
The Coordinated KNR6–AGAP–ARF1 Complex Modulates Vegetative and Reproductive Traits by Participating in Vesicle Trafficking in Maize

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
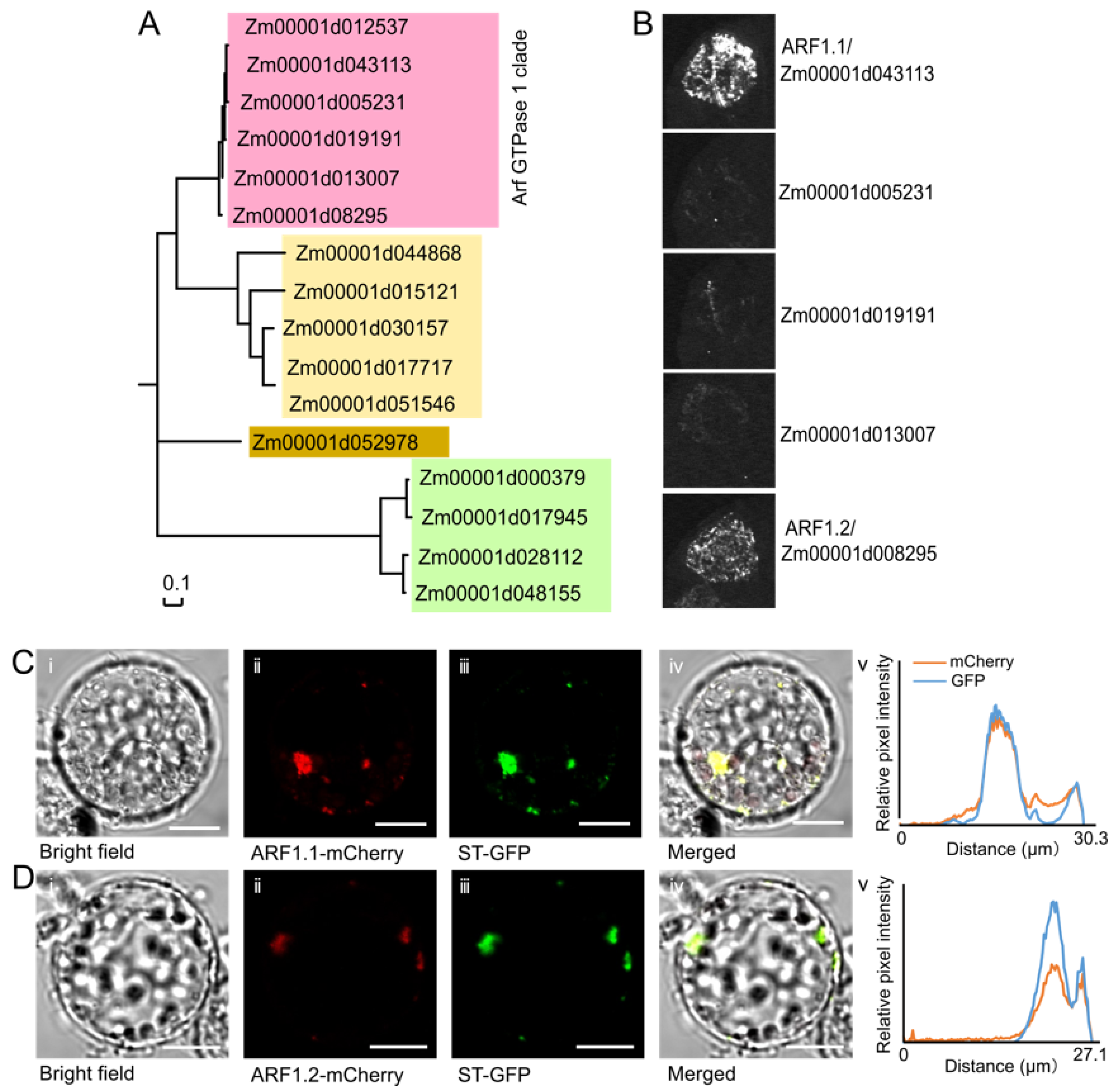
2.1. Sequence Extraction and Phylogenetic Analysis
2.2. Vector Construction and Genetic Transformation
2.3. Plant Materials and Phenotypic Identification
2.4. Luciferase (Luc) Complementation Image Assay
2.5. Yeast Three-Hybrid (Y3H) Assay
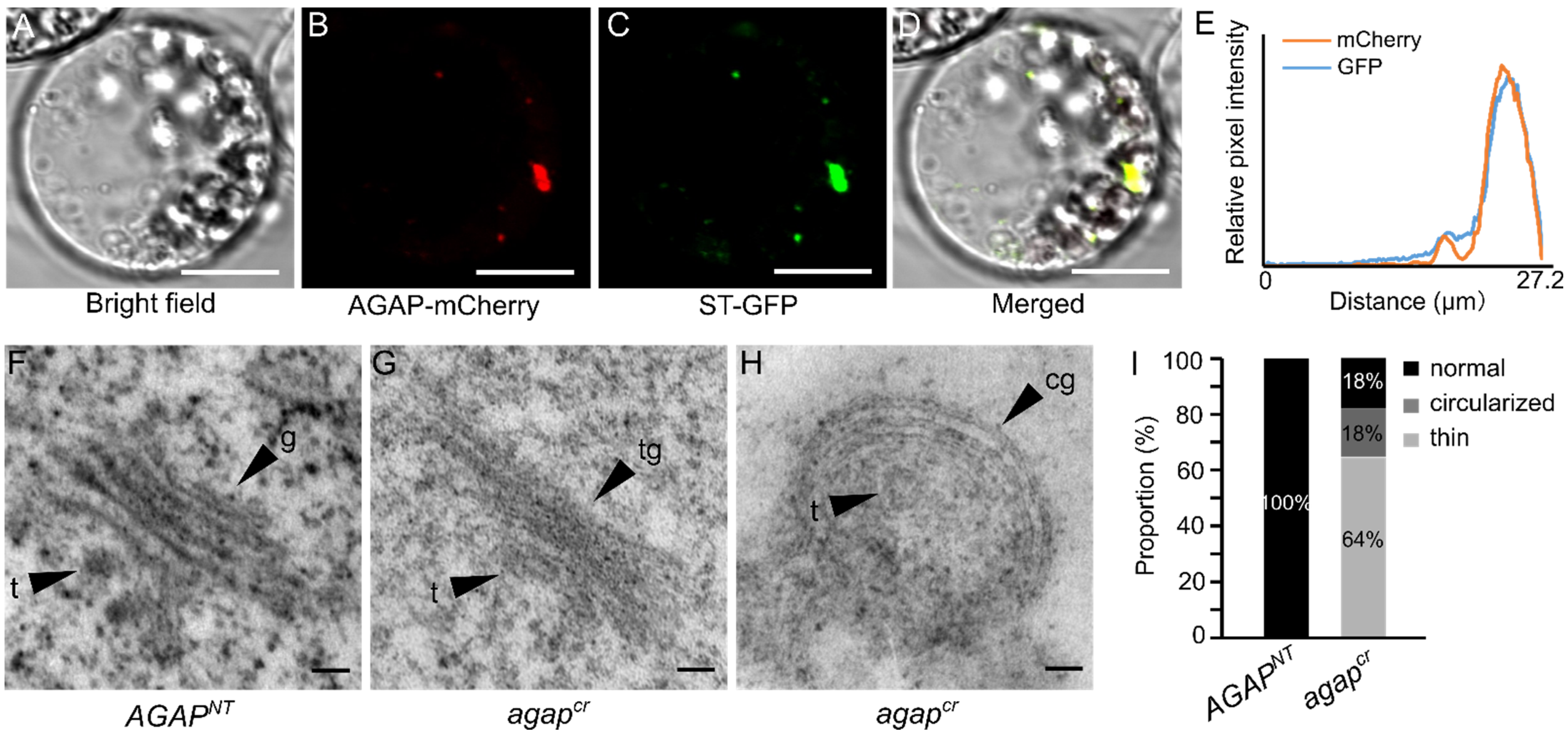
2.6. Subcellular Localization
2.7. Transmission Electron Microscopic Observation of Golgi Apparatus
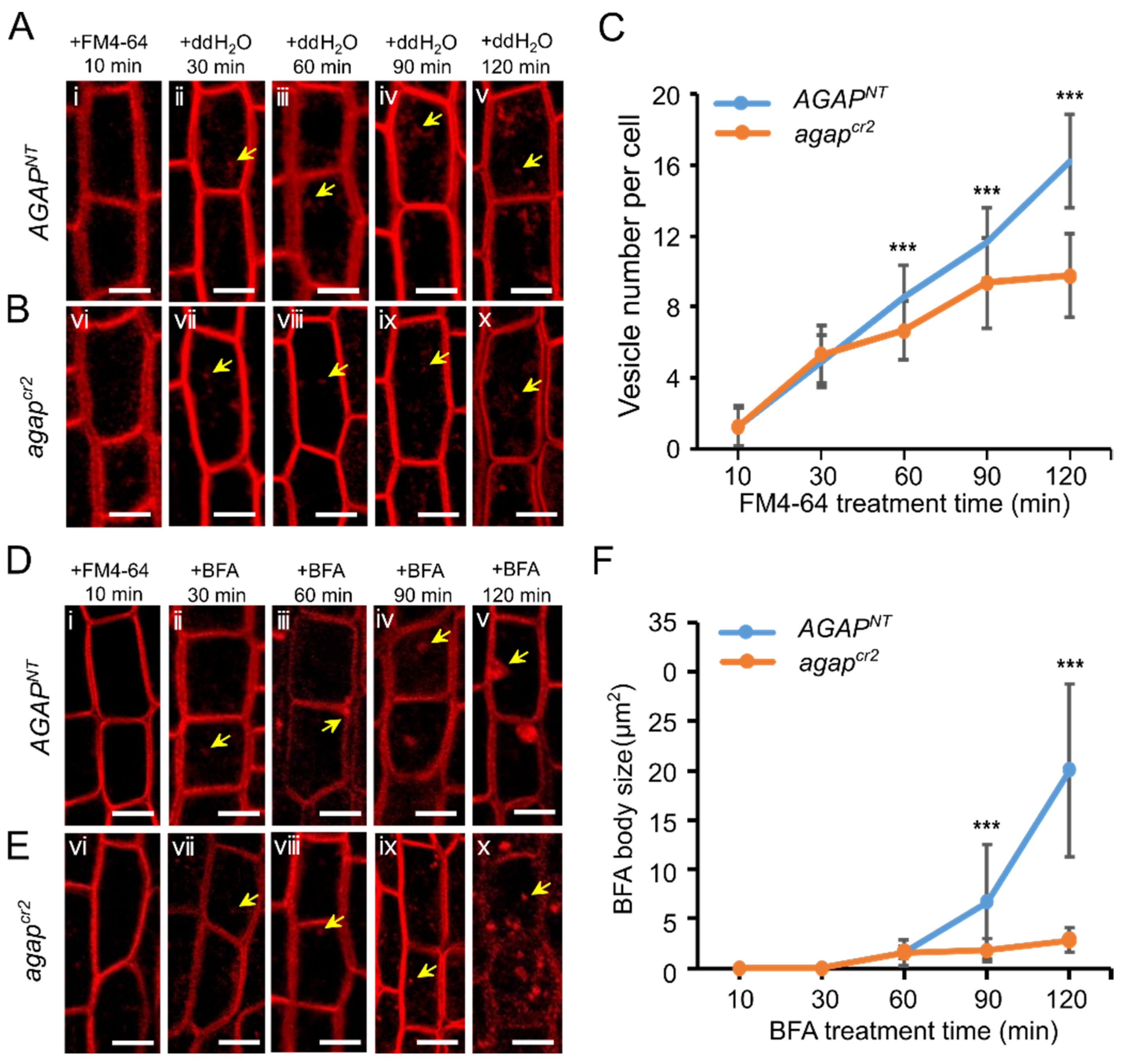
2.8. FM4-64 Internalization Assay
3. Results
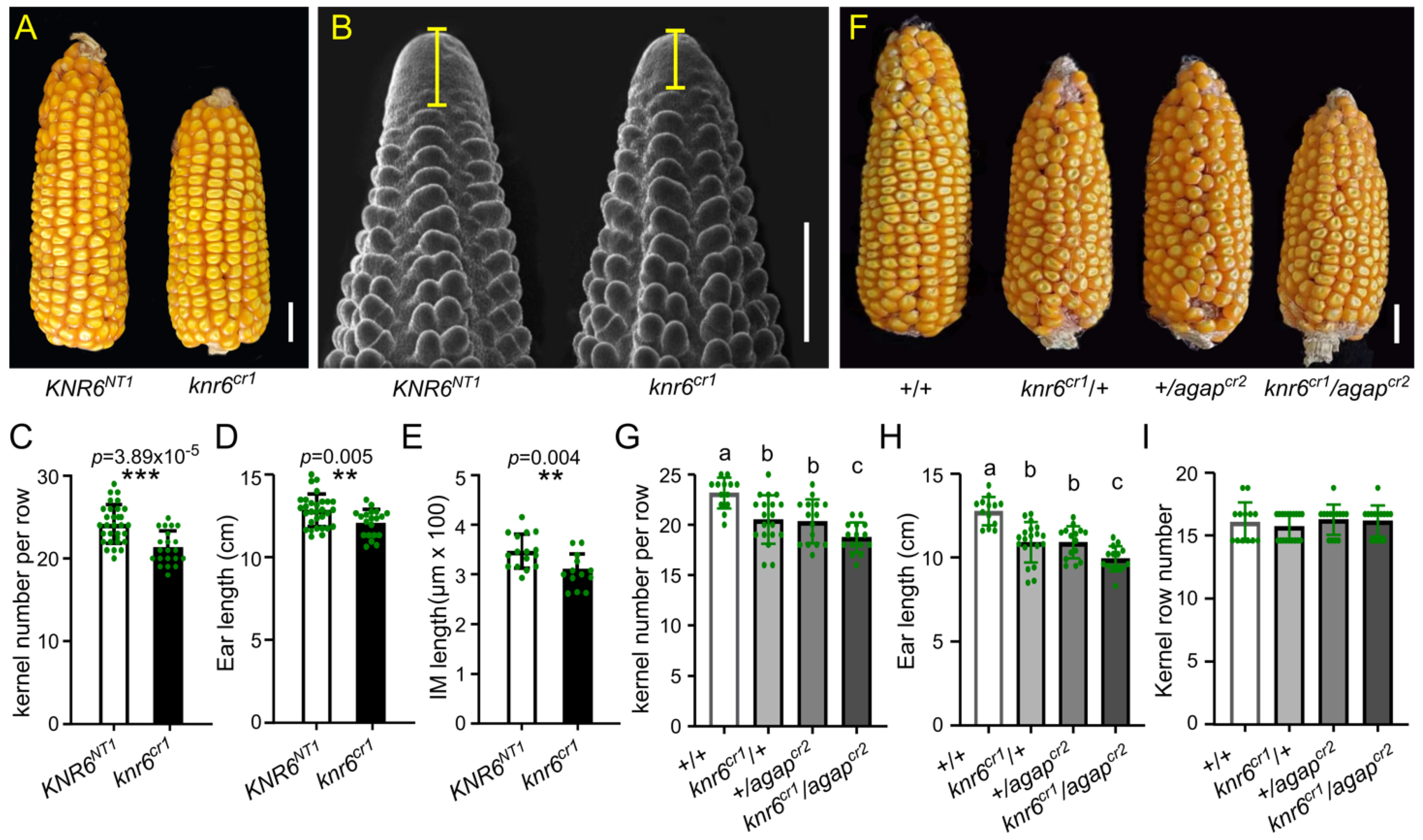
3.1. AGAP Gene Shows Pleiotropy in Vegetative and Reproductive Traits
3.2. AGAP Genetically Interacts with KNR6
3.3. AGAP Participates in Vesicle Trafficking
3.4. AGAP Physically Interacts with Two Members of the Arf GTPase 1 (ARF1) Protein Subfamily
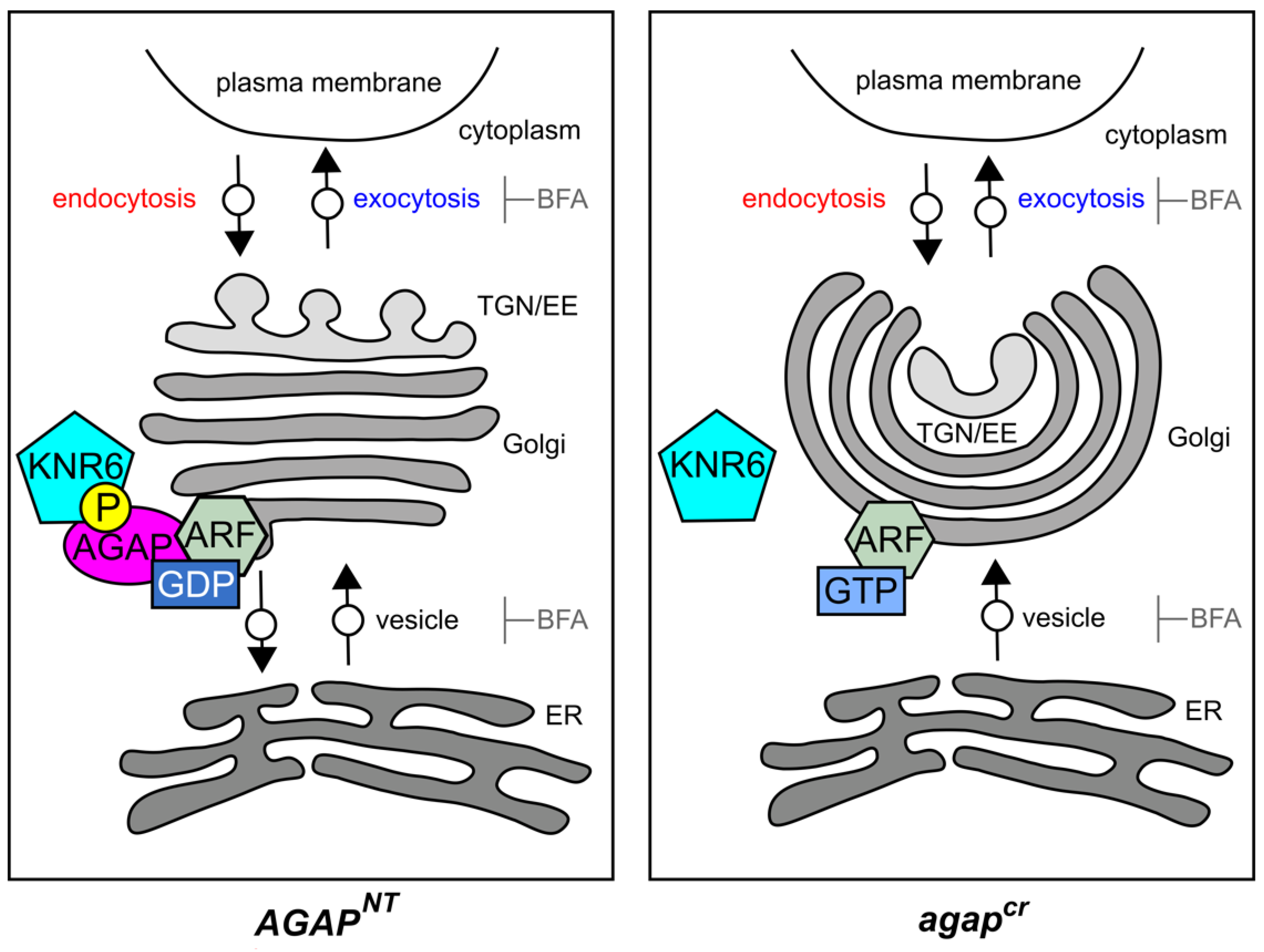
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, P.C.; Greyson, R.I.; Walden, D.B. Organ initiation and the development of unisexual flowers in the tassel and ear of zea mays. Am. J. Bot. 1983, 70, 450–462. [Google Scholar] [CrossRef]
- Irish, E.E. Class II tassel seed mutations provide evidence for multiple types of inflorescence meristems in maize (Poaceae). Am. J. Bot. 1997, 84, 1502–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSteen, P.; Laudencia-Chingcuanco, D.; Colasanti, J. A floret by any other name: Control of meristem identity in maize. Trends Plant Sci. 2000, 5, 61–66. [Google Scholar] [CrossRef]
- Vollbrecht, E.; Springer, P.S.; Goh, L.; Iv, E.S.B.; Martienssen, R. Architecture of floral branch systems in maize and related grasses. Nature 2005, 436, 1119–1126. [Google Scholar] [CrossRef]
- Gallavotti, A.; Barazesh, S.; Malcomber, S.; Hall, D.; Jackson, D.; Schmidt, R.J.; McSteen, P. sparse inflorescence1 encodes a monocot-specific YUCCA-like gene required for vegetative and reproductive development in maize. Proc. Natl. Acad. Sci. USA 2008, 105, 15196–15201. [Google Scholar] [CrossRef] [Green Version]
- Phillips, K.A.; Skirpan, A.L.; Liu, X.; Christensen, A.; Slewinski, T.L.; Hudson, C.; Barazesh, S.; Cohen, J.; Malcomber, S.; McSteen, P. vanishing tassel2 Encodes a Grass-Specific Tryptophan Aminotransferase Required for Vegetative and Reproductive Development in Maize. Plant Cell 2011, 23, 550–566. [Google Scholar] [CrossRef] [Green Version]
- Galli, M.; Liu, Q.; Moss, B.; Malcomber, S.; Li, W.; Gaines, C.; Federici, S.; Roshkovan, J.; Meeley, R.; Nemhauser, J.L.; et al. Auxin signaling modules regulate maize inflorescence architecture. Proc. Natl. Acad. Sci. USA 2015, 112, 13372–13377. [Google Scholar] [CrossRef] [Green Version]
- Kleine-Vehn, J.; Dhonukshe, P.; Swarup, R.; Bennett, M.; Friml, J. Subcellular Trafficking of the Arabidopsis Auxin Influx Carrier AUX1 Uses a Novel Pathway Distinct from PIN1. Plant Cell 2006, 18, 3171–3181. [Google Scholar] [CrossRef] [Green Version]
- Swarup, R.; Péret, B. AUX/LAX family of auxin influx carriers—an overview. Front. Plant Sci. 2012, 3, 225. [Google Scholar] [CrossRef] [Green Version]
- Carraro, N.; Forestan, C.; Canova, S.; Traas, J.; Varotto, S. ZmPIN1a and ZmPIN1bEncode Two Novel Putative Candidates for Polar Auxin Transport and Plant Architecture Determination of Maize. Plant Physiol. 2006, 142, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Skirpan, A.; Culler, A.H.; Gallavotti, A.; Jackson, D.; Cohen, J.D.; McSteen, P. BARREN INFLORESCENCE2 Interaction with ZmPIN1a Suggests a Role in Auxin Transport During Maize Inflorescence Development. Plant Cell Physiol. 2009, 50, 652–657. [Google Scholar] [CrossRef] [Green Version]
- McSteen, P.; Malcomber, S.; Skirpan, A.; Lunde, C.; Wu, X.; Kellogg, E.; Hake, S. barren inflorescence2Encodes a Co-Ortholog of thePINOIDSerine/Threonine Kinase and Is Required for Organogenesis during Inflorescence and Vegetative Development in Maize. Plant Physiol. 2007, 144, 1000–1011. [Google Scholar] [CrossRef] [Green Version]
- Steinmann, T.; Geldner, N.; Grebe, M.; Mangold, S.; Jackson, C.L.; Paris, S.; Gälweiler, L.; Palme, K.; Jürgens, G. Coordinated Polar Localization of Auxin Efflux Carrier PIN1 by GNOM ARF GEF. Science 1999, 286, 316–318. [Google Scholar] [CrossRef]
- Grebe, M.; Friml, J.; Swarup, R.; Ljung, K.; Sandberg, G.; Terlou, M.; Palme, K.; Bennett, M.; Scheres, B. Cell Polarity Signaling in Arabidopsis Involves a BFA-Sensitive Auxin Influx Pathway. Curr. Biol. 2002, 12, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Muday, G.K.; Peer, W.; Murphy, A.S. Vesicular cycling mechanisms that control auxin transport polarity. Trends Plant Sci. 2003, 8, 301–304. [Google Scholar] [CrossRef]
- Zhuang, X.; Jiang, J.; Li, J.; Ma, Q.; Xu, Y.; Xue, Y.; Xu, Z.; Chong, K. Over-expression of OsAGAP, an ARF-GAP, interferes with auxin influx, vesicle trafficking and root development. Plant J. 2006, 48, 581–591. [Google Scholar] [CrossRef]
- Cohen, M.S.; Chang, P. Insights into the biogenesis, function, and regulation of ADP-ribosylation. Nat. Chem. Biol. 2018, 14, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, L.; Mikoč, A.; Ahel, I. ADP-ribosylation: New facets of an ancient modification. FEBS J. 2017, 284, 2932–2946. [Google Scholar] [CrossRef]
- Moss, J.; Vaughan, M. Molecules in the ARF Orbit. J. Biol. Chem. 1998, 273, 21431–21434. [Google Scholar] [CrossRef] [Green Version]
- Naramoto, S.; Otegui, M.S.; Kutsuna, N.; de Rycke, R.; Dainobu, T.; Karampelias, M.; Fujimoto, M.; Feraru, E.; Miki, D.; Fukuda, H.; et al. Insights into the Localization and Function of the Membrane Trafficking Regulator GNOM ARF-GEF at the Golgi Apparatus in Arabidopsis. Plant Cell 2014, 26, 3062–3076. [Google Scholar] [CrossRef] [Green Version]
- Geldner, N.; Anders, N.; Wolters, H.; Keicher, J.; Kornberger, W.; Muller, P.; Delbarre, A.; Ueda, T.; Nakano, A.; Jürgens, G. The Arabidopsis GNOM ARF-GEF Mediates Endosomal Recycling, Auxin Transport, and Auxin-Dependent Plant Growth. Cell 2003, 112, 219–230. [Google Scholar] [CrossRef]
- Richter, S.; Anders, N.; Wolters, H.; Beckmann, H.; Thomann, A.; Heinrich, R.; Schrader, J.; Singh, M.K.; Geldner, N.; Mayer, U.; et al. Role of the GNOM gene in Arabidopsis apical-basal patterning—From mutant phenotype to cellular mechanism of protein action. Eur. J. Cell Biol. 2010, 89, 138–144. [Google Scholar] [CrossRef]
- Koizumi, K.; Naramoto, S.; Sawa, S.; Yahara, N.; Ueda, T.; Nakano, A.; Sugiyama, M.; Fukuda, H. VAN3 ARF–GAP-mediated vesicle transport is involved in leaf vascular network formation. Development 2005, 132, 1699–1711. [Google Scholar] [CrossRef] [Green Version]
- Sieburth, L.E.; Muday, G.K.; King, E.J.; Benton, G.; Kim, S.; Metcalf, K.E.; Meyers, L.; Seamen, E.; Van Norman, J.M. SCARFACE Encodes an ARF-GAP That Is Required for Normal Auxin Efflux and Vein Patterning in Arabidopsis. Plant Cell 2006, 18, 1396–1411. [Google Scholar] [CrossRef] [Green Version]
- Song, X.-F.; Yang, C.-Y.; Liu, J.; Yang, W.-C. RPA, a Class II ARFGAP Protein, Activates ARF1 and U5 and Plays a Role in Root Hair Development in Arabidopsis. Plant Physiol. 2006, 141, 966–976. [Google Scholar] [CrossRef] [Green Version]
- Yoo, C.-M.; Wen, J.; Motes, C.M.; Sparks, J.A.; Blancaflor, E.B. A Class I ADP-Ribosylation Factor GTPase-Activating Protein Is Critical for Maintaining Directional Root Hair Growth in Arabidopsis. Plant Physiol. 2008, 147, 1659–1674. [Google Scholar] [CrossRef] [Green Version]
- Min, M.K.; Kim, S.J.; Miao, Y.; Shin, J.; Jiang, L.; Hwang, I. Overexpression of Arabidopsis AGD7 Causes Relocation of Golgi-Localized Proteins to the Endoplasmic Reticulum and Inhibits Protein Trafficking in Plant Cells. Plant Physiol. 2007, 143, 1601–1614. [Google Scholar] [CrossRef] [Green Version]
- Liljegren, S.J.; Leslie, M.; Darnielle, L.; Lewis, M.W.; Taylor, S.M.; Luo, R.; Geldner, N.; Chory, J.; Randazzo, P.A.; Yanofsky, M.F.; et al. Regulation of membrane trafficking and organ separation by the NEVERSHED ARF-GAP protein. Development 2009, 136, 1909–1918. [Google Scholar] [CrossRef] [Green Version]
- Stefano, G.; Renna, L.; Hanton, S.L.; Chatre, L.; Haas, T.A.; Brandizzi, F. ARL1 Plays a Role in the Binding of the GRIP Domain of a Peripheral Matrix Protein to the Golgi Apparatus in Plant Cells. Plant Mol. Biol. 2006, 61, 431–449. [Google Scholar] [CrossRef]
- Matheson, L.A.; Hanton, S.L.; Rossi, M.; Latijnhouwers, M.; Stefano, G.; Renna, L.; Brandizzi, F. Multiple Roles of ADP-Ribosylation Factor 1 in Plant Cells Include Spatially Regulated Recruitment of Coatomer and Elements of the Golgi Matrix. Plant Physiol. 2007, 143, 1615–1627. [Google Scholar] [CrossRef] [Green Version]
- Matheson, L.A.; Suri, S.S.; Hanton, S.L.; Chatre, L.; Brandizzi, F. Correct Targeting of Plant ARF GTPases Relies on Distinct Protein Domains. Traffic 2008, 9, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Min, M.K.; Lee, Y.J.; Jin, J.B.; Shin, D.H.; Kim, D.H.; Lee, K.-H.; Hwang, I. ADP-Ribosylation Factor 1 of Arabidopsis Plays a Critical Role in Intracellular Trafficking and Maintenance of Endoplasmic Reticulum Morphology in Arabidopsis. Plant Physiol. 2002, 129, 1507–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, M.; Ueda, T.; Yahara, N.; Nakano, A. Arf1 GTPase plays roles in the protein traffic between the endoplasmic reticulum and the Golgi apparatus in tobacco andArabidopsiscultured cells. Plant J. 2002, 31, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Pimpl, P.; Hanton, S.L.; Taylor, J.P.; Dasilva, L.L.; Denecke, J. The GTPase ARF1p Controls the Sequence-Specific Vacuolar Sorting Route to the Lytic Vacuole. Plant Cell 2003, 15, 1242–1256. [Google Scholar] [CrossRef] [Green Version]
- Cukierman, E.; Huber, I.; Rotman, M.; Cassel, D. The ARF1 GTPase-Activating Protein: Zinc Finger Motif and Golgi Complex Localization. Science 1995, 270, 1999–2002. [Google Scholar] [CrossRef]
- Donaldson, J.G.; Jackson, C.L. Regulators and effectors of the ARF GTPases. Curr. Opin. Cell Biol. 2000, 12, 475–482. [Google Scholar] [CrossRef]
- Jensen, R.B.; Lykke-Andersen, K.; Frandsen, G.I.; Nielsen, H.B.; Haseloff, J.; Jespersen, H.M.; Mundy, J.; Skriver, K. Promiscuous and specific phospholipid binding by domains in ZAC, a membrane-associated Arabidopsis protein with an ARF GAP zinc finger and a C2 domain. Plant Mol. Biol. 2000, 44, 799–814. [Google Scholar] [CrossRef]
- Jia, H.; Li, M.; Li, W.; Liu, L.; Jian, Y.; Yang, Z.; Shen, X.; Ning, Q.; Du, Y.; Zhao, R.; et al. A serine/threonine protein kinase encoding gene KERNEL NUMBER PER ROW6 regulates maize grain yield. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef] [Green Version]
- Li, X. Infiltration of Nicotiana benthamiana Protocol for Transient Expression via Agrobacterium. Bio-Protocol 2011, 1. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Hao, H.; Xue, Y.; Zhang, L.; Song, K.; Ding, Z.; Botella, M.; Wang, H.; Lin, J. Dynamic analysis of Arabidopsis AP2 σ subunit reveals a key role in clathrin-mediated endocytosis and plant development. Development 2013, 140, 3826–3837. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, W.; Wang, H.; Du, Q.; Fu, Z.; Li, W.-X.; Tang, J. ZmEHD1 Is Required for Kernel Development and Vegetative Growth through Regulating Auxin Homeostasis. Plant Physiol. 2019, 182, 1467–1480. [Google Scholar] [CrossRef] [Green Version]
- Boevink, P.; Oparka, K.; Cruz, S.S.; Martin, B.; Betteridge, A.; Hawes, C. Stacks on tracks: The plant Golgi apparatus traffics on an actin/ER network. Plant J. 1998, 15, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Nebenführ, A.; Ritzenthaler, C.; Robinson, D.G. Brefeldin A: Deciphering an Enigmatic Inhibitor of Secretion. Plant Physiol. 2002, 130, 1102–1108. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Tejos, R.; Beck, M.; Himschoot, E.; Li, H.; Robatzek, S.; Vanneste, S.; Friml, J. Salicylic acid interferes with clathrin-mediated endocytic protein trafficking. Proc. Natl. Acad. Sci. USA 2013, 110, 7946–7951. [Google Scholar] [CrossRef] [Green Version]
- Poon, P.P.; Wang, X.; Rotman, M.; Huber, I.; Cukierman, E.; Cassel, D.; Singer, R.A.; Johnston, G.C. Saccharomyces cerevisiae Gcs1 is an ADP-ribosylation factor GTPase-activating protein. Proc. Natl. Acad. Sci. USA 1996, 93, 10074–10077. [Google Scholar] [CrossRef] [Green Version]
- A Kahn, R.; Gilman, A.G. The protein cofactor necessary for ADP-ribosylation of Gs by cholera toxin is itself a GTP binding protein. J. Biol. Chem. 1986, 261, 7906–7911. [Google Scholar] [CrossRef]
- Gommel, D.U.; Memon, A.R.; Heiss, A.; Lottspeich, F.; Pfannstiel, J.; Lechner, J.; Reinhard, C.; Helms, J.; Nickel, W.; Wieland, F.T. Recruitment to Golgi membranes of ADP-ribosylation factor 1 is mediated by the cytoplasmic domain of p23. EMBO J. 2001, 20, 6751–6760. [Google Scholar] [CrossRef] [Green Version]
- Vollbrecht, E.; Schmidt, R.J. Development of the inflorescences. In Handbook of Maize: Its Biology; Springer: New York, NY, USA, 2009; pp. 13–40. [Google Scholar]
- Li, M.; Zhong, W.; Yang, F.; Zhang, Z. Genetic and Molecular Mechanisms of Quantitative Trait Loci Controlling Maize Inflorescence Architecture. Plant Cell Physiol. 2018, 59, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Leal, D.; Xu, C.; Kwon, C.-T.; Soyars, C.; Demesa-Arevalo, E.; Man, J.; Liu, L.; Lemmon, Z.H.; Jones, D.S.; Van Eck, J.; et al. Evolution of buffering in a genetic circuit controlling plant stem cell proliferation. Nat. Genet. 2019, 51, 786–792. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, F.; Liu, L.; Char, S.N.; Ding, Y.; Je, B.I.; Schmelz, E.; Yang, B.; Jackson, D. The maize heterotrimeric G protein β subunit controls shoot meristem development and immune responses. Proc. Natl. Acad. Sci. USA 2019, 117, 1799–1805. [Google Scholar] [CrossRef]
- Pitcher, J.A.; Freedman, N.J.; Lefkowitz, R.J. G protein–coupled receptor kinases. Ann. Rev. Biochem. 1998, 67, 653–692. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.; Xu, Y.; Chong, K.; Lan, L.; Xu, Z.; Xue, Y. OsAGAP, an ARF-GAP from rice, regulates root development mediated by auxin in Arabidopsis. Plant Cell Environ. 2004, 28, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Chong, K. ARF-GTPase activating protein mediates auxin influx carrier AUX1 early endosome trafficking to regulate auxin dependent plant development. Plant Signal. Behav. 2011, 6, 1644–1646. [Google Scholar] [CrossRef] [Green Version]
- Jiang23, L.; Rogers, S.L.; Crews, S.T. The Drosophila Dead end Arf-like3 GTPase controls vesicle trafficking during tracheal fusion cell morphogenesis. Dev. Biol. 2007, 311, 487–499. [Google Scholar] [CrossRef] [Green Version]
- Min, M.K.; Jang, M.; Lee, M.; Lee, J.; Song, K.; Lee, Y.; Choi, K.Y.; Robinson, D.G.; Hwang, I. Recruitment of Arf1-GDP to Golgi by Glo3p-Type ArfGAPs Is Crucial for Golgi Maintenance and Plant Growth. Plant Physiol. 2012, 161, 676–691. [Google Scholar] [CrossRef] [Green Version]
- Bigay, J.; Gounon, P.; Robineau, S.; Antonny, B. Lipid packing sensed by ArfGAP1 couples COPI coat disassembly to membrane bilayer curvature. Nature 2003, 426, 563–566. [Google Scholar] [CrossRef]
- Nie, Z.; Hirsch, D.S.; A Randazzo, P. Arf and its many interactors. Curr. Opin. Cell Biol. 2003, 15, 396–404. [Google Scholar] [CrossRef]
- Inoue, H.; Randazzo, P.A. Arf GAPs and Their Interacting Proteins. Traffic 2007, 8, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.K.; Jürgens, G. Specificity of plant membrane trafficking–ARFs, regulators and coat proteins. Semin. Cell Dev. Biol. 2018, 80, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J. Structural Basis for Activation of ARF GTPase: Mechanisms of Guanine Nucleotide Exchange and GTP–Myristoyl Switching. Cell 1998, 95, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Weissman, J.T.; Béraud-Dufour, S.; Luan, P.; Wang, C.; Chen, W.; Aridor, M.; Wilson, I.A.; Balch, W.E. Crystal structure of Sar1-GDP at 1.7 Å resolution and the role of the NH2 terminus in ER export. J. Cell Biol. 2001, 155, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Gillingham, A.K.; Munro, S. The Small G Proteins of the Arf Family and Their Regulators. Annu. Rev. Cell Dev. Biol. 2007, 23, 579–611. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Zhao, R.; Du, Y.; Shen, X.; Ning, Q.; Li, Y.; Liu, D.; Xiong, Q.; Zhang, Z. The Coordinated KNR6–AGAP–ARF1 Complex Modulates Vegetative and Reproductive Traits by Participating in Vesicle Trafficking in Maize. Cells 2021, 10, 2601. https://doi.org/10.3390/cells10102601
Li M, Zhao R, Du Y, Shen X, Ning Q, Li Y, Liu D, Xiong Q, Zhang Z. The Coordinated KNR6–AGAP–ARF1 Complex Modulates Vegetative and Reproductive Traits by Participating in Vesicle Trafficking in Maize. Cells. 2021; 10(10):2601. https://doi.org/10.3390/cells10102601
Chicago/Turabian StyleLi, Manfei, Ran Zhao, Yanfang Du, Xiaomeng Shen, Qiang Ning, Yunfu Li, Dan Liu, Qing Xiong, and Zuxin Zhang. 2021. "The Coordinated KNR6–AGAP–ARF1 Complex Modulates Vegetative and Reproductive Traits by Participating in Vesicle Trafficking in Maize" Cells 10, no. 10: 2601. https://doi.org/10.3390/cells10102601