; Sandesc, D.; Dumache, R.; Nartita, R.; Sarandan, M.; Cradigati, A.C.; Luca, L.; Vernic, C.; Bedreag, O.H. Redox Changes Induced by General Anesthesia in Critically Ill Patients with Multiple Traumas. Mol. Biol. Int. 2015, 2015, 238586. [Google Scholar] [CrossRef] [PubMed]
- Denk, S.; Perl, M.; Huber-Lang, M. Damage- and Pathogen-Associated Molecular Patterns and Alarmins: Keys to Sepsis? Eur. Surg. Res. 2012, 48, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Oppenheim, J.J.; Yang, D. Alarmins: Chemotactic Activators of Immune Responses. Curr. Opin. Immunol. 2005, 17, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.K.; Roth, J.; Oppenheim, J.J.; Tracey, K.J.; Vogl, T.; Feldmann, M.; Horwood, N.; Nanchahal, J. Alarmins: Awaiting a Clinical Response. J. Clin. Invest. 2012, 122, 2711–2719. [Google Scholar] [CrossRef]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of Chromatin Protein HMGB1 by Necrotic Cells Triggers Inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef]
- Tbahriti, H.F.; Meknassi, D.; Moussaoui, R.; Messaoudi, A.; Zemour, L.; Kaddous, A.; Bouchenak, M.; Mekki, K. Inflammatory Status in Chronic Renal Failure: The Role of Homocysteinemia and Pro-Inflammatory Cytokines. World J. Nephrol. 2013, 2, 31–37. [Google Scholar] [CrossRef]
- Little, J.P.; Simtchouk, S.; Schindler, S.M.; Villanueva, E.B.; Gill, N.E.; Walker, D.G.; Wolthers, K.R.; Klegeris, A. Mitochondrial Transcription Factor A (Tfam) Is a pro-Inflammatory Extracellular Signaling Molecule Recognized by Brain Microglia. Mol. Cell. Neurosci. 2014, 60, 88–96. [Google Scholar] [CrossRef]
- Kirchhoff, C.; Biberthaler, P.; Mutschler, W.E.; Faist, E.; Jochum, M.; Zedler, S. Early Down-Regulation of the pro-Inflammatory Potential of Monocytes Is Correlated to Organ Dysfunction in Patients after Severe Multiple Injury: A Cohort Study. Crit. Care 2009, 13, R88. [Google Scholar] [CrossRef] [Green Version]
- Phillipson, M.; Kubes, P. The Neutrophil in Vascular Inflammation. Nat. Med. 2011, 17, 1381–1390. [Google Scholar] [CrossRef]
- Reis, G.S.; Augusto, V.S.; Silveira, A.P.C.; Jordão, A.A.; Baddini-Martinez, J.; Poli Neto, O.; Rodrigues, A.J.; Evora, P.R.B. Oxidative-Stress Biomarkers in Patients with Pulmonary Hypertension. Pulmonary 2013, 3, 856–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafner, S.; Radermacher, P.; Frick, M.; Dietl, P.; Calzia, E. Hyperglycemia, Oxidative Stress, and the Diaphragm: A Link between Chronic Co-Morbidity and Acute Stress? Crit. Care 2014, 18, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cellular, O.S.; Overload, C. Sevoflurane Protects Ventricular Myocytes against Oxidative Stress-Induced Cellular Ca2+ Overload and Hypercontracture. Anesthesiology 2013, 119, 606–620. [Google Scholar]
- Breitenbach, M.; Rinnerthaler, M.; Weber, M.; Breitenbach-Koller, H.; Karl, T.; Cullen, P.; Basu, S.; Haskova, D.; Hasek, J. The Defense and Signaling Role of NADPH Oxidases in Eukaryotic Cells: Review. Wien. Med. Wochenschr. 2018, 168, 286–299. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Wang, M.; Wang, Y.; Huang, A. Superoxide anion generation response to wound in Arabidopsis hypocotyl cutting. Plant Signal Behav. 2021, 16, 1848086. [Google Scholar] [CrossRef]
- Constantino, L.; Gonçalves, R.C.; Giombelli, V.R.; Tomasi, C.D.; Vuolo, F.; Kist, L.W.; Medeiros, G.; de Oliveira, T.; Augusto, M.; Pasquali, D.B.; et al. Regulation of Lung Oxidative Damage by Endogenous Superoxide Dismutase in Sepsis. Intensive Care Med. Exp. 2014, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Milkovic, L.; Gasparovic, A.C.; Cindric, M.; Mouthuy, P.A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef] [Green Version]
- Lv, P.; Xue, P.; Dong, J.; Peng, H.; Clewell, R.; Wang, A.; Wang, Y.; Peng, S.; Qu, W.; Zhang, Q.; et al. Keap1 Silencing Boosts Lipopolysaccharide-Induced Transcription of Interleukin 6 via Activation of Nuclear Factor ΚB in Macrophages. Toxicol. Appl. Pharmacol. 2013, 272, 697–702. [Google Scholar] [CrossRef]
- Pagano, G.; Talamanca, A.A.; Castello, G.; Cordero, M.D.; Ischia, M.; Gadaleta, M.N.; Pallardó, F.V.; Petrović, S.; Tiano, L.; Zatterale, A. Oxidative Stress and Mitochondrial Dysfunction across Broad-Ranging Pathologies: Toward Mitochondria-Targeted Clinical Strategies. Oxidative Med. Cell. Longev. 2014, 2014, 541230. [Google Scholar] [CrossRef]
- Waldbaum, S.; Patel, M. Mitochondria, Oxidative Stress, and Temporal Lobe Epilepsy. Epilepsy Res. 2010, 88, 23–45. [Google Scholar] [CrossRef] [Green Version]
- Zang, Q.S.; Martinez, B.; Yao, X.; Maass, D.L.; Ma, L.; Wolf, S.E.; Minei, J.P. Sepsis-Induced Cardiac Mitochondrial Dysfunction Involves Altered Mitochondrial-Localization of Tyrosine Kinase Src and Tyrosine Phosphatase SHP2. PLoS ONE 2012, 7, e43424. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.M.; Williams, J.A.; Ding, W.X. Mitochondrial Dynamics and Mitochondrial Quality Control. Redox Biol. 2015, 4, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, A.V.; Bahrami, S.; Calzia, E.; Dungel, P.; Gille, L.; Kuznetsov, A.V.; Troppmair, J. Mitochondrial Dysfunction and Biogenesis: Do ICU Patients Die from Mitochondrial Failure? Ann. Intensive Care 2011, 1, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalogeris, T.; Bao, Y.; Korthuis, R.J. Mitochondrial Reactive Oxygen Species: A Double Edged Sword in Ischemia/Reperfusion vs Preconditioning. Redox Biol. 2014, 2, 702–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Hu, F.; Wu, J.; Zhang, S. Cannabidiol Attenuates OGD/R-Induced Damage by Enhancing Mitochondrial Bioenergetics and Modulating Glucose Metabolism via Pentose-Phosphate Pathway in Hippocampal Neurons. Redox Biol. 2017, 11, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Gorelenkova Miller, O.; Behring, J.B.; Siedlak, S.L.; Jiang, S.; Matsui, R.; Bachschmid, M.M.; Zhu, X.; Mieyal, J.J. Upregulation of Glutaredoxin-1 Activates Microglia and Promotes Neurodegeneration: Implications for Parkinson’s Disease. Antioxid. Redox Signal. 2016, 25, 967–982. [Google Scholar] [CrossRef] [Green Version]
- Porfire, A.S.; Leucuţa, S.E.; Kiss, B.; Loghin, F.; Pârvu, A.E. Investigation into the Role of Cu/Zn-SOD Delivery System on Its Antioxidant and Antiinflammatory Activity in Rat Model of Peritonitis. Pharmacol. Rep. 2014, 66, 670–676. [Google Scholar] [CrossRef]
- Edem, V.F.; Kosoko, A.; Akinyoola, S.B.; Owoeye, O.; Rahamon, S.K.; Arinola, O.G. Plasma Antioxidant Enzymes, Lipid Peroxidation and Hydrogen Peroxide in Wistar Rats Exposed to Dichlorvos Insecticide. Sch. Res. Libr. Arch. Appl. Sci. Res. 2012, 4, 1778–1781. [Google Scholar]
- Risnes, S.F.; Hartwig, A. Impact of Cadmium on Antioxidant Enzymes in HCT116 Cells and Protective Interaction by Selenium. Perspect. Sci. 2015, 3, 55. [Google Scholar] [CrossRef] [Green Version]
- Samarghandian, S.; Afshari, R.; Farkhondeh, T. Effect of Long-Term Treatment of Morphine on Enzymes, Oxidative Stress Indices and Antioxidant Status in Male Rat Liver. Int. J. Clin. Exp. Med. 2014, 7, 1449–1453. [Google Scholar]
- Lee, B.-J.; Lin, J.-S.; Lin, Y.-C.; Lin, P.-T. Effects of L-Carnitine Supplementation on Oxidative Stress and Antioxidant Enzymes Activities in Patients with Coronary Artery Disease: A Randomized, Placebo-Controlled Trial. Nutr. J. 2014, 13, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, B.; Vicenzi, M.; Garrel, C.; Denis, F.M. Redox Biology Effects of N-Acetylcysteine, Oral Glutathione (GSH) and a Novel Sublingual Form of GSH on Oxidative Stress Markers: A Comparative Crossover Study. Redox Biol. 2015, 6, 198–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Babu, D.; Leclercq, G.; Goossens, V.; Vanden, T.; van Hamme, E.; Vandenabeele, P.; Lefebvre, R.A. Mitochondria and NADPH Oxidases Are the Major Sources of TNF-α/Cycloheximide-Induced Oxidative Stress in Murine Intestinal Epithelial MODE-K Cells. Cell. Signal. 2015, 27, 1141–1158. [Google Scholar] [CrossRef]
- Jaganjac, M.; Milkovic, L.; Zarkovic, N.; Zarkovic, K. Oxidative Stress and Regeneration. Free Radic. Biol. Med. 2022, 181, 154–165. [Google Scholar] [CrossRef]
- Li, P.; Stetler, R.A.; Leak, R.K.; Shi, Y.; Li, Y.; Yu, W.; Bennett, M.V.L.; Chen, J. Oxidative Stress and DNA Damage after Cerebral Ischemia: Potential Therapeutic Targets to Repair the Genome and Improve Stroke Recovery. Neuropharmacology 2018, 134, 208–217. [Google Scholar] [CrossRef]
- Carcy, R.; Cougnon, M.; Poet, M.; Durandy, M.; Sicard, A.; Counillon, L.; Blondeau, N.; Hauet, T.; Tauc, M.; Pisani, D.F. Targeting Oxidative Stress, a Crucial Challenge in Renal Transplantation Outcome. Free Radic. Biol. Med. 2021, 169, 258–270. [Google Scholar] [CrossRef]
- Yuan, Q.; Yuan, Y.; Zheng, Y.; Sheng, R.; Liu, L.; Xie, F.; Tan, J. Anti-Cerebral Ischemia Reperfusion Injury of Polysaccharides: A Review of the Mechanisms. Biomed. Pharmacother. 2021, 137, 111303. [Google Scholar] [CrossRef]
- Padmavathi, G.; Ramkumar, K.M. MicroRNA Mediated Regulation of the Major Redox Homeostasis Switch, Nrf2, and Its Impact on Oxidative Stress-Induced Ischemic/Reperfusion Injury. Arch. Biochem. Biophys. 2021, 698, 108725. [Google Scholar] [CrossRef]
- Hu, Y.; Deng, H.; Xu, S.; Zhang, J. MicroRNAs Regulate Mitochondrial Function in Cerebral Ischemia-Reperfusion Injury. Int. J. Mol. Sci. 2015, 16, 24895–24917. [Google Scholar] [CrossRef] [Green Version]
- Chi, O.Z.; Barsoum, S.; Rah, K.H.; Liu, X.; Weiss, H.R. Local O2 Balance in Cerebral Ischemia-Reperfusion Improved during Pentobarbital Compared with Isoflurane Anesthesia. J. Stroke Cerebrovasc. Dis. 2015, 24, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Zhai, F.; Zhang, X.; Guan, Y.; Yang, X.; Li, Y.; Song, G.; Guan, L. Expression Profiles of MicroRNAs after Focal Cerebral Ischemia/Reperfusion Injury in Rats. Neural Regen. Res. 2012, 7, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.R.; Keller, J.N. Oxidative Stress and Cerebral Endothelial Cells: Regulation of the Blood-Brain-Barrier and Antioxidant Based Interventions. Biochim. Biophys. Acta-Mol. Basis Dis. 2012, 1822, 822–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yan, J.; Shi, H. Neurobiology of Disease Role of Hypoxia Inducible Factor 1 in Hyperglycemia-Exacerbated Blood-Brain Barrier Disruption in Ischemic Stroke. Neurobiol. Dis. 2016, 95, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Cheng, Z.; Cheng, Y.; Xu, Y. Investigating the Passage of Tetramethylpyrazine-Loaded Liposomes across Blood-Brain Barrier Models in Vitro and Ex Vivo. Mater. Sci. Eng. C 2016, 69, 1010–1017. [Google Scholar] [CrossRef]
- Robertson, C.S.; Narayan, R.K.; Gokaslan, Z.L.; Pahwa, R.; Grossman, R.G.; Caram, P.J.; Allen, E. Cerebral Arteriovenous Oxygen Difference as an Estimate of Cerebral Blood Flow in Comatose Patients. J. Neurosurg. 1989, 70, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Bouzat, P.; Sala, N.; Payen, J.-F.; Oddo, M. Beyond Intracranial Pressure: Optimization of Cerebral Blood Flow, Oxygen, and Substrate Delivery after Traumatic Brain Injury. Ann. Intensive Care 2013, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Weiss, H.R.; Grayson, J.; Liu, X.; Barsoum, S.; Shah, H.; Chi, O.Z. Cerebral Ischemia and Reperfusion Increases the Heterogeneity of Local Oxygen Supply/Consumption Balance. Stroke 2013, 44, 2553–2558. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Sheng, H.; Yates, R.B.; Ishida, K.; Pearlstein, R.D.; Warner, D.S. Isoflurane Provides Long-Term Protection against Focal Cerebral Ischemia in the Rat. Anesthesiology 2007, 106, 92–99. [Google Scholar] [CrossRef]
- Gambim, M.; de Oliveira do Carmo, A.; Marti, L.; Veríssimo-Filho, S.; Lopes, L.; Janiszewski, M. Platelet-Derived Exosomes Induce Endothelial Cell Apoptosis through Peroxynitrite Generation: Experimental Evidence for a Novel Mechanism of Septic Vascular Dysfunction. Crit. Care 2007, 11, R107. [Google Scholar] [CrossRef] [Green Version]
- Carnes, C.A.; Chung, M.K.; Nakayama, T.; Nakayama, H.; Baliga, R.S.; Piao, S.; Kanderian, A.; Pavia, S.; Hamlin, R.L.; McCarthy, P.M.; et al. Ascorbate Attenuates Atrial Pacing-Induced Peroxynitrite Formation and Electrical Remodeling and Decreases the Incidence of Postoperative Atrial Fibrillation. Circ. Res. 2001, 89, e32–e38. [Google Scholar] [CrossRef] [PubMed]
- Erecinska, M.; Thoresen, M.; Silver, I.A. Effects of Hypothermia on Energy Metabolism in Mammalian Central Nervous System. J. Cereb. Blood Flow Metab. 2003, 23, 513–530. [Google Scholar] [CrossRef] [PubMed]
- Scheufler, K.M.; Lehnert, A.; Rohrborn, H.J.; Nadstawek, J.; Thees, C. Individual Value of Brain Tissue Oxygen Pressure, Microvascular Oxygen Saturation, Cytochrome Redox Level, and Energy Metabolites in Detecting Critically Reduced Cerebral Energy State during Acute Changes in Global Cerebral Perfusion. J. Neurosurg. Anesthesiol. 2004, 16, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Antoniades, C.; Tousoulis, D.; Vasiliadou, C.; Pitsavos, C.; Toutouza, M.; Tentolouris, C.; Marinou, K.; Stefanadis, C. Genetic Polymorphisms G894T on the ENOS Gene Is Associated with Endothelial Function and VWF Levels in Premature Myocardial Infarction Survivors. Int. J. Cardiol. 2006, 107, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Abd-elbaset, M.; Arafa, E.A. Quercetin Modulates INOS, ENOS and NOSTRIN Expressions and Attenuates Oxidative Stress in Warm Hepatic Ischemia-Reperfusion Injury in Rats. Beni-Suef Univ. J. Basic Appl. Sci. 2015, 4, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Silver, J.H.; Jaffe, R.A.; López, J.R. Plasma Nitrite as an Indicator of Cerebral Ischemia during Extracranial/Intracranial Bypass Surgery in Moyamoya Patients. J. Stroke Cerebrovasc. Dis. 2020, 29, 104830. [Google Scholar] [CrossRef]
- Suzuki, H.; Kanamaru, H.; Kawakita, F.; Asada, R.; Fujimoto, M.; Shiba, M. Cerebrovascular pathophysiology of delayed cerebral ischemia after aneurysmal subarachnoid hemorrhage. Histol. Histopathol. 2021, 36, 143–158. [Google Scholar] [CrossRef]
- Washington, C.W.; Zipfel, G.J. Detection and Monitoring of Vasospasm and Delayed Cerebral Ischemia: A Review and Assessment of the Literature. Neurocrit. Care 2011, 15, 312–317. [Google Scholar] [CrossRef]
- Liu, T.-J.; Zhang, J.-C.; Gao, X.-Z.; Tan, Z.-B.; Wang, J.-J.; Zhang, P.-P.; Cheng, A.-B.; Zhang, S.-B. Effect of Sevoflurane on the ATPase Activity of Hippocampal Neurons in a Rat Model of Cerebral Ischemia-Reperfusion Injury via the CAMP-PKA Signaling Pathway. Kaohsiung J. Med. Sci. 2018, 34, 22–33. [Google Scholar] [CrossRef]
- Coles, J.P.; Fryer, T.D.; Smielewski, P.; Chatfield, D.A.; Steiner, L.A.; Johnston, A.J.; Downey, S.P.; Williams, G.B.; Aigbirhio, F.; Hutchinson, P.J.; et al. Incidence and Mechanisms of Cerebral Ischemia in Early Clinical Head Injury. J. Cereb. Blood Flow Metab. 2004, 24, 202–211. [Google Scholar] [CrossRef]
- Drake, C.; Boutin, H.; Jones, M.S.; Denes, A.; Mccoll, B.W.; Selvarajah, J.R.; Hulme, S.; Georgiou, R.F.; Hinz, R.; Gerhard, A.; et al. Brain, Behavior, and Immunity Brain Inflammation Is Induced by Co-Morbidities and Risk Factors for Stroke. Brain Behav. Immun. 2011, 25, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Hartley, O.; Offord, R.E. Engineering Chemokines to Develop Optimized HIV Inhibitors. Curr. Protein Pept. Sci. 2005, 6, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Stamatovic, S.M.; Phillips, C.M.; Martinez-Revollar, G.; Keep, R.F.; Andjelkovic, A.V. Involvement of Epigenetic Mechanisms and Non-Coding RNAs in Blood-Brain Barrier and Neurovascular Unit Injury and Recovery after Stroke. Front. Neurosci. 2019, 13, 864. [Google Scholar] [CrossRef] [PubMed]
- De Perrot, M.; Liu, M.; Waddell, T.K.; Keshavjee, S. Ischemia-Reperfusion-Induced Lung Injury. Am. J. Respir. Crit. Care Med. 2003, 167, 490–511. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, Z.; Du, L.; Huang, Y.; Ge, J.; Deng, Y.; Mei, Z. The Potential Role of MicroRNA-124 in Cerebral Ischemia Injury. Int. J. Mol. Sci. 2020, 21, 120. [Google Scholar] [CrossRef] [Green Version]
- Stoica, L.; Dobrescu, A.; Isaic, A.; Verdeæ, G.; Taråa, C.; Lazãr, F. Metabolic and Hormonal Changes after Sleeve Gastrectomy and Mini Gastric Bypass in a Rat Model of Induced Type 2 Diabetes Mellitus and Obesity. Chirurgia 2019, 114, 732–738. [Google Scholar] [CrossRef]
- Rogobete, A.F.; Grintescu, I.M.; Bratu, T.; Bedreag, O.H.; Papurica, M.; Crainiceanu, Z.P.; Popovici, S.E.; Sandesc, D. Assessment of Metabolic and Nutritional Imbalance in Mechanically Ventilated Multiple Trauma Patients: From Molecular to Clinical Outcomes. Diagnostics 2019, 9, 171. [Google Scholar] [CrossRef] [Green Version]
- Eppinger, M.J.; Deeb, G.M.; Bolling, S.F.; Ward, P.A. Mediators of ischemia-reperfusion injury of rat lung. Am. J. Pathol. 1997, 150, 1773–1784. [Google Scholar]
- Qayumi, A.K.; Jamieson, W.R.E.; Godin, D.V.; Lam, S.; Ko, K.M.; Germann, E.; van den Broek, J. Response to Allopurinol Pretreatment in a Swine Model of Heart-Lung Transplantation. J. Investig. Surg. 1990, 3, 331–340. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, J.; Li, L.; Li, H.; Mao, S.; Zhang, F.; Zen, K.; Zhang, C.; Zhang, Q. MicroRNA-23a/b and MicroRNA-27a/b Suppress Apaf-1 Protein and Alleviate Hypoxia-Induced Neuronal Apoptosis. Cell Death Dis. 2014, 5, e1132. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell Death: Critical Control Points. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Borutaite, V.; Jekabsone, A.; Morkuniene, R.; Brown, G.C. Inhibition of Mitochondrial Permeability Transition Prevents Mitochondrial Dysfunction, Cytochrome c Release and Apoptosis Induced by Heart Ischemia. J. Mol. Cell. Cardiol. 2003, 35, 357–366. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial Formation of Reactive Oxygen Species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Cobelens, P.M.; van Putte, B.P.; Kavelaars, A.; Heijnen, C.J.; Kesecioglu, J. Inflammatory Consequences of Lung Ischemia-Reperfusion Injury and Low-Pressure Ventilation. J. Surg. Res. 2008, 153, 295–301. [Google Scholar] [CrossRef]
- Hickey, M.J.; Sharkey, K.A.; Sihota, E.G.; Reinhardt, P.H.; Macmicking, J.D.; Nathan, C.; Kubes, P. Inducible Nitric Oxide Synthase-Deficient Mice Have Enhanced Leukocyte-Endothelium Interactions in Endotoxemia. FASEB J. 1997, 11, 955–964. [Google Scholar] [CrossRef]
- Ovechkin, A.V.; Lominadze, D.; Sedoris, K.C.; Robinson, T.W.; Tyagi, S.C.; Roberts, A.M. Lung Ischemia-Reperfusion Injury: Implications of Oxidative Stress and Platelet-Arteriolar Wall Interactions. Arch. Physiol. Biochem. 2007, 113, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Saelens, X.; Festjens, N.; van de Walle, L.; van Gurp, M.; van Loo, G.; Vandenabeele, P. Toxic Proteins Released from Mitochondria in Cell Death. Oncogene 2004, 23, 2861–2874. [Google Scholar] [CrossRef] [Green Version]
- Scarabelli, T.M.; Stephanou, A.; Pasini, E.; Comini, L.; Raddino, R.; Knight, R.A.; Latchman, D.S. Different Signaling Pathways Induce Apoptosis in Endothelial Cells and Cardiac Myocytes during Ischemia/Reperfusion Injury. Circ. Res. 2002, 90, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Lemasters, J.J. Free Radical Biology and Medicine Translocation of Iron from Lysosomes to Mitochondria during Ischemia Predisposes to Injury after Reperfusion in Rat Hepatocytes. Free Radic. Biol. Med. 2013, 63, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Mikawa, K.; Akamatsu, H.; Nishina, K.; Shiga, M.; Maekawa, N.; Obara, H.; Niwa, Y. Propofol Inhibits Human Neutrophil Functions. Anesth. Analg. 1998, 87, 695–700. [Google Scholar] [CrossRef]
- Ohmizo, H.; Iwama, H.; Sugita, T. Complement Activation by Propofol and Its Effect during Propofol Anaesthesia. Anaesth. Intensive Care 1999, 27, 623–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoff, G.; Bauer, I.; Larsen, B.; Bauer, M. Modulation of Endotoxin-Stimulated TNF-Alpha Gene Expression by Ketamine and Propofol in Cultured Human Whole Blood. Anaesthesist 2001, 50, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhata, H.; Shimizu, R.; Yokoyama, M.M. Suppressive Effects of Volatile Anesthetics on Cytokine Release in Human Peripheral Blood Mononuclear Cells. Int. J. Immunopharmacol. 1995, 17, 529–534. [Google Scholar] [CrossRef]
- Giraud, O.; Molliex, S.; Rolland, C.; Leçon-Malas, V.; Desmonts, J.-M.; Aubier, M.; Dehoux, M. Halogenated Anesthetics Reduce Interleukin-1beta-Induced Cytokine Secretion by Rat Alveolar Type II Cells in Primary Culture. Anesthesiology 2003, 98, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Kotani, N.; Takahashi, S.; Sessler, D.I.; Hashiba, E.; Kubota, T.; Hashimoto, H.; Matsuki, A. Volatile Anesthetics Augment Expression of Proinflammatory Cytokines in Rat Alveolar Macrophages during Mechanical Ventilation. Anesthesiology 1999, 91, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, J.; Yang, L.; Dong, Y.; Zhang, Y.; Xie, Z. Isoflurane and Sevoflurane Increase Interleukin-6 Levels through the Nuclear Factor-Kappa B Pathway in Neuroglioma Cells. Br. J. Anaesth. 2013, 110 (Suppl. S1), i82–i91. [Google Scholar] [CrossRef] [Green Version]
- Hudetz, J.A.; Gandhi, S.D.; Iqbal, Z.; Patterson, K.M.; Pagel, P.S. Elevated Postoperative Inflammatory Biomarkers Are Associated with Short- and Medium-Term Cognitive Dysfunction after Coronary Artery Surgery. J. Anesth. 2011, 25, 1–9. [Google Scholar] [CrossRef]
- Manfreda, S.E.; Dunzendorfer, S.; Schratzberger, P.; Buratti, T.; Reinisch, N.; Kähler, C.M.; List, W.F.; Wiedermann, C.J. The Chemotaxis of Human Peripheral Blood B Lymphocytes by Beta-Endorphin Is Reversible by Naloxone. Anesth. Analg. 1998, 86, 670–672. [Google Scholar] [CrossRef]
- Yeager, M.P.; Yu, C.T.; Campbell, A.S.; Moschella, M.; Guyre, P.M. Effect of Morphine and Beta-Endorphin on Human Fc Receptor-Dependent and Natural Killer Cell Functions. Clin. Immunol. Immunopathol. 1992, 62, 336–343. [Google Scholar] [CrossRef]
- Baek, S.B.; Shin, M.S.; Han, J.H.; Moon, S.W.; Chang, B.; Jeon, J.W.; Yi, J.W.; Chung, J.Y. Rocuronium Bromide Inhibits Inflammation and Pain by Suppressing Nitric Oxide Production and Enhancing Prostaglandin E(2) Synthesis in Endothelial Cells. Int. Neurourol. J. 2016, 20, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Bargellini, A.; Rovesti, S.; Barbieri, A.; Vivoli, R.; Roncaglia, R.; Righi, E.; Borella, P. Effects of Chronic Exposure to Anaesthetic Gases on Some Immune Parameters. Sci. Total Environ. 2001, 270, 149–156. [Google Scholar] [CrossRef]
- Yamamoto, S.; Niida, S.; Azuma, E.; Yanagibashi, T.; Muramatsu, M.; Huang, T.T.; Sagara, H.; Higaki, S.; Ikutani, M.; Nagai, Y.; et al. Inflammation-Induced Endothelial Cell-Derived Extracellular Vesicles Modulate the Cellular Status of Pericytes. Sci. Rep. 2015, 5, 8505. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-ΚB, Inflammation, and Metabolic Disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xie, B.; Chen, S.; Jiang, F.; Meng, W. Association Study of Two Inflammation-Related Polymorphisms with Susceptibility to Hepatocellular Carcinoma: A Meta-Analysis. BMC Med. Genet. 2014, 15, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papurica, M.; Sandesc, D.; Rogobete, A.F.; Nartita, R.; Vernic, C.; Popovici, S.E.; Bedreag, O.H. Cardioprotective Effects Induced by Preconditioning with Halogenated Anesthetics. J. Interdiscip. Med. 2016, 1, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Tretter, V.; Hochreiter, B.; Zach, M.L.; Krenn, K.; Klein, K.U. Understanding Cellular Redox Homeostasis: A Challenge for Precision Medicine. Int. J. Mol. Sci. 2022, 23, 106. [Google Scholar] [CrossRef]
- Bedreag, O.H.; Rogobete, A.F.; Cradigati, C.A.; Sarandan, M.; Nartita, R.; Horhat, F.G.; Popovici, S.E.; Sandesc, D.; Papurica, M. A Novel Evaluation of Microvascular Damage in Critically Ill Polytrauma Patients by Using Circulating MicroRNAs. Rev. Română Med. Lab. 2016, 24, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, T.B.; Engel, A.; Sakamoto, H.; O’Shea, A.; O’Callaghan-Enright, S.; Shorten, G.D. The Effects of Propofol on Neutrophil Function, Lipid Peroxidation and Inflammatory Response during Elective Coronary Artery Bypass Grafting in Patients with Impaired Ventricular Function. Br. J. Anaesth. 2006, 97, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Potočnik, I.; Janković, V.N.; Šostarič, M.; Jerin, A.; Štupnik, T.; Skitek, M.; Markovič-Božič, J.; Klokočovnik, T. Antiinflammatory Effect of Sevoflurane in Open Lung Surgery with One-Lung Ventilation. Croat. Med. J. 2014, 55, 628–637. [Google Scholar] [CrossRef] [Green Version]
- Markovic-Bozic, J.; Karpe, B.; Potocnik, I.; Jerin, A.; Vranic, A.; Novak-Jankovic, V. Effect of Propofol and Sevoflurane on the Inflammatory Response of Patients Undergoing Craniotomy. BMC Anesthesiol. 2016, 16, 18. [Google Scholar] [CrossRef] [Green Version]
- Roh, G.U.; Song, Y.; Park, J.; Ki, Y.M.; Han, D.W. Effects of Propofol on the Inflammatory Response during Robot-Assisted Laparoscopic Radical Prostatectomy: A Prospective Randomized Controlled Study. Sci. Rep. 2019, 9, 5242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijeysundera, D.N.; Committee, E.R.; Duncan, D.; Nkonde-price, C.; Virani, S.S.; Washam, J.B.; Fleischmann, K.E.; Vice, P.G.; Fleisher, L.A.; Guideline, P.; et al. Perioperative Beta Blockade in Noncardiac Surgery: A Systematic Review for the 2014 ACC/AHA Guideline on Perioperative Cardiovascular Evaluation and Management of Patients Undergoing Noncardiac Surgery. Circulation 2014, 64, 2246–2264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julier, K.; da Silva, R.; Garcia, C.; Bestmann, L.; Frascarolo, P.; Zollinger, A.; Chassot, P.-G.; Schmid, E.R.; Turina, M.I.; von Segesser, L.K.; et al. Preconditioning by Sevoflurane Decreases Biochemical Markers for Myocardial and Renal Dysfunction in Coronary Artery Bypass Graft Surgery: A Double-Blinded, Placebo-Controlled, Multicenter Study. Anesthesiology 2003, 98, 1315–1327. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, K.; Lee, H.T. Volatile Anesthetics and AKI: Risks, Mechanisms, and a Potential Therapeutic Window. J. Am. Soc. Nephrol. 2014, 25, 884–892. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Xu, R.; Yu, X.; Fang, Y.; Ding, X. Volatile Anesthetics in Preventing Acute Kidney Injury after Cardiac Surgery: A Systematic Review and Meta-Analysis. J. Thorac. Cardiovasc. Surg. 2014, 148, 3127–3136. [Google Scholar] [CrossRef] [Green Version]
- Kempson, S.A.; Zhou, Y.; Danbolt, N.C. The Betaine/GABA Transporter and Betaine: Roles in Brain, Kidney, and Liver. Front. Physiol. 2014, 5, 159. [Google Scholar] [CrossRef] [Green Version]
- Bagley, E.E. Opioid and GABAB Receptors Differentially Couple to an Adenylyl Cyclase/Protein Kinase a Downstream Effector after Chronic Morphine Treatment. Front. Pharmacol. 2014, 5, 148. [Google Scholar] [CrossRef] [Green Version]
- Adermark, L.; Söderpalm, B.; Burkhardt, J.M. Brain Region Specific Modulation of Ethanol-Induced Depression of GABAergic Neurons in the Brain Reward System by the Nicotine Receptor Antagonist Mecamylamine. Alcohol 2014, 48, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Wagner, J.; Strosing, K.M.; Spassov, S.G.; Lin, Z.; Engelstaedter, H.; Tacke, S.; Hoetzel, A.; Faller, S. Sevoflurane Posttreatment Prevents Oxidative and Inflammatory Injury in Ventilator-Induced Lung Injury. PLoS ONE 2018, 13, e0192896. [Google Scholar] [CrossRef] [Green Version]
- Kalimeris, K.; Kouni, S.; Kostopanagiotou, G.; Nomikos, T.; Fragopoulou, E.; Kakisis, J.; Vasdekis, S.; Matsota, P.; Pandazi, A. Cognitive Function and Oxidative Stress after Carotid Endarterectomy: Comparison of Propofol to Sevoflurane Anesthesia. J. Cardiothorac. Vasc. Anesth. 2013, 27, 1246–1252. [Google Scholar] [CrossRef]
- Yang, P.; Du, Y.; Zeng, H.; Xing, H.; Tian, C.; Zou, X. Comparison of Inflammatory Markers between the Sevoflurane and Isoflurane Anesthesia in a Rat Model of Liver Ischemia/Reperfusion Injury. Transplant. Proc. 2019, 51, 2071–2075. [Google Scholar] [CrossRef] [PubMed]
- Gyires, K.; Budavñri, I.; Fürst, S.; Molnñr, I. Morphine Inhibits the Carrageenan-Induced Oedema and the Chemoluminescence of Leucocytes Stimulated by Zymosan. J. Pharm. Pharmacol. 1985, 37, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Sacerdote, P.; Bianchi, M.; Panerai, A.E. Involvement of β-Endorphin in the Modulation of Paw Inflammatory Edema in the Rat. Regul. Pept. 1996, 63, 79–83. [Google Scholar] [CrossRef]
- Planas, E.; Sánchez, S.; Rodriguez, L.; Pol, O.; Puig, M.M. Antinociceptive/Anti-Edema Effects of Liposomal Morphine during Acute Inflammation of the Rat Paw. Pharmacology 2000, 60, 121–127. [Google Scholar] [CrossRef]
- Joris, J.; Costello, A.; Dubner, R.; Hargreaves, K.M. Opiates Suppress Carrageenan-Induced Edema and Hyperthermia at Doses That Inhibit Hyperalgesia. Pain 1990, 43, 95–103. [Google Scholar] [CrossRef]
- Honoré, P.; Buritova, J.; Besson, J.M. Intraplantar Morphine Depresses Spinal C-Fos Expression Induced by Carrageenin Inflammation but Not by Noxious Heat. Br. J. Pharmacol. 1996, 118, 671–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.-X.; Lei, L.-G.; Wang, Y.; Da, D.-F.; Zhao, Z.-Q. Endomorphin-1 Reduces Carrageenan-Induced Fos Expression in the Rat Spinal Dorsal Horn. Neuropeptides 1999, 33, 281–284. [Google Scholar] [CrossRef]
- Fletcher, D.; Gentili, M.; Mazoit, J.X.; Samii, K. Additivity of Bupivacaine and Morphine for Peripheral Analgesia in Rats. Fundam. Clin. Pharmacol. 2000, 14, 327–334. [Google Scholar] [CrossRef]
- Walker, J.S. Anti-Inflammatory Effects of Opioids. Adv. Exp. Med. Biol. 2003, 521, 148–160. [Google Scholar]
- Hu, Q.; Wang, Q.; Han, C.; Yang, Y. Sufentanil Attenuates Inflammation and Oxidative Stress in Sepsisinduced Acute Lung Injury by Downregulating KNG1 Expression. Mol. Med. Rep. 2020, 22, 4298–4306. [Google Scholar] [CrossRef]
- Rahimi, S.; Dadfar, B.; Tavakolian, G.; Asadi Rad, A.; Rashid Shabkahi, A.; Siahposht-Khachaki, A. Morphine Attenuates Neuroinflammation and Blood-Brain Barrier Disruption Following Traumatic Brain Injury through the Opioidergic System. Brain Res. Bull. 2021, 176, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yang, X.; Shu, S.; Wang, S.; Guo, F.; Yin, Y.; Zhou, W.; Han, H.; Chai, X. Sufentanil Protects the Liver from Ischemia/Reperfusion-Induced Inflammation and Apoptosis by Inhibiting ATF4-Induced TP53BP2 Expression. Inflammation 2021, 44, 1160–1174. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, R.; Frass, M.; Gmeiner, B.; Sandor, N.; Schumann, R.; Wagner, O.; Kaye, A.D. Effects of Remifentanil on Neutrophil Adhesion, Transmigration, and Intercellular Adhesion Molecule Expression. Acta Anaesthesiol. Scand. 2000, 44, 1232–1237. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Zhang, Y.; Su, W.; Zhou, L.; Xu, J.; Xia, Z. Remifentanil Attenuates Lipopolysaccharide-Induced Oxidative Injury by Downregulating PKCβ2 Activation and Inhibiting Autophagy in H9C2 Cardiomyocytes. Life Sci. 2018, 213, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Shen, X.; Nan, H.; Yan, L.; Zhao, H.; Yu, J.; Lv, Y. Remifentanil Protects Liver against Ischemia/Reperfusion Injury through Activation of Anti-Apoptotic Pathways. J. Surg. Res. 2013, 183, 827–834. [Google Scholar] [CrossRef]
- Hyejin, J.; Mei, L.; Seongheon, L.; Cheolwon, J.; Seokjai, K.; Hongbeom, B.; Minsun, K.; Sungsu, C.; Sanghyun, K. Remifentanil Attenuates Human Neutrophils Activation Induced by Lipopolysaccharide. Immunopharmacol. Immunotoxicol. 2013, 35, 264–271. [Google Scholar] [CrossRef]
- Maeda, S.; Andoh, T.; Onishi, R.; Tomoyasu, Y.; Higuchi, H.; Miyawaki, T. Remifentanil Suppresses Increase in Interleukin-6 MRNA in the Brain by Inhibiting Cyclic AMP Synthesis. J. Anesth. 2018, 32, 731–739. [Google Scholar] [CrossRef]
- Hasegawa, A.; Iwasaka, H.; Hagiwara, S.; Hasegawa, R.; Kudo, K.; Kusaka, J.; Asai, N.; Noguchi, T. Remifentanil and Glucose Suppress Inflammation in a Rat Model of Surgical Stress. Surg. Today 2011, 41, 1617. [Google Scholar] [CrossRef]
- Lu, Y.; Ding, X.; Wu, X.; Huang, S. Ketamine Inhibits LPS-Mediated BV2 Microglial Inflammation via NMDA Receptor Blockage. Fundam. Clin. Pharmacol. 2020, 34, 229–237. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, Y.; Chen, X.; Zhu, M.; Miao, C. Propofol Attenuates BV2 Microglia Inflammation via NMDA Receptor Inhibition. Can. J. Physiol. Pharmacol. 2017, 96, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Inada, T.; Kamibayashi, T. Protective Effect of the Intravenous Anesthetic Propofol against a Local Inflammation in the Mouse Carrageenan-Induced Air Pouch Model. Immunopharmacol. Immunotoxicol. 2021, 43, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jin, Y.-W.; Wang, D.-Y.; Wang, Z.-G. Effect of propofol upon brain after whole-body hyperthermia in rats. Zhonghua Yi Xue Za Zhi 2009, 89, 2356–2359. [Google Scholar] [PubMed]
- Plachinta, R.V.; Hayes, J.K.; Cerilli, L.A.; Rich, G.F. Isoflurane Pretreatment Inhibits Lipopolysaccharide-Induced Inflammation in Rats. Anesthesiology 2003, 98, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Guo, L.; Wang, Y.; Song, S. Isoflurane Upregulates MicroRNA-9-3p to Protect Rats from Hepatic Ischemia-Reperfusion Injury through Inhibiting Fibronectin Type III Domain Containing 3B. Cell Cycle 2021, 20, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, X.; Feng, Z.; Chen, Y.; Wen, D.; Liu, Z. The Protective Effect of Isoflurane Pretreatment on Liver IRI by Suppressing Noncanonical Pyroptosis of Liver Macrophages. Int. Immunopharmacol. 2021, 99, 107977. [Google Scholar] [CrossRef]
- Blondonnet, R.; Simand, L.-A.; Vidal, P.; Borao, L.; Bourguignon, N.; Morand, D.; Bernard, L.; Roszyk, L.; Audard, J.; Godet, T.; et al. Design and Rationale of the Sevoflurane for Sedation in Acute Respiratory Distress Syndrome (SESAR) Randomized Controlled Trial. J. Clin. Med. 2022, 11, 2796. [Google Scholar] [CrossRef]
- Lv, G.; Li, C.; Wang, W.; Li, N.; Wang, K. Silencing SP1 Alleviated Sevoflurane-Induced POCD Development via Cholinergic Anti-Inflammatory Pathway. Neurochem. Res. 2020, 45, 2082–2090. [Google Scholar] [CrossRef]
- Ngamsri, K.-C.; Fabian, F.; Fuhr, A.; Gamper-Tsigaras, J.; Straub, A.; Fecher, D.; Steinke, M.; Walles, H.; Reutershan, J.; Konrad, F.M. Sevoflurane Exerts Protective Effects in Murine Peritonitis-Induced Sepsis via Hypoxia-Inducible Factor 1α/Adenosine A2B Receptor Signaling. Anesthesiology 2021, 135, 136–150. [Google Scholar] [CrossRef]
- Wang, F.; Li, C.; Shao, J.; Ma, J. Sevoflurane Induces Inflammation of Microglia in Hippocampus of Neonatal Rats by Inhibiting Wnt/β-Catenin/CaMKIV Pathway. J. Pharmacol. Sci. 2021, 146, 105–115. [Google Scholar] [CrossRef]
- Shen, Q.-Y.; Wu, L.; Wei, C.-S.; Zhou, Y.-N.; Wu, H.-M. Sevoflurane Prevents Airway Remodeling via Downregulation of VEGF and TGF-Β1 in Mice with OVA-Induced Chronic Airway Inflammation. Inflammation 2019, 42, 1015–1022. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, L.; Huang, Q.; Wu, G.; Weng, X.; Lai, Z.; Lin, P. Monitoring the End-Tidal Concentration of Sevoflurane for Preventing Awareness during Anesthesia (MEETS-PANDA): A Prospective Clinical Trial. Int. J. Surg. 2017, 41, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Papurica, M.; Rogobete, A.F.; Sandesc, D.; Cradigati, C.A.; Sarandan, M.; Crisan, D.C.; Horhat, F.G.; Boruga, O.; Dumache, R.; Nilima, K.R.; et al. The Expression of Nuclear Transcription Factor Kappa B (NF-KappaB) in the Case of Critically Ill Polytrauma Patients with Sepsis and Its Interactions with MicroRNAs. Biochem. Genet. 2016, 54, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Di Raimondo, D.; Pirera, E.; Rizzo, G.; Simonetta, I.; Musiari, G.; Tuttolomondo, A. Non-Coding RNA Networks as Potential Novel Biomarker and Therapeutic Target for Sepsis and Sepsis-Related Multi-Organ Failure. Diagnostics 2022, 12, 1355. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Chen, K.-Y.; Shih, H.-J.; Chiang, M.; Huang, I.-T.; Huang, Y.-H.; Huang, C.-J. Let-7i-5p Mediates the Therapeutic Effects of Exosomes from Human Placenta Choriodecidual Membrane-Derived Mesenchymal Stem Cells on Mitigating Endotoxin-Induced Mortality and Liver Injury in High-Fat Diet-Induced Obese Mice. Pharmaceuticals 2022, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Kim, M.; Yun, S.; Doh, J.; Greenberg, P.D.; Kim, T.-D.; Choi, I. MicroRNA-150 Regulates the Cytotoxicity of Natural Killers by Targeting Perforin-1. J. Allergy Clin. Immunol. 2014, 134, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Acta, A.; Twaroski, D.; Zj, B.; Bai, X. Pharmaceutica MicroRNAs: New Players in Anesthetic-Induced Developmental Neurotoxicity. Pharm. Anal. Acta 2015, 6, 357. [Google Scholar] [CrossRef] [Green Version]
- Manquat, E.; Ravaux, H.; Kindermans, M.; Joachim, J.; Serrano, J.; Touchard, C.; Mateo, J.; Mebazaa, A.; Gayat, E.; Vallée, F.; et al. Impact of Impaired Cerebral Blood Flow Autoregulation on Electroencephalogram Signals in Adults Undergoing Propofol Anaesthesia: A Pilot Study. BJA Open 2022, 1, 100004. [Google Scholar] [CrossRef]
- Gusar, V.A.; Timofeeva, A.V.; Zhanin, I.S.; Shram, S.I.; Pinelis, V.G. Estimation of Time-Dependent MicroRNA Expression Patterns in Brain Tissue, Leukocytes, and Blood Plasma of Rats under Photochemically Induced Focal Cerebral Ischemia. Mol. Biol. 2017, 51, 602–613. [Google Scholar] [CrossRef]
- Zheng, T.; Shi, Y.; Zhang, J.; Peng, J.; Zhang, X.; Chen, K.; Chen, Y.; Liu, L. MiR-130a Exerts Neuroprotective Effects against Ischemic Stroke through PTEN/PI3K/AKT Pathway. Biomed. Pharmacother. 2019, 117, 109117. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, Y.; Duan, Z.; Liang, J.; Xu, Y.; Zhang, S.; Tang, T. Restored MicroRNA-326-5p Inhibits Neuronal Apoptosis and Attenuates Mitochondrial Damage via Suppressing STAT3 in Cerebral Ischemia/Reperfusion Injury. Nanoscale Res. Lett. 2021, 16, 63. [Google Scholar] [CrossRef]
- Guo, F.; Han, X.; Zhang, J.; Zhao, X.; Lou, J.; Chen, H.; Huang, X. Repetitive Transcranial Magnetic Stimulation Promotes Neural Stem Cell Proliferation via the Regulation of MiR-25 in a Rat Model of Focal Cerebral Ischemia. PLoS ONE 2014, 9, e109267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Liu, X.; Cao, J.; Meng, F.; Li, M.; Chen, B.; Zhang, J. MiR-134 Regulates Ischemia/Reperfusion Injury-Induced Neuronal Cell Death by Regulating CREB Signaling. J. Mol. Neurosci. 2014, 55, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-F.; Shi, L.-L.; Zhang, L.; Zhao, Z.-H.; Liang, F.; Xu, X.; Zhao, L.-Y.; Yang, P.-B.; Zhang, J.-S.; Tian, Y.-f. MicroRNA-25 Negatively Regulates Cerebral Ischemia/Reperfusion Injury-Induced Cell Apoptosis Through Fas/FasL Pathway. J. Mol. Neurosci. 2016, 58, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Ren, P.; Wang, G.; Yan, F.; Zhang, Y. MicroRNA-128-3p Protects Mouse Against Cerebral Ischemia Through Reducing P38α Mitogen-Activated Protein Kinase Activity. J. Mol. Neurosci. 2017, 61, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-P.; Zhang, W.-J.; Yang, M.; Fang, H. Propofol Attenuates Lung Ischemia/Reperfusion Injury Though the Involvement of the MALAT1/MicroRNA-144/GSK3β Axis. Mol. Med. 2021, 27, 77. [Google Scholar] [CrossRef]
- Li, G.; Xu, M.; Wang, H.; Qi, X.; Wang, X.; Li, Y.; Sun, J.; Li, Y. MicroRNA-146a Overexpression Alleviates Intestinal Ischemia/Reperfusion-Induced Acute Lung Injury in Mice. Exp. Ther. Med. 2021, 22, 937. [Google Scholar] [CrossRef]
- Zhou, Y.; Dong, H.; Zhong, Y.; Huang, J.; Lv, J.; Li, J. The Cold-Inducible RNA-Binding Protein (CIRP) Level in Peripheral Blood Predicts Sepsis Outcome. PLoS ONE 2015, 674, e0137721. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Qi, W.; Dai, S.; Zou, G.; Liu, W.; Yu, B.; Tang, J. MicroRNA-223 Promotes Autophagy to Aggravate Lung Ischemia-Reperfusion Injury by Inhibiting the Expression of Transcription Factor HIF2α. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L1–L10. [Google Scholar] [CrossRef]
- Cai, J.; Gehrau, R.; Tu, Z.; Leroy, V.; Su, G.; Shang, J.; Mas, V.R.; Emtiazjoo, A.; Pelaez, A.; Atkinson, C.; et al. MicroRNA-206 AntagomiR—Enriched Extracellular Vesicles Attenuate Lung Ischemia—Reperfusion Injury through CXCL1 Regulation in Alveolar Epithelial Cells. J. Heart Lung Transpl. 2020, 39, 1476–1490. [Google Scholar] [CrossRef]
- Xiao, K.; Song, L.; Hu, Y.; He, W.; Hou, F.; Yan, P.; Xu, J.; Wang, K.; Tao, Y.; Li, D.; et al. Novel Role of MiR-18a-5p and Galanin in Rat Lung Ischemia Reperfusion-Mediated Response. Oxid. Med. Cell Longev. 2021, 2021, 6621921. [Google Scholar] [CrossRef]
- Qin, L.-B.; Li, Z.-Y.; Li, H.; Fan, X.-Q.; Liu, H.-G.; Dong, X.-M.; Jia, W.-Y. Inhibitive Effects of MicroRNA-34a on Protecting against Ischemia-Reperfusion Injury of Vital Organs in Hemorrhagic Shock Pregnant Mice. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1812–1818. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.; Wei, L.; Han, Z.; Chen, Z. Mesenchymal Stromal Cells-Derived Exosomes Alleviate Ischemia/Reperfusion Injury in Mouse Lung by Transporting Anti-Apoptotic MiR-21-5p. Eur. J. Pharmacol. 2019, 852, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Masaldan, S.; Bush, A.I.; Devos, D.; Rolland, A.S.; Moreau, C. Striking While the Iron Is Hot: Iron Metabolism and Ferroptosis in Neurodegeneration. Free Radic. Biol. Med. 2018, 133, 221–233. [Google Scholar] [CrossRef]
- Tuo, Q.-Z.; Lei, P.; Jackman, K.A.; Li, X.-L.; Xiong, H.; Li, X.-L.; Liuyang, Z.-Y.; Roisman, L.; Zhang, S.-T.; Ayton, S.; et al. Tau-Mediated Iron Export Prevents Ferroptotic Damage after Ischemic Stroke. Mol. Psychiatry 2017, 22, 1520–1530. [Google Scholar] [CrossRef]
- Dzikiewicz-Krawczyk, A.; Kok, K.; Slezak-Prochazka, I.; Robertus, J.-L.; Bruining, J.; Tayari, M.M.; Rutgers, B.; de Jong, D.; Koerts, J.; Seitz, A.; et al. ZDHHC11 and ZDHHC11B Are Critical Novel Components of the Oncogenic MYC-MiR-150-MYB Network in Burkitt Lymphoma. Leukemia 2017, 31, 1470–1473. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of Ferroptotic Cancer Cell Death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, X.; Cai, Z.; Wang, H.; Han, D.; Cheng, Q.; Zhang, P.; Gao, F.; Yu, Y.; Song, Z.; Wu, Q.; et al. Loss of Cardiac Ferritin H Facilitates Cardiomyopathy via Slc7a11-Mediated Ferroptosis. Circ. Res. 2020, 127, 486–501. [Google Scholar] [CrossRef]
- Drobna, M.; Szarzyńska, B.; Jaksik, R.; Sędek, Ł.; Kuchmiy, A.; Taghon, T.; van Vlierberghe, P.; Szczepański, T.; Witt, M.; Dawidowska, M. Hsa-MiR-20b-5p and Hsa-MiR-363-3p Affect Expression of PTEN and BIM Tumor Suppressor Genes and Modulate Survival of T-ALL Cells In Vitro. Cells 2020, 9, 1137. [Google Scholar] [CrossRef]
- Ding, C.; Ding, X.; Zheng, J.; Wang, B.; Li, Y.; Xiang, H.; Dou, M.; Qiao, Y.; Tian, P.; Xue, W. MiR-182-5p and MiR-378a-3p Regulate Ferroptosis in I/R-Induced Renal Injury. Cell Death Dis. 2020, 11, 929. [Google Scholar] [CrossRef]
- Chiba, T.; Cerqueira, D.M.; Li, Y.; Bodnar, A.J.; Mukherjee, E.; Pfister, K.; Phua, Y.L.; Shaikh, K.; Sanders, B.T.; Hemker, S.L.; et al. Endothelial-Derived MiR-17∽92 Promotes Angiogenesis to Protect against Renal Ischemia-Reperfusion Injury. J. Am. Soc. Nephrol. 2021, 32, 553–562. [Google Scholar] [CrossRef]
- Lorenzen, J.M.; Kielstein, J.T.; Hafer, C.; Gupta, S.K.; Ku, P. Circulating MiR-210 Predicts Survival in Critically Ill Patients with Acute Kidney Injury. Clin. J. Am. Soc. Nephrol. 2011, 6, 1540–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-Y.; Gao, J.; Song, Y.-H.; Cai, G.-Y.; Chen, X.-M. Identification of Potential Gene and MicroRNA Biomarkers of Acute Kidney Injury. Biomed. Res. Int. 2021, 2021, 8834578. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.-Y.; Wang, B.; Tang, T.-T.; Wen, Y.; Li, Z.-L.; Feng, S.-T.; Wu, M.; Liu, D.; Yin, D.; Ma, K.-L.; et al. Exosomal MiR-125b-5p Deriving from Mesenchymal Stem Cells Promotes Tubular Repair by Suppression of P53 in Ischemic Acute Kidney Injury. Theranostics 2021, 11, 5248–5266. [Google Scholar] [CrossRef]
- Wei, Q.; Liu, Y.; Liu, P.; Hao, J.; Liang, M.; Mi, Q.-S.; Chen, J.-K.; Dong, Z. MicroRNA-489 Induction by Hypoxia-Inducible Factor-1 Protects against Ischemic Kidney Injury. J. Am. Soc. Nephrol. 2016, 27, 2784–2796. [Google Scholar] [CrossRef] [Green Version]
- Aguado-fraile, E.; Ramos, E.; Conde, E.; Rodríguez, M.; Martín-, L. A Pilot Study Identifying a Set of MicroRNAs as Precise Diagnostic Biomarkers of Acute Kidney Injury. PLoS ONE 2015, 10, e0127175. [Google Scholar] [CrossRef]
- Bruno, N.; ter Maaten, J.M.; Ovchinnikova, E.S.; Vegter, E.L.; Valente, M.A.E.; van der Meer, P.; de Boer, R.A.; van der Harst, P.; Schmitter, D.; Metra, M.; et al. MicroRNAs Relate to Early Worsening of Renal Function in Patients with Acute Heart Failure. Int. J. Cardiol. 2015, 203, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, K.; Saikumar, J.; Bijol, V.; Koyner, J.L.; Qian, J.; Betensky, R.A.; Waikar, S.S.; Vaidya, V.S. Human MiRNome Profiling Identifies MicroRNAs Differentially Present in the Urine after Kidney Injury. Clin. Chem. 2013, 59, 1742–1752. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Lan, Y.; Li, H.; Cheng, C.; Lai, P. Urinary MiR-16 Transactivated by C / EBP β Reduces Kidney Function after Ischemia/Reperfusion—Induced Injury. Nat. Publ. Group 2016, 6, 27945. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.-M.; Huang, C.-M.; Zhu, X.-Y.; Bian, F.; Pan, S.-M. Differentially Expressed MiRNAs in Sepsis-Induced Acute Kidney Injury Target Oxidative Stress and Mitochondrial Dysfunction Pathways. PLoS ONE 2017, 12, e0173292. [Google Scholar] [CrossRef]
- Pavkovic, M.; Riefke, B.; Ellinger-Ziegelbauer, H. Urinary MicroRNA Profiling for Identification of Biomarkers after Cisplatin-Induced Kidney Injury. Toxicology 2014, 324, 147–157. [Google Scholar] [CrossRef]
- Gutiérrez-Escolano, A.; Santacruz-Vázquez, E.; Gómez-Pérez, F. Dysregulated MicroRNAs Involved in Contrast-Induced Acute Kidney Injury in Rat and Human. Ren. Fail. 2015, 37, 1498–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Che, L.; Wang, Y.; Zhou, H.; Gong, H.; Man, X.; Zhao, Q. Deregulated MicroRNA-22-3p in Patients with Sepsis-Induced Acute Kidney Injury Serves as a New Biomarker to Predict Disease Occurrence and 28-Day Survival Outcomes. Int. Urol. Nephrol. 2021, 53, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Shihana, F.; Joglekar, M.V.; Raubenheimer, J.; Hardikar, A.A.; Buckley, N.A.; Seth, D. Circulating Human MicroRNA Biomarkers of Oxalic Acid-Induced Acute Kidney Injury. Arch. Toxicol. 2020, 94, 1725–1737. [Google Scholar] [CrossRef] [PubMed]
- Collier, J.B.; Schnellmann, R.G. Extracellular Signal-Regulated Kinase 1/2 Regulates NAD Metabolism during Acute Kidney Injury through MicroRNA-34a-Mediated NAMPT Expression. Cell. Mol. Life Sci. 2020, 77, 3643–3655. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jing, Y.; Hao, J.; Frankfort, N.C.; Zhou, X.; Shen, B. MicroRNA-21 in the Pathogenesis of Acute Kidney Injury. Protein Cell 2013, 4, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, Y.; Xue, S.; Wang, X.; Dai, H.; Qian, J. Implications of Dynamic Changes in MiR-192 Expression in Ischemic Acute Kidney Injury. Int. Urol. Nephrol. 2017, 49, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Uhlenhaut, N.H.; Treier, M. Transcriptional Regulators in Kidney Disease: Gatekeepers of Renal Homeostasis. Trends Genet. 2008, 24, 361–371. [Google Scholar] [CrossRef]
- Tan, R.J.; Zhou, D.; Zhou, L.; Liu, Y. Wnt/β-Catenin Signaling and Kidney Fibrosis. Kidney Int. Suppl. 2014, 4, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Niehrs, C. Function and Biological Roles of the Dickkopf Family of Wnt Modulators. Oncogene 2006, 25, 7469–7481. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.-J.; Li, L.-L.; Tu, W.-B. MiR-214 Negatively Regulates Proliferation and WNT/β-Catenin Signaling in Breast Cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5148–5154. [Google Scholar]
- Zhu, X.; Li, W.; Li, H. MiR-214 Ameliorates Acute Kidney Injury via Targeting DKK3 and Activating of Wnt/β-Catenin Signaling Pathway. Biol. Res. 2018, 51, 31. [Google Scholar] [CrossRef] [PubMed]
- Marcil, V.; Lavoie, J.C.; Emonnot, L.; Seidman, E.; Levy, E. Analysis of the Effects of Iron and Vitamin C Co-Supplementation on Oxidative Damage, Antioxidant Response and Inflammation in THP-1 Macrophages. Clin. Biochem. 2011, 44, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Reyes-García, J.; Carbajal-García, A.; di Mise, A.; Zheng, Y.M.; Wang, X.; Wang, Y.X. Important Functions and Molecular Mechanisms of Mitochondrial Redox Signaling in Pulmonary Hypertension. Antioxidants 2022, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Ashok, A.; Andrabi, S.S.; Mansoor, S.; Kuang, Y.; Kwon, B.K.; Labhasetwar, V. Antioxidant Therapy in Oxidative Stress-Induced Neurodegenerative Diseases: Role of Nanoparticle-Based Drug Delivery Systems in Clinical Translation. Antioxidants 2022, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Baev, A.Y.; Vinokurov, A.Y.; Novikova, I.N.; Dremin, V.V.; Potapova, E.V.; Abramov, A.Y. Interaction of Mitochondrial Calcium and ROS in Neurodegeneration. Cells 2022, 11, 706. [Google Scholar] [CrossRef]
- Tripatara, P.; Patel, N.S.A.; Collino, M.; Gallicchio, M.; Kieswich, J.; Castiglia, S.; Benetti, E.; Stewart, K.N.; Brown, P.A.; Yaqoob, M.M.; et al. Generation of Endogenous Hydrogen Sulfide by Cystathionine γ-Lyase Limits Renal Ischemia/Reperfusion Injury and Dysfunction. Lab. Investig. 2008, 88, 1038–1048. [Google Scholar] [CrossRef] [Green Version]
- Bos, E.M.; Leuvenink, H.G.D.; Snijder, P.M.; Kloosterhuis, N.J.; Hillebrands, J.-L.; Leemans, J.C.; Florquin, S.; van Goor, H. Hydrogen Sulfide-Induced Hypometabolism Prevents Renal Ischemia/Reperfusion Injury. J. Am. Soc. Nephrol. 2009, 20, 1901–1905. [Google Scholar] [CrossRef] [Green Version]
- Granata, S.; Votrico, V.; Spadaccino, F.; Catalano, V.; Netti, G.S.; Ranieri, E.; Stallone, G.; Zaza, G. Oxidative Stress and Ischemia/Reperfusion Injury in Kidney Transplantation: Focus on Ferroptosis, Mitophagy and New Antioxidants. Antioxidants 2022, 11, 769. [Google Scholar] [CrossRef]
- Birk, A.V.; Liu, S.; Soong, Y.; Mills, W.; Singh, P.; Warren, J.D.; Seshan, S.V.; Pardee, J.D.; Szeto, H.H. The Mitochondrial-Targeted Compound SS-31 Re-Energizes Ischemic Mitochondria by Interacting with Cardiolipin. J. Am. Soc. Nephrol. 2013, 24, 1250–1261. [Google Scholar] [CrossRef]
- Szeto, H.H.; Liu, S.; Soong, Y.; Seshan, S.V.; Cohen-Gould, L.; Manichev, V.; Feldman, L.C.; Gustafsson, T. Mitochondria Protection after Acute Ischemia Prevents Prolonged Upregulation of IL-1β and IL-18 and Arrests CKD. J. Am. Soc. Nephrol. 2016, 28, 1437–1449. [Google Scholar] [CrossRef] [Green Version]
- Laight, D.W.; Andrews, T.J.; Haj-Yehia, A.I.; Carrier, M.J.; Änggård, E.E. Microassay of Superoxide Anion Scavenging Activity in Vitro. Environ. Toxicol. Pharmacol. 1997, 3, 65–68. [Google Scholar] [CrossRef]
- Chatterjee, P.; Bhattacharyya, M.; Bandyopadhyay, S.; Roy, D. Studying the System-Level Involvement of MicroRNAs in Parkinson’s Disease. PLoS ONE 2014, 9, e93751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, T.; Kurihara, S. Cystine and Theanine as Stress-Reducing Amino Acids—Perioperative Use for Early Recovery after Surgical Stress. Nutrients 2022, 14, 129. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.; Bissolati, M.; Rocchetti, S.; Beneduce, A.; Pecorelli, N.; di Carlo, V. Oral Preoperative Antioxidants in Pancreatic Surgery: A Double-Blind, Randomized, Clinical Trial. Nutrition 2012, 28, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Leong, J.-Y.; van der Merwe, J.; Pepe, S.; Bailey, M.; Perkins, A.; Lymbury, R.; Esmore, D.; Marasco, S.; Rosenfeldt, F. Perioperative Metabolic Therapy Improves Redox Status and Outcomes in Cardiac Surgery Patients: A Randomised Trial. Heart Lung Circ. 2010, 19, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, T.; Tsuchiya, T.; Oyama, A.; Tsuchiya, T.; Abe, N.; Sato, A.; Chiba, Y.; Kurihara, S.; Shibakusa, T.; Mikami, T. Perioperative Oral Administration of Cystine and Theanine Enhances Recovery after Distal Gastrectomy: A Prospective Randomized Trial. J. Parenter. Enteral. Nutr. 2012, 37, 384–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibakusa, T.; Mikami, T.; Kurihara, S.; Chiba, Y.; Tsuchiya, T.; Miyachi, T.; Oyama, A.; Tanaka, K.A.K.; Koyama, N. Enhancement of Postoperative Recovery by Preoperative Oral Co-Administration of the Amino Acids, Cystine and Theanine, in a Mouse Surgical Model. Clin. Nutr. 2012, 31, 555–561. [Google Scholar] [CrossRef]
- Tanaka, K.A.; Mazzeffi, M.; Durila, M. Role of Prothrombin Complex Concentrate in Perioperative Coagulation Therapy. J. Intensive Care 2014, 2, 60. [Google Scholar] [CrossRef] [Green Version]
- Dingchao, H.; Zhiduan, Q.; Liye, H.; Xiaodong, F. The Protective Effects of High-Dose Ascorbic Acid on Myocardium against Reperfusion Injury during and after Cardiopulmonary Bypass. Thorac. Cardiovasc. Surg. 1994, 42, 276–278. [Google Scholar] [CrossRef]
- Nathens, A.B.; Neff, M.J.; Jurkovich, G.J.; Klotz, P.; Farver, K.; Ruzinski, J.T.; Radella, F.; Garcia, I.; Maier, R.V. Randomized, Prospective Trial of Antioxidant Supplementation in Critically Ill Surgical Patients. Ann. Surg. 2002, 236, 814–822. [Google Scholar] [CrossRef]
- Tanaka, H.; Matsuda, T.; Miyagantani, Y.; Yukioka, T.; Matsuda, H.; Shimazaki, S. Reduction of Resuscitation Fluid Volumes in Severely Burned Patients Using Ascorbic Acid Administration: A Randomized, Prospective Study. Arch. Surg. 2000, 135, 326–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.A.; Beers, R.J.; Lentz, C.W. Resuscitation after Severe Burn Injury Using High-Dose Ascorbic Acid: A Retrospective Review. J. Burn Care Res. 2011, 32, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Shaw, G.M.; Fowler, A.A.; Natarajan, R. Ascorbate-Dependent Vasopressor Synthesis: A Rationale for Vitamin C Administration in Severe Sepsis and Septic Shock? Crit. Care 2015, 19, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakes, B.; Bolia, I.K.; Weber, A.E.; Petrigliano, F.A. Vitamin C in Orthopedic Practices: Current Concepts, Novel Ideas, and Future Perspectives. J. Orthop. Res. 2021, 39, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Suter, M. Positive End-Expiratory Pressure During Induction of General Anesthesia Increases Duration of Nonhypoxic Apnea in Morbidly Obese Patients. Anesth. Analg. 2005, 100, 580–584. [Google Scholar] [CrossRef]
- Chen, S.; Roffey, D.M.; Dion, C.-A.; Arab, A.; Wai, E.K. Effect of Perioperative Vitamin C Supplementation on Postoperative Pain and the Incidence of Chronic Regional Pain Syndrome: A Systematic Review and Meta-Analysis. Clin. J. Pain 2016, 32, 179–185. [Google Scholar] [CrossRef]
- Ngo, B.; van Riper, J.M.; Cantley, L.C.; Yun, J. Targeting Cancer Vulnerabilities with High-Dose Vitamin C. Nat. Rev. Cancer 2019, 19, 271–282. [Google Scholar] [CrossRef]
- Du, Y.T.; Long, Y.; Tang, W.; Liu, X.F.; Dai, F.; Zhou, B. Prooxidative Inhibition against NF-ΚB-Mediated Inflammation by Pharmacological Vitamin C. Free Radic. Biol. Med. 2022, 180, 85–94. [Google Scholar] [CrossRef]
- Brown, M.L.; Braun, M.; Cicalese, L.; Rastellini, C. Effect of Perioperative Antioxidant Therapy on Suboptimal Islet Transplantation in Rats. Transplant. Proc. 2005, 37, 217–219. [Google Scholar] [CrossRef]
- Moon, S.; Lim, S.H.; Cho, K.; Kim, M.H.; Lee, W.; Cho, Y.H. The Efficacy of Vitamin C on Postlaparoscopic Shoulder Pain: A Double-Blind Randomized Controlled Trial. Anesth. Pain Med. 2019, 14, 202–207. [Google Scholar] [CrossRef]
- Mohamed, M.H.; Hamawy, T.Y. Comparative Evaluation between Ascorbic Acid and N-Acetyl Cysteine for Preventing Tourniquet Induced Ischaemic Reperfusion Injury during Lower Limb Surgery, a Randomized Controlled Trial. Egypt. J. Anaesth. 2016, 32, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Rümelin, A.; Dörr, S.; Fauth, U. Single Preoperative Oral Application of Ascorbic Acid Does Not Affect Postoperative Plasma Levels of Ascorbic Acid. Ann. Nutr. Metab. 2002, 46, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.; Park, J.S.; Moon, S.; Yeo, J. Effect of Intravenous High Dose Vitamin C on Postoperative Pain and Morphine Use after Laparoscopic Colectomy: A Randomized Controlled Trial. Pain Res. Manag. 2016, 2016, 9147279. [Google Scholar] [CrossRef] [PubMed]
- Holford, P.; Carr, A.C.; Jovic, T.H.; Ali, S.R.; Whitaker, I.S.; Marik, P.E.; Smith, A.D. Vitamin C-An Adjunctive Therapy for Respiratory Infection, Sepsis and COVID-19. Nutrients 2020, 12, 3760. [Google Scholar] [CrossRef] [PubMed]
- Martín Giménez, V.M.; Chuffa, L.G.A.; Simão, V.A.; Reiter, R.J.; Manucha, W. Protective Actions of Vitamin D, Anandamide and Melatonin during Vascular Inflammation: Epigenetic Mechanisms Involved. Life Sci. 2022, 288, 120191. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef]
- Nieuwland, A.J.; Kokje, V.B.C.; Koning, O.H.; Hamming, J.F.; Szuhai, K.; Claas, F.H.J.; Lindeman, J.H.N. Activation of the Vitamin D Receptor Selectively Interferes with Calcineurin-Mediated Inflammation: A Clinical Evaluation in the Abdominal Aortic Aneurysm. Lab. Investig. 2016, 96, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Moreno, J.M.; Herencia, C.; de Oca, A.M.; Díaz-Tocados, J.M.; Vergara, N.; Gómez-Luna, M.J.; López-Argüello, S.D.; Camargo, A.; Peralbo-Santaella, E.; Rodríguez-Ortiz, M.E.; et al. High Phosphate Induces a Pro-Inflammatory Response by Vascular Smooth Muscle Cells and Modulation by Vitamin D Derivatives. Clin. Sci. 2017, 131, 1449–1463. [Google Scholar] [CrossRef]
- Bogunovic, L.; Kim, A.D.; Beamer, B.S.; Nguyen, J.; Lane, J.M. Hypovitaminosis D in Patients Scheduled to Undergo Orthopaedic Surgery: A Single-Center Analysis. J. Bone Joint Surg. Am. 2010, 92, 2300–2304. [Google Scholar] [CrossRef]
- Maniar, R.N.; Patil, A.M.; Maniar, A.R.; Gangaraju, B.; Singh, J. Effect of Preoperative Vitamin D Levels on Functional Performance after Total Knee Arthroplasty. Clin. Orthop. Surg. 2016, 8, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Mouli, V.H.; Schudrowitz, N.; Carrera, C.X.; Uzosike, A.C.; Fitz, W.; Rajaee, S.S. High-Dose Vitamin D Supplementation Can Correct Hypovitaminosis D Prior to Total Knee Arthroplasty. J. Arthroplast. 2022, 37, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Hajimohammadebrahim-Ketabforoush, M.; Shahmohammadi, M.; Keikhaee, M.; Eslamian, G.; Vahdat Shariatpanahi, Z. Single High-Dose Vitamin D3 Injection and Clinical Outcomes in Brain Tumor Resection: A Randomized, Controlled Clinical Trial. Clin. Nutr. ESPEN 2021, 41, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Alshafey, M.K.; Elrakhawy, H.M.; Rezk, M.E.; Moustafa, H.M. Role of Ascorbic Acid in Reduction of the Incidence of the Atrial Fibrillation in Patients under B-Blocker and Undergoing Coronary Artery Bypass Graft Operation in Early Post-Operative Period. J. Egypt. Soc. Cardio-Thorac. Surg. 2017, 25, 198–203. [Google Scholar] [CrossRef]
- Gunes, T.; Bozok, S.; Kestelli, M.; Yurekli, I.; Ilhan, G.; Ozpak, B.; Bademci, M.; Ozcem, B.; Sahin, A. α-Tocopherol and Ascorbic Acid in Early Postoperative Period of Cardiopulmonary Bypass. J. Cardiovasc. Med. 2012, 13, 691–699. [Google Scholar] [CrossRef]
- Castillo, R.L.; Loza, R.C. Pathophysiological Approaches of Acute Respiratory Distress Syndrome: Novel Bases for Study of Lung Injury. Open Respir. Med. J. 2015, 9, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Angdin, M.; Settergren, G.; Starkopf, J.; Zilmer, M.; Zilmer, K.; Vaage, J. Protective Effect of Antioxidants on Pulmonary Endothelial Function after Cardiopulmonary Bypass. J. Cardiothorac. Vasc. Anesth. 2003, 17, 314–320. [Google Scholar] [CrossRef]
- Antonic, M. Effect of Ascorbic Acid on Postoperative Acute Kidney Injury in Coronary Artery Bypass Graft Patients: A Pilot Study. Heart Surg. Forum 2017, 20, E214–E218. [Google Scholar] [CrossRef]
- Das, D.; Sen, C.; Goswami, A. Effect of Vitamin C on Adrenal Suppression by Etomidate Induction in Patients Undergoing Cardiac Surgery: A Randomized Controlled Trial. Ann. Card. Anaesth. 2016, 19, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Sadeghpour, A.; Alizadehasl, A.; Kyavar, M.; Sadeghi, T.; Moludi, J. Impact of Vitamin C Supplementation on Post-Cardiac Surgery ICU and Hospital Length of Stay. Anesthesiol. Pain Med. 2015, 5, e25337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samadikhah, J.; Golzari, S.E.J.; Sabermarouf, B.; Karimzadeh, I.; Tizro, P.; Khanli, H.M.; Ghabili, K. Efficacy of Combination Therapy of Statin and Vitamin C in Comparison with Statin in the Prevention of Post-CABG Atrial Fibrillation. Adv. Pharm. Bull. 2014, 4, 97–100. [Google Scholar] [CrossRef]
- Dehghani, M.R.; Majidi, N.; Rahmani, A.; Asgari, B.; Rezaei, Y. Effect of Oral Vitamin C on Atrial Fibrillation Development after Isolated Coronary Artery Bypass Grafting Surgery: A Prospective Randomized Clinical Trial. Cardiol. J. 2013, 21, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouybar, R.; Kabgani, H.; Kamalipour, H.; Shahbazi, S.; Allahyary, E.; Rasouli, M.; Hedayatallah Akhlagh, S.; Shafa, M.; Ghazinoor, M.; Taghi Moeinvaziri, M.; et al. The Perioperative Effect of Ascorbic Acid on Inflammatory Response in Coronary Artery Bypass Graft Surgery; A Randomized Controlled Trial Coronary Artery Bypass Graft Surgery. Int. Cardiovasc. Res. J. 2012, 6, e12835. [Google Scholar]
- Papoulidis, P.; Ananiadou, O.; Chalvatzoulis, E.; Ampatzidou, F.; Koutsogiannidis, C.; Karaiskos, T.; Madesis, A.; Drossos, G. The Role of Ascorbic Acid in the Prevention of Atrial Fibrillation after Elective On-Pump Myocardial Revascularization Surgery: A Single-Center Experience—A Pilot Study. Interact. CardioVascular Thorac. Surg. 2011, 12, 121–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collier, B.R.; Giladi, A.; Dossett, L.A.; Dyer, L.; Fleming, S.B.; Cotton, B.A. Impact of High-Dose Antioxidants on Outcomes in Acutely Injured Patients. J. Parenter. Enter. Nutr. 2008, 32, 384–388. [Google Scholar] [CrossRef]


{kind=link}
| Author | Anesthetic Drug | Study Type | Comments | References |
|---|---|---|---|---|
| Gyires et al. | Morphine | in vivo | ↓ edema; ↓ bradykinin activity; no influence on edema induced by histamine; | [122] |
| Sacerdote et al. | Morphine | in vivo | ↓ inflammatory response; ↑ activation of mu-opioid receptors; | [123] |
| Planas et al. | Liposomal morphine | in vivo | ↓ inflammatory edema; ↑duration of effects by encapsulation; | [124] |
| Joris et al. | Morphine | in vivo | ↓ vascular inflammation; ↓ vascular permeability; | [125] |
| Honore et al. | Morphine | in vivo | ↓ edema; ↓ Fos-like immunoreactive activity; | [126] |
| Jin et al. | Endomorphin-1 (Endogenous ligand for mu opioid receptor) | in vivo | ↓ peripheral edema; ↓ activity of Fos-like immunoreactive expression; | [127] |
| Fletcher et al. | Morphine | in vivo | ↓ inflammation; ↓ edema; | [128] |
| Walker et al. | Kappa-opioid drugs | in vivo | ↓ adhesion molecules; ↓ TNF expression; ↓ expression of inflammation cascade; | [129] |
| Hu et al. | Sufentanil | in vivo | ↑ super oxide dismutase (SOD) activity; ↑ catalase (CAT) activity; ↑ glutathione peroxidase (GSH-Px) activity; ↓ malondialdehyde (MDA) expression; ↓ Nuclear factor erythroid 2—related factor 2 (Nrf2) expression; ↓ inflammation and oxidative stress; | [130] |
| Rahimi et al. | Morphine | in vivo | ↓ neuroinflammation via opioid receptors; ↑ neurobehavioral function after traumatic brain injury; ↓ blood brain barrier (BBB) leakage; | [131] |
| Zhou et al. | Sufentanil | in vivo | ↓ Activating transcription factor 4 (AFT4) expression; ↓ liver inflammation; ↓ apoptosis injury induced by ischemia–reperfusion injury; | [132] |
| Hofbauer et al. | Remifentanil | in vitro | ↓ endothelial cell adhesion molecule; ↓ polymorphonuclear neutrophils migration; | [133] |
| Lei et al. | Remifentanil | in vitro | ↑ cardiomyocytes protection against oxidative stress; ↓ PKCβ expression; ↓ autophagy activity; | [134] |
| Zhao et al. | Remifentanil | in vivo | ↓ NF-kB expression; ↓ TNF-α expression; ↓ IAM-1 expression; ↑ protective effect against ischemia–reperfusion injury syndrome; | [135] |
| Hyejin et al. | Remifentanil | in vitro | ↓ human neutrophils activations induced by lipopolysaccharide (LPS); ↑ anti-inflammatory effects; | [136] |
| Maeda et al. | Remifentanil | in vivo | ↓ IL-6 expression in mouse brain; ↓ inflammation in cerebral tissue; | [137] |
| Hasegawa et al. | Remifentanil | in vivo | ↓ inflammation expression caused by surgical stress; | [138] |
| Lu et al. | Ketamine | in vitro | ↓ NMDA receptors activity; ↓ calmodulin-dependent protein kinase 2 (CAMK2) activity; ↓ NF-kB phosphorylation and nuclear translocation; ↑ protective against inflammation induced by LPS; | [139] |
| Wu et al. | Propofol | in vitro | ↑ NMDA expression; ↑ Ca2+ accumulation; ↓ NF-kB phosphorylation; ↓ LPS- induced pro-inflammatory cytokine; | [140] |
| Inada et al. | Propofol | in vitro | ↓ production of TNF-α and IL-6; ↓ production of neutrophil chemokines; ↑ anti-inflammatory effects; | [141] |
| Zhao et al. | Propofol | in vitro | ↓ activity of NLR family pyrin domain containing 3 (NLRP3); ↓ inflammation and apoptosis mechanisms; | [142] |
| Plachinta et al. | Isoflurane | in vivo | ↓ TNF-α activity; ↓ acidosis; ↓ damage to the vascular endothelium associated with LPS-induced inflammation; ↑ vascular protection during inflammation; | [143] |
| Wang et al. | Isoflurane | in vitro | ↑ microRNA-9-3p expression; ↓ activity of fibronectin type III domain containing 3B (FNDC3B); ↑ protection against hepatic ischemia–reperfusion injury; | [144] |
| Lu et al. | Isoflurane | in vivo | ↓ caspase-11, IL-1β and IL-18 expression; ↓ intracellular Ca2+ accumulation; ↑ protection against inflammation-induced by liver ischemia/ reperfusion injury; | [145] |
| Zhang et al. | Isoflurane | in vitro | ↓ heme-oxygenase-1 (HO1) activity; ↓ redox, inflammation and apoptosis expression in sepsis- induced brain damage; | [146] |
| Lv et al. | Sevoflurane | in vivo | ↑ neuro-inflammation and apoptosis by activation of cholinergic anti-inflammatory pathway; | [147] |
| Ngamsri et al. | Sevoflurane | in vivo | ↓ neutrophil expression in peritoneal lavage; ↑ protection against peritonitis- induced sepsis in rats; ↑ expression of hypoxia- inducible factor 1α and adenosine A2B receptor in liver, intestine and lung; | [148] |
| Wang et al. | Sevoflurane | in vivo | ↓ allergic airway inflammation; ↓ Th2 expression and NLRP3 activity; | [149] |
| Shen et al. | Sevoflurane | in vivo | ↓ acute lung inflammation in ovalbumin-induced allergic mices; ↑ protection against airway remodeling in mouse; ↓ VEGF and TGF-β1 activity in lung tissues; | [150] |
| Wang et al. | Sevoflurane | in vivo | ↑ inflammation of microglia in hippocampus of neonatal rats; ↓ activity of WNT/β-catenin/CaMKIV pathway; | [151] |
| Authors | Antioxidant | Study Description | Comments | Reference |
|---|---|---|---|---|
| Alshafey et al. | Vitamin C |
| ↓ incidence of atrial fibrillation; ↓ ventilator stay; ↓ need for inotropic support; | [243] |
| Gunes et al. | Vitamin C + α-tocopherol |
| ↓ C-reactive protein (CRP) in patients who received antioxidant therapy; ↓ white blood cell count in control group; ↓ systemic inflammatory response; | [244] |
| Castillo et al. | Vitamin C + omega 3 polyunsaturated fatty acids (n-3PUFA) |
| ↓ reduced/oxidized glutathione (GSH/GSSG) ratio; ↓ malondialdehyde (MDA) expression; ↓ lipid peroxidation; ↓ protein carbonylation; ↓ NF-kB activation; | [245] |
| Angdin et al. | Vitamin E + vitamin C + allopurinol + acetylcysteine |
| ↑ pulmonary vasodilatation protection; ↑ protection of endothelium-dependent vasodilation after cardio-pulmonary by-pass; ↓ endothelial dysfunction; | [246] |
| Antonic et al. | Vitamin C |
| -no protective effects on the incidence of acute renal injury; | [247] |
| Das et al. | Vitamin C |
| ↓ requirement of adrenaline; -no effects off length stay in ICU, and time of extubation; | [248] |
| Sadeghpour et al. | Vitamin C |
| ↓ hospital length of stay; ↓ intubation time; ↓ drainage volume in the ICU; ↓ inflammatory factors; | [249] |
| Samadikhah et al. | Vitamin C + atorvastatin |
| ↓ atrial fibrillation incidence post coronary artery by-pass graft (CABG); | [250] |
| Dehghani et al. | Vitamin C |
| ↓ incidence postoperative atrial fibrillation; ↓ hospital stay; ↓ ICU stay; | [251] |
| Jouybar et al. | Vitamin C |
|
| [252] |
| Papoulidis et al. | Vitamin C |
| ↓ atrial fibrillation incidence; ↓ hospitalization time; ↓ ICU stay; | [253] |
| Collier et al. | Vitamin C + vitamin E + selenium |
| ↓ ICU stay; ↓ hospital stay; | [254] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ritiu, S.A.; Rogobete, A.F.; Sandesc, D.; Bedreag, O.H.; Papurica, M.; Popovici, S.E.; Toma, D.; Ivascu, R.I.; Velovan, R.; Garofil, D.N.; et al. The Impact of General Anesthesia on Redox Stability and Epigenetic Inflammation Pathways: Crosstalk on Perioperative Antioxidant Therapy. Cells 2022, 11, 1880. https://doi.org/10.3390/cells11121880
Ritiu SA, Rogobete AF, Sandesc D, Bedreag OH, Papurica M, Popovici SE, Toma D, Ivascu RI, Velovan R, Garofil DN, et al. The Impact of General Anesthesia on Redox Stability and Epigenetic Inflammation Pathways: Crosstalk on Perioperative Antioxidant Therapy. Cells. 2022; 11(12):1880. https://doi.org/10.3390/cells11121880
Chicago/Turabian StyleRitiu, Stelian Adrian, Alexandru Florin Rogobete, Dorel Sandesc, Ovidiu Horea Bedreag, Marius Papurica, Sonia Elena Popovici, Daiana Toma, Robert Iulian Ivascu, Raluca Velovan, Dragos Nicolae Garofil, and et al. 2022. "The Impact of General Anesthesia on Redox Stability and Epigenetic Inflammation Pathways: Crosstalk on Perioperative Antioxidant Therapy" Cells 11, no. 12: 1880. https://doi.org/10.3390/cells11121880