Study on Transglucosylation Properties of Amylosucrase from Xanthomonas campestris pv. Campestris and Its Application in the Production of α-Arbutin


Abstract
:1. Introduction
2. Results
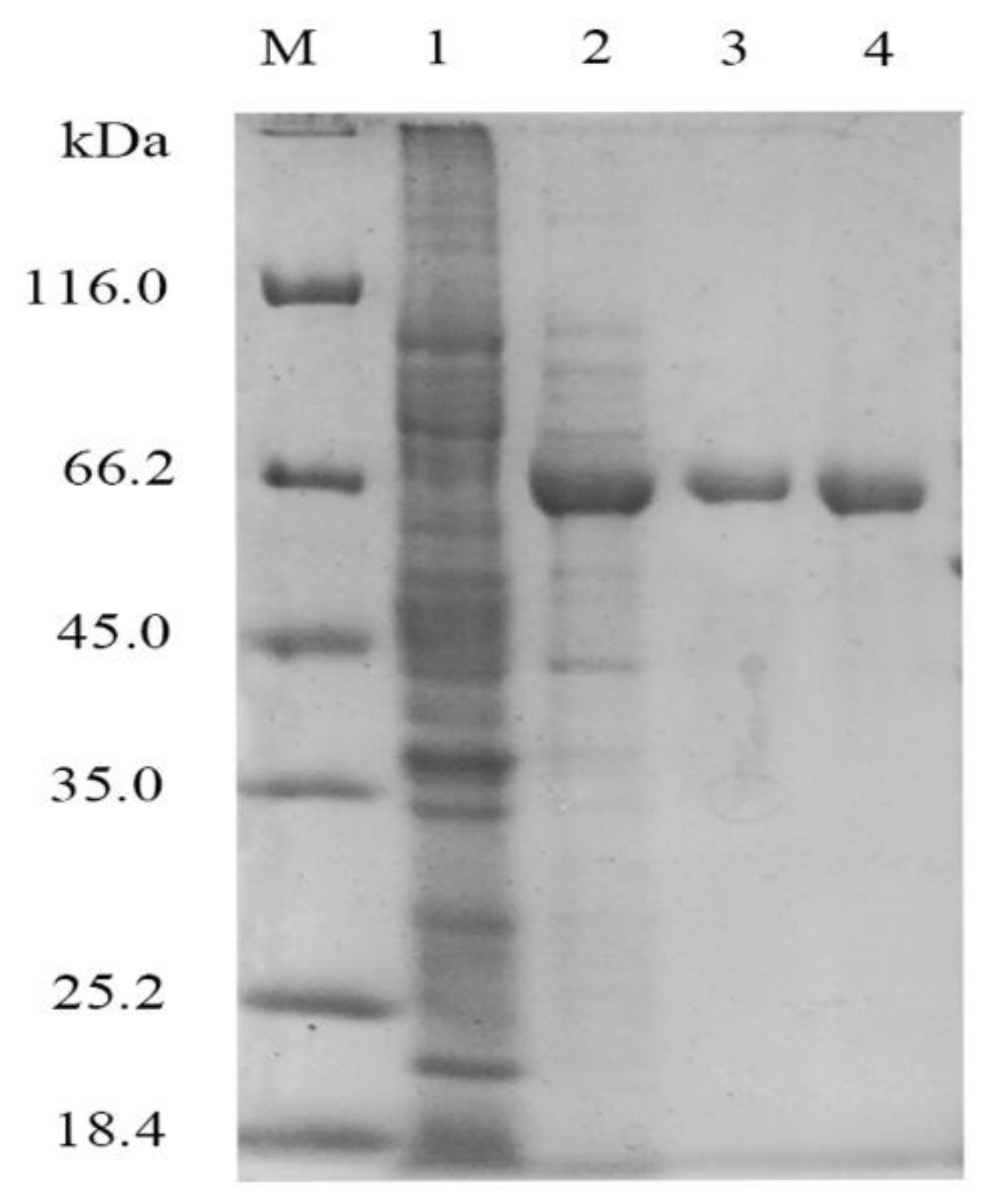
2.1. XcAS Heterologous Expression and Protein Purification
2.2. Bioconversion of HQ to α-Arbutin by Purified XcAS
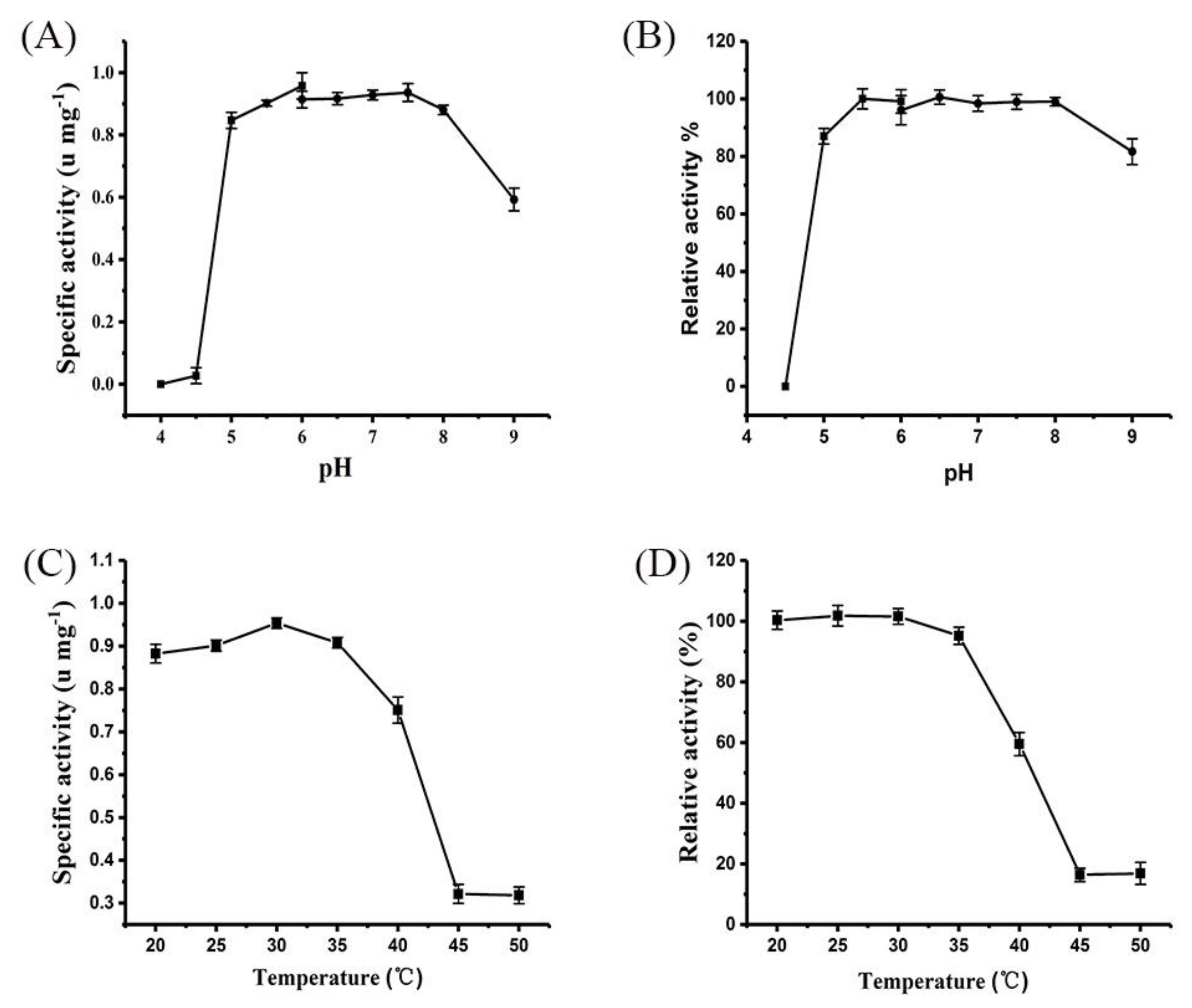
2.3. Catalysis Characterization of Purified XcAS on Transglucoside Reaction
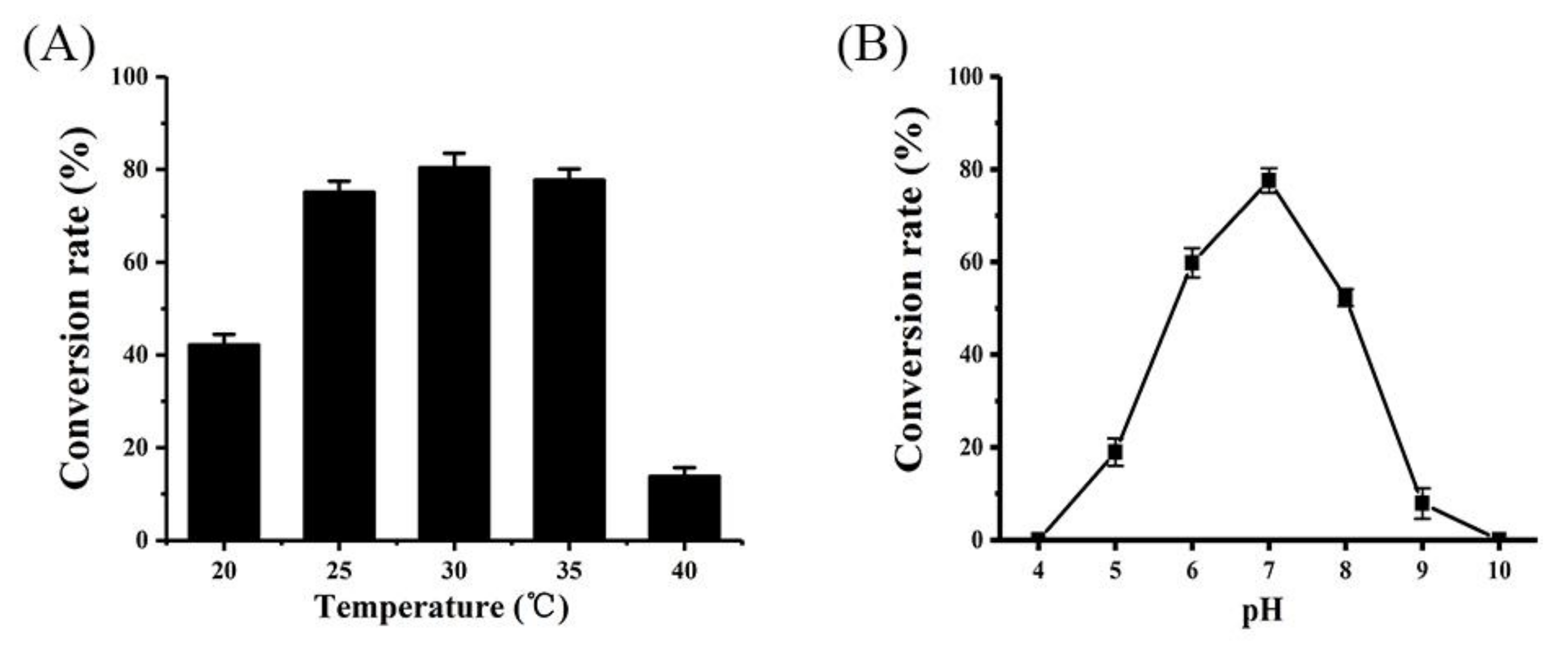
2.4. Effects of pH and Temperature on α-Arbutin Production by Whole-Cell Biocatalysis
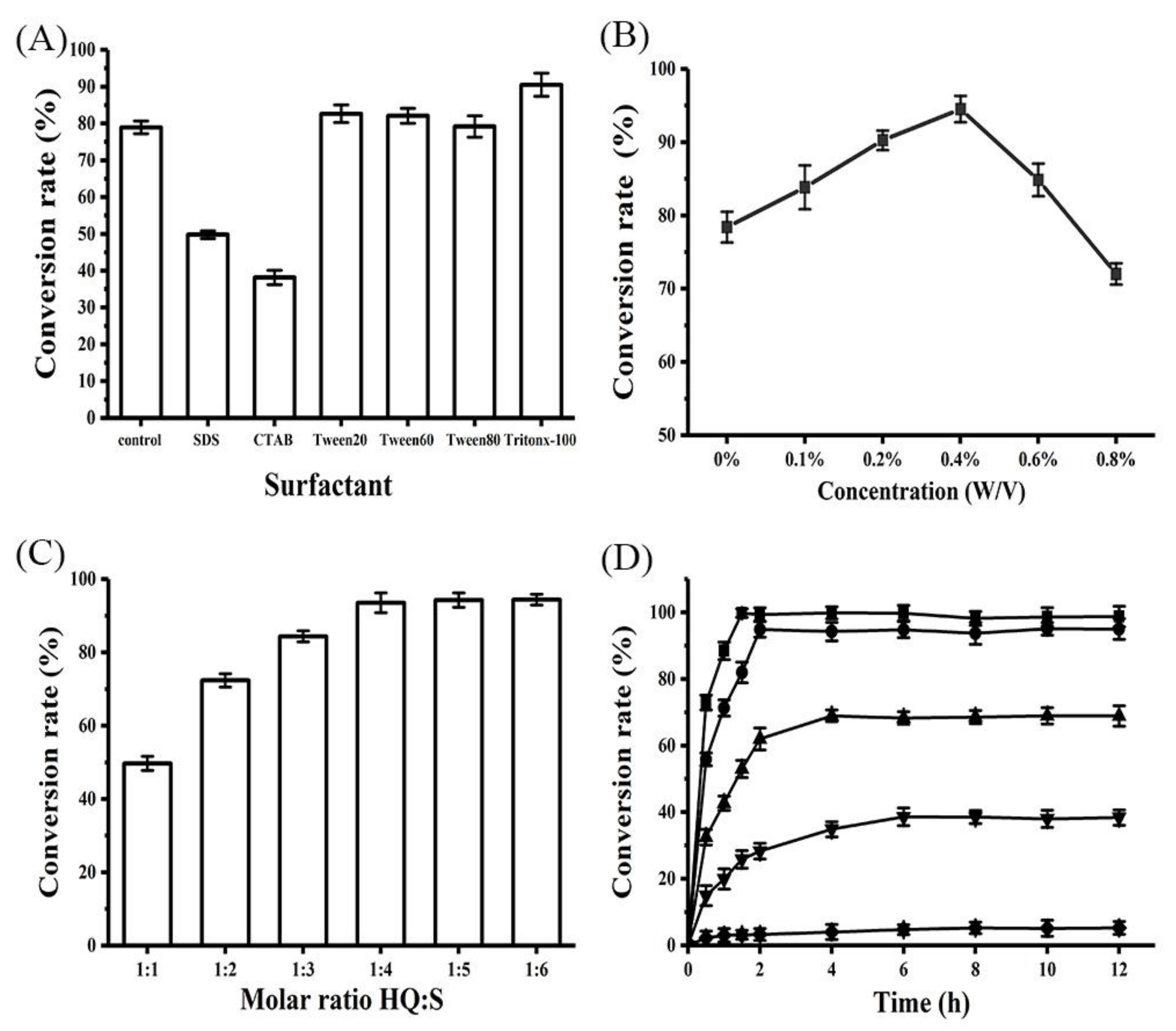
2.5. Effects of Surfactant and Its Concentration on α-Arbutin Production
2.6. Effects of Substrate Ratio and Concentration on α-Arbutin Production
2.7. Efficient Production of α-Arbutin through a Batch and Fed-Batch Strategy
3. Discussion
4. Materials and Methods
4.1. Microorganism Plasmids and Chemicals
4.2. Cloning and Recombinant Expression of ASase Gene from X. campestris pv. campestris
4.3. Protein Purification
4.4. Enzymatic Activity Assay and SDS-PAGE
4.5. Enzyme Properties of Purified XcAS on Transglucosylation
4.6. Bioconversion of HQ to α-Arbutin by Whole-Cell Biocatalysis
4.7. Analytical Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seo, D.H.; Jung, J.H.; Lee, J.E.; Jeon, E.J.; Kim, W.; Park, C.S. Biotechnological production of arbutins (alpha- and beta-arbutins), skin-lightening agents, and their derivatives. Appl. Microbiol. Biotechnol. 2012, 95, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Nishimura, T.; Nomura, K.; Sugimoto, K.; Kuriki, T. Inhibitory effects of alpha-arbutin on melanin synthesis in cultured human melanoma cells and a three-dimensional human skin model. Biol. Pharm. Bull. 2004, 27, 510–514. [Google Scholar] [CrossRef]
- Funayama, M.; Arakawa, H.; Yamamoto, R.; Nishino, T.; Shin, T.; Murao, S. Effects of alpha- and beta-arbutin on activity of tyrosinases from mushroom and mouse melanoma. Biosci. Biotechnol. Biochem. 1995, 59, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Kometani, T.; Takii, H.; Terada, Y.; Okada, S. Purifiction and some properties of alpha-amylase from bacillus-subtilis X-23 that glucosylates phenolic-compounds such as hydroquinone. J. Ferment. Bioeng. 1994, 78, 31–36. [Google Scholar] [CrossRef]
- Kurosu, J.; Sato, T.; Yoshida, K.; Tsugane, T.; Shimura, S.; Kirimura, K.; Kino, K.; Usami, S. Enzymatic synthesis of alpha-arbutin by alpha-anomer-selective-glucosylation of hydroquinone using lyophilized cells of Xanthomonas campestris WU-9701. J. Biosci. Bioeng. 2002, 93, 328–330. [Google Scholar] [CrossRef]
- Liu, C.Q.; Deng, L.; Zhang, P.; Zhang, S.R.; Liu, L.; Xu, T.; Wang, F.; Tan, T.W. Screening of high alpha-arbutin producing strains and production of alpha-arbutin by fermentation. World J. Microbiol. Biotechnol. 2013, 29, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Q.; Deng, L.; Zhing, P.; Zhang, S.R.; Xu, T.; Wang, F.; Tan, T.W. Efficient production of alpha-arbutin by whole-cell biocatalysis using immobilized hydroquinone as a glucosyl acceptor. J. Mol. Catal. B-Enzym. 2013, 91, 1–7. [Google Scholar] [CrossRef]
- Liu, C.Q.; Zhang, P.; Zhang, S.R.; Xu, T.; Wang, F.; Deng, L. Feeding strategies for the enhanced production of alpha-arbutin in the fed-batch fermentation of Xanthomonas maltophilia BT-112. Bioprocess. Biosyst. Eng. 2014, 37, 325–329. [Google Scholar] [CrossRef]
- Prodanovic, R.; Milosavic, N.; Sladic, D.; Zlatovic, M.; Bozic, B.; Velickovic, T.C.; Vujcic, Z. Transglucosylation of hydroquinone catalysed by alpha-glucosidase from baker’s yeast. J. Mol. Catal. B-Enzym. 2005, 35, 142–146. [Google Scholar] [CrossRef]
- Sato, T.; Hasegawa, N.; Saito, J.; Umezawa, S.; Honda, Y.; Kino, K.; Kirimura, K. Purification, characterization, and gene identification of an alpha-glucosyl transfer enzyme, a novel type alpha-glucosidase from Xanthomonas campestris WU-9701. J. Mol. Catal. B-Enzym. 2012, 80, 20–27. [Google Scholar] [CrossRef]
- Seo, D.H.; Jung, J.H.; Ha, S.J.; Cho, H.K.; Jung, D.H.; Kim, T.J.; Baek, N.I.; Yoo, S.H.; Park, C.S. High-yield enzymatic bioconversion of hydroquinone to alpha-arbutin, a powerful skin lightening agent, by amylosucrase. Appl. Microbiol. Biotechnol. 2012, 94, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.H.; Wang, Y.C.; Tian, Y.Q.; Xu, W.; Bai, Y.X.; Zhang, T.; Mu, W.M. Highly efficient biosynthesis of alpha-arbutin from hydroquinone by an amylosucrase from Cellulomonas carboniz. Process Biochem. 2018, 68, 93–99. [Google Scholar] [CrossRef]
- Kitao, S.; Sekine, H. Alpha-D-Glucosyl Transfer to Phenolic Compounds by Sucrose Phosphorylase from Leuconostoc mesenteroides and Production of alpha-Arbutin. Biosci. Biotechnol. Biochem. 1994, 58, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.S.; Kang, J.; Lee, J.H.; Kin, G.E.; Kim, G.J.; Kim, D. Synthesis and characterization of hydroquinone glucoside using Leuconostoc mesenteroides dextransucrase. Enzym. Microb. Technol. 2009, 45, 355–360. [Google Scholar] [CrossRef]
- Mathew, S.; Adlercreutz, P. Regioselective glycosylation of hydroquinone to α-arbutin by cyclodextrin glucanotransferase from Thermoanaerobacter sp. Biochem. Eng. J. 2013, 79, 187–193. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, Y.T.; Wei, X.M.; Yang, K.D.; Yang, X.K.; Wang, Y.T.; Xu, L.M.; Du, L.Q.; Huang, R.B. Sucrose Isomerase and Its Mutants from Erwinia rhapontici Can Synthesise alpha-Arbutin. Protein Pept. Lett. 2011, 18, 1028–1034. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, W.; Zhang, W.; Zhang, T.; Guang, C.; Mu, W. Amylosucrase as a transglucosylation tool: From molecular features to bioengineering applications. Biotechnol. Adv. 2018, 36, 1540–1552. [Google Scholar] [CrossRef]
- De Montalk, G.P.; Remaud-Simeon, M.; Willemot, R.M.; Sarcabal, P.; Planchot, V.; Monsan, P. Amylosucrase from Neisseria polysaccharea: Novel catalytic properties. FEBS Lett. 2000, 471, 219–223. [Google Scholar] [CrossRef]
- Skov, L.K.; Mirza, O.; Henriksen, A.; De Montalk, G.P.; Remaud-Simeon, M.; Sarcabal, P.; Willemot, R.M.; Monsan, P.; Gajhede, M. Amylosucrase, a glucan-synthesizing enzyme from the alpha-amylase family. J. Biol. Chem. 2001, 276, 25273–25278. [Google Scholar] [CrossRef]
- Seo, D.-H.; Jung, J.-H.; Ha, S.-J.; Yoo, S.-H.; Kim, T.-J.; Cha, J.; Park, C.-S. Molecular Cloning of the Amylosucrase Gene from a Moderate Thermophilic Bacterium Deinococcus Geothermalis and Analysis of its Dual Enzyme Activity. Carbohydr. Act. Enzym. 2008, 125–140. [Google Scholar] [CrossRef]
- Wu, P.H.; Giridhar, R.; Wu, W.T. Surface display of transglucosidase on Escherichia coli by using the ice nucleation protein of Xanthomonas campestris and its application in glucosylation of hydroquinone. Biotechnol. Bioeng. 2006, 95, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- King, A.T.; Davey, M.R.; Mellor, I.R.; Mulligan, B.J.; Lowe, K.C. Surfactant Effects on Yeast-Cells. Enzym. Microb. Technol. 1991, 13, 148–153. [Google Scholar] [CrossRef]
- Khan, S.; Altaf, I.; Rasheed, M.A.; Ashraf, M.; Ishaq, S.; Bashir, R.; Munir, N. Protection of vitamin A and E against toxicity induced by hydroquinone. Indian J. Exp. Biol. 2017, 55, 383–388. [Google Scholar]
- Shao, M.; Zhang, X.; Rao, Z.; Xu, M.; Yang, T.; Li, H.; Xu, Z.; Yang, S. Efficient testosterone production by engineered Pichia pastoris co-expressing human 17β-hydroxysteroid dehydrogenase type 3 and Saccharomyces cerevisiae glucose 6-phosphate dehydrogenase with NADPH regeneration. Green Chem. 2016, 18, 1774–1784. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, T.S.; Parajuli, P.; Pandey, R.P.; Sohng, J.K. Sustainable Production of Dihydroxybenzene Glucosides Using Immobilized Amylosucrase from Deinococcus geothermalis. J. Microbiol. Biotechnol. 2018, 28, 1447–1456. [Google Scholar] [PubMed]
- Ramshini, H.; Parrini, C.; Relini, A.; Zampagni, M.; Mannini, B.; Pesce, A.; Saboury, A.A.; Nemat-Gorgani, M.; Chiti, F. Large Proteins Have a Great Tendency to Aggregate but a Low Propensity to form Amyloid Fibrils. PLoS ONE 2011, 6, e16075. [Google Scholar] [CrossRef] [PubMed]
- Jhamb, K.; Sahoo, D.K. Production of soluble recombinant proteins in Escherichia coli: Effects of process conditions and chaperone co-expression on cell growth and production of xylanase. Bioresour. Technol. 2012, 123, 135–143. [Google Scholar] [CrossRef]
- Pinsach, J.; de Mas, C.; Lopez-Santin, J.; Striedner, G.; Bayer, K. Influence of process temperature on recombinant enzyme activity in Escherichia coli fed-batch Cultures. Enzym. Microb. Technol. 2008, 43, 507–512. [Google Scholar] [CrossRef]
- Zhu, X.; Tian, Y.; Xu, W.; Bai, Y.; Zhang, T.; Mu, W. Biochemical characterization of a highly thermostable amylosucrase from Truepera radiovictrix DSM 17093. Int. J. Biol. Macromol. 2018, 116, 744–752. [Google Scholar] [CrossRef]
- Chen, J.Y.; Kaleem, I.; He, D.M.; Liu, G.Y.; Li, C. Efficient production of glycyrrhetic acid 3-O-mono-beta-d-glucuronide by whole-cell biocatalysis in an ionic liquid/buffer biphasic system. Process Biochem. 2012, 47, 908–913. [Google Scholar] [CrossRef]
- Shen, D.X.; Shi, X.; Wang, Y.; Juan-Ling, F.U.; Zhou, Z.C. Inhibitory effect of thioredoxin on cytotoxicity of hydroquinone. Chin. J. Pharmacol. Toxicol. 2003, 17, 55–60. [Google Scholar]
- Wu, P.H.; Nair, G.R.; Chu, I.M.; Wu, W.T. High cell density cultivation of Escherichia coli with surface anchored transglucosidase for use as whole-cell biocatalyst for alpha-arbutin synthesis. J. Ind. Microbiol. Biotechnol. 2008, 35, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.D.; Cheng, H.J.; Ma, X.Q.; Xu, N.; Liu, J.; Ma, Y.H. Expression, characterization and mutagenesis of a novel glutamate decarboxylase from Bacillus megaterium. Biotechnol. Lett. 2016, 38, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ma, X.; Cheng, H.; Xu, N.; Liu, J.; Ma, Y. Co-expression of L-glutamate oxidase and catalase in Escherichia coli to produce alpha-ketoglutaric acid by whole-cell biocatalyst. Biotechnol. Lett. 2017, 39, 913–919. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Biotransformation Model | Donor | Biotransformation Period (h) | Final Concentration (g/L) | Molar Yield (%) | Productivity (g/L/h) | Reference |
|---|---|---|---|---|---|---|
| Biocatalysis | ||||||
| Xanthomonas campestris WU-9701 | Maltose | 36 | 11.43 | 93 | 0.32 | 5 |
| B. subtilis strain X-23 | Maltopentaose | 0.6 | 1.50 | 24.8 | 0.4 | 13 |
| Xanthomonas maltophilia BT-112 | Sucrose | 48 | 65.9 | 95.2 | 1.4 | 7 |
| Escherichia coli JM109 (Amylosucrase) | Sucrose | 8 | 60.9 | 95.5 | 7.6 | This study |
| Enzymatic catalysis | ||||||
| Amylosucrase (Cellulomonas carboniz T26 ) | Sucrose | 2 | 0.608 | 44.7 | 0.31 | 12 |
| Amylosucrase (Deinococcus geothermalis) | Sucrose | 24 | 5.78 | 90 | 0.24 | 11 |
| α-Glucosidase (Saccharomyces cerevisiae) | Maltose | 20 | 1 | 4.6 | 0.05 | 9 |
| Dextransucrase (Leuconostoc mesenteroides) | Sucrose | 6 | 0.544 | 0.4 | 0.09 | 14 |
| Fermentation | ||||||
| Escherichia coli BL21 (α-glucosidase) | Maltose | 36 | 21 | 76 | 0.58 | 25 |
| Xanthomonas maltophilia BT-112 | Sucrose | 72 | 61.7 | 94.5 | 0.86 | 8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Fan, W.; Zhang, R.; Shi, J.; Knežević-Jugović, Z.; Zhang, B. Study on Transglucosylation Properties of Amylosucrase from Xanthomonas campestris pv. Campestris and Its Application in the Production of α-Arbutin. Catalysts 2019, 9, 5. https://doi.org/10.3390/catal9010005
Yang C, Fan W, Zhang R, Shi J, Knežević-Jugović Z, Zhang B. Study on Transglucosylation Properties of Amylosucrase from Xanthomonas campestris pv. Campestris and Its Application in the Production of α-Arbutin. Catalysts. 2019; 9(1):5. https://doi.org/10.3390/catal9010005
Chicago/Turabian StyleYang, Chengyu, Weiming Fan, Ruijie Zhang, Jiping Shi, Zorica Knežević-Jugović, and Baoguo Zhang. 2019. "Study on Transglucosylation Properties of Amylosucrase from Xanthomonas campestris pv. Campestris and Its Application in the Production of α-Arbutin" Catalysts 9, no. 1: 5. https://doi.org/10.3390/catal9010005