
Effect of Drought and Heavy Metal Contamination on Growth and Photosynthesis of Silver Birch Trees Growing on Post-Industrial Heaps

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
- Birches on the control site are characterised by better growth and physiological condition than birches growing on heaps. Many years of growth in extreme habitats should exclude the occurrence of the phenomenon of hormesis [18], while the cumulative toxic effect of environmental factors should translate into a worse physiological condition of trees, including reduced annual growth of wood.
- There are modifications in the photosynthesis mechanism, enabling the adaptation of birch trees to the extreme conditions of the habitat. Recent scientific work [19,20,21,22] indicates the role of non-photochemical quenching (NPQ) in the adaptation to drought conditions, which is a key stress factor in heaps.
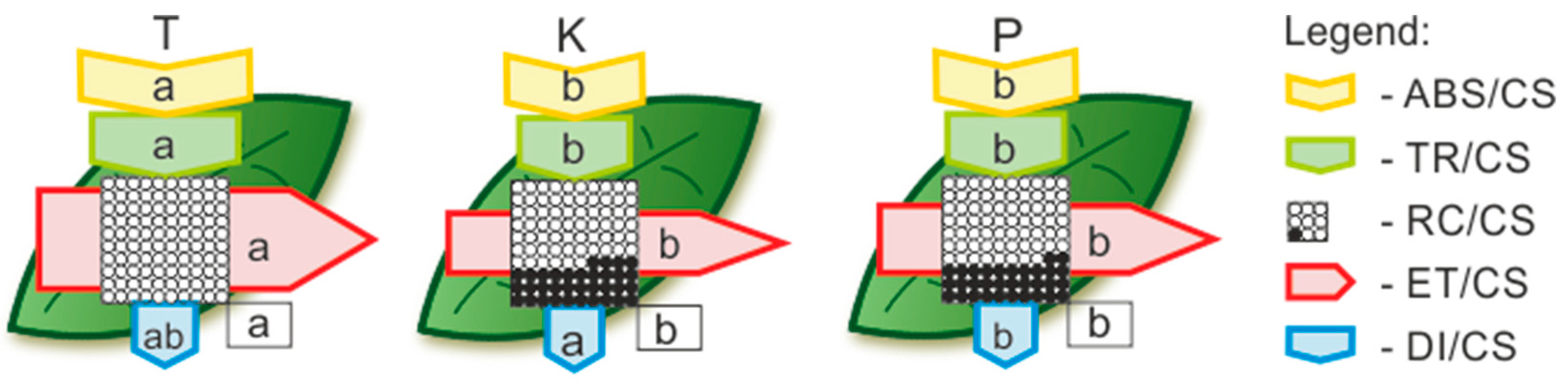
- It is possible to demonstrate physiological parameters, the modification of which enables birch trees to colonise extreme habitats as one of the first species.
2. Materials and Methods
2.1. Site Description
2.2. Plant Material and Sampling
2.3. Statistical Analysis
3. Results
3.1. Growth Parameters
3.2. Pigment Content and Gas Exchange Parameters
3.3. Element Accumulation in Plant Tissues
3.4. Chlorophyll Fluorescence Characteristics
3.5. Dendroclimatological Analysis
3.6. Determining the Correctness of Selection and Correlation between the Tested Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bussotti, F.; Pollastrini, M. Revisiting the concept of stress in forest trees at the time of global change and issues for stress monitoring. Plant Stress 2021, 2, 100013. [Google Scholar] [CrossRef]
- Aspelmeier, S.; Leuschner, C. Genotypic variation in drought response of silver birch (Betula pendula): Leaf water status and carbon gain. Tree Physiol. 2004, 24, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Franiel, I. The Biology and Ecology of Betula pendula Roth on Post-Industrial Waste Dumping Grounds: The Variability Range of Life History Traits; Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2012; ISBN 5555555555. [Google Scholar]
- Franiel, I.; Więski, K. Leaf features of silver birch (Betula pendula Roth). Variability within and between two populations (uncontaminated vs Pb-contaminated and Zn-contaminated site). Trees- Struct. Funct. 2005, 19, 81–88. [Google Scholar] [CrossRef]
- Marguí, E.; Queralt, I.; Carvalho, M.L.; Hidalgo, M. Assessment of metal availability to vegetation (Betula pendula) in Pb-Zn ore concentrate residues with different features. Environ. Pollut. 2007, 145, 179–184. [Google Scholar] [CrossRef]
- Czaja, M.; Kołton, A.; Baran, A.; Muszyńska, E.; Muras, P. Physiological responses of Betula pendula Roth growing in polluted areas. Ecol. Quest. 2015, 22, 39–46. [Google Scholar] [CrossRef]
- Kosiorek, M.; Modrzewska, B.; Wyszkowski, M. Levels of selected trace elements in Scots pine (Pinus sylvestris L.), silver birch (Betula pendula L.), and Norway maple (Acer platanoides L.) in an urbanized environment. Environ. Monit. Assess. 2016, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franiel, I.; Babczyńska, A. The growth and reproductive effort of Betula pendula Roth in a heavy-metals polluted area. Polish J. Environ. Stud. 2011, 20, 1097–1101. [Google Scholar]
- Alagic, S.Č.; Šerbula, S.S.; Tǒic, S.B.; Pavlović, A.N.; Petrovic, J.V. Bioaccumulation of arsenic and cadmium in birch and lime from the Bor region. Arch. Environ. Contam. Toxicol. 2013, 65, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Pełka-Gościniak, J. Plany zagospodarowania zwałowisk odpadów powęglowych KWK Murcki w Katowicach. Acta Geogr. Silesiana 2015, 18, 59–67. [Google Scholar]
- Fabiańska, M.J.; Ciesielczuk, J.; Kruszewski, Ł.; Misz-Kennan, M.; Blake, D.R.; Stracher, G.; Moszumańska, I. Gaseous compounds and efflorescences generated in self-heating coal-waste dumps—A case study from the Upper and Lower Silesian Coal Basins (Poland). Int. J. Coal Geol. 2013, 116–117, 247–261. [Google Scholar] [CrossRef]
- Misz-Kennan, M.; Fabiańska, M.J. Application of organic petrology and geochemistry to coal waste studies. Int. J. Coal Geol. 2011, 88, 1–23. [Google Scholar] [CrossRef]
- Ciesielczuk, J.; Misz-Kennan, M.; Hower, J.C.; Fabiańska, M.J. Mineralogy and geochemistry of coal wastes from the Starzykowiec coal-waste dump (Upper Silesia, Poland). Int. J. Coal Geol. 2014, 127, 42–55. [Google Scholar] [CrossRef]
- Skrȩt, U.; Fabiańska, M.J.; Misz-Kennan, M. Simulated water-washing of organic compounds from self-heated coal wastes of the Rymer Cones Dump (Upper Silesia Coal Region, Poland). Org. Geochem. 2010, 41, 1009–1012. [Google Scholar] [CrossRef]
- Kaja, S. Vascular flora of sites contaminated with heavy metals on the example of two post-industrial spoil heaps connected with manufacturing of zinc and lead products in upper silesia. Arch. Environ. Prot. 2011, 37, 57–74. [Google Scholar]
- Jędrzejczyk, M.; Bzdęga, K.; Rostański, A. Obecny stan zasobów introdukowanych fiołków galmanowych Viola guestphalica Nauenb. i Viola calaminaria (Ging.) Lej. na zwałowisku w Katowicach—Wełnowcu. Arch. Ochr. Środowiska 2003, 29, 21–30. [Google Scholar]
- Tyszka, R.; Kierczak, J.; Pietranik, A.; Ettler, V.; Mihaljevič, M. Extensive weathering of zinc smelting slag in a heap in Upper Silesia (Poland): Potential environmental risks posed by mechanical disturbance of slag deposits. Appl. Geochem. 2014, 40, 70–81. [Google Scholar] [CrossRef]
- Małkowski, E.; Sitko, K.; Szopiński, M.; Gieroń, Ż.; Pogrzeba, M.; Kalaji, H.M.; Zieleźnik-Rusinowska, P. Hormesis in Plants: The Role of Oxidative Stress, Auxins and Photosynthesis in Corn Treated with Cd or Pb. Int. J. Mol. Sci. 2020, 21, 2099. [Google Scholar] [CrossRef] [Green Version]
- Bano, H.; Athar, H.u.-R.; Zafar, Z.U.; Ogbaga, C.C.; Ashraf, M. Peroxidase activity and operation of photo-protective component of NPQ play key roles in drought tolerance of mung bean [Vigna radiata (L.) Wilcziek]. Physiol. Plant. 2021, 172, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.H.; Zhu, F.; He, Z.X.; Chen, X.Y.; Wang, G.J.; Liu, M.S.; Xu, H.Y. Photosynthesis Performance and Antioxidative Enzymes Response of Melia azedarach and Ligustrum lucidum Plants Under Pb–Zn Mine Tailing Conditions. Front. Plant Sci. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- D’Odorico, P.; Schönbeck, L.; Vitali, V.; Meusburger, K.; Schaub, M.; Ginzler, C.; Zweifel, R.; Velasco, V.M.E.; Gisler, J.; Gessler, A.; et al. Drone-based physiological index reveals long-term acclimation and drought stress responses in trees. Plant Cell Environ. 2021, 44, 3552–3570. [Google Scholar] [CrossRef]
- Bashir, N.; Athar, H.; Kalaji, H.M.; Wr, J.; Mahmood, S.; Zafar, Z.U.; Ashraf, M. Is Photoprotection of PSII One of the Key Mechanisms for Drought Tolerance in Maize? Int. J. Mol. Sci. 2021, 22, 13490. [Google Scholar] [CrossRef]
- Strzyszcz, Z.; Magiera, T.; Heller, F. The influence of industrial immissions on the magnetic susceptibility of soils in Upper Silesia. Stud. Geophys. Geod. 1996, 40, 276–286. [Google Scholar] [CrossRef]
- Leśniok, M.; Małarzewski, Ł.; Niedźwiedź, T. Classification of circulation types for Southern Poland with an application to air pollution concentration in Upper Silesia. Phys. Chem. Earth 2010, 35, 516–522. [Google Scholar] [CrossRef]
- Czaja, S. Mining and hydrological transformations in Upper Silesia from the fifteenth to the nineteenth century. Geogr. J. 2001, 167, 57–71. [Google Scholar] [CrossRef]
- Dulias, R. The Impact of Mining on the Landscape. A Study of the Upper Silesian Coal Basin in Poland; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-29539-8. [Google Scholar]
- Szopiński, M.; Sitko, K.; Rusinowski, S.; Zieleźnik-Rusinowska, P.; Corso, M.; Rostański, A.; Rojek-Jelonek, M.; Verbruggen, N.; Małkowski, E. Different strategies of Cd tolerance and accumulation in Arabidopsis halleri and Arabidopsis arenosa. Plant Cell Environ. 2020, 43, 3002–3019. [Google Scholar] [CrossRef]
- Bury, M.; Rusinowski, S.; Sitko, K.; Krzyżak, J.; Kitczak, T.; Możdżer, E.; Siwek, H.; Włodarczyk, M.; Zieleźnik-Rusinowska, P.; Szada-Borzyszkowska, A.; et al. Physiological status and biomass yield of Sida hermaphrodita (L.) Rusby cultivated on two distinct marginal lands in Southern and Northern Poland. Ind. Crops Prod. 2021, 167, 113502. [Google Scholar] [CrossRef]
- Cook, E.; Kairiukstis, L. Methods of Dendrochronology: Applications in the Environmental Sciences; Springer: New York, NY, USA, 1990; ISBN 978-90-481-4060-2. [Google Scholar]
- Holmes, R. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-ring Bull. 1983, 43, 51–67. [Google Scholar]
- Kitao, M.; Lei, T.T.; Koike, T.; Tobita, H.; Maruyama, Y. Higher electron transport rate observed at low intercellular CO2 concentration in long-term drought-acclimated leaves of Japanese mountain birch (Betula ermanii). Physiol. Plant. 2003, 118, 406–413. [Google Scholar] [CrossRef]
- Opała, M. The 443-Year Tree-Ring Chronology for the Scots Pine from Upper Silesia (Poland) as A Dating Tool and Climate Proxy. Geochronometria 2015, 42, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Sensuła, B.; Opała, M.; Wilczyński, S.; Pawełczyk, S. Long- and short-term incremental response of Pinus sylvestris L. from industrial area nearby steelworks in Silesian Upland, Poland. Dendrochronologia 2015, 36, 1–12. [Google Scholar] [CrossRef]
- Franiel, I.; Kompała-Bąba, A. Reproduction strategies of the silver birch (Betula pendula Roth) at post-industrial sites. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Wierzbicka, M.; Pielichowska, M. Adaptation of Biscutella laevigata L, a metal hyperaccumulator, to growth on a zinc-lead waste heap in southern Poland: I—Differences between waste-heap and mountain populations. Chemosphere 2004, 54, 1663–1674. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC press: Boca Raton, FL, USA, 2010; ISBN 9781420093704. [Google Scholar]
- Li, J.; Jia, Y.; Dong, R.; Huang, R.; Liu, P.; Li, X.; Wang, Z.; Liu, G.; Chen, Z. Advances in the mechanisms of plant tolerance to manganese toxicity. Int. J. Mol. Sci. 2019, 20, 5096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimann, C.; Arnoldussen, A.; Finne, T.E.; Koller, F.; Nordgulen, Ø.; Englmaier, P. Element contents in mountain birch leaves, bark and wood under different anthropogenic and geogenic conditions. Appl. Geochem. 2007, 22, 1549–1566. [Google Scholar] [CrossRef]
- Kitao, M.; Lei, T.T.; Nakamura, T.; Koike, T. Manganese toxicity as indicated by visible foliar symptoms of Japanese white birch (Betula platyphylla var. japonica). Environ. Pollut. 2001, 111, 89–94. [Google Scholar] [CrossRef]
- Sitko, K.; Rusinowski, S.; Kalaji, H.M.; Szopiński, M.; Małkowski, E. Photosynthetic efficiency as bioindicator of environmental pressure in A. halleri. Plant Physiol. 2017, 175, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusinowski, S.; Krzyżak, J.; Sitko, K.; Kalaji, H.M.; Jensen, E.; Pogrzeba, M. Cultivation of C4 perennial energy grasses on heavy metal contaminated arable land: Impact on soil, biomass, and photosynthetic traits. Environ. Pollut. 2019, 250, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Pilarska, M.; Niewiadomska, E.; Sychta, K.; Słomka, A. Differences in the functioning of photosynthetic electron transport between metallicolous and non-metallicolous populations of the pseudometallophyte Viola tricolor. J. Plant Physiol. 2020, 250. [Google Scholar] [CrossRef]
- Kitao, M.; Lei, T.T.; Koike, T.; Kayama, M.; Tobita, H.; Maruyama, Y. Interaction of drought and elevated CO2 concentration on photosynthetic down-regulation and susceptibility to photoinhibition in Japanese white birch seedlings grown with limited N availability. Tree Physiol. 2007, 27, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Pšidová, E.; Ditmarová, Ľ.; Jamnická, G.; Kurjak, D.; Majerová, J.; Czajkowski, T.; Bolte, A. Photosynthetic response of beech seedlings of different origin to water deficit. Photosynthetica 2015, 53, 187–194. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Račková, L.; Paganová, V.; Swoczyna, T.; Rusinowski, S.; Sitko, K. Can chlorophyll-a fluorescence parameters be used as bio-indicators to distinguish between drought and salinity stress in Tilia cordata Mill? Environ. Exp. Bot. 2018, 152, 149–157. [Google Scholar] [CrossRef]
- Szopiński, M.; Sitko, K.; Gieroń, Ż.; Rusinowski, S.; Corso, M.; Hermans, C.; Verbruggen, N.; Małkowski, E. Toxic effects of cd and zn on the photosynthetic apparatus of the Arabidopsis halleri and Arabidopsis arenosa pseudo-metallophytes. Front. Plant Sci. 2019, 10, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gieroń, Ż.; Sitko, K.; Zieleźnik-Rusinowska, P.; Szopiński, M.; Rojek-Jelonek, M.; Rostański, A.; Rudnicka, M.; Małkowski, E. Ecophysiology of Arabidopsis arenosa, a new hyperaccumulator of Cd and Zn. J. Hazard. Mater. 2021, 412, 125052. [Google Scholar] [CrossRef] [PubMed]
- Gallé, A.; Feller, U. Changes of photosynthetic traits in beech saplings (Fagus sylvatica) under severe drought stress and during recovery. Physiol. Plant. 2007, 131, 412–421. [Google Scholar] [CrossRef]
- Lamoreaux, R.J.; Chaney, W.R. The Effect of Cadmium on Net Photosynthesis, Transpiration, and Dark Respiration of Excised Silver Maple Leaves. Physiol. Plant. 1978, 43, 231–236. [Google Scholar] [CrossRef]
- Salisbury, A.B.; Gallagher, F.J.; Caplan, J.S.; Grabosky, J.C. Maintenance of photosynthesis by Betula populifolia in metal contaminated soils. Sci. Total Environ. 2018, 625, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Bussotti, F.; Gerosa, G.; Digrado, A.; Pollastrini, M. Selection of chlorophyll fluorescence parameters as indicators of photosynthetic efficiency in large scale plant ecological studies. Ecol. Indic. 2020, 108, 105686. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T | K | P | |
|---|---|---|---|
| Growth and Physiological Characteristics | |||
| Leaf thickness (mm) | 0.08 ± 0.01 c | 0.31 ± 0.01 a | 0.20 ± 0.03 b |
| TRW5 (mm) | 3.10 ± 0.b | 3.77 ± 0.a | 2.21 ± 0.c |
| TRWT (mm) | 3.06 ± 0.b | 3.83 ± 0.a | 2.35 ± 0.c |
| Chlorophyll content (r.u.) | 35.6 ± 0.9 a | 31.04 ± 1.25 b | 28.22 ± 0.71 c |
| Flavonol content (r.u.) | 1.38 ± 0.04 c | 1.76 ± 0.02 a | 1.67 ± 0.03 b |
| Anthocyanin content (r.u.) | 0.110 ± 0.002 b | 0.137 ± 0.005 a | 0.141 ± 0.003 a |
| A (μmol CO2 m−2 s−1) | 13.8 ± 0.4 a | 11.0 ± 0.5 b | 9.5 ± 0.3 c |
| Ci (μmol CO2 mol−1) | 244 ± 5 b | 257 ± 3 b | 280 ± 5 a |
| E (mmol H2O m−2 s−1) | 2.98 ± 0.15 a | 2.20 ± 0.07 b | 1.81 ± 0.05 c |
| gs (μmol H2O m−2 s−1) | 205 ± 13 a | 208 ± 13 a | 202 ± 7 a |
| Element Accumulation in Leaves (μg g−1 DW) | |||
| Ca | 10,200 ± 560 a | 7520 ± 610 b | 11,580 ± 1050 a |
| Cd | BDL | BDL | 1.43 ± 0.19 a |
| Cu | 5.4 ± 0.4 a | 3.0 ± 0.4 b | 2.0 ± 0.3 b |
| Fe | 86 ± 5 a | 82 ± 6 a | 89 ± 6 a |
| K | 5960 ± 460 a | 4100 ± 420 b | 3720 ± 270 b |
| Mg | 2200 ± 110 c | 2910 ± 180 b | 3980 ± 180 a |
| Mn | 290 ± 90 b | 3180 ± 130 a | 110 ± 10 b |
| Zn | 195 ± 27 b | 148 ± 8 b | 550 ± 61 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sitko, K.; Opała-Owczarek, M.; Jemioła, G.; Gieroń, Ż.; Szopiński, M.; Owczarek, P.; Rudnicka, M.; Małkowski, E. Effect of Drought and Heavy Metal Contamination on Growth and Photosynthesis of Silver Birch Trees Growing on Post-Industrial Heaps. Cells 2022, 11, 53. https://doi.org/10.3390/cells11010053
Sitko K, Opała-Owczarek M, Jemioła G, Gieroń Ż, Szopiński M, Owczarek P, Rudnicka M, Małkowski E. Effect of Drought and Heavy Metal Contamination on Growth and Photosynthesis of Silver Birch Trees Growing on Post-Industrial Heaps. Cells. 2022; 11(1):53. https://doi.org/10.3390/cells11010053
Chicago/Turabian StyleSitko, Krzysztof, Magdalena Opała-Owczarek, Gabriela Jemioła, Żaneta Gieroń, Michał Szopiński, Piotr Owczarek, Małgorzata Rudnicka, and Eugeniusz Małkowski. 2022. "Effect of Drought and Heavy Metal Contamination on Growth and Photosynthesis of Silver Birch Trees Growing on Post-Industrial Heaps" Cells 11, no. 1: 53. https://doi.org/10.3390/cells11010053