
The Flagellin:Allergen Fusion Protein rFlaA:Betv1 Induces a MyD88− and MAPK-Dependent Activation of Glucose Metabolism in Macrophages

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Recombinant Proteins
2.2. Determination of Beta-Glucans
2.3. Mice
2.4. In Vitro Generation of Mouse Bone Marrow-Derived Macrophages, Stimulation, and Flow Cytometry
2.5. Preparation of BMDM:CD4+ T Cell Co-Cultures
2.6. Inhibitors
2.7. Western Blotting
2.8. Analysis of Cell Metabolic State
2.9. Metabolic Flux Analysis
2.10. RNA-Seq and Bioinformatics
2.11. Statistical Analysis
3. Results
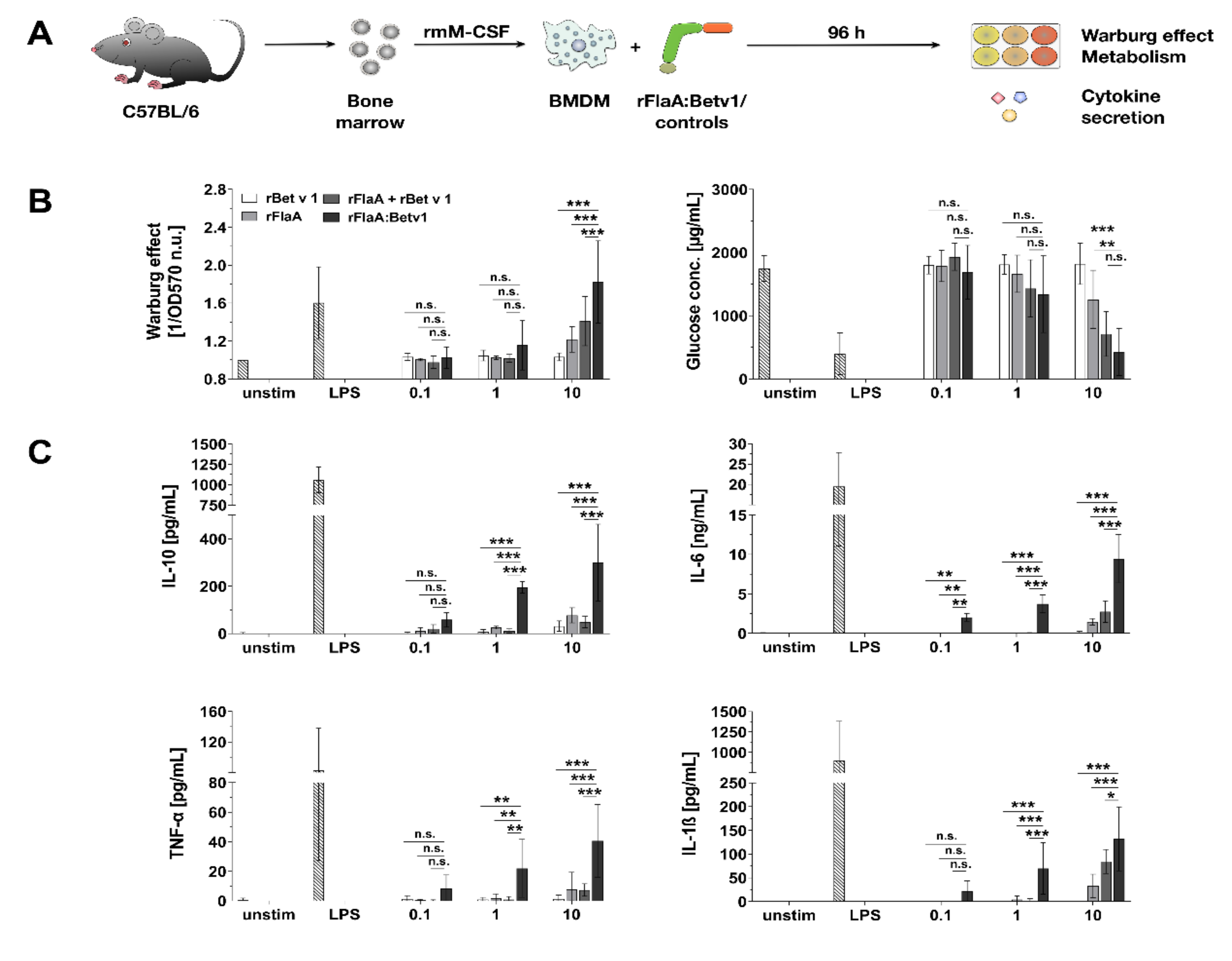
3.1. Activation of BMDM Metabolism by rFlaA:Betv1 Is Accompanied by Both Pro- and Anti-inflammatory Cytokine Secretion
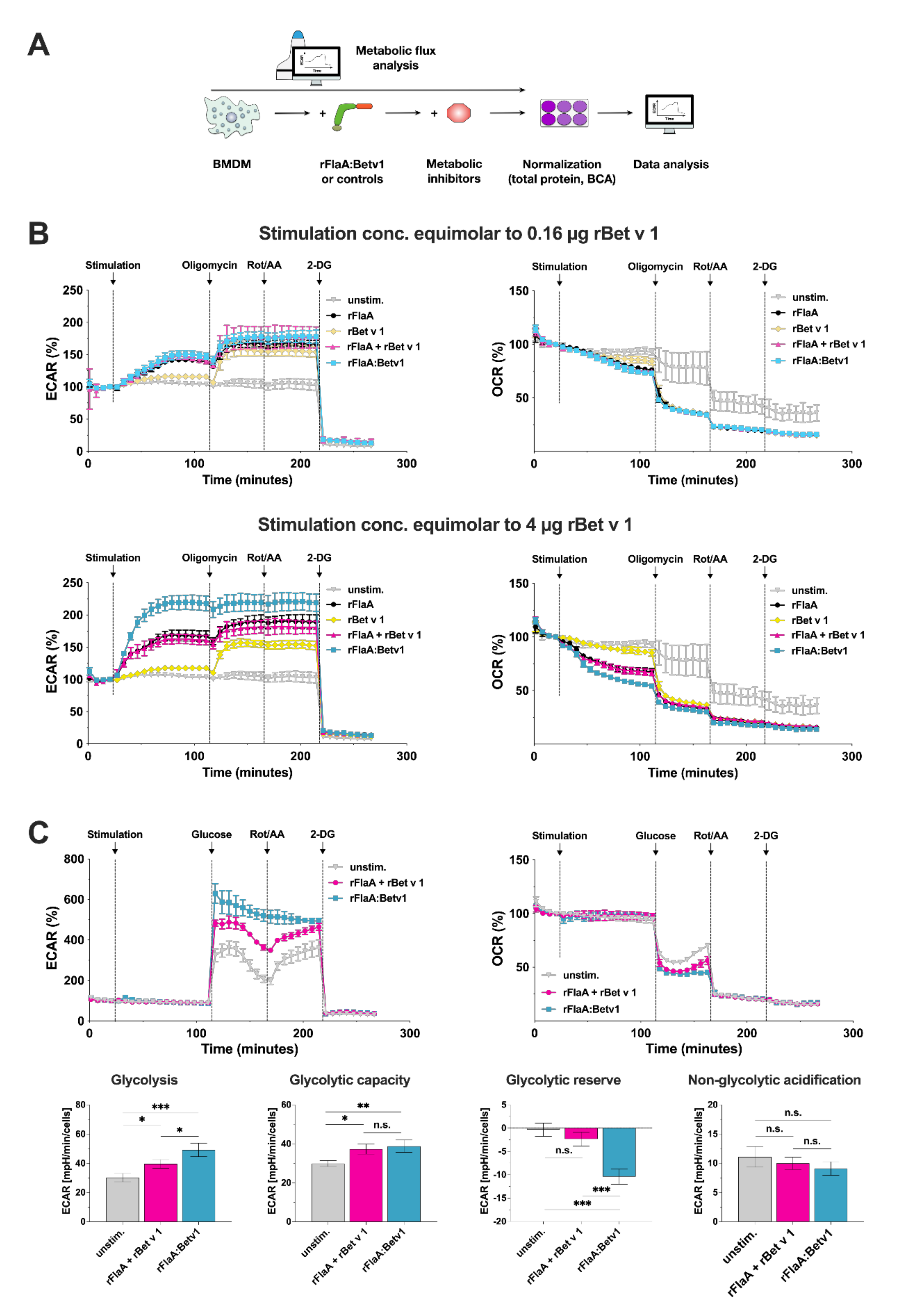
3.2. rFlaA:Betv1 Triggers a Pronounced Shift Towards Glycolysis in BMDMs
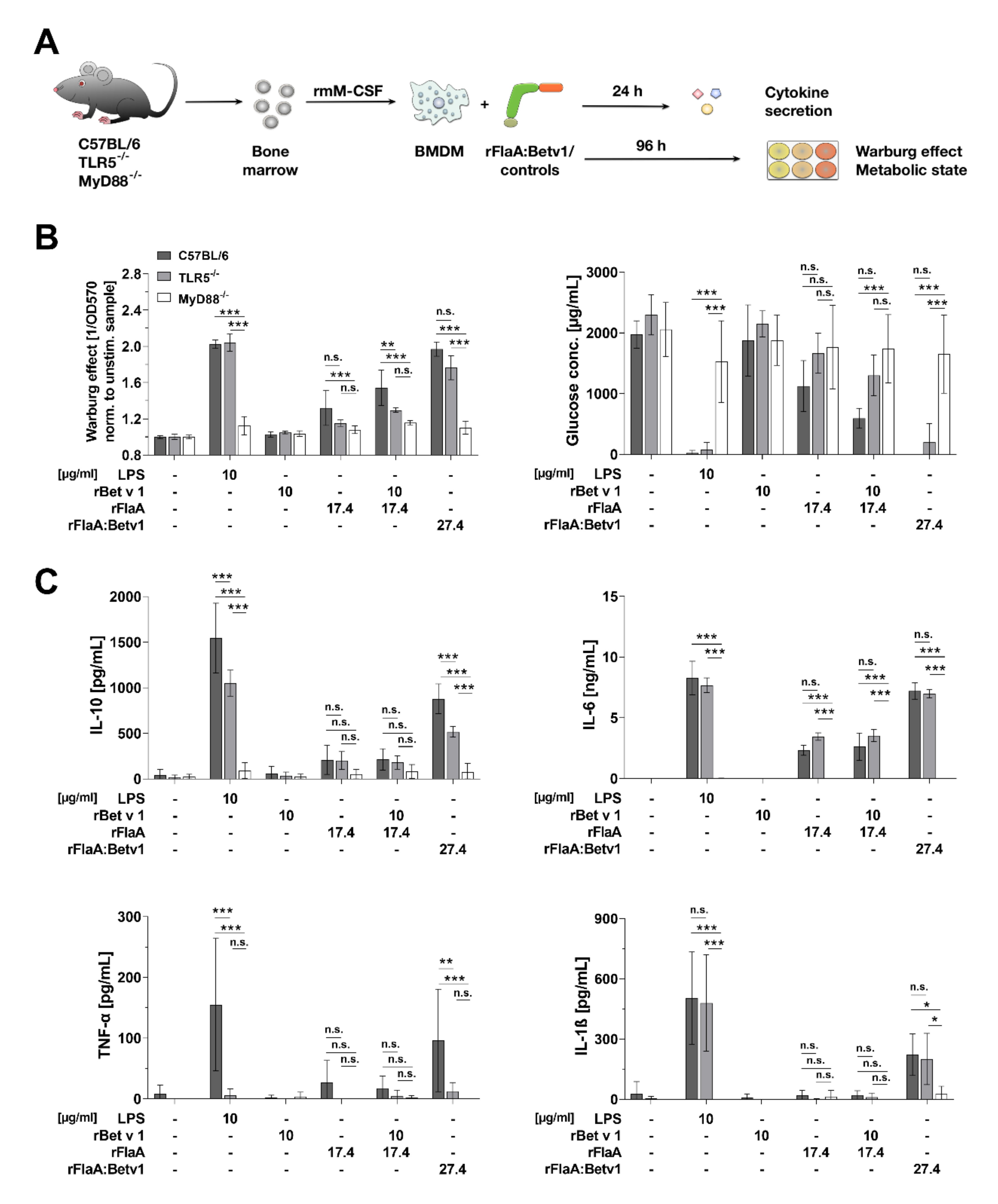
3.3. rFlaA:Betv1-Induced BMDM Metabolism and Inflammatory Cytokine Secretion Depend on MyD88 While Only Being Partially TLR5-Dependent
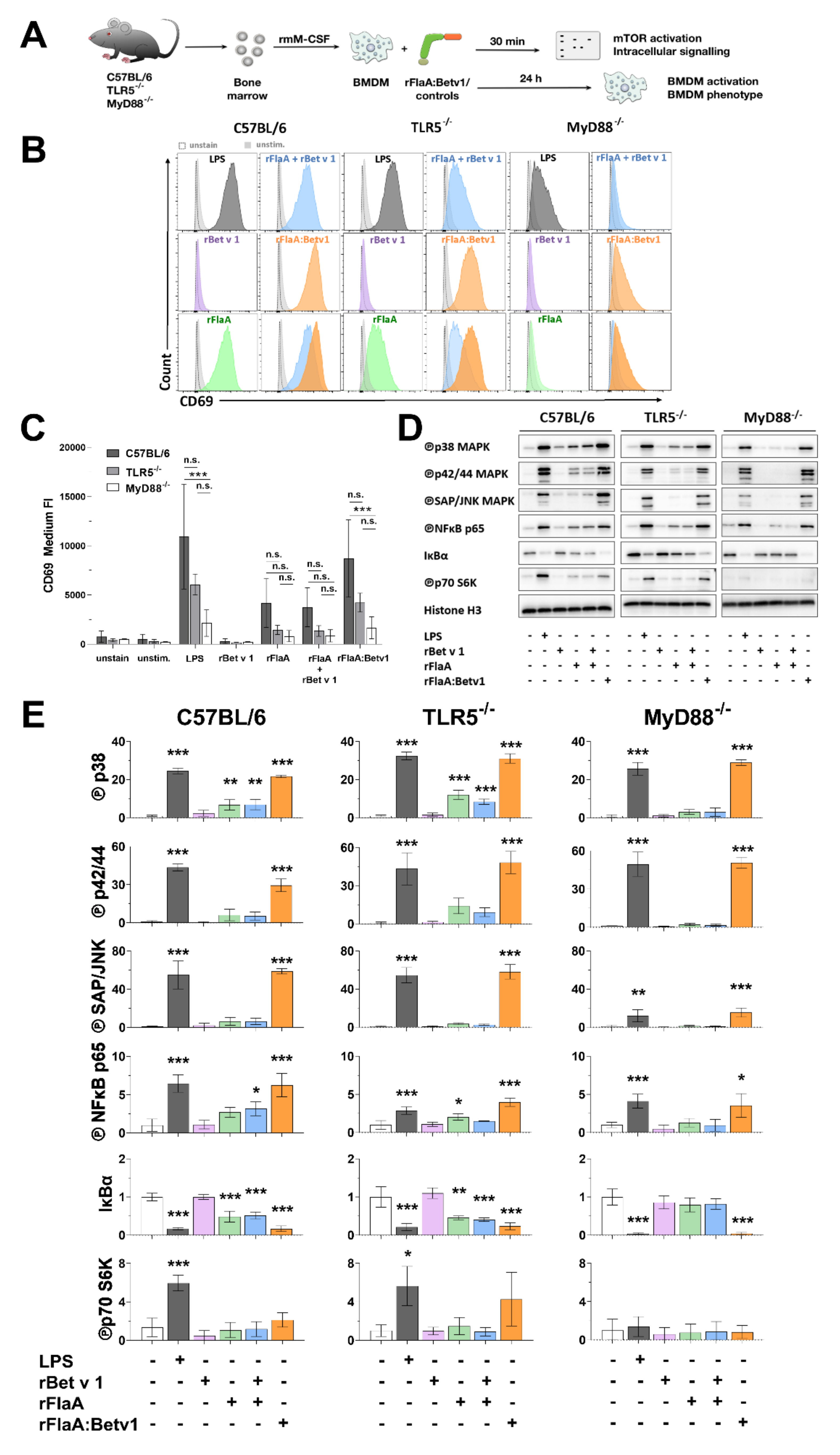
3.4. rFlaA:Betv1 Induces MAPK, NFκB, and mTOR Signaling in BMDMs
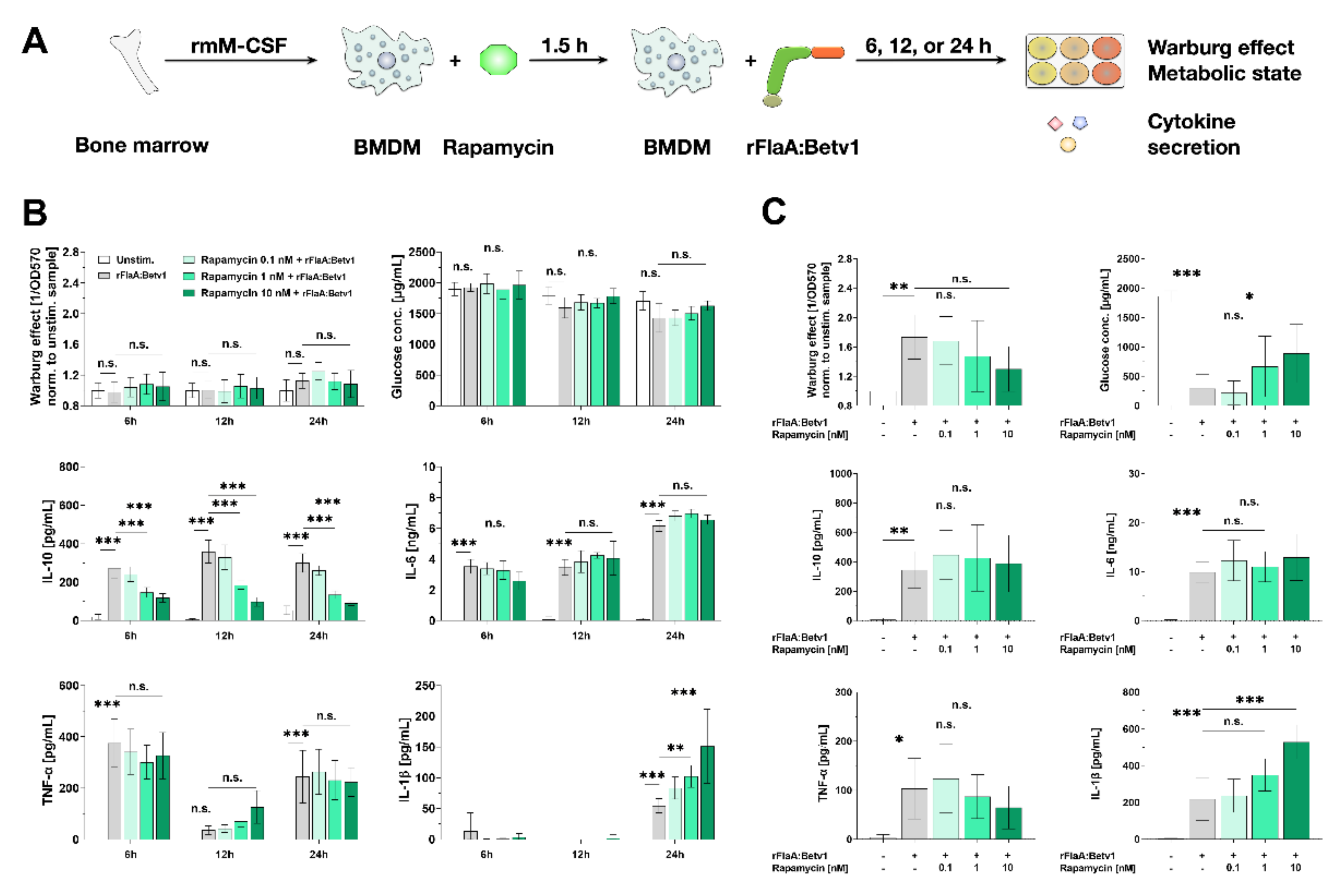
3.5. Activation of BMDM Metabolism and IL-10 Secretion are Partly mTOR-Dependent while Pro-Inflammatory Cytokine Secretion Is mTOR-Independent
3.6. MAP Kinase Signaling Contributes to rFlaA:Betv1-Induced Activation of BMDM Metabolism as well as Both Pro- and Anti-Inflammatory Cytokine Secretion
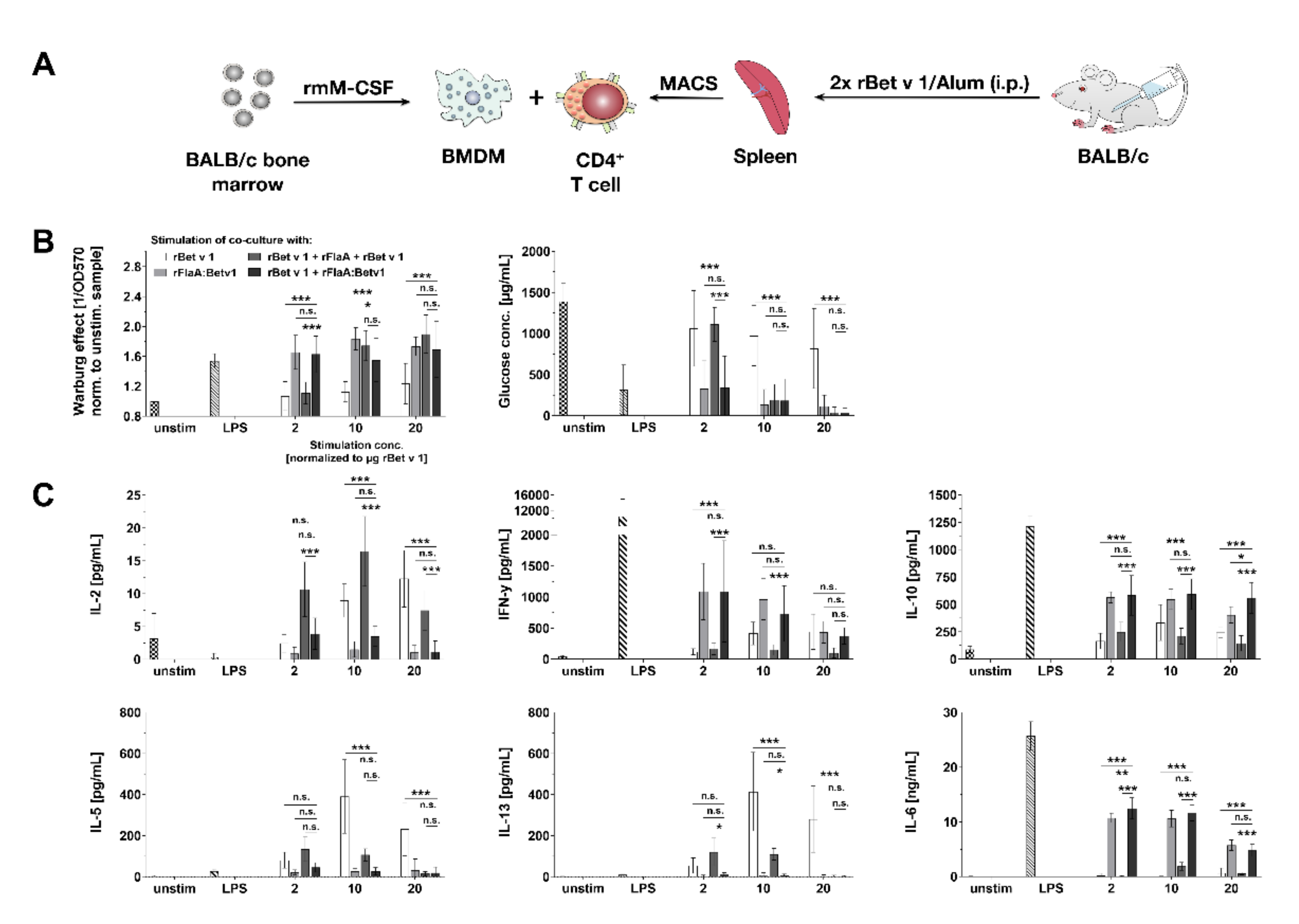
3.7. rFlaA:Betv1 Suppresses Th2 Responses from Bet v 1-specific CD4+ T Cells In Vitro
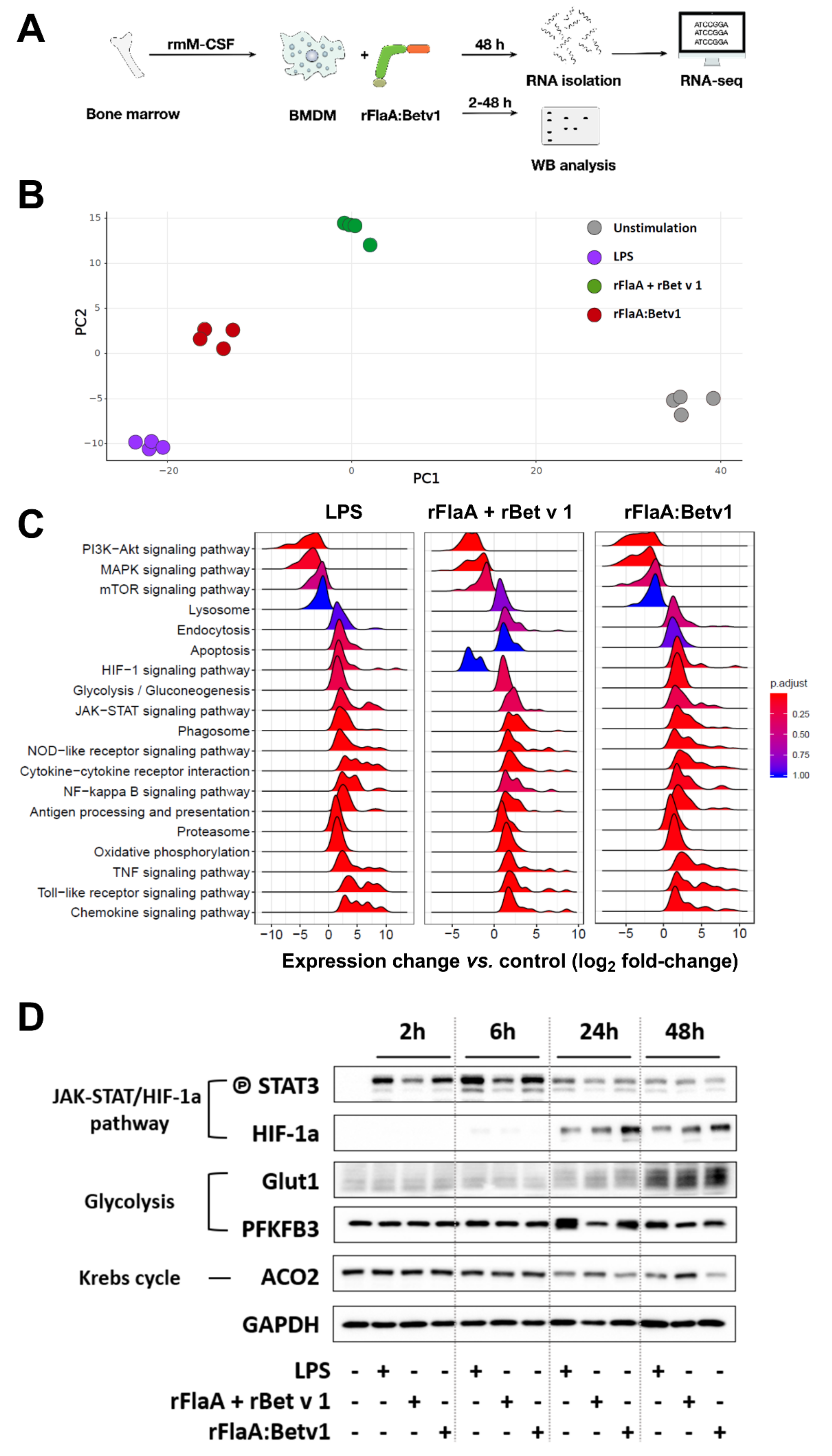
3.8. rFlaA:Betv1 Induces a Transcriptional Shift Towards HIF-1a-Mediated Glycolytic Metabolism in BMDMs
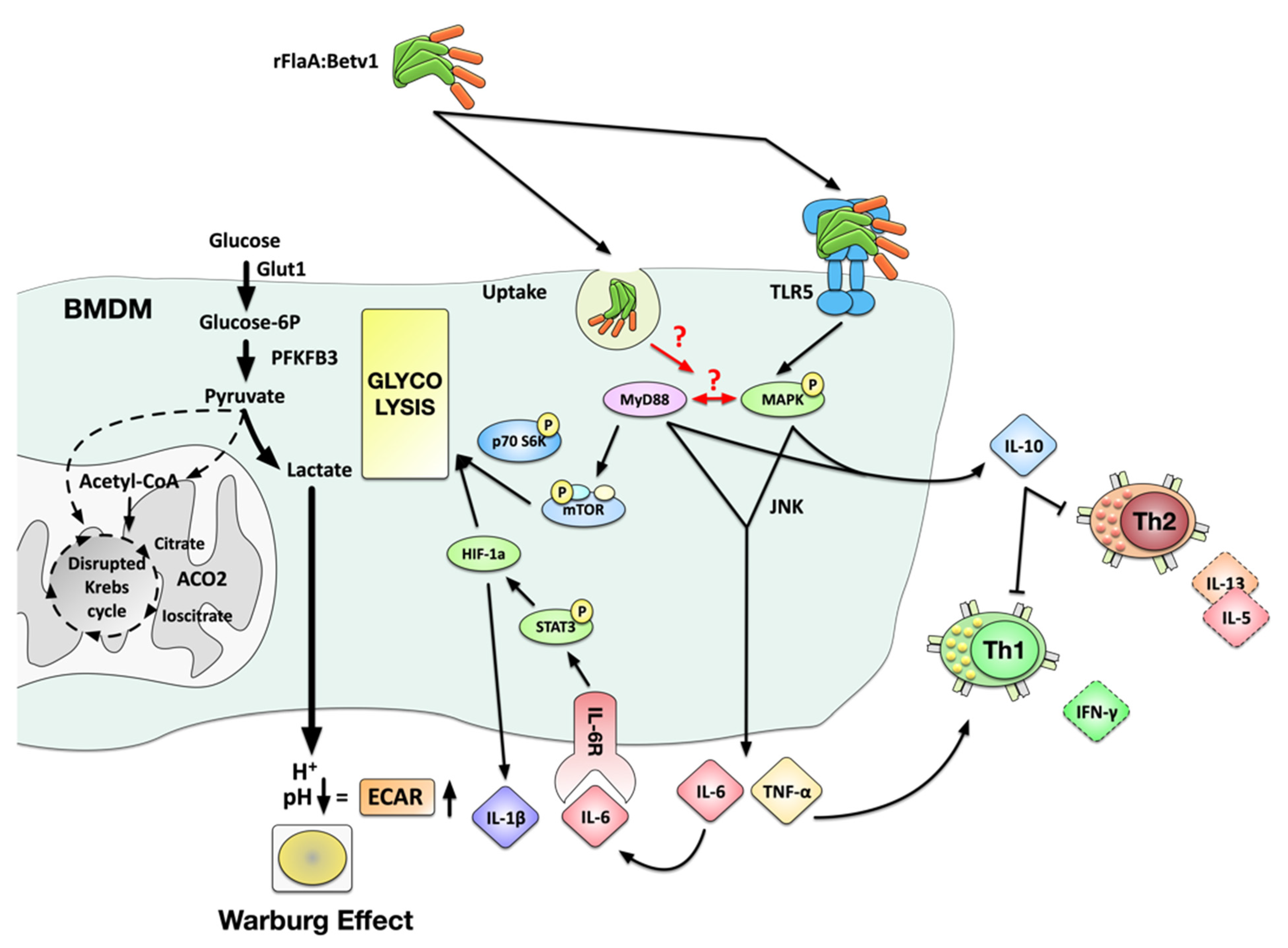
4. Discussion
4.1. rFlaA:Betv1 Induces a Partly TLR5-Dependent Activation of BMDMs Characterized by Both Pro- and Anti-Inflammatory Cytokine Secretion
4.2. rFlaA:Betv1-Stimulated BMDMs Efficiently Modulate Allergen-Specific T Cell Responses
4.3. rFlaA:Betv1 Activates Both Glycolytic Metabolism and HIF-1a Signaling in BMDMs
4.4. MAPK, NFκB, and mTOR Signaling Contribute to rFlaA:Betv1-Mediated BMDM Activation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-DG | 2-deoxy-glucose |
| ACO2 | Aconitase 2 |
| BMDM | Bone marrow-derived macrophage |
| CD | Cluster of differentiation |
| C/EBPβ | CCAAT enhancer-binding protein |
| ECAR | Extracellular acidification rate |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IFN-γ | Interferon gamma |
| IL | Interleukin |
| Ipaf | ICE-protease activating factor |
| MAP(K) | Mitogen-activated protein (kinase) |
| (m)DC | (Myeloid) Dendritic cell |
| mTOR | Mammalian target of rapamycin |
| mTORC1 | mTOR complex 1 |
| MyD88 | Myeloid differentiation primary response 88 |
| NFκB | Nuclear factor ‘kappa-light-chain-enhancer’ of activated B-cells |
| OCR | Oxygen consumption rate |
| PI3K | Phosphatidylinositol 3 kinase |
| PDK:1 | Pyruvate dehydrogenase kinase 1 |
| PFK | Phospho-fructo-kinase |
| PFKFB3 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 |
| PGM | Phosphoglycerate mutase |
| PKM | Pyruvate kinase isozyme |
| rBet v 1 | Recombinant major birch pollen allergen number one from Betula verrucosa |
| rFlaA | Recombinant Listeria monocytogenes flagellin A |
| rFlaA:Betv1 | Recombinant fusion protein of FlaA and Bet v 1 |
| rmM-CSF | Recombinant mouse macrophage colony-stimulating factor |
| RAPTOR | Regulatory associated protein of mTOR |
| TSC | Tuberous sclerosis complex |
| Rot/AA | Rotenone/ antimycin A |
| SAP/JNK MAPK | Stress-activated protein kinase/c-Jun NH2-terminal kinase |
| Th1/2 | T helper 1/2 cell |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor alpha |
References
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The Innate Immune Response to Bacterial Flagellin Is Mediated by Toll-like Receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Lee, S.E.; Kim, S.Y.; Jeong, B.C.; Kim, Y.R.; Bae, S.J.; Ahn, O.S.; Lee, J.J.; Song, H.C.; Kim, J.M.; Choy, H.E.; et al. A Bacterial Flagellin, Vibrio Vulnificus FlaB, Has a Strong Mucosal Adjuvant Activity to Induce Protective Immunity. Infect. Immun. 2006, 74, 694–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honko, A.N.; Sriranganathan, N.; Lees, C.J.; Mizel, S.B. Flagellin Is an Effective Adjuvant for Immunization against Lethal Respiratory Challenge with Yersinia Pestis. Infect. Immun. 2006, 74, 1113–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huleatt, J.W.; Jacobs, A.R.; Tang, J.; Desai, P.; Kopp, E.B.; Huang, Y.; Song, L.; Nakaar, V.; Powell, T.J. Vaccination with Recombinant Fusion Proteins Incorporating Toll-like Receptor Ligands Induces Rapid Cellular and Humoral Immunity. Vaccine 2007, 25, 763–775. [Google Scholar] [CrossRef]
- Turley, C.B.; Rupp, R.E.; Johnson, C.; Taylor, D.N.; Wolfson, J.; Tussey, L.; Kavita, U.; Stanberry, L.; Shaw, A. Safety and Immunogenicity of a Recombinant M2e-Flagellin Influenza Vaccine (STF2.4xM2e) in Healthy Adults. Vaccine 2011, 29, 5145–5152. [Google Scholar] [CrossRef]
- Treanor, J.J.; Taylor, D.N.; Tussey, L.; Hay, C.; Nolan, C.; Fitzgerald, T.; Liu, G.; Kavita, U.; Song, L.; Dark, I.; et al. Safety and Immunogenicity of a Recombinant Hemagglutinin Influenza-Flagellin Fusion Vaccine (VAX125) in Healthy Young Adults. Vaccine 2010, 28, 8268–8274. [Google Scholar] [CrossRef]
- Song, L.; Xiong, D.; Kang, X.; Yang, Y.; Wang, J.; Guo, Y.; Xu, H.; Chen, S.; Peng, D.; Pan, Z.; et al. An Avian Influenza A (H7N9) Virus Vaccine Candidate Based on the Fusion Protein of Hemagglutinin Globular Head and Salmonella Typhimurium Flagellin. BMC Biotechnol. 2015, 15, 79. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, L.A.; Kotlyarov, R.Y.; Kovaleva, A.A.; Potapchuk, M.V.; Korotkov, A.V.; Sergeeva, M.V.; Kasianenko, M.A.; Kuprianov, V.V.; Ravin, N.V.; Tsybalova, L.M.; et al. Protection against Multiple Influenza A Virus Strains Induced by Candidate Recombinant Vaccine Based on Heterologous M2e Peptides Linked to Flagellin. PLoS ONE 2015, 10, e0119520. [Google Scholar] [CrossRef]
- Wang, B.-Z.; Gill, H.S.; He, C.; Ou, C.; Wang, L.; Wang, Y.-C.; Feng, H.; Zhang, H.; Prausnitz, M.R.; Compans, R.W. Microneedle Delivery of an M2e-TLR5 Ligand Fusion Protein to Skin Confers Broadly Cross-Protective Influenza Immunity. J. Control Release 2014, 178, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Delaney, K.N.; Phipps, J.P.; Johnson, J.B.; Mizel, S.B. A Recombinant Flagellin-Poxvirus Fusion Protein Vaccine Elicits Complement-Dependent Protection against Respiratory Challenge with Vaccinia Virus in Mice. Viral Immunol. 2010, 23, 201–210. [Google Scholar] [CrossRef] [PubMed]
- McDonald, W.F.; Huleatt, J.W.; Foellmer, H.G.; Hewitt, D.; Tang, J.; Desai, P.; Price, A.; Jacobs, A.; Takahashi, V.N.; Huang, Y.; et al. A West Nile Virus Recombinant Protein Vaccine That Coactivates Innate and Adaptive Immunity. J. Infect. Dis. 2007, 195, 1607–1617. [Google Scholar] [CrossRef]
- Lee, S.E.; Nguyen, C.T.; Kim, S.Y.; Thi, T.N.; Rhee, J.H. Tetanus Toxin Fragment C Fused to Flagellin Makes a Potent Mucosal Vaccine. Clin. Exp. Vaccine Res. 2015, 4, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Weimer, E.T.; Lu, H.; Kock, N.D.; Wozniak, D.J.; Mizel, S.B. A Fusion Protein Vaccine Containing OprF Epitope 8, OprI, and Type A and B Flagellins Promotes Enhanced Clearance of Nonmucoid Pseudomonas Aeruginosa. Infect. Immun. 2009, 77, 2356–2366. [Google Scholar] [CrossRef] [Green Version]
- Kitzmüller, C.; Kalser, J.; Mutschlechner, S.; Hauser, M.; Zlabinger, G.J.; Ferreira, F.; Bohle, B. Fusion Proteins of Flagellin and the Major Birch Pollen Allergen Bet v 1 Show Enhanced Immunogenicity, Reduced Allergenicity, and Intrinsic Adjuvanticity. J. Allergy Clin. Immunol. 2018, 141, 293–299.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schülke, S.; Fiedler, A.-H.; Ann-Christine, J.; Flaczyk, A.; Wolfheimer, S.; Anke, H.; Hendrik, B.; Birgit, N.; Bohle, B.; Vieths, S.; et al. Critical Role of Mammalian Target of Rapamycin for IL-10 DC Induction by a Flagellin FlaA-Conjugate Preventing Allergic Sensitization. J. Allergy Clin. Immunol. 2018, 141, 1786–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schülke, S.; Burggraf, M.; Waibler, Z.; Wangorsch, A.; Wolfheimer, S.; Kalinke, U.; Vieths, S.; Toda, M.; Scheurer, S. A Fusion Protein of Flagellin and Ovalbumin Suppresses the TH2 Response and Prevents Murine Intestinal Allergy. J. Allergy Clin. Immunol. 2011, 128, 1340–1348.e12. [Google Scholar] [CrossRef]
- Schülke, S.; Kuttich, K.; Wolfheimer, S.; Duschek, N.; Wangorsch, A.; Reuter, A.; Briza, P.; Pablos, I.; Gadermaier, G.; Ferreira, F.; et al. Author Correction: Conjugation of Wildtype and Hypoallergenic Mugwort Allergen Art v 1 to Flagellin Induces IL-10-DC and Suppresses Allergen-Specific TH2-Responses in Vivo. Sci. Rep. 2018, 8, 2745. [Google Scholar] [CrossRef]
- Schülke, S.; Wolfheimer, S.; Gadermaier, G.; Wangorsch, A.; Siebeneicher, S.; Briza, P.; Spreitzer, I.; Schiller, D.; Loeschner, B.; Uematsu, S.; et al. Prevention of Intestinal Allergy in Mice by RflaA:Ova Is Associated with Enforced Antigen Processing and TLR5-Dependent IL-10 Secretion by MDC. PLoS ONE 2014, 9, e87822. [Google Scholar] [CrossRef]
- Kay, A.B. Immunomodulation in Asthma: Mechanisms and Possible Pitfalls. Curr. Opin. Pharmacol. 2003, 3, 220–226. [Google Scholar] [CrossRef]
- Koya, T.; Matsuda, H.; Takeda, K.; Matsubara, S.; Miyahara, N.; Balhorn, A.; Dakhama, A.; Gelfand, E.W. IL-10–Treated Dendritic Cells Decrease Airway Hyperresponsiveness and Airway Inflammation in Mice. J. Allergy Clin. Immunol. 2007, 119, 1241–1250. [Google Scholar] [CrossRef]
- Gentile, D.A.; Patel, A.; Ollila, C.; Fireman, P.; Zeevi, A.; Doyle, W.J.; Skoner, D.P. Diminished IL-10 Production in Subjects with Allergy after Infection with Influenza A Virus. J. Allergy Clin. Immunol. 1999, 103, 1045–1048. [Google Scholar] [CrossRef]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage Plasticity and Polarization in Tissue Repair and Remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, X.-H.; Jin, L. Macrophage Polarization in Physiological and Pathological Pregnancy. Front. Immunol. 2019, 10, 792. [Google Scholar] [CrossRef] [PubMed]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.S. Macrophages: Time to Take Notice. Ann. Allergy Asthma Immunol. 2019, 123, 229. [Google Scholar] [CrossRef] [PubMed]
- Saradna, A.; Do, D.C.; Kumar, S.; Fu, Q.-L.; Gao, P. Macrophage Polarization and Allergic Asthma. Transl. Res. 2018, 191, 1–14. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhu, L. Update on the Role of Alternatively Activated Macrophages in Asthma. J. Asthma Allergy 2016, 9, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, N.; Terawaki, S.; Shimizu, K.; Oikawa, D.; Sakamoto, H.; Sunami, K.; Tokunaga, F. Th2 Cells and Macrophages Cooperatively Induce Allergic Inflammation through Histamine Signaling. PLoS ONE 2021, 16, e0248158. [Google Scholar] [CrossRef] [PubMed]
- Robbe, P.; Draijer, C.; Borg, T.R.; Luinge, M.; Timens, W.; Wouters, I.M.; Melgert, B.N.; Hylkema, M.N. Distinct Macrophage Phenotypes in Allergic and Nonallergic Lung Inflammation. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 308, L358–L367. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.; Secor, E.R.; Silbart, L.K. Broncho-Alveolar Macrophages Express Chemokines Associated with Leukocyte Migration in a Mouse Model of Asthma. Cell Immunol. 2013, 281, 159–169. [Google Scholar] [CrossRef]
- Schülke, S.; Waibler, Z.; Mende, M.-S.; Zoccatelli, G.; Vieths, S.; Toda, M.; Scheurer, S. Fusion Protein of TLR5-Ligand and Allergen Potentiates Activation and IL-10 Secretion in Murine Myeloid DC. Mol. Immunol. 2010, 48, 341–350. [Google Scholar] [CrossRef]
- Siebeneicher, S.; Reuter, S.; Krause, M.; Wangorsch, A.; Maxeiner, J.; Wolfheimer, S.; Schülke, S.; Naito, S.; Heinz, A.; Taube, C.; et al. Epicutaneous Immune Modulation with Bet v 1 plus R848 Suppresses Allergic Asthma in a Murine Model. Allergy 2014, 69, 328–337. [Google Scholar] [CrossRef]
- Levin, J.Z.; Yassour, M.; Adiconis, X.; Nusbaum, C.; Thompson, D.A.; Friedman, N.; Gnirke, A.; Regev, A. Comprehensive Comparative Analysis of Strand-Specific RNA Sequencing Methods. Nat. Methods 2010, 7, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New Perspectives on Genomes, Pathways, Diseases and Drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Sorgi, C.A.; Rose, S.; Court, N.; Carlos, D.; Paula-Silva, F.W.G.; Assis, P.A.; Frantz, F.G.; Ryffel, B.; Quesniaux, V.; Faccioli, L.H. GM-CSF Priming Drives Bone Marrow-Derived Macrophages to a Pro-Inflammatory Pattern and Downmodulates PGE2 in Response to TLR2 Ligands. PLoS ONE 2012, 7, e40523. [Google Scholar] [CrossRef]
- Verma, N.K.; Ziegler, H.K.; Stocker, B.; Schoolnik, G.K. Induction of a Cellular Immune Response to a Defined T-Cell Epitope as an Insert in the Flagellin of a Live Vaccine Strain of Salmonella. Vaccine 1995, 13, 235–244. [Google Scholar] [CrossRef]
- Bao, W.; Wang, Y.; Fu, Y.; Jia, X.; Li, J.; Vangan, N.; Bao, L.; Hao, H.; Wang, Z. MTORC1 Regulates Flagellin-Induced Inflammatory Response in Macrophages. PLoS ONE 2015, 10, e0125910. [Google Scholar] [CrossRef] [Green Version]
- Hawn, T.R.; Berrington, W.R.; Smith, I.A.; Uematsu, S.; Akira, S.; Aderem, A.; Smith, K.D.; Skerrett, S.J. Altered Inflammatory Responses in TLR5-Deficient Mice Infected with Legionella Pneumophila. J. Immunol. 2007, 179, 6981–6987. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, F.A.; Aitken, J.D.; Gewirtz, A.T.; Vijay-Kumar, M. TLR5 Activation Induces Secretory Interleukin-1 Receptor Antagonist (SIL-1Ra) and Reduces Inflammasome-Associated Tissue Damage. Mucosal Immunol. 2011, 4, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Zhu, L.; Zhai, Y.; Zhao, Q.; Peng, J.; Zhang, H.; Yang, Z.; Zhang, L.; Ding, W.; Zhao, Y. TSC1 Controls IL-1β Expression in Macrophages via MTORC1-Dependent C/EBPβ Pathway. Cell Mol. Immunol. 2016, 13, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Ryan, D.G.; O’Neill, L.A.J. Krebs Cycle Rewired for Macrophage and Dendritic Cell Effector Functions. FEBS Lett. 2017, 591, 2992–3006. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, K.; Erathodiyil, N.; Gopalan, B.; Chong, S.; Wan, A.C.A.; Ying, J.Y. Targeting Warburg Effect in Cancers with PEGylated Glucose. Adv. Healthc. Mater. 2016, 5, 696–701. [Google Scholar] [CrossRef]
- Feingold, K.R.; Shigenaga, J.K.; Kazemi, M.R.; McDonald, C.M.; Patzek, S.M.; Cross, A.S.; Moser, A.; Grunfeld, C. Mechanisms of Triglyceride Accumulation in Activated Macrophages. J. Leukoc. Biol. 2012, 92, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Wang, Y.; Dong, L.; He, Y.; Liu, R.; Yang, Q.; Cao, Y.; Wang, Y.; Jia, A.; Bi, Y.; et al. Regulations of Glycolytic Activities on Macrophages Functions in Tumor and Infectious Inflammation. Front. Cell Infect. Microbiol. 2020, 10, 287. [Google Scholar] [CrossRef]
- Suzuki, H.; Hisamatsu, T.; Chiba, S.; Mori, K.; Kitazume, M.T.; Shimamura, K.; Nakamoto, N.; Matsuoka, K.; Ebinuma, H.; Naganuma, M.; et al. Glycolytic Pathway Affects Differentiation of Human Monocytes to Regulatory Macrophages. Immunol. Lett. 2016, 176, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, C.M.; Holowka, T.; Sun, J.; Blagih, J.; Amiel, E.; DeBerardinis, R.J.; Cross, J.R.; Jung, E.; Thompson, C.B.; Jones, R.G.; et al. Toll-like Receptor-Induced Changes in Glycolytic Metabolism Regulate Dendritic Cell Activation. Blood 2010, 115, 4742–4749. [Google Scholar] [CrossRef] [Green Version]
- Everts, B.; Amiel, E.; van der Windt, G.J.W.; Freitas, T.C.; Chott, R.; Yarasheski, K.E.; Pearce, E.L.; Pearce, E.J. Commitment to Glycolysis Sustains Survival of NO-Producing Inflammatory Dendritic Cells. Blood 2012, 120, 1422–1431. [Google Scholar] [CrossRef] [Green Version]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate Is an Inflammatory Signal That Induces IL-1β through HIF-1α. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef]
- Michl, J.; Ohlbaum, D.J.; Silverstein, S.C. 2-Deoxyglucose Selectively Inhibits Fc and Complement Receptor-Mediated Phagocytosis in Mouse Peritoneal Macrophages II. Dissociation of the Inhibitory Effects of 2-Deoxyglucose on Phagocytosis and ATP Generation. J. Exp. Med. 1976, 144, 1484–1493. [Google Scholar] [CrossRef]
- Cramer, T.; Yamanishi, Y.; Clausen, B.E.; Förster, I.; Pawlinski, R.; Mackman, N.; Haase, V.H.; Jaenisch, R.; Corr, M.; Nizet, V.; et al. HIF-1alpha Is Essential for Myeloid Cell-Mediated Inflammation. Cell 2003, 112, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Wang, Y.; Li, Y.; Yu, Q.; Jin, X.; Wang, X.; Jia, A.; Hu, Y.; Han, L.; Wang, J.; et al. HIF1α-Dependent Glycolysis Promotes Macrophage Functional Activities in Protecting against Bacterial and Fungal Infection. Sci. Rep. 2018, 8, 3603. [Google Scholar] [CrossRef]
- Jha, A.K.; Huang, S.C.-C.; Sergushichev, A.; Lampropoulou, V.; Ivanova, Y.; Loginicheva, E.; Chmielewski, K.; Stewart, K.M.; Ashall, J.; Everts, B.; et al. Network Integration of Parallel Metabolic and Transcriptional Data Reveals Metabolic Modules That Regulate Macrophage Polarization. Immunity 2015, 42, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Vats, D.; Mukundan, L.; Odegaard, J.I.; Zhang, L.; Smith, K.L.; Morel, C.R.; Wagner, R.A.; Greaves, D.R.; Murray, P.J.; Chawla, A. Oxidative Metabolism and PGC-1beta Attenuate Macrophage-Mediated Inflammation. Cell Metab. 2006, 4, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Talreja, J.; Talwar, H.; Bauerfeld, C.; Grossman, L.I.; Zhang, K.; Tranchida, P.; Samavati, L. HIF-1α Regulates IL-1β and IL-17 in Sarcoidosis. Elife 2019, 8. [Google Scholar] [CrossRef]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Lian, G.; Li, X.; Zhang, L.; Zhang, Y.; Sun, L.; Zhang, X.; Liu, H.; Pang, Y.; Kong, W.; Zhang, T.; et al. Macrophage Metabolic Reprogramming Aggravates Aortic Dissection through the HIF1α-ADAM17 Pathway. EBioMedicine 2019, 49, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-MTOR Pathways: Cross-Talk and Compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Kodaki, T.; Woscholski, R.; Hallberg, B.; Rodriguez-Viciana, P.; Downward, J.; Parker, P.J. The Activation of Phosphatidylinositol 3-Kinase by Ras. Curr. Biol. 1994, 4, 798–806. [Google Scholar] [CrossRef]
- Rodriguez-Viciana, P.; Warne, P.H.; Dhand, R.; Vanhaesebroeck, B.; Gout, I.; Fry, M.J.; Waterfield, M.D.; Downward, J. Phosphatidylinositol-3-OH Kinase as a Direct Target of Ras. Nature 1994, 370, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Suire, S.; Hawkins, P.; Stephens, L. Activation of Phosphoinositide 3-Kinase Gamma by Ras. Curr. Biol. 2002, 12, 1068–1075. [Google Scholar] [CrossRef] [Green Version]
- Roux, P.P.; Ballif, B.A.; Anjum, R.; Gygi, S.P.; Blenis, J. Tumor-Promoting Phorbol Esters and Activated Ras Inactivate the Tuberous Sclerosis Tumor Suppressor Complex via P90 Ribosomal S6 Kinase. Proc. Natl. Acad. Sci. USA 2004, 101, 13489–13494. [Google Scholar] [CrossRef] [Green Version]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. MTOR: From Growth Signal Integration to Cancer, Diabetes and Ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, L.R.; Komander, D.; Alessi, D.R. The Nuts and Bolts of AGC Protein Kinases. Nat. Rev. Mol. Cell Biol. 2010, 11, 9–22. [Google Scholar] [CrossRef]
- Carriere, A.; Romeo, Y.; Acosta-Jaquez, H.A.; Moreau, J.; Bonneil, E.; Thibault, P.; Fingar, D.C.; Roux, P.P. ERK1/2 Phosphorylate Raptor to Promote Ras-Dependent Activation of MTOR Complex 1 (MTORC1). J. Biol. Chem. 2011, 286, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Foster, K.G.; Acosta-Jaquez, H.A.; Romeo, Y.; Ekim, B.; Soliman, G.A.; Carriere, A.; Roux, P.P.; Ballif, B.A.; Fingar, D.C. Regulation of MTOR Complex 1 (MTORC1) by Raptor Ser863 and Multisite Phosphorylation. J. Biol. Chem. 2010, 285, 80–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo Name | Sequence |
|---|---|
| NNSR_RT | gctcttccgatctctNNNNNN |
| NNSR_2 | gctcttccgatctgaNNNNNN |
| NNSRnest_ind_N | CAAGCAGAAGACGGCATACGAGATNNNNNNGTGACTGGAGTTCAGACGTGTGCTCTTCCGATCTGA (N stands for a 6mer TruSeq index (Illumina)) |
| NNSR_Illumina | AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATCTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-J.; Papp, G.; Miskey, C.; Fiedler, A.; Goretzki, A.; Wolfheimer, S.; Zimmermann, J.; Crauwels, P.; Ivics, Z.; van Zandbergen, G.; et al. The Flagellin:Allergen Fusion Protein rFlaA:Betv1 Induces a MyD88− and MAPK-Dependent Activation of Glucose Metabolism in Macrophages. Cells 2021, 10, 2614. https://doi.org/10.3390/cells10102614
Lin Y-J, Papp G, Miskey C, Fiedler A, Goretzki A, Wolfheimer S, Zimmermann J, Crauwels P, Ivics Z, van Zandbergen G, et al. The Flagellin:Allergen Fusion Protein rFlaA:Betv1 Induces a MyD88− and MAPK-Dependent Activation of Glucose Metabolism in Macrophages. Cells. 2021; 10(10):2614. https://doi.org/10.3390/cells10102614
Chicago/Turabian StyleLin, Yen-Ju, Garibald Papp, Csaba Miskey, Anna Fiedler, Alexandra Goretzki, Sonja Wolfheimer, Jennifer Zimmermann, Peter Crauwels, Zoltán Ivics, Ger van Zandbergen, and et al. 2021. "The Flagellin:Allergen Fusion Protein rFlaA:Betv1 Induces a MyD88− and MAPK-Dependent Activation of Glucose Metabolism in Macrophages" Cells 10, no. 10: 2614. https://doi.org/10.3390/cells10102614