
SIRT3: Oncogene and Tumor Suppressor in Cancer


1
Grupo Multidisciplinar de Oncología Traslacional, Institut Universitari d´Investigació en Ciències de la Salut (IUNICS), Universitat de les Illes Balears. Cra de Valldemossa, km 7.5, 07122 Palma, Illes Balears 07122, Spain
2
Ciber Fisiopatología Obesidad y Nutrición (CB06/03) Instituto Salud Carlos III, Madrid 28029, Spain
3
Instituto de Investigación Sanitaria de Baleares (IdISBa), Hospital Universitario, Son Espases, edificio S. E-07120 Palma, Palma de Mallorca, Illes Balears 07120, Spain
*
Author to whom correspondence should be addressed.
Cancers 2017, 9(7), 90; https://doi.org/10.3390/cancers9070090
Submission received: 4 May 2017
/
Revised: 2 July 2017
/
Accepted: 7 July 2017
/
Published: 12 July 2017
{kind=link}
{kind=link}
Abstract
:Sirtuin 3 (SIRT3), the major deacetylase in mitochondria, plays a crucial role in modulating oxygen reactive species (ROS) and limiting the oxidative damage in cellular components. SIRT3 targets different enzymes which regulate mitochondrial metabolism and participate in ROS detoxification, such as the complexes of the respiratory chain, the isocitrate dehydrogenase, or the manganese superoxide dismutase. Thus, SIRT3 activity is essential in maintaining mitochondria homeostasis and has recently received great attention, as it is considered a fidelity protein for mitochondrial function. In some types of cancer, SIRT3 functions as a tumoral promoter, since it keeps ROS levels under a certain threshold compatible with cell viability and proliferation. On the contrary, other studies describe SIRT3 as a tumoral suppressor, as SIRT3 could trigger cell death under stress conditions. Thus, SIRT3 could have a dual role in cancer. In this regard, modulation of SIRT3 activity could be a new target to develop more personalized therapies against cancer.
1. Introduction
Sirtuins are a family of NAD+-dependent deacetylases which regulate signaling pathways involved in cellular proliferation and differentiation, metabolism, response to stress, and cancer [1,2]. Seven isoforms have been described in mammals, and sirtuins 3, 4, and 5 are exclusively located in the mitochondria [3,4,5]. Among these, sirtuin 3 (SIRT3) has recently gained great attention, as it presents the most robust mitochondrial deacetylase activity and targets key proteins for the proper function and metabolism of these organelles [6].
SIRT3 is synthesized as a 44 kDa peptide with an N-terminal sequence as a targeting signal for mitochondrial localization. Under stress conditions, SIRT3 translocates to the mitochondrial matrix, where is activated by a protease, which yield the 28-kDa active form of SIRT3 [7,8,9]. Recent studies point out a possible nuclear localization of SIRT3 in the nucleus, where it may regulate gene expression in response to stress [7,10].
2. SIRT3 as a Mitochondrial Fidelity Protein
Mitochondria are crucial organelles for several cellular processes, such as energy production through oxidative phosphorylation, cell cycle and proliferation, apoptosis, etc. [2,11]. Nevertheless, the mitochondrial respiratory chain is responsible for the production of reactive oxygen species or oxygen reactive species (ROS) [12]. When ROS production is controlled and ROS levels are under a certain threshold, they act as second messengers and stimulate cellular proliferation. However, high ROS levels can be harmful to the cell, damaging proteins, lipids, and DNA, thus contributing to mitochondrial dysfunction and carcinogenesis [13,14,15,16].
For this reason, fidelity proteins are essential for the maintenance and proper function of mitochondria [17,18,19]. Around 35% of mitochondrial proteins are estimated to be regulated by acetylation, and the majority of them are involved in energy metabolism. This observation suggests that acetylation may be a critical post-translational modification in these organelles [11,20,21].
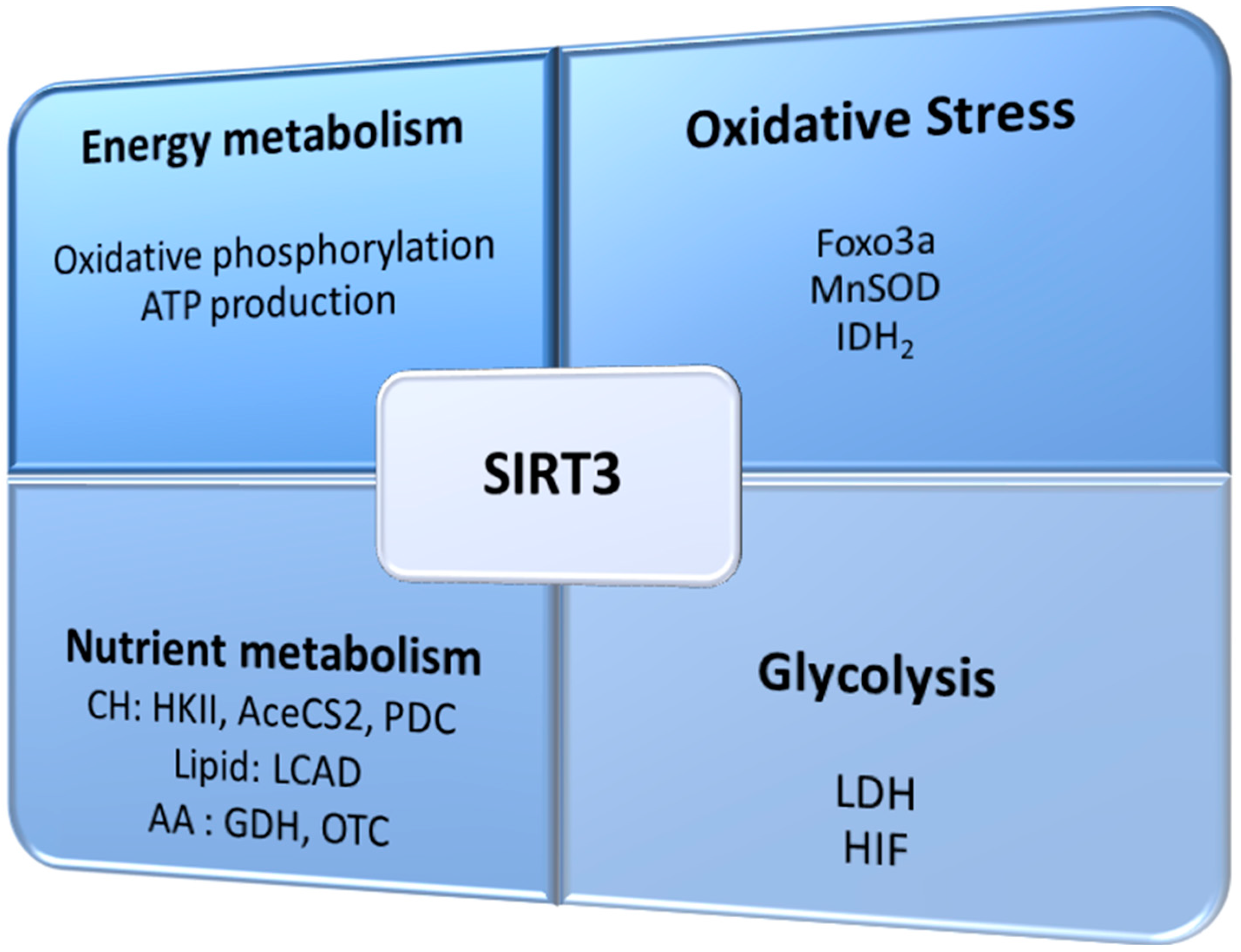
SIRT3 has been reported to deacetylate and activate proteins that are essential against oxidative stress, including antioxidant enzymes and enzymes involved in mitochondrial function and ATP synthesis [22,23,24]. Loss of SIRT3 function is linked to the development of a permissive phenotype for carcinogenesis, as mitochondrial proteins become hyperacetylated and cells show elevated oxidative stress, which leads to mitochondrial dysfunction [5,11]. Furthermore, some studies in tumors report a correlation between SIRT3 expression and the evolution of several types of cancer [25,26,27,28,29,30,31]. As a stress-responsive protein, SIRT3 function is to regulate ROS production to avoid damage to cellular components [5,32]. Thus, SIRT3 is crucial in the maintenance of mitochondrial function and integrity of these organelles, and has been referred to as the guardian of the mitochondria, similarly to the consideration of p53 as the guardian of the genome [33]. The main processes in which SIRT3 takes part, directly and indirectly, are schematized in Figure 1.
3. SIRT3 Regulates Oxidative Stress and Cellular Metabolism
As mentioned above, SIRT3 is key for the mitochondrial integrity and proper function under stress conditions [2,34]. Several studies show that mice lacking SIRT3 show no important affections under basal conditions, although after fasting, exercise or calorie restriction these mice develop several diseases, including some types of cancer. These conditions have an increase in ROS production in common that leads to oxidative stress, which is proposed to be the cause of disease [18,35,36].
SIRT3 limits ROS production as well as reduces the resulting oxidative damage. In this regard, SIRT3 is considered a key ROS scavenger in the cell, since it targets two main oxidative stress-responsive proteins: manganese superoxide dismutase (MnSOD) and isocitrate dehydrogenase (IDH) [37,38,39,40]. MnSOD is a mitochondrial antioxidant enzyme crucial for the cell, as it converts highly reactive superoxide ions into hydrogen peroxide, which is later neutralized to water by catalase and other mitochondrial peroxidases [38,41,42]. On the other hand, deacetylation of IDH increases its activity, stimulating the conversion of isocitrate to α-ketoglutarate. This reaction generates nicotinamide adenine dinucleotide phosphate (NADPH), which is required for the regeneration of reduced glutathione, another main antioxidant of the cell [43,44,45]. Some recent studies also indicate that SIRT3 could affect gene expression of these antioxidant enzymes, presumably through deacetylation and activation of transcriptional factor FoxO3a, which regulates essential proteins for mitochondrial homeostasis such as MnSOD and catalase [39,44,46,47,48].
SIRT3 also modulates energy metabolism by directly deacetylating and activating several enzymes involved in regulation of mitochondrial metabolism, including the Krebs cycle, the respiratory chain and the oxidation of fatty acids [6,49], contributing to proper mitochondrial function. This way, SIRT3 induces higher oxidative phosphorylation and ATP production [50].
For instance, SIRT3 deacetylates and inactivates cyclophilin D, which results in the detachment of hexokinase II from mitochondria, thereby inhibiting glycolysis [51]. Furthermore, SIRT3 can increase acetyl-CoA availability for the tricarboxylic acid cycle (TCA) in mitochondria activating acetyl-CoA synthase 2 (AceCS2) [52,53] and some activator enzymes of the pyruvate dehydrogenase complex (PDC) [54]. A recent study suggests that SIRT3 could also deacetylate and enhance the activity of lactate dehydrogenase (LDH), promoting anaerobic glycolysis [55].
SIRT3 also participates in the regulation of lipid metabolism through deacetylation and activation of long chain acyl-CoA dehydrogenase (LCAD), thus stimulating fatty acid oxidation [56]. It has also been described that SIRT3 enhances the synthesis of β-hydroxybutyrate by activating 2-hydroxy-3-methylglutaryl CoA synthase 2 [57].
Glutamate dehydrogenase (GDH) is another promising target of SIRT3. GDH is involved in amino acid metabolism, driving them into the Krebs cycle to be used as fuel, and is affected in tumoral cells, contributing to its survival and proliferation [37,58]. SIRT3 can also promote the urea cycle by deacetylating and activating ornithine transcarbamylase (OTC) under energy restriction [59].
SIRT3 also takes part in the coordination of the TCA cycle, the electron transport chain (ETC) and ATP synthesis through deacetylation and activation of different complexes of the oxidative phosphorylation [50]. For instance, it has been reported that SIRT3 deacetylates and activates different subunits of all ETC complexes [50,60,61,62,63] and ATPase [64], thus enhancing mitochondrial function. Furthermore, SIRT3 activates liver kinase B1 (LKB1), which in turn activates AMP-activated protein kinase (AMPK), leading to ATP synthesis [65].
SIRT3 is involved in mitochondrial biogenesis and mitochondrial dynamics. Under stress conditions, the peroxisome proliferator-activated receptor γ co-activator 1-α (PGC-1α) along with the estrogen receptor-related alpha (ERRα), involved in mitochondrial biogenesis, can regulate expression and protein levels of SIRT3 [66,67]. On the other hand, SIRT3 indirectly participates in PGC1-α expression through activating AMPK signaling pathway, which results in cAMP response element-binding protein (CREB) phosphorylation and increased PGC1-α gene expression [68]. The establishment of this positive feedback loop is crucial for the regulation of mitochondrial biogenesis and function, activating mitochondrial enzymes involved in antioxidant defenses and metabolism [63,66,68]. Finally, SIRT3 also deacetylates and activates optic atrophy 1 (OPA1), which is involved in mitochondrial fusion [69].
SIRT3 also contributes to mitochondrial quality control. For instance, SIRT3 has been described to coordinate the mitochondrial unfolded protein response and upregulate mitophagy machinery [70]. Furthermore, SIRT3 can suppress the formation and opening of the mitochondrial permeability transition pore, preventing mitochondrial dysfunction [71]. SIRT3 also deacetylates 8-oxoguanine glycosylase 1 (OGG1), which prevents its degradation. This way, OGG1 contributes to mitochondrial DNA repair, protecting integrity of mitochondria, and preventing apoptosis [72].
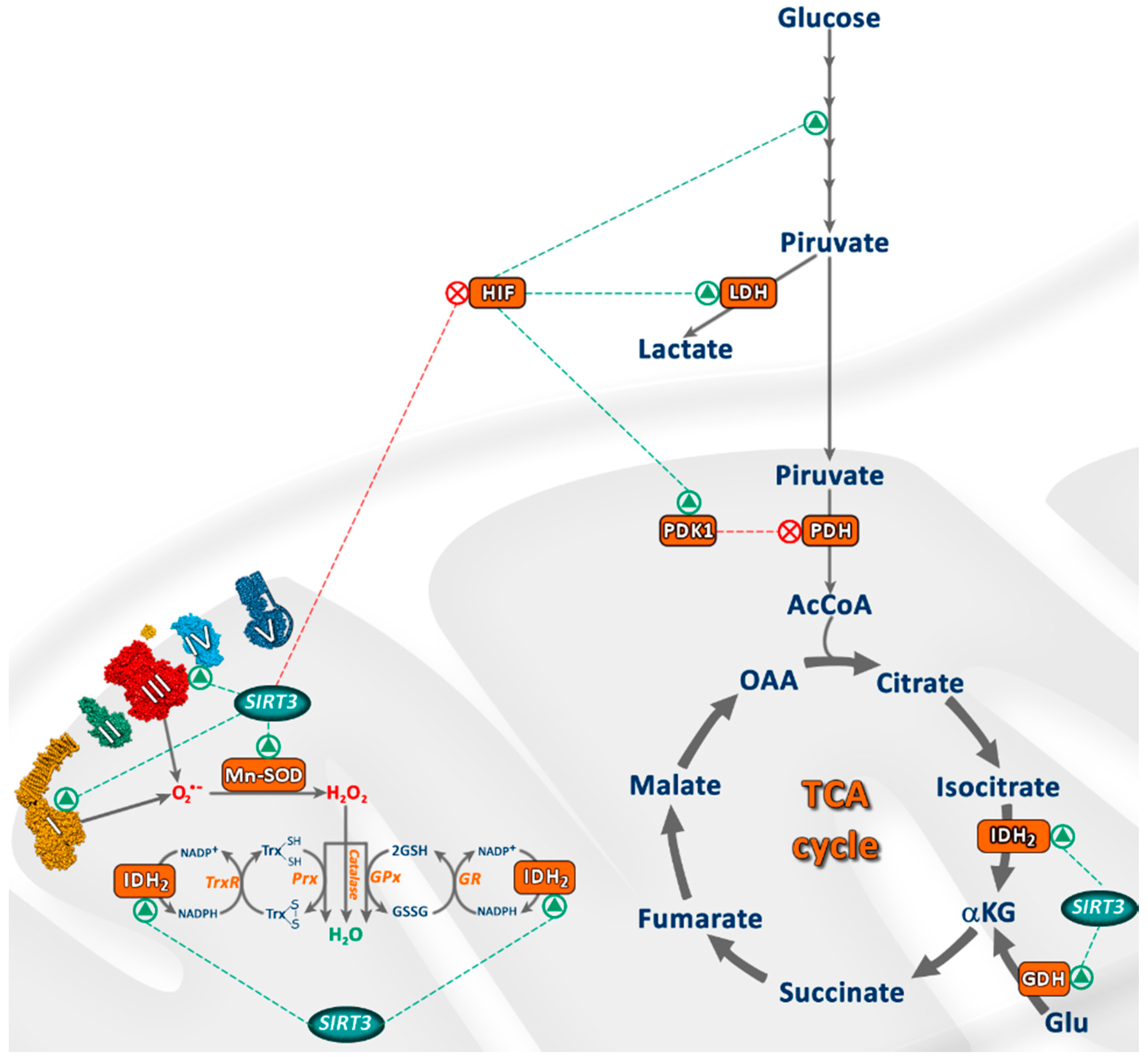
These observations indicate the role of SIRT3 in energy metabolism, ATP synthesis, mitochondrial function and mitochondrial ROS scavenging, which are crucial for the proper function of mitochondria. The main targets of SIRT3 are shown in Figure 2.
Metabolism can be crucial in cancer development and progression, as tumoral cells undergo a metabolic reprogramming to meet energy demands for continued growth and proliferation, known as the Warburg effect [73,74]. For this reason, cancer cells show a shift from oxidative phosphorylation to glycolysis as the main metabolic process to obtain ATP [55,75,76]. Apart from a rapid energy supply, cells also obtain different intermediates to meet the requirements for the synthesis of macromolecules and sustain cell division [77,78].
SIRT3 plays a pivotal role in this metabolic reprogramming through regulation of all the targets mentioned before. Moreover, SIRT3 leads to destabilization and inactivation of hypoxia inducible factor 1-alpha (HIF-1 α). This nuclear factor induces expression of genes involved in glycolysis, directly promoting the Warburg effect, and genes involved in angiogenesis, contributing to tumor evolution [1,35,36]. Thus, loss of SIRT3 is associated to high oxidative stress and ROS production, as well as to metabolic reprogramming, which contributes to carcinogenesis.
4. SIRT3 Is Involved in Proliferation and Apoptosis Pathways
Besides its function in metabolism regulation, SIRT3 has been referred to as an oncogene in some types of cancer, as SIRT3 can maintain ROS production at the appropriate levels to prevent apoptosis and promote cell proliferation [18,33,70]. Furthermore, SIRT3 regulates mitochondrial homeostasis and preserves mitochondrial membrane integrity, thus increasing cellular resistance to oxidative stress [24,51,72]. In accordance to this proliferative function, in cervical cancer cells SIRT3 has been reported to interact with the protein Ku70, involved in DNA repair, which results in apoptosis avoidance under stress [79]. Additionally, p53 has been recently described as a target of SIRT3. In bladder cancer, deacetylation of p53 induces cellular proliferation, rescuing cells from growth arrest caused by p53 [80]. Some reports suggest that SIRT3 could have a crucial role in the development and progression of some types of cancer acting as a tumour promoter, such as breast cancer [46], colon cancer [30], gastric cancer [55], esophageal cancer [81], oral squamous cell carcinoma [82], melanoma [83], and renal cancer [58]. These studies show a correlation between high SIRT3 expression and poorer clinical prognosis. Furthermore, SIRT3 knockdown sensitizes cancer cells to cytotoxic treatments [46,82] and reduces cell proliferation [46,82,83].
On the contrary, some studies suggest a tumour suppressor role for SIRT3. It has been described that SIRT3 induces cell arrest and apoptosis, by regulating proteins such as Bcl-2, p53 of HIF-1α [35,80,84,85]. In agreement with this protective function, mice lacking SIRT3 develop tumors more quickly than control mice. Additionally, tumors lacking SIRT3 grow faster and show greater volume than control tumors in xenograft models [18,35]. SIRT3 has been described as a tumor suppressor in breast cancer [25], hepatocellular carcinoma [86,87,88], B cell malignancies and leukemia [85,89], and metastatic ovarian cancer [90]. In these reports, SIRT3 expression correlates with a good outcome and a general increase in overall survival of cancer patients. The mechanisms described include inhibition of proliferation when SIRT3 is overexpressed [86,89,91], and limiting metabolic reprogramming [51,89].
5. Conclusions
As it has been discussed throughout this review, SIRT3 seems to prevent cell death under oxidative stress, while other studies report a pro-apoptotic function for this deacetylase. Therefore, SIRT3 shows a dual role in cancer, as it can act as a tumour suppressor or a tumour promoter, depending on the cellular context. Cancer cells usually show higher ROS levels than normal cells, which confers advantages in tumor promotion and progression, as well as resistance to chemotherapy. However, some anticancer therapies are based on its ability to further increase ROS production to reach toxic levels, resulting in cell death and overcoming treatment resistance [92,93].
For this reason, SIRT3 rises as a possible target to develop new therapeutic strategies against cancer. For instance, in breast cancer SIRT3 could confer resistance to tamoxifen, a commonly used therapy to block the estrogen receptor in this type of cancer [29]. In this way, some studies report that SIRT3 silencing results in a decreased cellular proliferation and induces cell death, hence it could improve chemotherapy efficacy for some cancers [4,46,70]. Therefore, modulation of SIRT3 activity could be an approach to improve therapies against cancer, especially to overcome acquired resistance to treatment.
Acknowledgments
This work was supported by a grant from Fondo de Investigaciones Sanitarias of Instituto de Salud Carlos III (PI14/01434) of the Spanish Government, cofinanced by FEDER-Unión Europea (“Una manera de hacer Europa”). Margalida Torrens-Mas was funded by an FPU (Formación profesorado universitario) grant of Ministerio de Educación, Cultura y Deporte of Spanish Government.
Conflicts of Interest
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
References
- Haigis, M.C.; Deng, C.-X.; Finley, L.W.S.; Kim, H.-S.; Gius, D. SIRT3 is a mitochondrial tumor suppressor: A scientific tale that connects aberrant cellular ROS, the Warburg effect, and carcinogenesis. Cancer Res. 2012, 72, 2468–2472. [Google Scholar] [CrossRef] [PubMed]
- Sack, M.N.; Finkel, T. Mitochondrial metabolism, sirtuins, and aging. Cold Spring Harbor Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Michan, S.; Sinclair, D. Sirtuins in mammals: Insights into their biological function. Biochem. J. 2007, 404, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Alhazzazi, T.Y.; Kamarajan, P.; Verdin, E.; Kapila, Y.L. SIRT3 and cancer: Tumor promoter or suppressor? Biochim. Biophys. Acta 2011, 1816, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Hirschey, M.D.; Shimazu, T.; Huang, J.-Y.; Schwer, B.; Verdin, E. SIRT3 regulates mitochondrial protein acetylation and intermediary metabolism. Cold Spring Harbor Symp. Quant. Biol. 2011, 76, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Verdin, E.; Hirschey, M.D.; Finley, L.W.S.; Haigis, M.C. Sirtuin regulation of mitochondria: Energy production, apoptosis, and signaling. Trends Biochem. Sci. 2010, 35, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Scher, M.B.; Vaquero, A.; Reinberg, D. SirT3 is a nuclear NAD+-dependent histone deacetylase that translocates to the mitochondria upon cellular stress. Genes Dev. 2007, 21, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Schwer, B.; North, B.J.; Frye, R.A.; Ott, M.; Verdin, E. The human silent information regulator (Sir)2 homologue hSIRT3 is a mitochondrial nicotinamide adenine dinucleotide-dependent deacetylase. J. Cell Biol. 2002, 158, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.M.; Huang, J.-Y.; Verdin, E.; Spelbrink, J.N. A new splice variant of the mouse SIRT3 gene encodes the mitochondrial precursor protein. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Iwahara, T.; Bonasio, R.; Narendra, V.; Reinberg, D. SIRT3 functions in the nucleus in the control of stress-related gene expression. Mol. Cell. Biol. 2012, 32, 5022–5034. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.A.; Hirschey, M.D. Mitochondrial protein acetylation regulates metabolism. Essays Biochem. 2012, 52, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Laurent, A.; Nicco, C.; Chéreau, C.; Goulvestre, C.; Alexandre, J.; Alves, A.; Lévy, E.; Goldwasser, F.; Panis, Y.; Soubrane, O.; et al. Controlling tumor growth by modulating endogenous production of reactive oxygen species. Cancer Res. 2005, 65, 948–956. [Google Scholar] [PubMed]
- Kongara, S.; Karantza, V. The interplay between autophagy and ROS in tumorigenesis. Front. Oncol. 2012, 2, 171. [Google Scholar] [CrossRef] [PubMed]
- Waris, G.; Ahsan, H. Reactive oxygen species: Role in the development of cancer and various chronic conditions. J. Carcinog. 2006, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Sainz, R.M.; Lombo, F.; Mayo, J.C. Radical decisions in cancer: Redox control of cell growth and death. Cancers 2012, 4, 442–474. [Google Scholar] [CrossRef] [PubMed]
- Pelicano, H.; Carney, D.; Huang, P. ROS stress in cancer cells and therapeutic implications. Drug Resist. Updat. 2004, 7, 97–110. [Google Scholar] [CrossRef]
- Kim, H.S.; Patel, K.; Muldoon-Jacobs, K.; Bisht, K.S.; Aykin-Burns, N.; Pennington, J.D.; van der Meer, R.; Nguyen, P.; Savage, J.; Owens, K.M.; et al. SIRT3 is a mitochondria-localized tumor suppressor required for maintenance of mitochondrial integrity and metabolism during stress. Cancer Cell 2010, 17, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, C.; Kumar, C.; Gnad, F.; Nielsen, M.L.; Rehman, M.; Walther, T.C.; Olsen, J.V.; Mann, M. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 2009, 325, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Sprung, R.; Chen, Y.; Xu, Y.; Ball, H.; Pei, J.; Cheng, T.; Kho, Y.; Xiao, H.; Xiao, L.; et al. Substrate and functional diversity of lysine acetylation revealed by a proteomics survey. Mol. Cell. 2006, 23, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Hallows, W.C.; Albaugh, B.N.; Denu, J.M. Where in the cell is SIRT3?—Functional localization of an NAD+-dependent protein deacetylase. Biochem. J. 2008, 411. [Google Scholar] [CrossRef] [PubMed]
- Buler, M.; Aatsinki, S.-M.; Izzi, V.; Hakkola, J. Metformin reduces hepatic expression of SIRT3, the mitochondrial deacetylase controlling energy metabolism. PLoS ONE 2012, 7, e49863. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, L.; Li, C.Y.; Pei, Z.; Zhou, M.; Li, N. SIRT3 protects cells from hypoxia via PGC-1α- and MnSOD-dependent pathways. Neuroscience 2015, 286, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Desouki, M.M.; Doubinskaia, I.; Gius, D.; Abdulkadir, S.A. Decreased mitochondrial SIRT3 expression is a potential molecular biomarker associated with poor outcome in breast cancer. Hum. Pathol. 2014, 45. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, N.; Zino, S.; Macintyre, A.; Kingsmore, D.; Payne, P.; George, W.D.; Shiels, P.G. Altered sirtuin expression is associated with node-positive breast cancer. Br. J. Cancer. 2006, 95, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- He, S.; He, C.; Yuan, H.; Xiong, S.; Xiao, Z.; Chen, L. The SIRT3 expression profile is associated with pathological and clinical outcomes in human breast cancer patients. Cell Physiol. Biochem. 2014, 34, 2061–2069. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fu, L.L.; Wen, X.; Wang, X.Y.; Liu, J.; Cheng, Y.; Huang, J. Sirtuin-3 (SIRT3), a therapeutic target with oncogenic and tumor-suppressive function in cancer. Cell Death Dis. 2014, 5, e1047. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ren, X.; Cheng, Y.; Huber-Keener, K.; Liu, X.; Zhang, Y.; Yuan, Y.-S.; Yang, J.-W.; Liu, C.-G.; Yang, J.-M. Identification of Sirtuin 3, a mitochondrial protein deacetylase, as a new contributor to tamoxifen resistance in breast cancer cells. Biochem. Pharmacol. 2013, 86, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Huang, Z.; Jiang, H.; Shi, F. The Sirtuin 3 expression profile is associated with pathological and clinical outcomes in colon cancer patients. Biomed. Res. Int. 2014, 2014, 87126. [Google Scholar] [CrossRef] [PubMed]
- Sastre-Serra, J.; Nadal-Serrano, M.; Pons, D.G.; Valle, A.; Garau, I.; Garcia-Bonafe, M; Oliver, J.; Roca, P. The oxidative stress in breast tumors of postmenopausal women is ERalpha/ERbeta ratio dependent. Free Radic. Biol. Med. 2013, 61, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Mas, M.; Roca, P.; Sastre-Serra, J. Role of sirtuin 3 in oxidative stress and cancer. Med. Balear 2017, 32, 47–52. [Google Scholar]
- Park, S.-H.; Ozden, O.; Jiang, H.; Cha, Y.I.; Pennington, J.D.; Aykin-Burns, N.; Spitz, D.R.; Gius, D.; Kim, H.-S. Sirt3, mitochondrial ROS, ageing, and carcinogenesis. Int. J. Mol. Sci. 2011, 12, 6226–6239. [Google Scholar] [CrossRef] [PubMed]
- Alhazzazi, T.Y.; Kamarajan, P.; Verdin, E.; Kapila, Y. Sirtuin-3 (SIRT3) and the hallmarks of cancer. Genes Cancer 2013, 4, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Finley, L.W.S.; Carracedo, A.; Lee, J.; Souza, A.; Egia, A.; Zhang, J.; Teruya-Feldstein, J.; Moreira, P.I.; Cardoso, S.M.; Clish, C.B.; et al. SIRT3 opposes reprogramming of cancer cell metabolism through HIF1α destabilization. Cancer Cell 2011, 19, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.L.; Emerling, B.M.; Ricoult, S.J.H.; Guarente, L. SirT3 suppresses hypoxia inducible factor 1α and tumor growth by inhibiting mitochondrial ROS production. Oncogene 2011, 30, 2986–2996. [Google Scholar] [CrossRef] [PubMed]
- Schlicker, C.; Gertz, M.; Papatheodorou, P.; Kachholz, B.; Becker, C.F.W.; Steegborn, C. Substrates and regulation mechanisms for the human mitochondrial sirtuins Sirt3 and Sirt5. J. Mol. Biol. 2008, 382, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Coleman, M.C.; Pennington, J.D.; Ozden, O.; Park, S.-H.; Jiang, H.; Kim, H.-S.; Flynn, C.R.; Hill, S.; McDonald, W.H.; et al. Sirt3-mediated deacetylation of evolutionarily conserved lysine 122 regulates MnSOD activity in response to stress. Mol. Cell 2010, 40, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Bause, A.S.; Haigis, M.C. SIRT3 regulation of mitochondrial oxidative stress. Exp. Gerontol. 2013, 48, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yan, Y.; Principe, D.R.; Zou, X.; Vassilopoulos, A.; Gius, D. SIRT3 and SIRT4 are mitochondrial tumor suppressor proteins that connect mitochondrial metabolism and carcinogenesis. Cancer Metab. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Ozden, O.; Park, S.H.; Kim, H.S.; Jiang, H.; Coleman, M.C.; Spitz, D.R.; Gius, D. Acetylation of MnSOD directs enzymatic activity responding to cellular nutrient status or oxidative stress. Aging 2011, 3, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Santa-Maria, C.A.; O’Brien, J.; Gius, D.; Zhu, Y. Manganese superoxide dismutase acetylation and dysregulation, due to loss of SIRT3 activity, promote a Luminal B-like breast carcinogenic-permissive phenotype. Antioxid. Redox Signal. 2016, 25, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Someya, S.; Yu, W.; Hallows, W.C.; Xu, J.; Vann, J.M.; Leeuwenburgh, C.; Tanokura, M.; Denu, J.M.; Prolla, T.A. Sirt3 mediates reduction of oxidative damage and prevention of age-related hearing loss under caloric restriction. Cell 2010, 143, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, N.R.; Gupta, M.; Kim, G.; Rajamohan, S.B.; Isbatan, A.; Gupta, M.P. Sirt3 blocks the cardiac hypertrophic response by augmenting Foxo3a-dependent antioxidant defense mechanisms in mice. J. Clin. Investig. 2009, 119, 2758–2771. [Google Scholar] [CrossRef] [PubMed]
- Weir, H.J.M.; Lane, J.D.; Balthasar, N. SIRT3: A central regulator of mitochondrial adaptation in health and disease. Genes Cancer 2013, 4, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Mas, M.; Pons, D.G.; Sastre-Serra, J.; Oliver, J.; Roca, P. SIRT3 silencing sensitizes breast cancer cells to cytotoxic treatments through an increment in ROS production. J. Cell. Biochem. 2016, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tseng, A.H.H.; Shieh, S.S.; Wang, D.L. SIRT3 deacetylates FOXO3 to protect mitochondria against oxidative damage. Free Radic. Biol. Med. 2013, 63, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, P.P.; Karthikeyan, A.; Lu, J.; Ling, E.-A.; Dheen, S.T. Sirtuin 3 regulates Foxo3a-mediated antioxidant pathway in microglia. Neuroscience 2015, 311, 398–414. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-Y.; Hirschey, M.D.; Shimazu, T.; Ho, L.; Verdin, E. Mitochondrial sirtuins. Biochim. Biophys. Acta Proteins Proteomics 2010, 1804, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.-H.; Kim, H.-S.; Song, S.; Lee, I.H.; Liu, J.; Vassilopoulos, A.; Deng, C.-X.; Finkel, T. A role for the mitochondrial deacetylase Sirt3 in regulating energy homeostasis. Proc. Natl. Acad. Sci. USA 2008, 105, 14447–14452. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Zhou, Y.; Dai, Q.; Qiao, C.; Zhao, L.; Hui, H.; Lu, N.; Gio, L.-Q. Oroxylin A induces dissociation of hexokinase II from the mitochondria and inhibits glycolysis by SIRT3-mediated deacetylation of cyclophilin D in breast carcinoma. Cell Death Dis. 2013, 4, e601. [Google Scholar] [CrossRef] [PubMed]
- Hallows, W.C.; Lee, S.; Denu, J.M. Sirtuins deacetylate and activate mammalian acetyl-CoA synthetases. Proc. Natl. Acad. Sci. USA 2006, 103, 10230–10235. [Google Scholar] [CrossRef] [PubMed]
- Schwer, B.; Bunkenborg, J.; Verdin, R.O.; Andersen, J.S.; Verdin, E. Reversible lysine acetylation controls the activity of the mitochondrial enzyme acetyl-CoA synthetase 2. Proc. Natl. Acad. Sci. USA 2006, 103, 10224–10229. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Kang, H.B.; Elf, S.; Xie, J.; Gu, T.L.; Aguiar, M.; Lonning., S.; Hitosugi, T.; Chung, T.-W.; Arellano, M.; et al. Tyr-94 phosphorylation inhibits pyruvate dehydrogenase phosphatase 1 and promotes tumor growth. J. Biol. Chem. 2014, 289, 21413–21422. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Qin, L.; Wu, J.; Qu, X.; Hou, C.; Sun, W.; Li, S.; Vaughan, A.T.M.; Li, J.J.; Liu, J. SIRT3 enhances glycolysis and proliferation in SIRT3-expressing gastric cancer cells. PLoS ONE 2015, 10, e0129834. [Google Scholar] [CrossRef] [PubMed]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.O.; Ilkayeva, R. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 2010, 464, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, T.; Hirschey, M.D.; Hua, L.; Dittenhafer-Reed, K.E.; Schwer, B.; Lombard, D.B.; Li, Y.; Bunkenborg, J.; Alt, F.W.; Denu, J.M.; Jacobson, M.P.; et al. SIRT3 deacetylates mitochondrial 3-hydroxy-3-methylglutaryl CoA synthase 2 and regulates ketone body production. Cell Metab. 2010, 12, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Koh, E.; Lee, Y.S.; Lee, H.-W.; Kang, H.G.; Yoon, Y.E.; Hanb, W.K.; Choic, K.H.; Kimet, K.-S. Mitochondrial Sirt3 supports cell proliferation by regulating glutamine-dependent oxidation in renal cell carcinoma. Biochem. Biophys. Res. Commun. 2016, 474, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Allows, W.C.; Yu, W.; Smith, B.C.; Devires, M.K.; Ellinger, J.J.; Someya, S.; Shortreed, M.R.; Prolla, T.; Markley, J.L.; Smith, L.M.; et al. Sirt3 promotes the urea cycle and fatty acid oxidation during dietary restriction. Mol. Cell 2011, 41, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Cimen, H.; Han, M.J.; Yang, Y.; Tong, Q.; Koc, H.; Koc, E.C. Regulation of succinate dehydrogenase activity by SIRT3 in mammalian mitochondria. Biochemistry 2010, 49, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Finley, L.W.S.; Haas, W.; Desquiret-Dumas, V.; Wallace, D.C.; Procaccio, V.; Gygi, S.P.; Haigis, M.C. Succinate dehydrogenase is a direct target of sirtuin 3 deacetylase activity. PLoS ONE 2011, 6, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Jing, E.; Emanuelli, B.; Hirschey, M.D.; Boucher, J.; Lee, K.Y.; Lombard, D.; Haigis, M.C. Sirtuin-3 (Sirt3) regulates skeletal muscle metabolism and insulin signaling via altered mitochondrial oxidation and reactive oxygen species production. Proc. Natl. Acad. Sci. USA 2011, 108, 14608–14613. [Google Scholar] [CrossRef] [PubMed]
- Abdel Khalek, W.; Cortade, F.; Ollendorff, V.; Lapasset, L.; Tintignac, L.; Chabi, B.; Verdin, E.M.; Kahn, C.R. SIRT3, a mitochondrial NAD+-dependent deacetylase, is involved in the regulation of myoblast differentiation. PLoS ONE 2014, 9, e114388. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lee, H.; Liao, C.; Wei, Y. Regulation of mitochondrial FoF1ATPase activity by Sirt3-catalyzed deacetylation and its de ficiency in human cells harboring 4977 bp deletion of mitochondrial DNA. BBA Mol. Basis Dis. 2013, 1832, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Pillai, V.B.; Sundaresan, N.R.; Kim, G.; Gupta, M.; Rajamohan, S.B.; Pillai, J.B.; Samant, S.; Ravindra, P.V.; Isbatan, A.; Gupta, M.P. Exogenous NAD blocks cardiac hypertrophic response via activation of the SIRT3-LKB1-AMP-activated kinase pathway. J. Biol. Chem. 2010, 285, 3133–3144. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wang, R.; Xue, Y.; Liu, X.; Zhang, H.; Chen, Y.; Fang, F.; Chang, Y. Sirtuin 3, a new target of PGC-1alpha, plays an important role in the suppression of ROS and mitochondrial biogenesis. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Giralt, A.; Hondares, E.; Villena, J.A.; Ribas, F.; Díaz-Delfín, J.; Giralt, M.; Villarroya, F. Peroxisome proliferator-activated receptor-gamma coactivator-1alpha controls transcription of the Sirt3 gene, an essential component of the thermogenic brown adipocyte phenotype. J. Biol. Chem. 2011, 286, 16958–16966. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Wang, F.; Stieren, E.; Tong, Q. SIRT3, a mitochondrial sirtuin deacetylase, regulates mitochondrial function and thermogenesis in brown adipocytes. J. Biol. Chem. 2005, 280, 13560–13567. [Google Scholar] [CrossRef] [PubMed]
- Samant, S.A.; Zhang, H.J.; Hong, Z.; Pillai, V.B.; Sundaresan, N.R.; Wolfgeher, D.; Archerc, S.L.; Chanf, D.C.; Guptaa, M.P. SIRT3 deacetylates and activates OPA1 to regulate mitochondrial dynamics during stress. Mol. Cell. Biol. 2014, 34, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Papa, L.; Germain, D. SirT3 regulates the mitochondrial unfolded protein response. Mol. Cell. Biol. 2014, 34, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Hafner, A.V.; Dai, J.; Gomes, A.P.; Xiao, C.Y.; Palmeira, K.C.M.; Rosenzweig, A. Regulation of the mPTP by SIRT3-mediated deacetylation of CypD at lysine 166 suppresses age-related cardiac hypertrophy. Aging (Albany. NY). 2010, 2, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Ren, X.; Gowda, S.P.; Shan, Y.; Zhang, L.; Yuan, Y.-S.; Patel1, R.; Wu, H.; Huber-Keener, K.; Yang, J.W.; et al. Interaction of Sirt3 with OGG1 contributes to repair of mitochondrial DNA and protects from apoptotic cell death under oxidative stress. Cell Death Dis. 2013, 4, e731. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.S.; Thompson, C.B. Metabolic reprogramming: A cancer hallmark even warburg did not anticipate. Cancer Cell 2012, 21, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.M.R. Cancer metabolism: The Warburg effect today. Exp. Mol. Pathol. 2010, 89, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Lum, J.J.; Hatzivassiliou, G.; Thompson, C.B. The biology of cancer: Metabolic reprogramming fuels cell growth and proliferation. Cell Metab. 2008, 7, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Heiden, M.G.; Cantley, L.C.; Thompson, C.B.; Mammalian, P.; Exhibit, C.; Metabolism, A. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, N.R.; Samant, S.A.; Pillai, V.B.; Rajamohan, S.B.; Gupta, M.P. SIRT3 is a stress-responsive deacetylase in cardiomyocytes that protects cells from stress-mediated cell death by deacetylation of Ku70. Mol. Cell. Biol. 2008, 28, 6384–6401. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Banck, M.; Mujtaba, S.; Zhou, M.M.; Sugrue, M.M.; Walsh, M.J. p53-Induced growth arrest is regulated by the Mitochondrial SirT3 deacetylase. PLoS ONE 2010, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, H.; Wang, X.; Zhang, R.; Wang, C.; Guo, Z. Sirtuin-3 (SIRT3) expression is associated with overall survival in esophageal cancer. Ann. Diagn. Pathol. 2013, 17, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Alhazzazi, T.Y.; Kamarajan, P.; Joo, N.; Huang, J.-Y.; Verdin, E.; D’Silva, N.J. Sirtuin-3 (SIRT3), a novel potential therapeutic target for oral cancer. Cancer 2011, 117, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Nihal, M.; Singh, C.K.; Zhong, W.; Liu, X.; Ahmad, N. Pro-proliferative function of mitochondrial sirtuin deacetylase SIRT3 in human melanoma. J. Investig. Dermatol. 2016, 136, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.J.; Milner, J. SIRT3 is pro-apoptotic and participates in distinct basal apoptotic pathways. Cell Cycle 2007, 6, 2669–2677. [Google Scholar] [CrossRef] [PubMed]
- Marfe, G.; Tafani, M.; Indelicato, M.; Sinibaldi-Salimei, P.; Reali, V.; Pucci, B.; Fini, M.; Russo, M.A. Kaempferol induces apoptosis in two different cell lines via Akt inactivation, Bax and SIRT3 activation, and mitochondrial dysfunction. J. Cell. Biochem. 2009, 106, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.-L.; Cheng, W.; Yin, X.-M.; Jiang, B. The expression of SIRT3 in primary hepatocellular carcinoma and the mechanism of its tumor suppressing effects. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 978–998. [Google Scholar] [PubMed]
- Zhang, B.; Qin, L.; Zhou, C.J.; Liu, Y.L.; Qian, H.X.; He, S.B. SIRT3 expression in hepatocellular carcinoma and its impact on proliferation and invasion of hepatoma cells. Asian Pac. J. Trop. Med. 2013, 6, 649–652. [Google Scholar] [CrossRef]
- Wang, J.-X.; Yi, Y.; Li, Y.-W.; Cai, X.-Y.; He, H.-W.; Ni, X.-C.; Zhou, J.; Cheng, Y.-F.; Jin, J.-J.; Fan, J.; et al. Down-regulation of sirtuin 3 is associated with poor prognosis in hepatocellular carcinoma after resection. BMC Cancer 2014, 14, 297. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Denu, R.A.; Krautkramer, K.A.; Grindle, K.M.; Yang, D.T.; Asimakopoulos, F.; Hematti, P.; Denu, J.M. Loss of SIRT3 provides growth advantage for B-cell malignancies. J. Biol. Chem. 2016, 291, 3268–3279. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.C.; Jing, L.M.; Wang, W.X.; Gao, Y.X. Down-regulation of SIRT3 promotes ovarian carcinoma metastasis. Biochem. Biophys. Res. Commun. 2016, 475, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Zhou, L.M. SIRT3 inhibits hepatocellular carcinoma cell growth through reducing Mdm2-mediated p53 degradation. Biochem. Biophys. Res. Commun. 2012, 423, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L.B.; Chandel, N.S. Mitochondrial reactive oxygen species and cancer. Cancer Metab. 2014, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sirtuin 3 (SIRT3) is involved in several processes, including metabolism and oxidative stress, which can be crucial for tumor development and promotion. Foxo3a: forkhead box O3; MnSOD: manganese superoxide dismutase; IDH2: isocitrate dehydrogenase; CH: carbohydrate; HKII: hexokinase II; AceCS2: acetyl-CoA synthase 2; PDC: pyruvate dehydrogenase complex; LCAD: long chain acyl-CoA dehydrogenase; GDH: glutamate dehydrogenase; OTC: ornithine transcarbamylase; LDH: lactate dehydrogenase; HIF: hypoxia-inducible factor.
Figure 1.
Sirtuin 3 (SIRT3) is involved in several processes, including metabolism and oxidative stress, which can be crucial for tumor development and promotion. Foxo3a: forkhead box O3; MnSOD: manganese superoxide dismutase; IDH2: isocitrate dehydrogenase; CH: carbohydrate; HKII: hexokinase II; AceCS2: acetyl-CoA synthase 2; PDC: pyruvate dehydrogenase complex; LCAD: long chain acyl-CoA dehydrogenase; GDH: glutamate dehydrogenase; OTC: ornithine transcarbamylase; LDH: lactate dehydrogenase; HIF: hypoxia-inducible factor.

Figure 2.
Main targets of SIRT3 in mitochondria. Green triangles represent protein activation, and red crosses represent target inactivation.
Figure 2.
Main targets of SIRT3 in mitochondria. Green triangles represent protein activation, and red crosses represent target inactivation.

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Torrens-Mas, M.; Oliver, J.; Roca, P.; Sastre-Serra, J. SIRT3: Oncogene and Tumor Suppressor in Cancer. Cancers 2017, 9, 90. https://doi.org/10.3390/cancers9070090
AMA Style
Torrens-Mas M, Oliver J, Roca P, Sastre-Serra J. SIRT3: Oncogene and Tumor Suppressor in Cancer. Cancers. 2017; 9(7):90. https://doi.org/10.3390/cancers9070090
Chicago/Turabian StyleTorrens-Mas, Margalida, Jordi Oliver, Pilar Roca, and Jorge Sastre-Serra. 2017. "SIRT3: Oncogene and Tumor Suppressor in Cancer" Cancers 9, no. 7: 90. https://doi.org/10.3390/cancers9070090
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.