

The Membrane Protein Sortilin Is a Potential Biomarker and Target for Glioblastoma
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. SORT1 (Sortilin) mRNA Data Mining
2.2. Patient Samples
2.3. Immunohistochemical Detection and Quantification of Sortilin Expression
2.4. Sortilin Quantification in Patient Plasma Samples
2.5. Cell lines and Culture Conditions
2.6. Western Blotting
2.7. Measurement of Cell Growth and Invasions
2.8. Statistics
3. Results
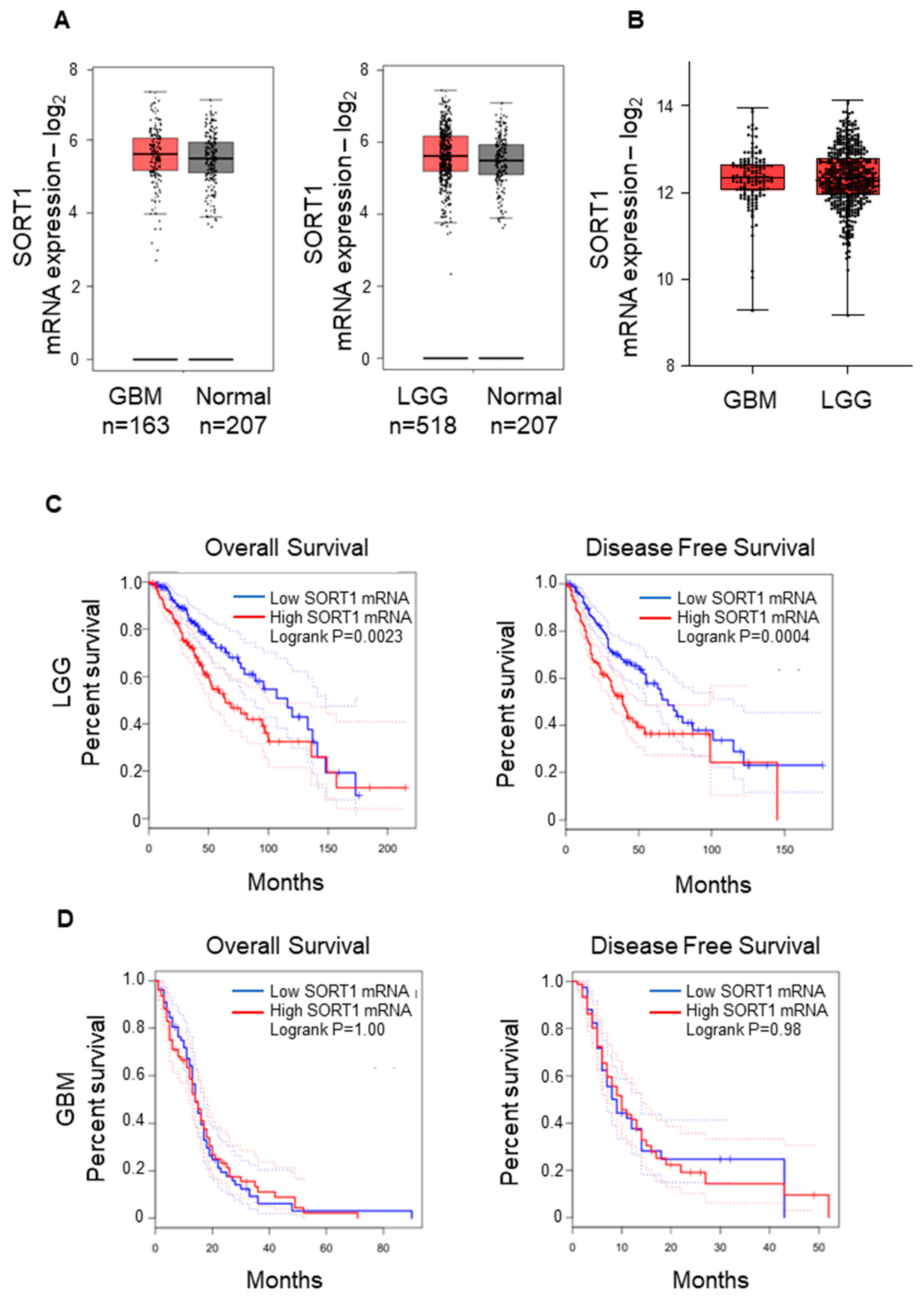
3.1. SORT1 (Sortilin) mRNA Expression Is Not Increased in GBM Tissues
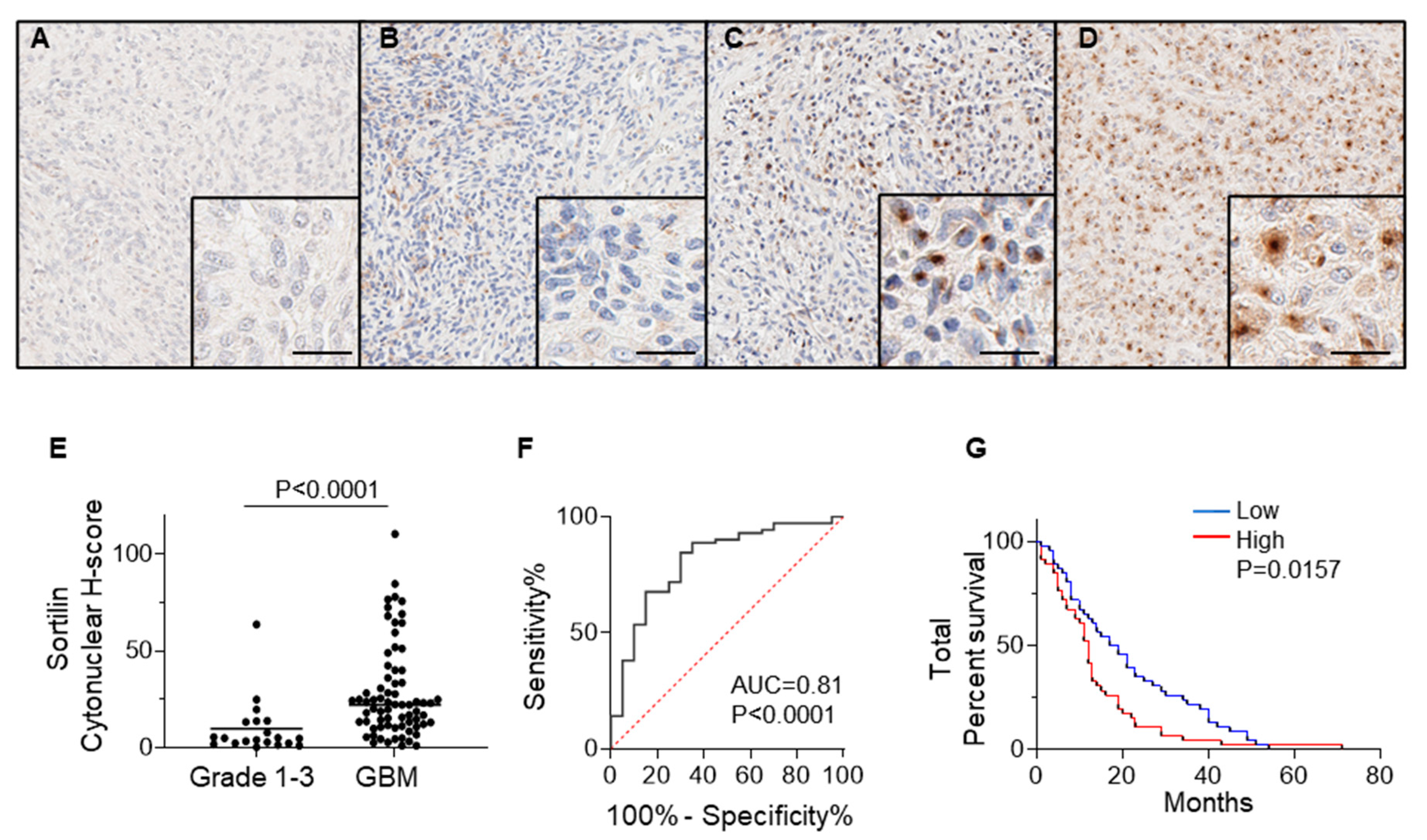
3.2. Sortilin Protein Expression Is Increased in GBM Tissues Compared to Grade 1–3 Glioma
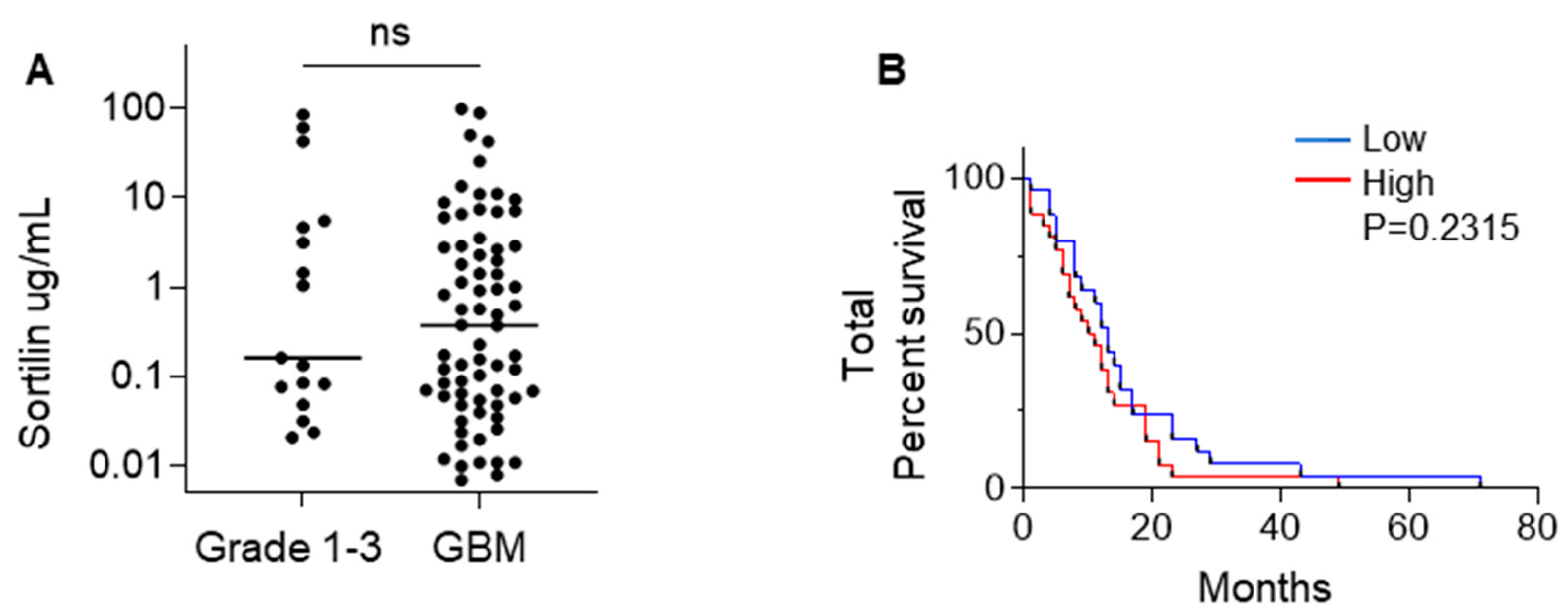
3.3. Sortilin Is Detectable in the Plasma of GBM Patients
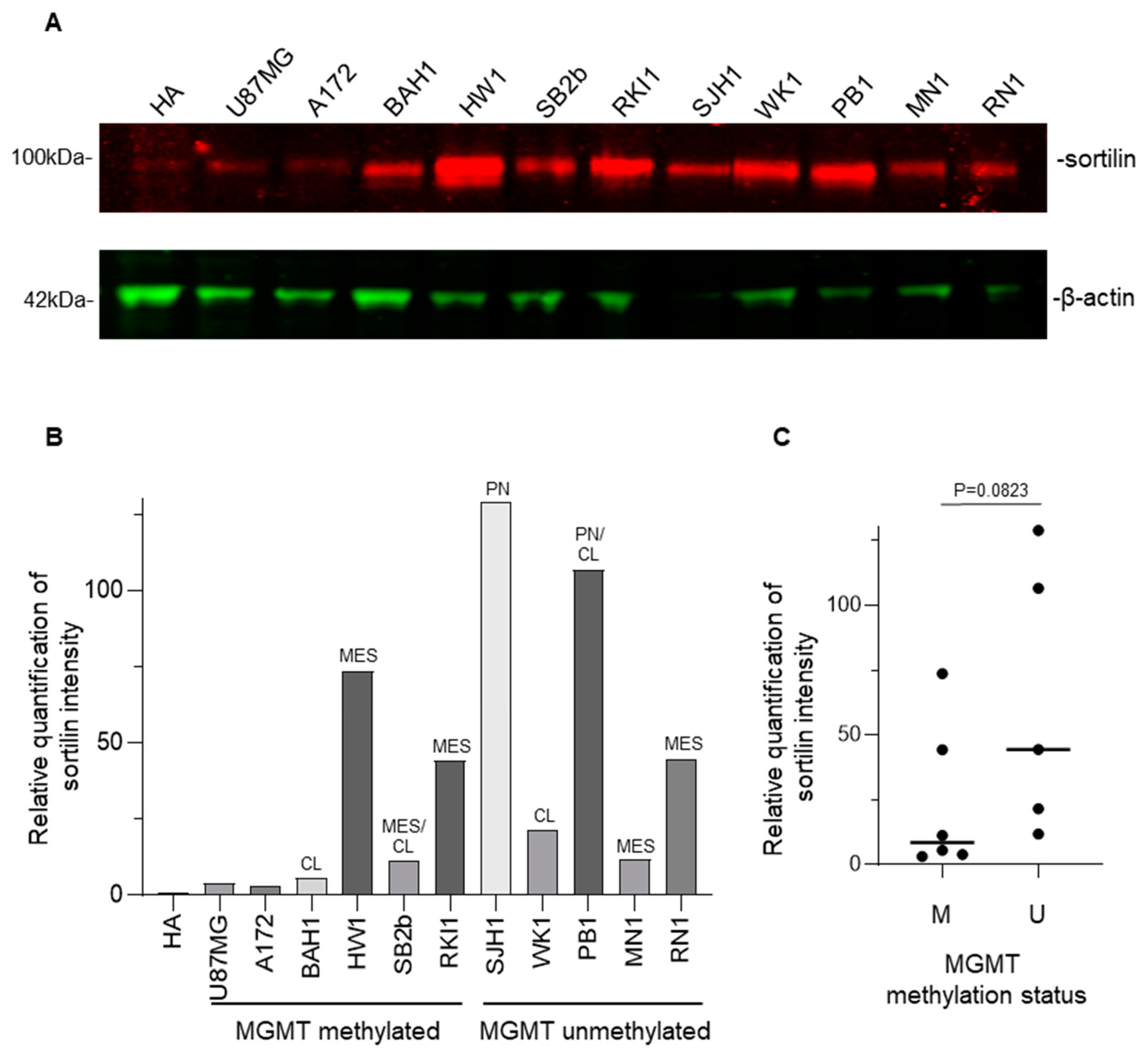
3.4. Sortilin Overexpression in Patient-Derived GBM Cell Lines
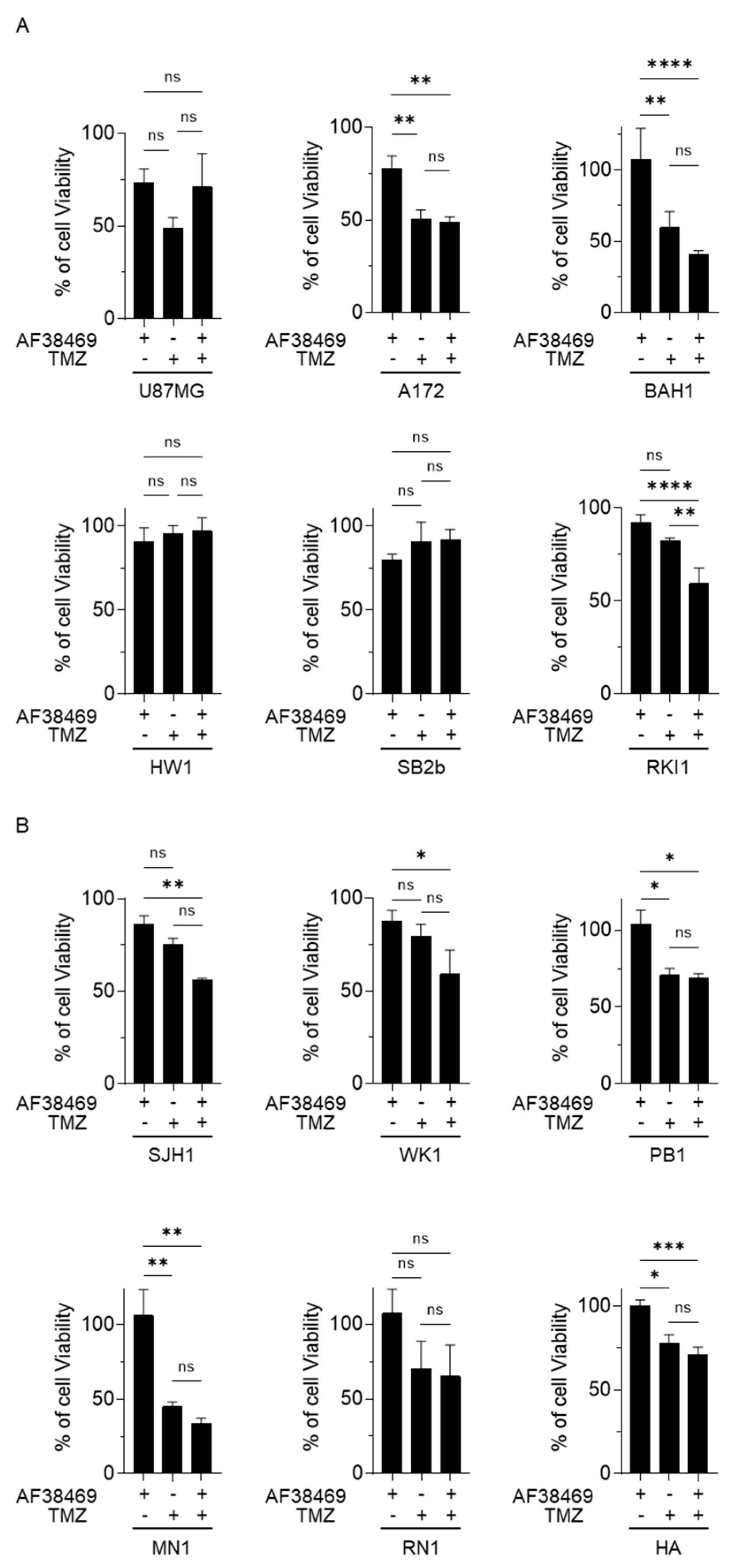
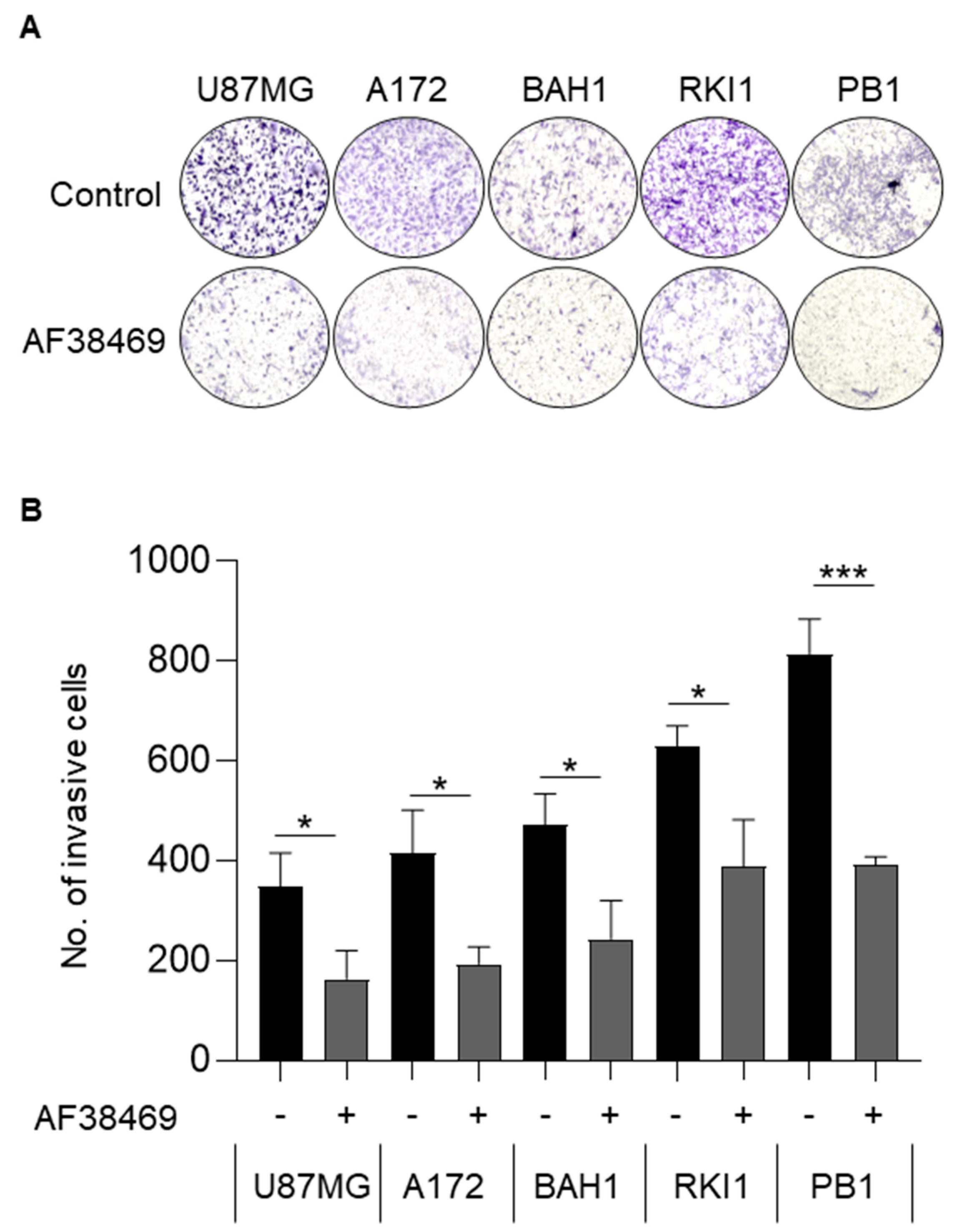
3.5. Targeting Sortilin with Small Molecule Inhibitor AF38469 Inhibits GBM Cell Invasion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Gittleman, H.; Farah, P.; Ondracek, A.; Chen, Y.; Wolinsky, Y.; Stroup, N.E.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2006–2010. Neuro Oncol. 2013, 15 (Suppl. S2), ii1–ii56. [Google Scholar] [CrossRef] [PubMed]
- Skaga, E.; Skretteberg, M.A.; Johannesen, T.B.; Brandal, P.; Vik-Mo, E.O.; Helseth, E.; Langmoen, I.A. Real-world validity of randomized controlled phase III trials in newly diagnosed glioblastoma: To whom do the results of the trials apply? Neurooncol. Adv. 2021, 3, vdab008. [Google Scholar] [CrossRef] [PubMed]
- Iacob, G.; Dinca, E.B. Current data and strategy in glioblastoma multiforme. J. Med. Life 2009, 2, 386–393. [Google Scholar] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef]
- Mazella, J. Deciphering Mechanisms of Action of Sortilin/Neurotensin Receptor-3 in the Proliferation Regulation of Colorectal and Other Cancers. Int. J. Mol. Sci. 2022, 23, 11888. [Google Scholar] [CrossRef]
- Ye, S.; Wang, B.; Zhou, Y.; Sun, Q.; Yang, X. Sortilin 1 regulates hepatocellular carcinoma progression by activating the PI3K/AKT signaling. Hum. Exp. Toxicol. 2022, 41, 09603271221140111. [Google Scholar] [CrossRef]
- Gao, Y.; Li, Y.; Song, Z.; Jin, Z.; Li, X.; Yuan, C. Sortilin 1 Promotes Hepatocellular Carcinoma Cell Proliferation and Migration by Regulating Immune Cell Infiltration. J. Oncol. 2022, 2022, 6509028. [Google Scholar] [CrossRef]
- Gao, F.; Griffin, N.; Faulkner, S.; Li, X.; King, S.J.; Jobling, P.; Denham, J.W.; Jiang, C.C.; Hondermarck, H. The Membrane Protein Sortilin Can Be Targeted to Inhibit Pancreatic Cancer Cell Invasion. Am. J. Pathol. 2020, 190, 1931–1942. [Google Scholar] [CrossRef]
- Demeule, M.; Charfi, C.; Currie, J.C.; Larocque, A.; Zgheib, A.; Kozelko, S.; Béliveau, R.; Marsolais, C.; Annabi, B. TH1902, a new docetaxel-peptide conjugate for the treatment of sortilin-positive triple-negative breast cancer. Cancer Sci. 2021, 112, 4317–4334. [Google Scholar] [CrossRef]
- Rhost, S.; Hughes, É.; Harrison, H.; Rafnsdottir, S.; Jacobsson, H.; Gregersson, P.; Magnusson, Y.; Fitzpatrick, P.; Andersson, D.; Berger, K.; et al. Sortilin inhibition limits secretion-induced progranulin-dependent breast cancer progression and cancer stem cell expansion. Breast Cancer Res. 2018, 20, 137. [Google Scholar] [CrossRef]
- Marsland, M.; Dowdell, A.; Jiang, C.C.; Wilmott, J.S.; Scolyer, R.A.; Zhang, X.D.; Hondermarck, H.; Faulkner, S. Expression of NGF/proNGF and Their Receptors TrkA, p75(NTR) and Sortilin in Melanoma. Int. J. Mol. Sci. 2022, 23, 4260. [Google Scholar] [CrossRef] [PubMed]
- Blondy, S.; Talbot, H.; Saada, S.; Christou, N.; Battu, S.; Pannequin, J.; Jauberteau, M.O.; Lalloué, F.; Verdier, M.; Mathonnet, M.; et al. Overexpression of sortilin is associated with 5-FU resistance and poor prognosis in colorectal cancer. J. Cell. Mol. Med. 2021, 25, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wu, P.-f.; Ma, J.-x.; Liao, M.-j.; Wang, X.-h.; Xu, L.-s.; Xu, M.-h.; Yi, L. Sortilin promotes glioblastoma invasion and mesenchymal transition through GSK-3β/β-catenin/twist pathway. Cell Death Dis. 2019, 10, 208. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Xiang, Y.; Liao, M.-J.; Wu, P.-F.; Yang, L.; Huang, G.-H.; Shi, B.-Z.; Yi, L.; Lv, S.-Q. Presenilin1 inhibits glioblastoma cell invasiveness via promoting Sortilin cleavage. Cell Commun. Signal. 2021, 19, 112. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Carithers, L.J.; Moore, H.M. The Genotype-Tissue Expression (GTEx) Project. Biopreserv. Biobank 2015, 13, 307–308. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Marsland, M.; Dowdell, A.; Faulkner, S.; Jobling, P.; Rush, R.A.; Gedye, C.; Lynam, J.; Griffin, C.P.; Baker, M.; Marsland, J.; et al. ProNGF Expression and Targeting in Glioblastoma Multiforme. Int. J. Mol. Sci. 2023, 24, 1616. [Google Scholar] [CrossRef]
- March, B.; Lockhart, K.R.; Faulkner, S.; Smolny, M.; Rush, R.; Hondermarck, H. ELISA-based quantification of neurotrophic growth factors in urine from prostate cancer patients. FASEB Bioadv. 2021, 3, 888–896. [Google Scholar] [CrossRef]
- Stringer, B.W.; Day, B.W.; D’Souza, R.C.J.; Jamieson, P.R.; Ensbey, K.S.; Bruce, Z.C.; Lim, Y.C.; Goasdoué, K.; Offenhäuser, C.; Akgül, S.; et al. A reference collection of patient-derived cell line and xenograft models of proneural, classical and mesenchymal glioblastoma. Sci. Rep. 2019, 9, 4902. [Google Scholar] [CrossRef] [PubMed]
- Festuccia, C.; Mancini, A.; Colapietro, A.; Gravina, G.L.; Vitale, F.; Marampon, F.; Delle Monache, S.; Pompili, S.; Cristiano, L.; Vetuschi, A.; et al. The first-in-class alkylating deacetylase inhibitor molecule tinostamustine shows antitumor effects and is synergistic with radiotherapy in preclinical models of glioblastoma. J. Hematol. Oncol. 2018, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Pundavela, J.; Demont, Y.; Jobling, P.; Lincz, L.F.; Roselli, S.; Thorne, R.F.; Bond, D.; Bradshaw, R.A.; Walker, M.M.; Hondermarck, H. ProNGF correlates with Gleason score and is a potential driver of nerve infiltration in prostate cancer. Am. J. Pathol. 2014, 184, 3156–3162. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.C.; Marsland, M.; Wang, Y.; Dowdell, A.; Eden, E.; Gao, F.; Faulkner, S.; Jobling, P.; Li, X.; Liu, L.; et al. Tumor innervation is triggered by endoplasmic reticulum stress. Oncogene 2022, 41, 586–599. [Google Scholar] [CrossRef]
- Burgess, D.J. Reaching completion for GTEx. Nat. Rev. Genet. 2020, 21, 717. [Google Scholar] [CrossRef]
- Søreide, K. Receiver-operating characteristic curve analysis in diagnostic, prognostic and predictive biomarker research. J. Clin. Pathol. 2009, 62, 1–5. [Google Scholar] [CrossRef]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef]
- Xiong, J.; Zhou, L.; Yang, M.; Lim, Y.; Zhu, Y.H.; Fu, D.L.; Li, Z.W.; Zhong, J.H.; Xiao, Z.C.; Zhou, X.F. ProBDNF and its receptors are upregulated in glioma and inhibit the growth of glioma cells in vitro. Neuro Oncol. 2013, 15, 990–1007. [Google Scholar] [CrossRef]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef]
- Roselli, S.; Pundavela, J.; Demont, Y.; Faulkner, S.; Keene, S.; Attia, J.; Jiang, C.C.; Zhang, X.D.; Walker, M.M.; Hondermarck, H. Sortilin is associated with breast cancer aggressiveness and contributes to tumor cell adhesion and invasion. Oncotarget 2015, 6, 10473–10486. [Google Scholar] [CrossRef]
- Angelucci, C.; D’Alessio, A.; Lama, G.; Binda, E.; Mangiola, A.; Vescovi, A.L.; Proietti, G.; Masuelli, L.; Bei, R.; Fazi, B.; et al. Cancer stem cells from peritumoral tissue of glioblastoma multiforme: The possible missing link between tumor development and progression. Oncotarget 2018, 9, 28116–28130. [Google Scholar] [CrossRef] [PubMed]
- Molgaard, S.; Demontis, D.; Nicholson, A.M.; Finch, N.A.; Petersen, R.C.; Petersen, C.M.; Rademakers, R.; Nykjaer, A.; Glerup, S. Soluble sortilin is present in excess and positively correlates with progranulin in CSF of aging individuals. Exp. Gerontol. 2016, 84, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Goettsch, C.; Iwata, H.; Hutcheson, J.D.; O’Donnell, C.J.; Chapurlat, R.; Cook, N.R.; Aikawa, M.; Szulc, P.; Aikawa, E. Serum Sortilin Associates With Aortic Calcification and Cardiovascular Risk in Men. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Li, L.; Ren, X.; Sun, Z.; Bao, Z.; Yuan, G.; Cai, H.; Wang, L.; Shao, C.; Wang, Z. Role of Sortilin and Matrix Vesicles in Nε-Carboxymethyl-Lysine-Induced Diabetic Atherosclerotic Calcification. Diabetes Metab. Syndr. Obes. 2020, 13, 4141–4151. [Google Scholar] [CrossRef] [PubMed]
- Schrøder, T.J.; Christensen, S.; Lindberg, S.; Langgård, M.; David, L.; Maltas, P.J.; Eskildsen, J.; Jacobsen, J.; Tagmose, L.; Simonsen, K.B.; et al. The identification of AF38469: An orally bioavailable inhibitor of the VPS10P family sorting receptor Sortilin. Bioorg. Med. Chem. Lett. 2014, 24, 177–180. [Google Scholar] [CrossRef]
- Dong, Z.; Lei, Q.; Yang, R.; Zhu, S.; Ke, X.X.; Yang, L.; Cui, H.; Yi, L. Inhibition of neurotensin receptor 1 induces intrinsic apoptosis via let-7a-3p/Bcl-w axis in glioblastoma. Br. J. Cancer 2017, 116, 1572–1584. [Google Scholar] [CrossRef]
- Faulkner, S.; Jobling, P.; Rowe, C.W.; Rodrigues Oliveira, S.M.; Roselli, S.; Thorne, R.F.; Oldmeadow, C.; Attia, J.; Jiang, C.C.; Zhang, X.D.; et al. Neurotrophin Receptors TrkA, p75(NTR), and Sortilin Are Increased and Targetable in Thyroid Cancer. Am. J. Pathol. 2018, 188, 229–241. [Google Scholar] [CrossRef]
- Currie, J.C.; Demeule, M.; Charfi, C.; Zgheib, A.; Larocque, A.; Danalache, B.A.; Ouanouki, A.; Béliveau, R.; Marsolais, C.; Annabi, B. The Peptide-Drug Conjugate TH1902: A New Sortilin Receptor-Mediated Cancer Therapeutic against Ovarian and Endometrial Cancers. Cancers 2022, 14, 1877. [Google Scholar] [CrossRef]
- Demeule, M.; Charfi, C.; Currie, J.C.; Zgheib, A.; Danalache, B.A.; Béliveau, R.; Marsolais, C.; Annabi, B. The TH1902 Docetaxel Peptide-Drug Conjugate Inhibits Xenografts Growth of Human SORT1-Positive Ovarian and Triple-Negative Breast Cancer Stem-like Cells. Pharmaceutics 2022, 14, 1910. [Google Scholar] [CrossRef]
- Charfi, C.; Demeule, M.; Currie, J.C.; Larocque, A.; Zgheib, A.; Danalache, B.A.; Ouanouki, A.; Béliveau, R.; Marsolais, C.; Annabi, B. New Peptide-Drug Conjugates for Precise Targeting of SORT1-Mediated Vasculogenic Mimicry in the Tumor Microenvironment of TNBC-Derived MDA-MB-231 Breast and Ovarian ES-2 Clear Cell Carcinoma Cells. Front. Oncol. 2021, 11, 760787. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Subgroup | Total |
|---|---|---|
| Participants | n | 91 |
| Sex | Female | 37 (40%) |
| Male | 55 (60%) | |
| Age at diagnosis | Median (min, max) | 63 (17, 82) |
| Median (Q1, Q3) | 63 (56.5, 72) | |
| Grade | 1 | 2 (2.2%) |
| 2 | 6 (6.6%) | |
| 3 | 12 (13.2%) | |
| GBM | 71 (78%) | |
| Tumor site | Frontal | 39 (42%) |
| Temporal | 30 (33%) | |
| Parietal | 15 (16%) | |
| Other | 8 (9%) |
| Parameter | Sortilin Intensity | p-Value | |
|---|---|---|---|
| Low | High | ||
| Sex | 0.5219 | ||
| Female | 16 (44%) | 20 (56%) | |
| Male | 29 (53%) | 26 (47%) | |
| Age | 0.4043 | ||
| ≤63 | 25 (54%) | 21 (46%) | |
| >63 | 20 (44%) | 25 (56%) | |
| Grade | <0.0001 | ||
| 1–3 | 17 (85%) | 3 (15%) | |
| GBM | 28 (39%) | 43 (61%) | |
| Tumor site | 0.7241 | ||
| Frontal | 21 (54%) | 18 (46%) | |
| Temporal | 14 (48%) | 15 (52%) | |
| Other | 10 (43%) | 13 (57%) | |
| Parameter | Sortilin Conc. | p-Value | |
|---|---|---|---|
| Low | High | ||
| Sex | >0.9999 | ||
| Female | 18 (51%) | 17 (49%) | |
| Male | 26 (50%) | 26 (50% | |
| Age | 0.6700 | ||
| ≤63 | 23 (53%) | 20 (47%) | |
| >63 | 21 (48%) | 23 (52%) | |
| Grade | 0.6927 | ||
| 1–3 | 8 (47%) | 9 (53%) | |
| GBM | 35 (50%) | 35 (50%) | |
| Tumor site | 0.6622 | ||
| Frontal | 20 (56%) | 16 (44%) | |
| Temporal | 14 (50%) | 14 (50%) | |
| Other | 10 (43%) | 13 (57%) | |
| Parameter | Sortilin Conc. | Total | p-Value | |
|---|---|---|---|---|
| Low | High | |||
| Sortilin H-score | ||||
| Low | 23 (55%) | 19 (45%) | 87 | <0.0001 |
| High | 5 (11%) | 40 (89%) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marsland, M.; Dowdell, A.; Faulkner, S.; Gedye, C.; Lynam, J.; Griffin, C.P.; Marsland, J.; Jiang, C.C.; Hondermarck, H. The Membrane Protein Sortilin Is a Potential Biomarker and Target for Glioblastoma. Cancers 2023, 15, 2514. https://doi.org/10.3390/cancers15092514
Marsland M, Dowdell A, Faulkner S, Gedye C, Lynam J, Griffin CP, Marsland J, Jiang CC, Hondermarck H. The Membrane Protein Sortilin Is a Potential Biomarker and Target for Glioblastoma. Cancers. 2023; 15(9):2514. https://doi.org/10.3390/cancers15092514
Chicago/Turabian StyleMarsland, Mark, Amiee Dowdell, Sam Faulkner, Craig Gedye, James Lynam, Cassandra P. Griffin, Joanne Marsland, Chen Chen Jiang, and Hubert Hondermarck. 2023. "The Membrane Protein Sortilin Is a Potential Biomarker and Target for Glioblastoma" Cancers 15, no. 9: 2514. https://doi.org/10.3390/cancers15092514