
Critical Roles of the Sphingolipid Metabolic Pathway in Liver Regeneration, Hepatocellular Carcinoma Progression and Therapy

Abstract
:Simple Summary
Abstract
1. Introduction
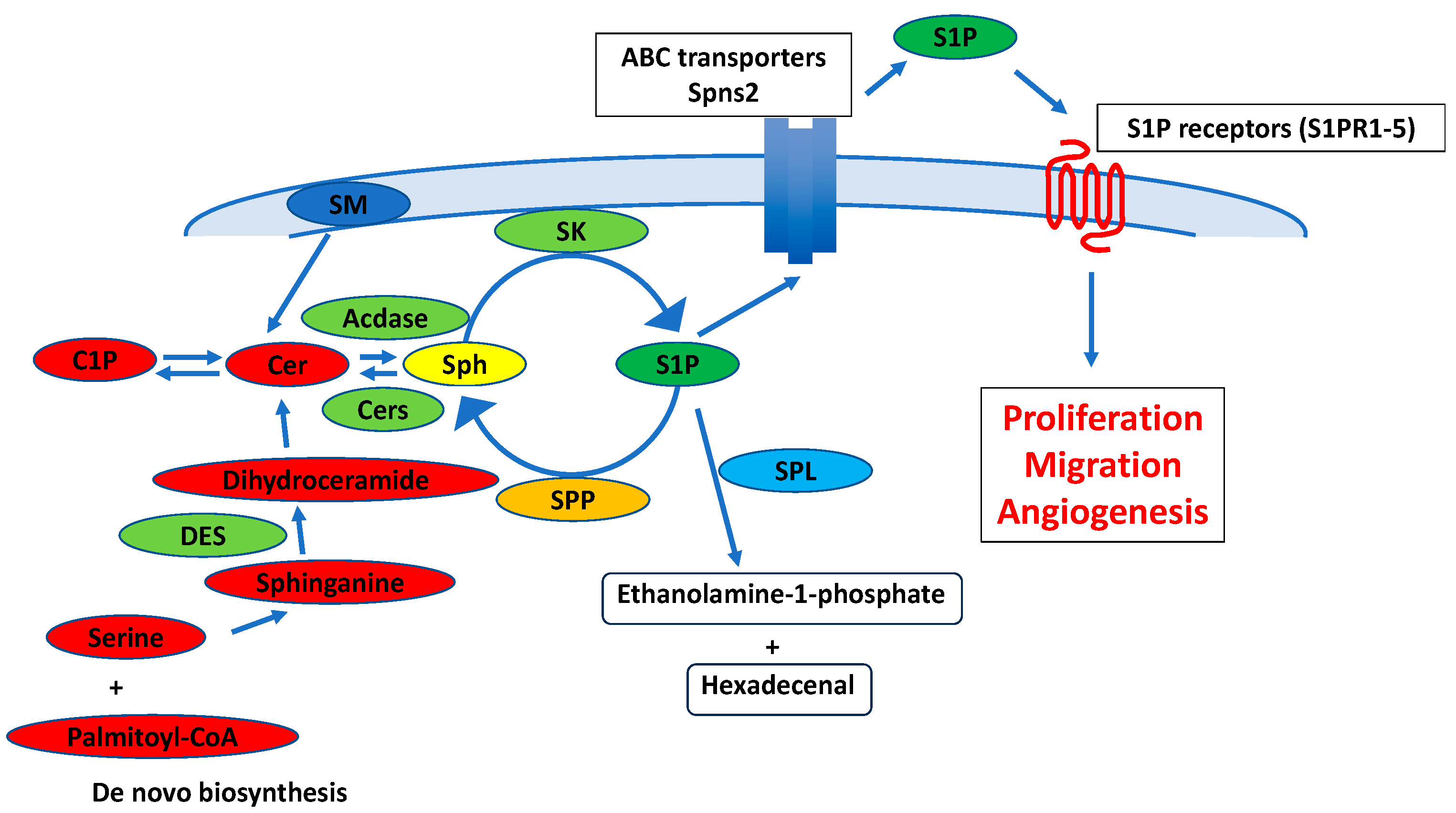
2. Metabolism and Physiological Functions in Sphingolipids
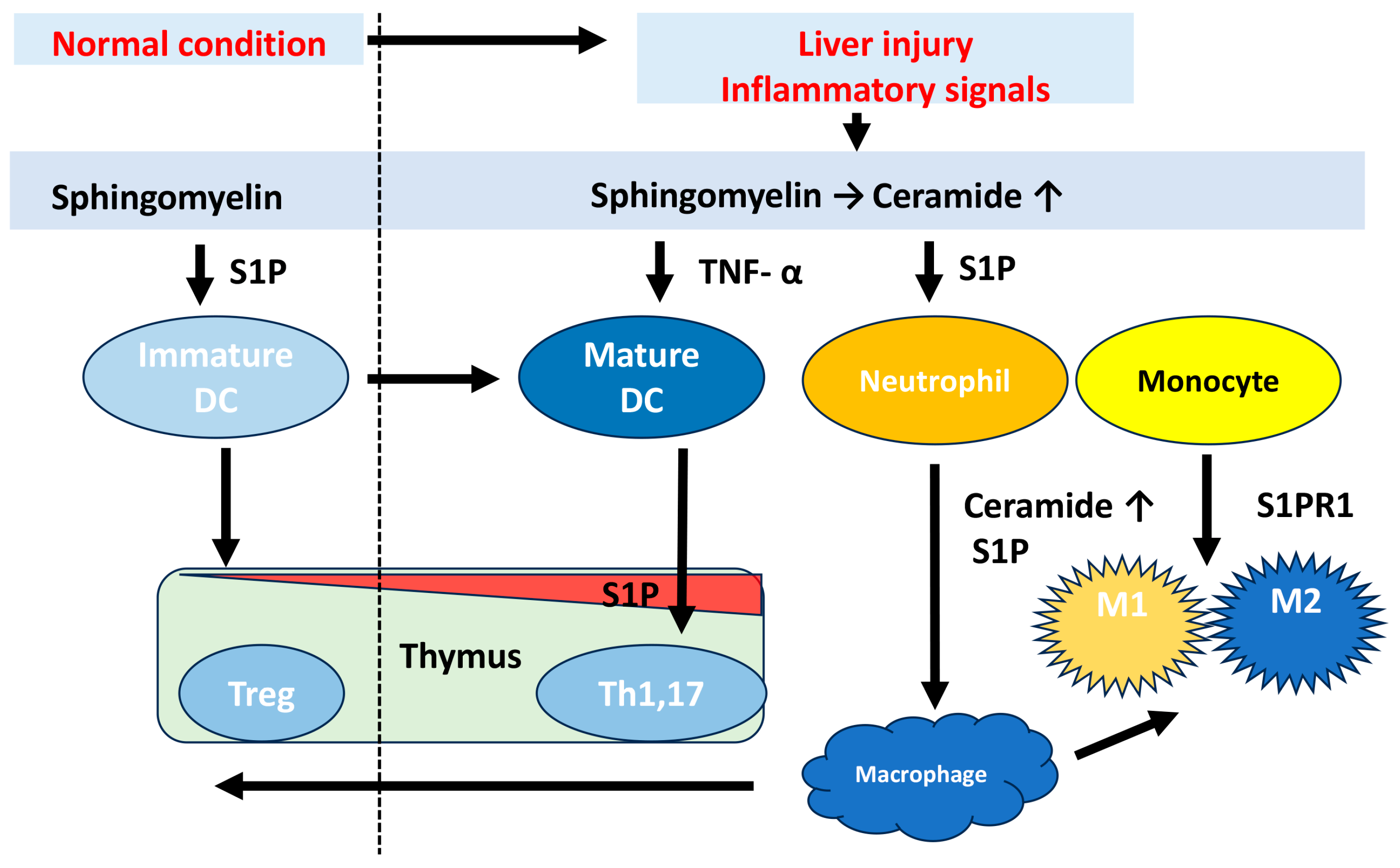
3. Sphingolipid Metabolic Pathway in Liver Regeneration
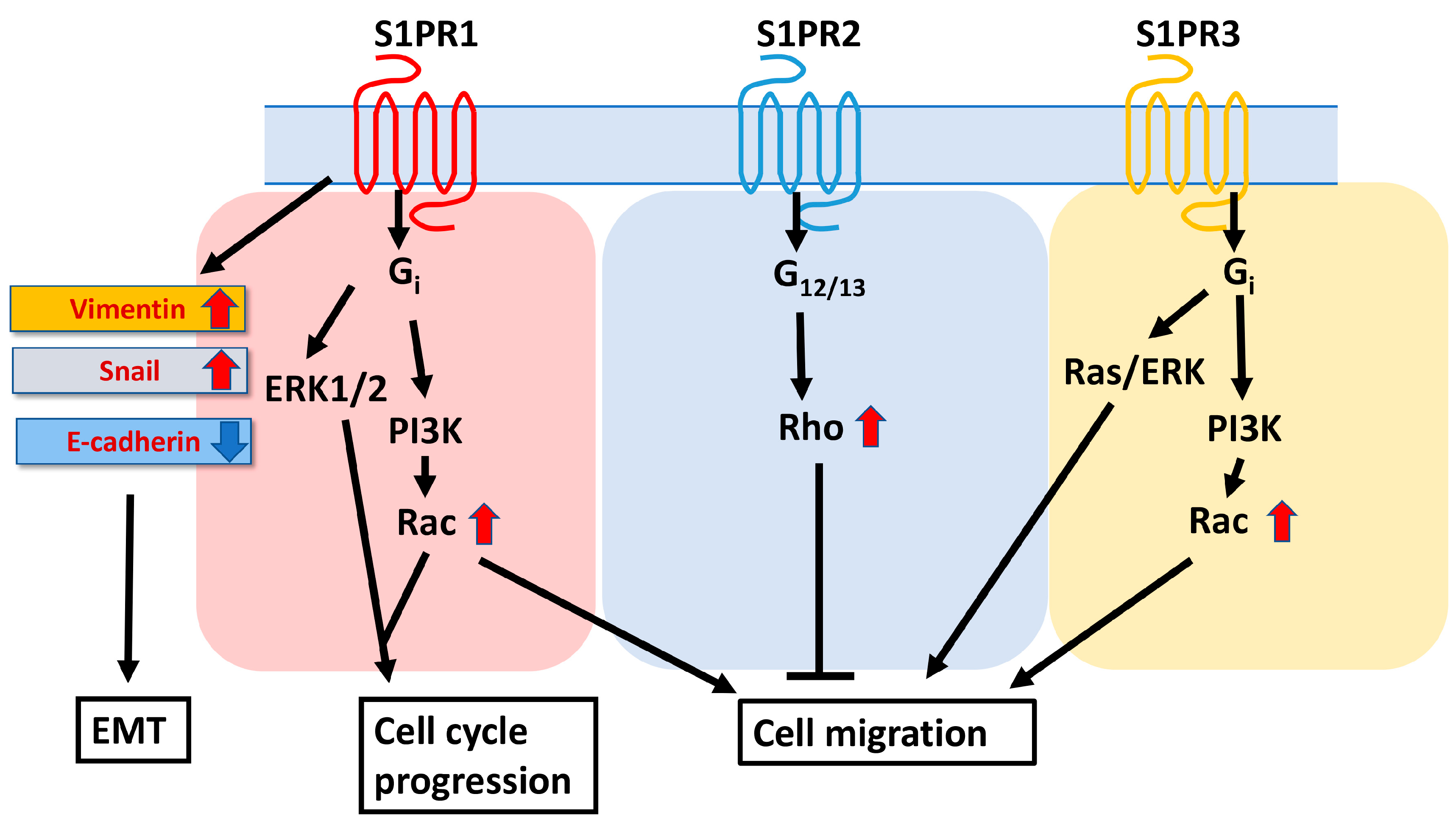
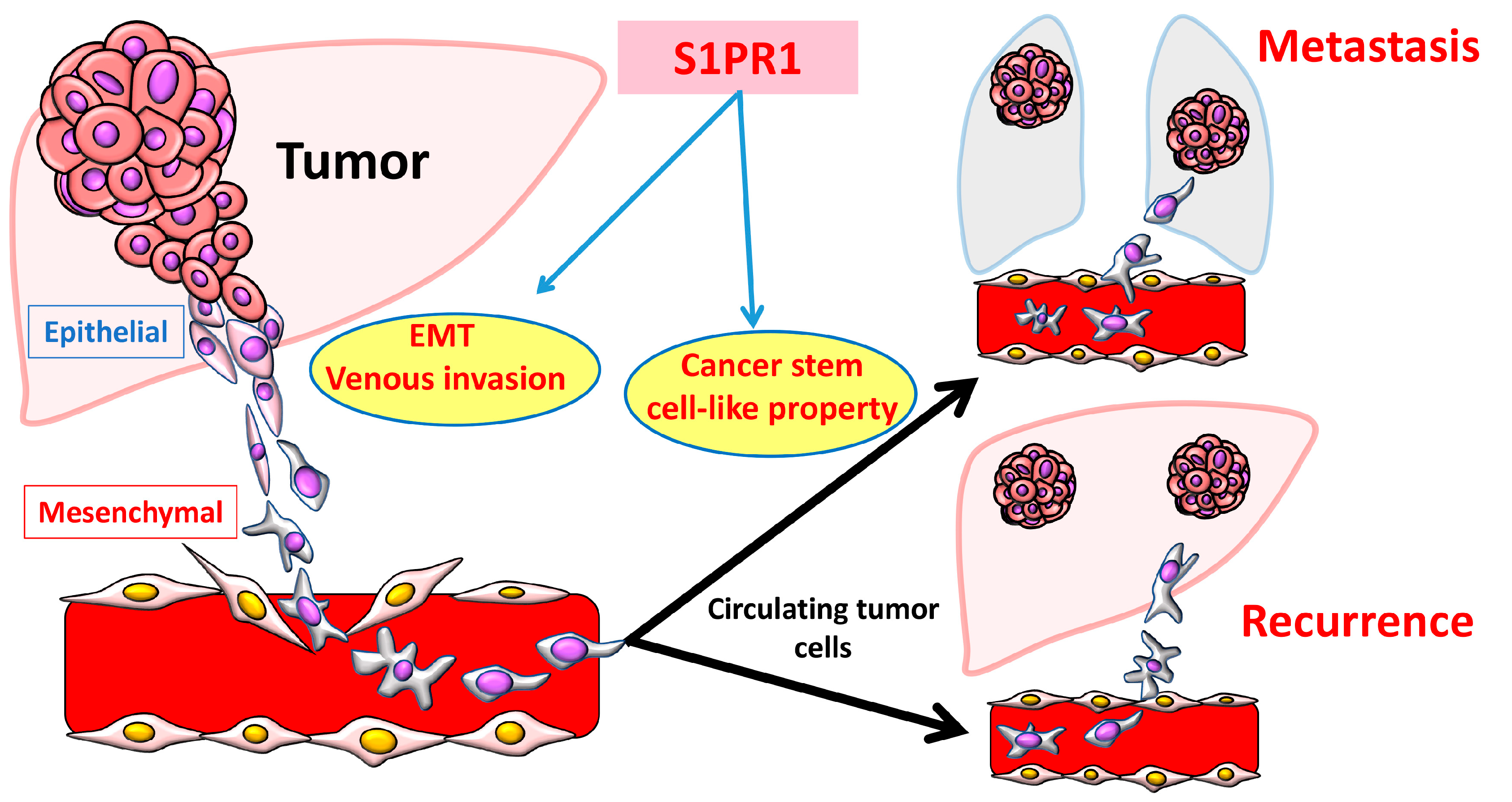
4. Sphingolipid Metabolic Pathway in Liver Fibrosis and HCC Progression
5. Sphingolipid Metabolic Pathway in HCC Therapy
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Quinville, B.M.; Deschenes, N.M.; Ryckman, A.E.; Walia, J.S. A Comprehensive Review: Sphingolipid Metabolism and Implications of Disruption in Sphingolipid Homeostasis. Int. J. Mol. Sci. 2021, 22, 5793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sun, C.; Zhao, Y.; Ye, B.; Yu, G. Signaling pathways of liver regeneration: Biological mechanisms and implications. iScience 2024, 27, 108683. [Google Scholar] [CrossRef]
- Nojima, H.; Kuboki, S.; Shinoda, K.; Shimizu, H.; Ohtsuka, M.; Kato, A.; Yoshitomi, H.; Furukawa, K.; Takayashiki, T.; Miyazaki, M. Activation of peroxisome proliferator-activated receptor-gamma inhibits tumor growth by negatively regulating nuclear factor-κB activation in patients with hepatocellular carcinoma. J. Hepatobiliary Pancreat. Sci. 2016, 23, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lin, H.; Wu, G.; Zhu, M.; Li, M. IL-6/STAT3 Is a Promising Therapeutic Target for Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 760971. [Google Scholar] [CrossRef] [PubMed]
- Shoda, L.K.; Battista, C.; Siler, S.Q.; Pisetsky, D.S.; Watkins, P.B.; Howell, B.A. Mechanistic Modelling of Drug-Induced Liver Injury: Investigating the Role of Innate Immune Responses. Gene Regul. Syst. Biol. 2017, 11, 1177625017696074. [Google Scholar] [CrossRef]
- Pralhada Rao, R.; Vaidyanathan, N.; Rengasamy, M.; Mammen Oommen, A.; Somaiya, N.; Jagannath, M.R. Sphingolipid metabolic pathway: An overview of major roles played in human diseases. J. Lipids 2013, 2013, 178910. [Google Scholar] [CrossRef] [PubMed]
- Nojima, H.; Freeman, C.M.; Gulbins, E.; Lentsch, A.B. Sphingolipids in liver injury, repair and regeneration. Biol. Chem. 2015, 396, 633–643. [Google Scholar] [CrossRef]
- Satyananda, V.; Oshi, M.; Tokumaru, Y.; Maiti, A.; Hait, N.; Matsuyama, R.; Endo, I.; Takabe, K. Sphingosine 1-phosphate (S1P) produced by sphingosine kinase 1 (SphK1) and exported via ABCC1 is related to hepatocellular carcinoma (HCC) progression. Am. J. Cancer Res. 2021, 11, 4394–4407. [Google Scholar]
- Shimizu, H.; Miyazaki, M.; Wakabayashi, Y.; Mitsuhashi, N.; Kato, A.; Ito, H.; Nakagawa, K.; Yoshidome, H.; Kataoka, M.; Nakajima, N. Vascular endothelial growth factor secreted by replicating hepatocytes induces sinusoidal endothelial cell proliferation during regeneration after partial hepatectomy in rats. J. Hepatol. 2001, 34, 683–689. [Google Scholar] [CrossRef]
- Kong, X.; Horiguchi, N.; Mori, M.; Gao, B. Cytokines and STATs in Liver Fibrosis. Front. Physiol. 2012, 3, 69. [Google Scholar] [CrossRef] [PubMed]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [CrossRef]
- Jin, J.; Hou, Q.; Mullen, T.D.; Zeidan, Y.H.; Bielawski, J.; Kraveka, J.M.; Bielawska, A.; Obeid, L.M.; Hannun, Y.A.; Hsu, Y.T. Ceramide generated by sphingomyelin hydrolysis and the salvage pathway is involved in hypoxia/reoxygenation-induced Bax redistribution to mitochondria in NT-2 cells. J. Biol. Chem. 2008, 283, 26509–26517. [Google Scholar] [CrossRef] [PubMed]
- Stith, J.L.; Velazquez, F.N.; Obeid, L.M. Advances in determining signaling mechanisms of ceramide and role in disease. J. Lipid Res. 2019, 60, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Cannavo, A.; Liccardo, D.; Komici, K.; Corbi, G.; de Lucia, C.; Femminella, G.D.; Elia, A.; Bencivenga, L.; Ferrara, N.; Koch, W.J.; et al. Sphingosine Kinases and Sphingosine 1-Phosphate Receptors: Signaling and Actions in the Cardiovascular System. Front. Pharmacol. 2017, 8, 556. [Google Scholar] [CrossRef] [PubMed]
- Pitson, S.M. Regulation of sphingosine kinase and sphingolipid signaling. Trends Biochem. Sci. 2011, 36, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sugiura, M.; Nava, V.E.; Edsall, L.C.; Kono, K.; Poulton, S.; Milstien, S.; Kohama, T.; Spiegel, S. Molecular cloning and functional characterization of a novel mammalian sphingosine kinase type 2 isoform. J. Biol. Chem. 2000, 275, 19513–19520. [Google Scholar] [CrossRef] [PubMed]
- Gulbins, E.; Kolesnick, R. Raft ceramide in molecular medicine. Oncogene 2003, 22, 7070–7077. [Google Scholar] [CrossRef]
- Chi, H. Sphingosine-1-phosphate and immune regulation: Trafficking and beyond. Trends Pharmacol. Sci. 2011, 32, 16–24. [Google Scholar] [CrossRef]
- Kroll, A.; Cho, H.E.; Kang, M.H. Antineoplastic Agents Targeting Sphingolipid Pathways. Front. Oncol. 2020, 10, 833. [Google Scholar] [CrossRef]
- Hajduch, E.; Lachkar, F.; Ferré, P.; Foufelle, F. Roles of Ceramides in Non-Alcoholic Fatty Liver Disease. J. Clin. Med. 2021, 10, 792. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Huai, Q.; Zhang, X.; Dai, H.; Li, X.; Wang, H. Insights into the roles and pathomechanisms of ceramide and sphigosine-1-phosphate in nonalcoholic fatty liver disease. Int. J. Biol. Sci. 2023, 19, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Z.; Wang, X.R.; Wang, J.; Xie, C.; Wang, X.X.; Pan, H.D.; Meng, W.Y.; Liang, T.L.; Li, J.X.; Yan, P.Y.; et al. The key role of sphingolipid metabolism in cancer: New therapeutic targets, diagnostic and prognostic values, and anti-tumor immunotherapy resistance. Front. Oncol. 2022, 12, 941643. [Google Scholar] [CrossRef] [PubMed]
- Garris, C.S.; Blaho, V.A.; Hla, T.; Han, M.H. Sphingosine-1-phosphate receptor 1 signalling in T cells: Trafficking and beyond. Immunology 2014, 142, 347–353. [Google Scholar] [CrossRef]
- Zhang, T.; de Waard, A.A.; Wuhrer, M.; Spaapen, R.M. The Role of Glycosphingolipids in Immune Cell Functions. Front. Immunol. 2019, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Carlsten, M.; Bryceson, Y.T. Rough operators: Sphingomyelinase inhibitors spike NK cells to kill cancer. Signal Transduct. Target. Ther. 2023, 8, 316. [Google Scholar] [CrossRef] [PubMed]
- Ishay, Y.; Nachman, D.; Khoury, T.; Ilan, Y. The role of the sphingolipid pathway in liver fibrosis: An emerging new potential target for novel therapies. Am. J. Physiol. Cell Physiol. 2020, 318, C1055–C1064. [Google Scholar] [CrossRef]
- Maceyka, M.; Harikumar, K.B.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate signaling and its role in disease. Trends Cell Biol. 2012, 22, 50–60. [Google Scholar] [CrossRef]
- Strub, G.M.; Maceyka, M.; Hait, N.C.; Milstien, S.; Spiegel, S. Extracellular and intracellular actions of sphingosine-1-phosphate. Adv. Exp. Med. Biol. 2010, 688, 141–155. [Google Scholar] [CrossRef]
- Wang, P.; Yuan, Y.; Lin, W.; Zhong, H.; Xu, K.; Qi, X. Roles of sphingosine-1-phosphate signaling in cancer. Cancer Cell Int. 2019, 19, 295. [Google Scholar] [CrossRef]
- Yan, J.; Chen, Y.; Wu, Q.; Shao, L.; Zhou, X. Expression of sphingosine-1-phosphate receptor 2 is correlated with migration and invasion of human colon cancer cells: A preliminary clinical study. Oncol. Lett. 2022, 24, 241. [Google Scholar] [CrossRef]
- Yokota, T.; Nojima, H.; Kuboki, S.; Yoshitomi, H.; Furukawa, K.; Takayashiki, T.; Takano, S.; Ohtsuka, M. Sphingosine-1-phosphate Receptor-1 Promotes Vascular Invasion and EMT in Hepatocellular Carcinoma. J. Surg. Res. 2021, 259, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Zabielski, P.; Baranowski, M.; Zendzian-Piotrowska, M.; Blachnio, A.; Gorski, J. Partial hepatectomy activates production of the pro-mitotic intermediates of the sphingomyelin signal transduction pathway in the rat liver. Prostaglandins Other Lipid Mediat. 2007, 83, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Sun, S.; Chen, G.; Xie, H.; Yu, S.; Lin, X.; Qian, J.; Mao, C.; Peng, H.; Chen, H.; et al. Ceramides and sphingosine-1-phosphate mediate the distinct effects of M1/M2-macrophage infusion on liver recovery after hepatectomy. Cell Death Dis. 2021, 12, 324. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Lazzarini, A.; Lazzarini, R.; Floridi, A.; Damaskopoulou, E.; Curcio, F.; Cataldi, S. Nuclear lipid microdomain as place of interaction between sphingomyelin and DNA during liver regeneration. Int. J. Mol. Sci. 2013, 14, 6529–6541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, X.; Ma, P.; Xiong, L.; Bai, W.; Zhang, G.; Xu, Y.; Song, W.; Yu, K. Establishment of a Rat Model of Liver Venous Deprivation: Simultaneous Portal and Hepatic Vein Ligation. J. Clin. Transl. Hepatol. 2023, 11, 393–404. [Google Scholar] [CrossRef]
- Nojima, H.; Freeman, C.M.; Schuster, R.M.; Japtok, L.; Kleuser, B.; Edwards, M.J.; Gulbins, E.; Lentsch, A.B. Hepatocyte exosomes mediate liver repair and regeneration via sphingosine-1-phosphate. J. Hepatol. 2016, 64, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Ji, A.L.; Wang, Z.X.; Qiang, G.H.; Qu, Z.; Wu, J.H.; Jiang, C.P. Exosome-Mimetic Nanovesicles from Hepatocytes promote hepatocyte proliferation in vitro and liver regeneration in vivo. Sci. Rep. 2018, 8, 2471. [Google Scholar] [CrossRef]
- Du, Y.; Li, D.; Han, C.; Wu, H.; Xu, L.; Zhang, M.; Zhang, J.; Chen, X. Exosomes from Human-Induced Pluripotent Stem Cell-Derived Mesenchymal Stromal Cells (hiPSC-MSCs) Protect Liver against Hepatic Ischemia/Reperfusion Injury via Activating Sphingosine Kinase and Sphingosine-1-Phosphate Signaling Pathway. Cell Physiol. Biochem. 2017, 43, 611–625. [Google Scholar] [CrossRef]
- Serriere-Lanneau, V.; Teixeira-Clerc, F.; Li, L.; Schippers, M.; de Wries, W.; Julien, B.; Tran-Van-Nhieu, J.; Manin, S.; Poelstra, K.; Chun, J.; et al. The sphingosine 1-phosphate receptor S1P2 triggers hepatic wound healing. FASEB J. 2007, 21, 2005–2013. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, L.; Chang, N.; Hou, L.; Zhou, X.; Yang, L.; Li, L. Neutrophils undergo switch of apoptosis to NETosis during murine fatty liver injury via S1P receptor 2 signaling. Cell Death Dis. 2020, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zeng, L.; Hu, M. Serum sphingosine-1 phosphate level is increased in patients with hepatitis B and displays a positive association with liver fibrosis. Am. J. Transl. Res. 2022, 14, 4964–4976. [Google Scholar] [PubMed]
- Li, C.; Zheng, S.; You, H.; Liu, X.; Lin, M.; Yang, L.; Li, L. Sphingosine 1-phosphate (S1P)/S1P receptors are involved in human liver fibrosis by action on hepatic myofibroblasts motility. J. Hepatol. 2011, 54, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yue, S.; Yang, L.; Liu, X.; Han, Z.; Zhang, Y.; Li, L. Sphingosine kinase/sphingosine 1-phosphate (S1P)/S1P receptor axis is involved in liver fibrosis-associated angiogenesis. J. Hepatol. 2013, 59, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yue, S.; Li, C.; Yang, L.; You, H.; Li, L. Essential roles of sphingosine 1-phosphate receptor types 1 and 3 in human hepatic stellate cells motility and activation. J. Cell. Physiol. 2011, 226, 2370–2377. [Google Scholar] [CrossRef]
- Hou, L.; Yang, L.; Chang, N.; Zhao, X.; Zhou, X.; Dong, C.; Liu, F.; Yang, L.; Li, L. Macrophage Sphingosine 1-Phosphate Receptor 2 Blockade Attenuates Liver Inflammation and Fibrogenesis Triggered by NLRP3 Inflammasome. Front. Immunol. 2020, 11, 1149. [Google Scholar] [CrossRef]
- Yang, L.; Han, Z.; Tian, L.; Mai, P.; Zhang, Y.; Wang, L.; Li, L. Sphingosine 1-Phosphate Receptor 2 and 3 Mediate Bone Marrow-Derived Monocyte/Macrophage Motility in Cholestatic Liver Injury in Mice. Sci. Rep. 2015, 5, 13423. [Google Scholar] [CrossRef]
- Ge, J.; Chang, N.; Zhao, Z.; Tian, L.; Duan, X.; Yang, L.; Li, L. Essential Roles of RNA-binding Protein HuR in Activation of Hepatic Stellate Cells Induced by Transforming Growth Factor-β1. Sci. Rep. 2016, 6, 22141. [Google Scholar] [CrossRef]
- Miura, K.; Nagahashi, M.; Prasoon, P.; Hirose, Y.; Kobayashi, T.; Sakata, J.; Abe, M.; Sakimura, K.; Matsuda, Y.; Butash, A.L.; et al. Dysregulation of sphingolipid metabolic enzymes leads to high levels of sphingosine-1-phosphate and ceramide in human hepatocellular carcinoma. Hepatol. Res. 2021, 51, 614–626. [Google Scholar] [CrossRef]
- Ji, Y.; Chen, H.; Gow, W.; Ma, L.; Jin, Y.; Hui, B.; Yang, Z.; Wang, Z. Potential biomarkers Ang II/AT1R and S1P/S1PR1 predict the prognosis of hepatocellular carcinoma. Oncol. Lett. 2020, 20, 208. [Google Scholar] [CrossRef]
- Wang, F.; Wu, Z. Sphingosine kinase 1 overexpression is associated with poor prognosis and oxaliplatin resistance in hepatocellular carcinoma. Exp. Ther. Med. 2018, 15, 5371–5376. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, D.; de Verdier, P.J.; Ryk, C.; Lunqe, O.; Stål, P.; Flygare, J. FTY720 (Fingolimod) sensitizes hepatocellular carcinoma cells to sorafenib-mediated cytotoxicity. Pharmacol. Res. Perspect. 2015, 3, e00171. [Google Scholar] [CrossRef]
- Liu, X.T.; Chung, L.H.; Liu, D.; Chen, J.; Huang, Y.; Teo, J.D.; Han, X.D.; Zhao, Y.; Guan, F.H.X.; Tran, C.; et al. Ablation of sphingosine kinase 2 suppresses fatty liver-associated hepatocellular carcinoma via downregulation of ceramide transfer protein. Oncogenesis 2022, 11, 67. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.L.; Kang, Y.K.; Chen, Z.; Tsao, C.J.; Qin, S.; Kim, J.S.; Luo, R.; Feng, J.; Ye, S.; Yang, T.S.; et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: A phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009, 10, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Momchilova, A.; Nikolaev, G.; Pankov, S.; Vassileva, E.; Krastev, N.; Robev, B.; Krastev, D.; Pinkas, A.; Pankov, R. Effect of Quercetin and Fingolimod, Alone or in Combination, on the Sphingolipid Metabolism in HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 3916. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Zhang, S.; Ma, D.; Yan, D.; Zhang, G.; Cao, Y.; Wang, Z.; Wu, J.; Jiang, C. Targeting SphK2 Reverses Acquired Resistance of Regorafenib in Hepatocellular Carcinoma. Front. Oncol. 2020, 10, 694. [Google Scholar] [CrossRef]
- Uranbileg, B.; Kurano, M.; Kano, K.; Sakai, E.; Arita, J.; Hasegawa, K.; Nishikawa, T.; Ishihara, S.; Yamashita, H.; Seto, Y.; et al. Sphingosine 1-phosphate lyase facilitates cancer progression through converting sphingolipids to glycerophospholipids. Clin. Transl. Med. 2022, 12, e1056. [Google Scholar] [CrossRef]
- Matsushima-Nishiwaki, R.; Yamada, N.; Fukuchi, K.; Kozawa, O. Sphingosine 1-phosphate (S1P) reduces hepatocyte growth factor-induced migration of hepatocellular carcinoma cells via S1P receptor 2. PLoS ONE 2018, 13, e0209050. [Google Scholar] [CrossRef]
- Yin, Y.; Xu, M.; Gao, J.; Li, M. Alkaline ceramidase 3 promotes growth of hepatocellular carcinoma cells via regulating S1P/S1PR2/PI3K/AKT signaling. Pathol. Res. Pract. 2018, 214, 1381–1387. [Google Scholar] [CrossRef]
- Hoffmann, K.; Nagel, A.J.; Tanabe, K.; Fuchs, J.; Dehlke, K.; Ghamarnejad, O.; Lemekhova, A.; Mehrabi, A. Markers of liver regeneration-the role of growth factors and cytokines: A systematic review. BMC Surg. 2020, 20, 31. [Google Scholar] [CrossRef]
- Pedone, E.; Olteanu, V.A.; Marucci, L.; Muñoz-Martin, M.I.; Youssef, S.A.; de Bruin, A.; Cosma, M.P. Modeling Dynamics and Function of Bone Marrow Cells in Mouse Liver Regeneration. Cell Rep. 2017, 18, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Verma, B.K.; Subramaniam, P.; Vadigepalli, R. Model-based virtual patient analysis of human liver regeneration predicts critical perioperative factors controlling the dynamic mode of response to resection. BMC Syst. Biol. 2019, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Sparrelid, E.; Olthof, P.B.; Dasari, B.V.M.; Erdmann, J.I.; Santol, J.; Starlinger, P.; Gilg, S. Current evidence on posthepatectomy liver failure: Comprehensive review. BJS Open 2022, 6, zrac142. [Google Scholar] [CrossRef] [PubMed]
- Nowatari, T.; Murata, S.; Nakayama, K.; Sano, N.; Maruyama, T.; Nozaki, R.; Ikeda, N.; Fukunaga, K.; Ohkohchi, N. Sphingosine 1-phosphate has anti-apoptotic effect on liver sinusoidal endothelial cells and proliferative effect on hepatocytes in a paracrine manner in human. Hepatol. Res. 2015, 45, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Alessenko, A.; Chatterjee, S. Neutral sphingomyelinase: Localization in rat liver nuclei and involvement in regeneration/proliferation. Mol. Cell. Biochem. 1995, 143, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Zabielski, P.; Blachnio-Zabielska, A.; Baranowski, M.; Zendzian-Piotrowska, M.; Gorski, J. Activation of PPARα by bezafibrate negatively affects de novo synthesis of sphingolipids in regenerating rat liver. Prostaglandins Other Lipid Mediat. 2010, 93, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Nojima, H.; Konishi, T.; Freeman, C.M.; Schuster, R.M.; Japtok, L.; Kleuser, B.; Edwards, M.J.; Gulbins, E.; Lentsch, A.B. Chemokine Receptors, CXCR1 and CXCR2, Differentially Regulate Exosome Release in Hepatocytes. PLoS ONE 2016, 11, e0161443. [Google Scholar] [CrossRef] [PubMed]
- Cartier, A.; Hla, T. Sphingosine 1-phosphate: Lipid signaling in pathology and therapy. Science 2019, 366, eaar5551. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, Z.; Dong, W.; Yang, Z.; Wang, J.; Xu, H.; Sun, T.; Huang, Z.; Jin, J. S1PR1 induces metabolic reprogramming of ceramide in vascular endothelial cells, affecting hepatocellular carcinoma angiogenesis and progression. Cell Death Dis. 2022, 13, 768. [Google Scholar] [CrossRef]
- Ikeda, H.; Watanabe, N.; Ishii, I.; Shimosawa, T.; Kume, Y.; Tomiya, T.; Inoue, Y.; Nishikawa, T.; Ohtomo, N.; Tanoue, Y.; et al. Sphingosine 1-phosphate regulates regeneration and fibrosis after liver injury via sphingosine 1-phosphate receptor 2. J. Lipid Res. 2009, 50, 556–564. [Google Scholar] [CrossRef]
- Gonzalez-Carmona, M.A.; Sandhoff, R.; Tacke, F.; Vogt, A.; Weber, S.; Canbay, A.E.; Rogler, G.; Sauerbruch, T.; Lammert, F.; Yildiz, Y. Beta-glucosidase 2 knockout mice with increased glucosylceramide show impaired liver regeneration. Liver Int. 2012, 32, 1354–1362. [Google Scholar] [CrossRef]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]
- Dutta-Moscato, J.; Solovyev, A.; Mi, Q.; Nishikawa, T.; Soto-Gutierrez, A.; Fox, I.J.; Vodovotz, Y. A Multiscale Agent-Based in silico Model of Liver Fibrosis Progression. Front. Bioeng. Biotechnol. 2014, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Kawai, H.; Osawa, Y.; Matsuda, M.; Tsunoda, T.; Yanagida, K.; Hishikawa, D.; Okawara, M.; Sakamoto, Y.; Shimagaki, T.; Tsutsui, Y.; et al. Sphingosine-1-phosphate promotes tumor development and liver fibrosis in mouse model of congestive hepatopathy. Hepatology 2022, 76, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Shin, E.J.; Yang, J.H.; Ki, S.H. Integrative roles of sphingosine kinase in liver pathophysiology. Toxicol. Res. 2023, 39, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Quillin, R.C., 3rd; Wilson, G.C.; Nojima, H.; Freeman, C.M.; Wang, J.; Schuster, R.M.; Blanchard, J.A.; Edwards, M.J.; Gandhi, C.R.; Gulbins, E.; et al. Inhibition of acidic sphingomyelinase reduces established hepatic fibrosis in mice. Hepatol. Res. 2015, 45, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Kleuser, B. Divergent Role of Sphingosine 1-Phosphate in Liver Health and Disease. Int. J. Mol. Sci. 2018, 19, 722. [Google Scholar] [CrossRef] [PubMed]
- Cuzzocrea, S.; Di Paola, R.; Genovese, T.; Mazzon, E.; Esposito, E.; Crisafulli, C.; Bramanti, P.; Salvemini, D. Anti-inflammatory and anti-apoptotic effects of fumonisin B1, an inhibitor of ceramide synthase, in a rodent model of splanchnic ischemia and reperfusion injury. J. Pharmacol. Exp. Ther. 2008, 327, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Ouro, A.; Ala-Ibanibo, L.; Presa, N.; Delgado, T.C.; Martínez-Chantar, M.L. Sphingolipids in Non-Alcoholic Fatty Liver Disease and Hepatocellular Carcinoma: Ceramide Turnover. Int. J. Mol. Sci. 2019, 21, 40. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.C.; Patel, S.H.; Wang, J.; Xu, K.; Turner, K.M.; Becker, K.A.; Carpinteiro, A.; Szabo, I.; Ahmad, S.A.; Gulbins, E. Acid sphingomyelinase expression is associated with survival in resectable pancreatic ductal adenocarcinoma. J. Mol. Med. 2023, 101, 891–903. [Google Scholar] [CrossRef]
- Zeng, Y.; Liu, X.; Yan, Z.; Xie, L. Sphingosine 1-phosphate regulates proliferation, cell cycle and apoptosis of hepatocellular carcinoma cells via syndecan-1. Prog. Biophys. Mol. Biol. 2019, 148, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Moro, K.; Nagahashi, M.; Gabriel, E.; Takabe, K.; Wakai, T. Clinical application of ceramide in cancer treatment. Breast Cancer 2019, 26, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, G.; Schoell, N.; Ferreirós, N.; Bon, D.; Herrmann, E.; Farnik, H.; Köberle, V.; Piiper, A.; Zeuzem, S.; Kronenberger, B.; et al. Serum sphingolipidomic analyses reveal an upregulation of C16-ceramide and sphingosine-1-phosphate in hepatocellular carcinoma. Oncotarget 2016, 7, 18095–18105. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kabir, I.; Jiang, H.; Zhou, H.; Libien, J.; Zeng, J.; Stanek, A.; Ou, P.; Li, K.R.; Zhang, S.; et al. Liver serine palmitoyltransferase activity deficiency in early life impairs adherens junctions and promotes tumorigenesis. Hepatology 2016, 64, 2089–2102. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.P.; Xiao, Z.L.; Yang, Z.; Li, J.; Feng, G.X.; Chen, F.Q.; Li, Y.H.; Feng, J.Y.; Gao, Y.E.; Ye, L.H.; et al. Hepatitis B virus X protein promotes human hepatoma cell growth via upregulation of transcription factor AP2α and sphingosine kinase 1. Acta Pharmacol. Sin. 2015, 36, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Rohrbach, T.; Milstien, S.; Spiegel, S. Role of Sphingosine Kinase 1 and Sphingosine-1-Phosphate Axis in Hepatocellular Carcinoma. Handb. Exp. Pharmacol. 2020, 259, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qi, Y.; Zhao, Y.; Kaczorowski, D.; Couttas, T.A.; Coleman, P.R.; Don, A.S.; Bertolino, P.; Gamble, J.R.; Vadas, M.A.; et al. Deletion of sphingosine kinase 1 inhibits liver tumorigenesis in diethylnitrosamine-treated mice. Oncotarget 2018, 9, 15635–15649. [Google Scholar] [CrossRef]
- Lu, Z.; Xiao, Z.; Liu, F.; Cui, M.; Li, W.; Yang, Z.; Li, J.; Ye, L.; Zhang, X. Long non-coding RNA HULC promotes tumor angiogenesis in liver cancer by up-regulating sphingosine kinase 1 (SPHK1). Oncotarget 2016, 7, 241–254. [Google Scholar] [CrossRef]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Kudo, M.; Finn, R.S.; Qin, S.; Han, K.H.; Ikeda, K.; Piscaglia, F.; Baron, A.; Park, J.W.; Han, G.; Jassem, J.; et al. Lenvatinib versus sorafenib in first-line treatment of patients with unresectable hepatocellular carcinoma: A randomised phase 3 non-inferiority trial. Lancet 2018, 391, 1163–1173. [Google Scholar] [CrossRef]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef] [PubMed]
- Maestri, M.; Pallozzi, M.; Santopaolo, F.; Cerrito, L.; Pompili, M.; Gasbarrini, A.; Ponziani, F.R. Durvalumab: An investigational agent for unresectable hepatocellular carcinoma. Expert Opin. Investig. Drugs 2022, 31, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Bruix, J.; Qin, S.; Merle, P.; Granito, A.; Huang, Y.H.; Bodoky, G.; Pracht, M.; Yokosuka, O.; Rosmorduc, O.; Breder, V.; et al. Regorafenib for patients with hepatocellular carcinoma who progressed on sorafenib treatment (RESORCE): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 389, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.X.; Finn, R.S.; Edeline, J.; Cattan, S.; Ogasawara, S.; Palmer, D.; Verslype, C.; Zagonel, V.; Fartoux, L.; Vogel, A.; et al. Pembrolizumab in patients with advanced hepatocellular carcinoma previously treated with sorafenib (KEYNOTE-224): A non-randomised, open-label phase 2 trial. Lancet Oncol. 2018, 19, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.S.; Ryoo, B.Y.; Merle, P.; Kudo, M.; Bouattour, M.; Lim, H.Y.; Breder, V.; Edeline, J.; Chao, Y.; Ogasawara, S.; et al. Pembrolizumab As Second-Line Therapy in Patients With Advanced Hepatocellular Carcinoma in KEYNOTE-240: A Randomized, Double-Blind, Phase III Trial. J. Clin. Oncol. 2020, 38, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Abou-Alfa, G.K.; Meyer, T.; Cheng, A.L.; El-Khoueiry, A.B.; Rimassa, L.; Ryoo, B.Y.; Cicin, I.; Merle, P.; Chen, Y.; Park, J.W.; et al. Cabozantinib in Patients with Advanced and Progressing Hepatocellular Carcinoma. N. Engl. J. Med. 2018, 379, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.X.; Kang, Y.K.; Yen, C.J.; Finn, R.S.; Galle, P.R.; Llovet, J.M.; Assenat, E.; Brandi, G.; Pracht, M.; Lim, H.Y.; et al. Ramucirumab after sorafenib in patients with advanced hepatocellular carcinoma and increased α-fetoprotein concentrations (REACH-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 2019, 20, 282–296. [Google Scholar] [CrossRef]
- Yau, T.; Kang, Y.K.; Kim, T.Y.; El-Khoueiry, A.B.; Santoro, A.; Sangro, B.; Melero, I.; Kudo, M.; Hou, M.M.; Matilla, A.; et al. Efficacy and Safety of Nivolumab Plus Ipilimumab in Patients With Advanced Hepatocellular Carcinoma Previously Treated With Sorafenib: The CheckMate 040 Randomized Clinical Trial. JAMA Oncol. 2020, 6, e204564. [Google Scholar] [CrossRef]
- Qin, S.; Chen, Z.; Fang, W.; Ren, Z.; Xu, R.; Ryoo, B.Y.; Meng, Z.; Bai, Y.; Chen, X.; Liu, X.; et al. Pembrolizumab Versus Placebo as Second-Line Therapy in Patients From Asia With Advanced Hepatocellular Carcinoma: A Randomized, Double-Blind, Phase III Trial. J. Clin. Oncol. 2023, 41, 1434–1443. [Google Scholar] [CrossRef]
- Correction: Virtual clinical trials of anti-PD-1 and anti-CTLA-4 immunotherapy in advanced hepatocellular carcinoma using a quantitative systems pharmacology model. J. Immunother. Cancer 2023, 11, e005414corr1. [CrossRef]
- Janneh, A.H.; Ogretmen, B. Targeting Sphingolipid Metabolism as a Therapeutic Strategy in Cancer Treatment. Cancers 2022, 14, 2183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Huang, D.; Shao, D.; Liu, H.; Zhou, Q.; Gui, S.; Wei, W.; Wang, Y. Fenretinide inhibits the proliferation and migration of human liver cancer HepG2 cells by downregulating the activation of myosin light chain kinase through the p38-MAPK signaling pathway. Oncol. Rep. 2018, 40, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Adiseshaiah, P.P.; Clogston, J.D.; McLeland, C.B.; Rodriguez, J.; Potter, T.M.; Neun, B.W.; Skoczen, S.L.; Shanmugavelandy, S.S.; Kester, M.; Stern, S.T.; et al. Synergistic combination therapy with nanoliposomal C6-ceramide and vinblastine is associated with autophagy dysfunction in hepatocarcinoma and colorectal cancer models. Cancer Lett 2013, 337, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Grbčić, P.; Tomljanović, I.; Klobučar, M.; Kraljević Pavelić, S.; Lučin, K.; Sedić, M. Dual sphingosine kinase inhibitor SKI-II enhances sensitivity to 5-fluorouracil in hepatocellular carcinoma cells via suppression of osteopontin and FAK/IGF-1R signalling. Biochem. Biophys. Res. Commun. 2017, 487, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Chen, Y.Z.; Peng, Y.; Yi, N.; Gu, X.S.; Jin, Y.; Bai, X.M. Ceramide production mediates cinobufotalin-induced growth inhibition and apoptosis in cultured hepatocellular carcinoma cells. Tumour Biol. 2015, 36, 5763–5771. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhang, X.; He, S.; Miao, Y.; Wu, N.; Li, J.; Gan, Y. Sphk2 RNAi nanoparticles suppress tumor growth via downregulating cancer cell derived exosomal microRNA. J. Control. Release 2018, 286, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; He, H.; Zhang, H.; Yu, D.; Zhao, W.; Chen, Y.; Shao, R. The blockage of Ras/ERK pathway augments the sensitivity of SphK1 inhibitor SKI II in human hepatoma HepG2 cells. Biochem. Biophys. Res. Commun. 2013, 434, 35–41. [Google Scholar] [CrossRef]
- Jakobi, K.; Beyer, S.; Koch, A.; Thomas, D.; Schwalm, S.; Zeuzem, S.; Pfeilschifter, J.; Grammatikos, G. Sorafenib Treatment and Modulation of the Sphingolipid Pathway Affect Proliferation and Viability of Hepatocellular Carcinoma In Vitro. Int. J. Mol. Sci. 2020, 21, 2409. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Sphingolipids | Pharmalogical Approach | References | |
|---|---|---|---|---|
| Hepatectomy | Ceramide ↑, ASM ↑ | Liver regenerarion ↑ | [33] | |
| Hepatectomy | Ceramide ↑, S1P ↑ | Liver regenerarion ↑ | [34] | |
| Hepatectomy | sphingomyelinase ↑ | Liver regenerarion ↑ | [35] | |
| Hepatectomy (ALPPS) | S1P (serum and liver) ↑ | [36] | ||
| Hepatectomy | SK2 ↑, S1P ↑ | Hepatocyte-derived exosomes | Liver regenerarion ↑ | [37] |
| Hepatectomy | SK2 ↑ | Nanovesicles from hepatocytes | Liver regenerarion ↑ | [38] |
| Ischemia reperfusion | SK1 ↑, S1P ↑ | Human-induced pluripotent stem cell-derived mesenchymal stromal cells | Liver regenerarion ↑ | [39] |
| Acute liver injury | S1PR2 ↑ | Liver regenerarion ↑ | [40] | |
| Liver fibrosis | S1PR2 inhibition | Liver regenerarion ↑ fibrosis ↓ | [41] | |
| Liver fibrosis | S1P ↑ | fibrosis ↑ | [42] | |
| Liver fibrosis | S1PR1 ↑, S1PR3 ↑, | fibrosis ↑ | [43,44] | |
| Liver fibrosis | S1PR1 ↑, S1PR3 ↑, S1P ↑ | fibrosis ↑ | [45] | |
| Liver fibrosis | S1PR2 inhibition | fibrosis ↓ | [46] | |
| Liver fibrosis | S1PR2 ↑, S1PR3 ↑, | fibrosis ↑ | [47] | |
| Liver fibrosis | SK1 ↑ | fibrosis ↑ | [48] | |
| HCC | Ceramide ↑, S1P ↑, SK1 ↑ | [49] | ||
| HCC | S1PR1↑ | S1PR1 inhibition | HCC Progression ↑ | [32] |
| HCC | S1PR1 ↑ | HCC Progression ↑ | [50] | |
| HCC | S1PR1 ↑, Ceramide ↑ | HCC Progression ↑ | [51] | |
| HCC | FTY720 | Sorafenib-mediated cytotoxicity | [52] | |
| HCC | SK2 ↑ | HCC Progression ↓ | [53] | |
| HCC | SK1 ↓ | Cinobufotalin | HCC cells ↓ | [54] |
| HCC | SK1 ↓ | Fingolimod | HCC cells ↓ | [55] |
| HCC | SK2 ↓ | ABC294640 | HCC cells ↓ | [56] |
| HCC | S1P lyase ↑ | HCC Progression ↑ | [57] | |
| HCC | S1PR2 ↑ | CYM5520 | HCC Progression ↓ | [58] |
| HCC | S1PR2 ↑ | Alkaline ceramidase 3 inhibition | HCC Progression ↓ | [59] |
| ↑ up-regulation | ↑ up-regulation | |||
| ↓ down-regulation | ↓ down-regulation |
| References | Trials | Agent | Targets | Indication | Overall Survival (Months) |
|---|---|---|---|---|---|
| [70] | SHARP | Sorafenib | VEGFR, c-KIT, PDGFR, RET and Ras/Raf/MEK/ERK | First-line | 10.7 |
| [40] | Asia–Pacific | Lenvatinib | VEGFR, PDGFR, FGFR, KIT and RET | First-line | 6.5 |
| [71] | REFLECT | Bevacizumab Atezolizumab | VEGF and PD-L1 | First-line | 13.6 |
| [72] | IMbrave150 | Durvalumab Tremelimumab | PDL-1 and CTLA-4 | First-line | 16.4 |
| [73] | HIMALAYA | Atezolizumab Cabozantinib | PD-L1 and VEGF | First-line | 15.4 |
| [74] | RESORCE | Regorafenib | VEGFR, FGFR, PDGFR, B-RAF, RET and KIT | Second | 10.6 |
| [41] | KEYNOTE 224 | Pembrolizumab | PD-1 | Second | 12.9 |
| [75] | KEYNOTE 240 | Pembrolizumab | PD-1 | Second | 13.9 |
| [76] | CELESTIAL | Cabozantinib | VEGFR, AXL, c-MET, KIT and RET | Second | 10.2 |
| [77] | REACH-2 | Ramucirumab | VEGFR2 | Second | 8.5 |
| [78] | CHECKMATE 040 | Nivolumab and Ipilimumab | PD-1 and CTLA-4 | Second | arm A: 22.8 B: 12.5 C: 12.7 |
| [42] | KEYNOTE 394 | Pembrolizumab | PD-1 | Second | 14.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nojima, H.; Shimizu, H.; Murakami, T.; Shuto, K.; Koda, K. Critical Roles of the Sphingolipid Metabolic Pathway in Liver Regeneration, Hepatocellular Carcinoma Progression and Therapy. Cancers 2024, 16, 850. https://doi.org/10.3390/cancers16050850
Nojima H, Shimizu H, Murakami T, Shuto K, Koda K. Critical Roles of the Sphingolipid Metabolic Pathway in Liver Regeneration, Hepatocellular Carcinoma Progression and Therapy. Cancers. 2024; 16(5):850. https://doi.org/10.3390/cancers16050850
Chicago/Turabian StyleNojima, Hiroyuki, Hiroaki Shimizu, Takashi Murakami, Kiyohiko Shuto, and Keiji Koda. 2024. "Critical Roles of the Sphingolipid Metabolic Pathway in Liver Regeneration, Hepatocellular Carcinoma Progression and Therapy" Cancers 16, no. 5: 850. https://doi.org/10.3390/cancers16050850