

Nanoparticle Enhancement of Natural Killer (NK) Cell-Based Immunotherapy
by
 , and
, and
Dhanashree Murugan
1,2,
Vasanth Murugesan
2,3,
Balaji Panchapakesan
4,* and
Loganathan Rangasamy
2,*
1
School of Biosciences & Technology (SBST), Vellore Institute of Technology (VIT), Vellore 632014, India
2
Drug Discovery Unit (DDU), Centre for Biomaterials, Cellular and Molecular Theranostics (CBCMT), Vellore Institute of Technology (VIT), Vellore 632014, India
3
School of Advanced Sciences (SAS), Vellore Institute of Technology (VIT), Vellore 632014, India
4
Small Systems Laboratory, Department of Mechanical Engineering, Worcester Polytechnic Institute, Worcester, MA 01609, USA
*
Authors to whom correspondence should be addressed.
Cancers 2022, 14(21), 5438; https://doi.org/10.3390/cancers14215438
Submission received: 9 October 2022
/
Revised: 1 November 2022
/
Accepted: 2 November 2022
/
Published: 4 November 2022
(This article belongs to the Special Issue Natural Killer Cells in Cancer Biology and Therapy)
Abstract
:Simple Summary
Natural killer cells are a part of the native immune response to cancer. NK cell-based immunotherapies are an emerging strategy to kill tumor cells. This paper reviews the role of NK cells, their mechanism of action for killing tumor cells, and the receptors which could serve as potential targets for signaling. In this review, the role of nanoparticles in NK cell activation and increased cytotoxicity of NK cells against cancer are highlighted.
Abstract
Natural killer (NK) cells are one of the first lines of defense against infections and malignancies. NK cell-based immunotherapies are emerging as an alternative to T cell-based immunotherapies. Preclinical and clinical studies of NK cell-based immunotherapies have given promising results in the past few decades for hematologic malignancies. Despite these achievements, NK cell-based immunotherapies have limitations, such as limited performance/low therapeutic efficiency in solid tumors, the short lifespan of NK cells, limited specificity of adoptive transfer and genetic modification, NK cell rejection by the patient’s immune system, insignificant infiltration of NK cells into the tumor microenvironment (TME), and the expensive nature of the treatment. Nanotechnology could potentially assist with the activation, proliferation, near-real time imaging, and enhancement of NK cell cytotoxic activity by guiding their function, analyzing their performance in near-real time, and improving immunotherapeutic efficiency. This paper reviews the role of NK cells, their mechanism of action in killing tumor cells, and the receptors which could serve as potential targets for signaling. Specifically, we have reviewed five different areas of nanotechnology that could enhance immunotherapy efficiency: nanoparticle-assisted immunomodulation to enhance NK cell activity, nanoparticles enhancing homing of NK cells, nanoparticle delivery of RNAi to enhance NK cell activity, genetic modulation of NK cells based on nanoparticles, and nanoparticle activation of NKG2D, which is the master regulator of all NK cell responses.
1. Introduction
Our immune system is a living, breathing medicine that protects the body from various diseases, including cancer [1,2]. Three types of immunity have been identified: innate, adaptive, and passive immunity. Both innate and adaptive immunity systemically fight against the development of neoplastic cells and bacterial and viral infections in the body on a constant basis, which we term “immunosurveillance” [3].
The innate immune system can identify various pathogenic organisms through a molecular immunosurveillance process called pathogen-assisted molecular patterns or “PAMPs” that are not produced by the host (e.g., polysaccharides) [4,5,6]. PAMPs are evolutionarily conserved molecules that are present only in pathogenic organisms. They are recognized by the binding of PAMPs to cell surface and endosomal receptors (e.g., Toll-like receptors) [7]. Immune cells such as macrophages, dendritic cells, NK cells, and some epithelial cells, which form a network of immunosurveillance processes, recognize PAMPs [8]. The second mechanism by which the immune system is alerted during endogenous immunosurveillance is through danger-assisted molecular patterns or “DAMPs” (e.g., calreticulin or CD91) [4].
Cells that are physically, chemically, and biologically stressed or damaged/dying, which can be due to pathogens or other stressors (e.g., chemo or radiation), emit danger signals called DAMPs that alert the immune system [4]. DAMPs can be highly diverse, and their diversity is responsible for immune stimulation, immune modulation, and producing systemic antitumor immunity [4]. Together, PAMPs and alarmins form a network family of DAMPs that aids the immunosurveillance process [9]. Still, cancer cells evade the immunosurveillance process in several ways: loss of cell-adhesion antigens, impairment of cytotoxic T-lymphocytes and NK cells, generation of ligands that block recognition, and secretion of cytokines (e.g., VEGF, IL-10, and TGF-β) that inhibit maturation of dendritic cells [10,11]. In many cancers, tumor growth is supported through immunosuppression that hampers the effective antitumor response and tumor eradication. The tumor microenvironment is often “cold” or “desert-like” meaning they don’t secrete any molecules that could be identified by immune cells [12]. Additionally, tumor-associated macrophages (TAMs) and myeloid-derived suppressor cells (MSDC) envelop the tumor mass, which effectively keeps the infiltration of T-cells at bay [13]. TAMs also keep the immune system suppressed through the following additional pathways: regulation of PD-1 and CTLA-4 [14], T-cell suppression and exhaustion [15], recruitment of Tregs through CCL2 [16], and promotion of inflammation of the tumor microenvironment [16]. TAMs also secrete large amounts of cytokines, such as TGF-β and IL10+, that can impair the cytotoxicity of NK cells [17]. Inflammatory cytokines induced by regulatory or suppressive immune cells promote cancer cell proliferation and suppress the antitumor immune response in the tumor microenvironment. Cells expressing indoleamine 2,3-dioxygenase (IDO) can inhibit the T cell response and lead to immunosuppression of the TME [18,19,20]. Arginase is produced by myeloid-derived suppressor cells and is released into circulation in patients with cancer [21,22]. These enzymes deplete the amino acids necessary for the proper functioning of T-cells. Together, the TME, along with immunosuppressive cells and inhibitory molecules, produces a significant barrier preventing immune attack, making it difficult to treat cancer. This explains some of the reasons for the failure of immunotherapy in many patients.
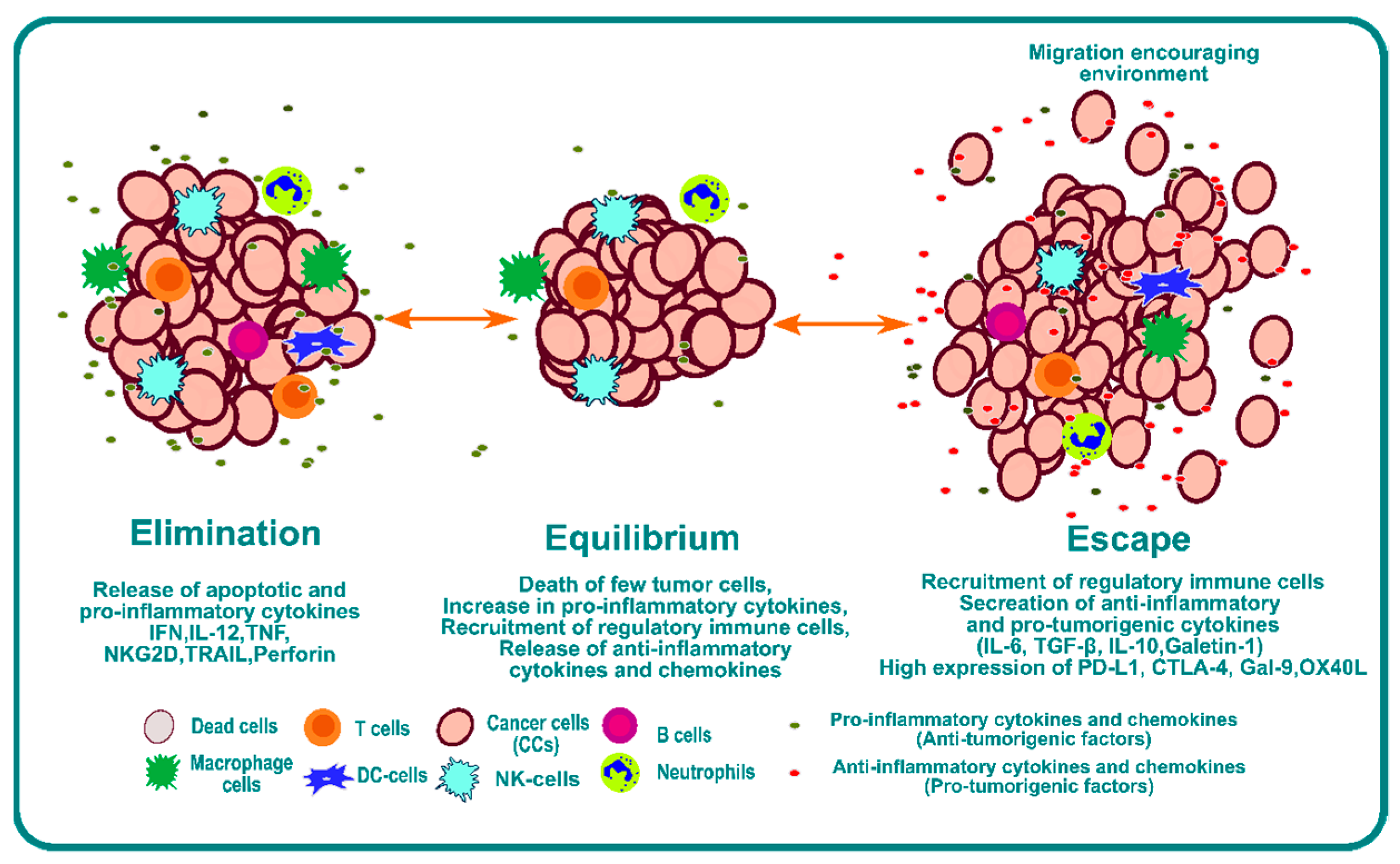
One of the mechanisms of tumor cell escape and subsequent metastasis involves immunoediting of the TME [3,23]. Cancer immunoediting is a dynamic process involving the interplay between tumor cells and immune cells where the tumor cells become less immunogenic over a period of time and develop the capability to escape [24]. Immunoediting can result in protection against cancer through immune-cell-mediated destruction of cancer cells, but it also can result in the escape of cancer cells beyond immune system control, which can lead to proliferation and metastasis. Three phases or “Es” of immunoediting have been identified: elimination, equilibrium, and escape [25] (Figure 1).
The elimination phase presents a phase where a strong immune response is observed due to a combination of both adaptive and innate immunity. This immunoediting phase is beneficial for systemic protection against cancer. Tumor cells have a high expression of surface calreticulin that acts as “eat me” signals for the dendritic cells [26,27,28,29]. Dendritic cells will process tumor antigens and present them on the surface to educate T and NKT cells, which is one of the most critical steps for the success of any form of immunotherapy [30,31]. The effector cells secrete IFN-γ; anti-angiogenic chemokines such as CXCL9, CXCL10, and CXCL11; and cytokines like IL-12 and IFN-γ that enhance macrophage polarization and NK cell activation [32]. Furthermore, Langerhans cells (LCs) secrete IL-15 and can cause the activation of NK and CD8+ T cells [33,34,35].
The second phase is the equilibrium phase. As the name suggests, it creates a microenvironment that contains a balance of pro-inflammatory cytokines and anti-inflammatory cytokines [36]. This phase of immunoediting results in dormancy of tumor growth. Nothing happens in this phase; the tumor is not eradicated, but it also does not grow. This phase is poorly understood, but it is known that the immune system keeps the tumor in a state of functional dormancy in this phase [36].
The third phase is called escape, and it occurs due to the following reasons: the loss of tumor antigens and subsequent loss of immune recognition of tumor cells [37,38]; an increase in secretion of cytokines such as TGF-β [39,40,41], IL-10 [42,43,44], and VEGF [45,46,47]; an increase in transcription factors such as STAT3 [48,49] and BACH2 [50,51]; and overexpression of molecules such as PDL1 and CTLA4, which act as a bridge to inhibit recognition by immune cells. Together, they provide an overarching barrier that prevents immune attack, and they impair the function of immune cells by inducing checkpoint blockade of the immune system. This environment provides enhanced opportunity for invasion and metastasis.
It is now known that cancers are immunogenic. The discovery of immune checkpoint blockade has revolutionized our understanding and created a new field of immuno-oncology. The initial discovery of PD-1 [52] and CTLA-4 [53] led to several promising therapeutic drugs (e.g., Ipilimumab, the first immunotherapy drug that targets CTLA4) for cancer treatment [54]. Ipilimumab blocks the CTLA-4 pathway, which is a T-cell inhibitor that results in the infiltration of cytotoxic T-cells into the TME [53]. Early investigations with Ipilimumab as a monotherapy in advanced metastatic melanoma were highly promising, showing an objective response rate of 40–60% for two years despite adverse side effects in 10–15% of patients [54,55]. The hefty price tag makes it hard to prescribe this as the first line of treatment for most individuals [55]. However, immunotherapy does not work for everyone, and only a fraction of patients benefits from immune-checkpoint-blockade-based therapies. The percentage of patients estimated to respond to checkpoint inhibitor drugs was 0.14% in 2011 and increased to 12.46% in 2018 [56]. Nanotechnology can assist with increasing the efficacy of immunotherapies in a variety of ways. The small size of nanoparticles, 10–100 nm, makes them ideal for modulating immunotherapies in a variety of ways that may not be possible using traditional antibody delivery approaches. These include delivery of immune checkpoint inhibitors (ICI) or antibodies to the TME using encapsulated nanoparticles, creation of tumor antigens through nanoparticle-mediated local ablation of cancer cells and necroptotic cell death, creation of designer nanoparticles with PAMPs and DAMPs that are customized to the TME, education of T cells and NK cells in vitro using nanoparticles based on the mutational status of the TME, delivery of pro-inflammatory cytokines using nanoparticles to neutralize the immunosuppressive environment, and many others. Thus, nanotechnology could potentially enhance immunotherapies and overcome the immunosuppressive environment in a variety of solid cancers. In this review, we focus on the roles NK cells and how nanotechnology can be used to modulate NK cell activity in tumor eradication.
1.1. Natural Killer Cells
Natural Killer (NK) cells are one of the first lines of defense against infections and neoplasms. NK cells are lymphocytes like B cells and T cells, that originate from common lymphoid progenitor cells. NK cells originate from bone marrow and from some secondary lymphoid tissues like the spleen, lymph nodes, or tonsils [57]. NK cells constitute 10–15% of circulating lymphocytes [58]. They are also regulatory cells and interact and communicate with dendritic cells, macrophages, T cells, and endothelial cells [59].
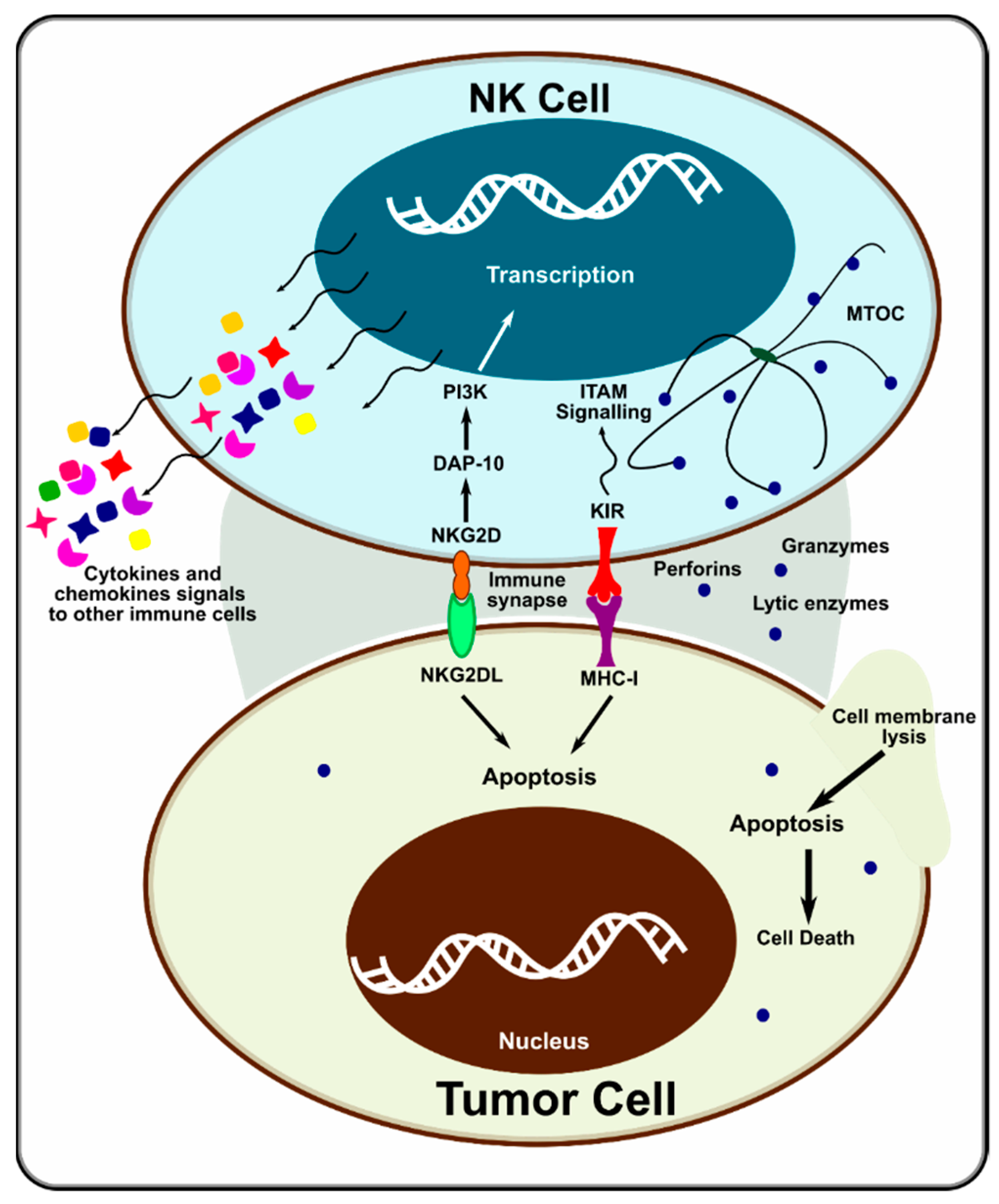
NK cells express MHC class I receptors, which are needed for self-tolerance and functional competence [60]. When encountering MHC class I-deficient hematopoietic cells (e.g., tumor cells), they can recognize the “missing self”. An immunological synapse is formed between the NK and target cell. The polarized lysosomes of the NK cell are activated, and they deliver their cytotoxic contents (e.g., granzymes and perforins) at the synapse, killing the target cell (Figure 2). Downregulation of HLA class I antigen expression in tumor cells also activates NK cells [61]. Many activating receptors have been identified in NK cells in the past decade, including NKG2D, SLAM, DNAX, and NCRs [62]. NK cells use a variety of receptors and pathways to sense their environment, synergistically communicate, and modulate their natural cytotoxicity. Not just one receptor activates NK cells. Only when multiple activating receptors are engaged, and information is acted on synergistically does the cytotoxic activity of NK cells come into effect [62]. This suggests that NK cells have a much more sophisticated control mechanism than T and B cells.
While early ICI-based immunotherapies have focused on T cell proliferation to eradicate the TME, and while recent FDA approval of CAR-T cells is promising, studies have revealed the following disadvantages of CAR-T cells: T cell exhaustion [63,64]; CAR-T cell-associated toxicities may be severe as normal cells surrounding the tumor may also express similar antigens [65]; the response is limited in solid tumors such as breast cancer (ORR: 19%) [66]; and that MHC class I-deficient cells can escape T cells, resulting in poor tumor infiltration by cytotoxic T cells [67]. Due to these shortcomings of T cells, NK cells have recently become more actively researched, with the aim of understanding their functions and controlling their response to selectively work against tumors.
In addition to the direct cytotoxic activity of NK cells, they could also convert the immunologically “cold” microenvironment into a “hot” microenvironment that could render immunotherapy effective.
Figure 2.
Mechanism of action of NK cell on tumor cells. Adapted from [68].
Figure 2.
Mechanism of action of NK cell on tumor cells. Adapted from [68].

1.2. Potential NK Cell Activation Targets
NK cells belong to the innate immune system and thus do not require any prerequisite stimulation like T or B cells [69]. In NK cell-based immunotherapies, strategies should be developed to increase NK cell proliferation, expression levels of cytotoxic agents, infiltration capability, inhibition of inhibitory signaling pathways, and inhibition of NK cell exhaustion. Thus, there are two strategies to develop NK cell-based immunotherapies: to upregulate or activate molecules that would enable the activation of NK cells (such as activation of the NKG2D receptor or delivery of NK cell-activating cytokines (e.g., IL-2, IL-15, or IL-18)); and to inhibit molecules that restrict NK cell activity. For instance, NKG2D, DNAM1, NCR, CD16, NCAM/CD56, CD96, CXCR1, CX3CR1, IL-2/IL-15, CD11, and CCR7 are NK cell-activating receptors; while CD94/NKG2A, TIM-3, TIGIT, KIRs, LAG-3, and PD-1 are NK cell-inhibitory receptors [70]. A list of activating and inhibitory NK cell receptors and their functions is provided in a concise form in the table below (Table 1).
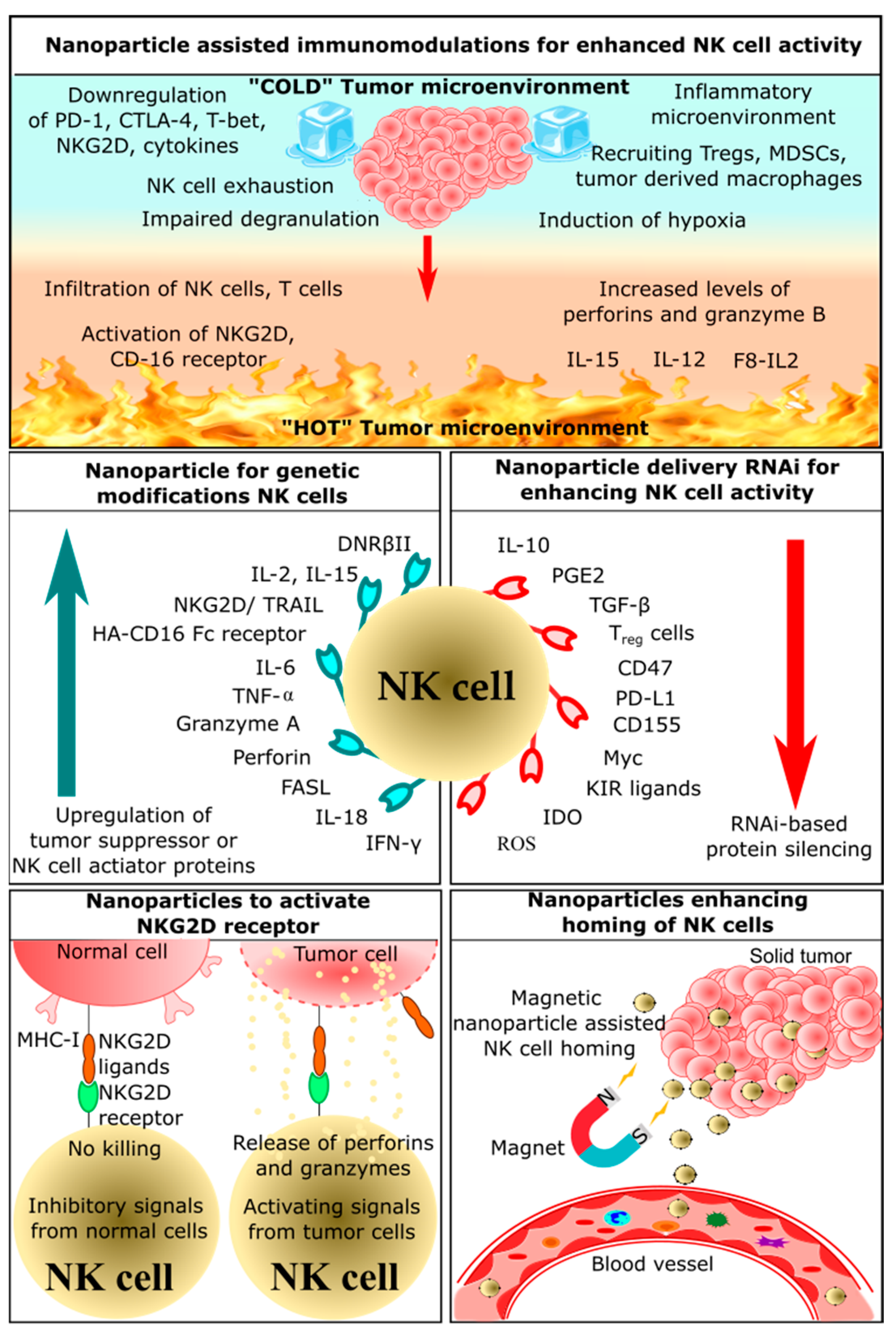
2. Application of Nanoparticles in NK Cell-Based Immunotherapy
The small size of nanoparticles compared to a cell is conducive for delivering drug molecules or cargo. Nanoparticles are not restricted to a delivery vehicle, and they play a pivotal role in the activation of different immune cells, including NK cells. This section of the review summarizes the potential of nanoparticles to assist NK cell-based immunotherapy. Based on the mechanism of action or the target of the immunotherapy, NK cell-based immunotherapies are further subdivided into the following five categories: nanoparticle-assisted immunomodulation to enhance NK cell activity, nanoparticle enhancing homing of NK cells, nanoparticles delivering RNAi to enhance NK cell function, nanoparticles for genetic modification of NK cells, nanoparticles activating NKG2D receptor (Figure 3). NK cell-based nano-immunotherapies are a nascent and developing field, and these therapies might move toward the clinical phase in the coming decades. Currently, CytoSen Therapeutics and KBI Biopharma have developed nanoparticle-based NK cell therapy, and it has entered phase two of a clinical trial [87]. Phase one clinical trials are ongoing for NK cell-based treatment of esophageal cancer [88]. PRECIOUS-01 is a natural killer T cell (iNKT) activating agent that consists of threitolceramide-6 (ThrCer6, IMM60) and New York Esophageal Squamous Cell Carcinoma-1 (NY-ESO-1) cancer-testis antigen peptides encapsulated in a biodegradable polymer poly (lactic-co-glycolic acid) (PLGA) nanoparticle [88]. NY-ESO-1 is overexpressed in many advanced cancers, including lung cancers, bladder cancers, melanoma, and ovarian cancers.
2.1. Nanoparticle-Assisted Immunomodulation for Enhanced NK Cell Activity
Immunomodulation-based cancer therapeutics have recently emerged, as this approach employs the host’s own defense mechanisms to recognize and eliminate cancerous cells. Nanoparticles acting as immunomodulators can improve therapeutic effects and overcome limitations of conventional methods of cancer treatment [92]. There is a broad scope of nanoparticles for drug delivery and an increase in immunotherapeutic efficiency. Some nanoparticles can be used to deliver anticancer drugs, chemokines, and cytokines [93,94,95]. Lipid-based nanoparticles could encapsulate these molecules and effectively deliver them to the tumor site [96,97,98]. Another strategy involves the use of surface-engineered nanoparticles; polymeric or metal-based nanoparticles can be modified such that tumor antigens or antibodies can be docked onto the surface (Table 2) [93,99,100]. For instance, liposome-loaded immunomodulatory agents like TGF-β and IL-2 has also been shown to enhance immune cell infiltration in the tumor microenvironment [101,102]. Similarly, the use of selenium-based nanoparticles that could enhance the function of NK cells has been reported [103]. Selenium-based nanoparticles not only enhance NK cell function but also induce non-specific humoral and cell-mediated immune responses and elevate IL-17 and IFN-γ [103,104,105]. Ruthenium nanoparticles were decorated with bispecific antibodies (SS-Fc, anti-CD16, and anti-CEA), and these triggered immune responses by stimulating NK cells to induce necrosis and apoptosis [106,107].
2.2. Nanoparticles Enhancing Homing of NK Cells
Homing is a mode of signaling between homing receptors present on immune cells and homing receptor ligands on the affected tissues or tumor-secreted molecules [128]. These interactions in a tumor microenvironment could recruit more effector immune cells and enhance immune cell infiltration [128,129]. Cancer cells secrete cytokines and chemokines like TNF-α and ROS, which attracts other effector immune cells. Depending on the function of the cytokines and chemokines, different immunological signals are triggered [130]. These signals could either induce inflammation or activate cytotoxic activity. Chronic inflammation induces tumor cell proliferation, survival, and metastasis [131]. Immune cells involved in inflammation include neutrophils, macrophages, and myeloid-derived suppressor cells [132]. Some of these cytokines induce effector immune cells that could kill cancer cells by activating T cells, B cells, and NK cells [133]. NK cells have a crucial role in the cytotoxicity-mediated killing of tumor cells, and they secrete various cytokines and chemokines that could recruit other immune cells [132]. Once NK cells infiltrate the tumor microenvironment, multiple signaling pathways are triggered by a ligand-receptor interaction, which results in the release of perforin, granzymes, and apoptosis-inducing factors [134]. Hence, it is essential to induce NK cell homing in the tumor microenvironment for effective tumor reduction.
The use of nanoparticles, especially magnetic nanoparticles, for homing immune cells has been explored extensively. For instance, the conjugation of iron oxide nanoparticles on the surface of primary NK cells would significantly enhance the homing of NK cells in the tumor microenvironment. These nanoparticles showed significant antitumor efficacy by increasing the expression of granzymes and perforins in neuroblastoma cells compared with unmodified NK cells [135]. Moreover, the killing of cancer cells by NK cells was associated with the homing of NK cells as well as by external magnetic guidance [135]. These nanoparticles have attracted significant interest for the delivery of immunotherapeutic drugs. Similarly, immunomodulatory magnetic microspheres also increased NK cell infiltration at the tumor site [136]. These microspheres consist of iron oxide nanocubes and recombinant interferon gamma encapsulated in biodegradable poly(lactide-co-glycolide). This architecture allows sustained release of IFN-γ and sensitive MRI T2 contrast agents, which enables both the homing of immune cells and MRI imaging. These microspheres were evaluated on an orthotropic liver tumor VX2 rabbit model, which showed enhanced proliferation and infiltration of NK cells. There are various strategies being adopted for the development of nanoparticle-based NK cell homing. Some of these strategies involve delivering effector molecules such as granzymes, membrane-bound heat shock proteins, and cytokines such as IL-12 using magnetic nanoparticles, which not only enhance homing but also activate NK cells or kill tumor cells (Table 3) [136].
2.3. Nanoparticles Delivering RNAi for Enhancing NK Cells Activity
RNA effectors such as siRNA, miRNA, and shRNA could silence specific genes, which could alter genomic function and enhance antitumor activity. The use of these RNA effectors to enhance NK cell activity is classified under RNAi-mediated immunotherapy [142]. A manganese dioxide (MnO2) nanoparticle system was used to deliver small interfering RNA (siRNA) targeting transforming growth factor-β receptor-2 (TGFBR2), which is known to inhibit the function of NK cells. These nanoparticles loaded with TGFBR2 siRNA protected NK cells from immunosuppression by inhibiting TGFBR2. Thus, silencing TGFBR2 in NK cells made the tumor microenvironment more immunoresponsive by activating NK cells. This suggests that these nanoparticles can be used to enhance the antitumor effects of NK cells through TGFBR2 knockdown and increased expression of IFN-γ [143]. Thus, RNAi-based adoptive NK cell therapy has great potential to improve the survival of cancer patients (Table 4).
EpCAM (epithelial cell adhesion molecule)-targeted cationic liposomes containing si-CD47 and si-PD-L1 were used to knock down immunosuppressive CD47 and PD-L1 [144]. These liposomes effectively prevented the growth of tumors and reduced lung metastasis in 4T1 tumor-bearing mice. These siRNAs could slightly increase the percentage of NK cells, promote the NK cell response, and also increase antibody production. It was also reported that this dual blockade of innate and adaptive immune checkpoint increased the expression of IFN-γ and IL-6 in vivo and in vitro. This suggests that this dual blocking system can be employed to stimulate both adaptive and innate immunity to fight breast cancer [144]. Similarly, another study utilized cationic lipid-assisted nanoparticles encapsulated with siCD155. These nanoparticles were efficient in delivering siCD155 to B16-F10 melanoma cells and macrophages in vitro and in vivo [145]. Downregulation of CD155 promoted the activation of NK and T cells while inhibiting the proliferation of melanoma cells. This suggests that nanoparticle-delivered siCD155 can be used to inhibit melanoma cell proliferation and reprogram the tumor microenvironment with proliferated NK cells and T cells [145].
A novel cocktail strategy was developed by combining NK cell-derived exosomes with miRNA-loaded biomimetic nanoparticles for targeting and therapeutic delivery of miRNA to neuroblastoma cells. NK cell-derived exosomes induced miRNA-loaded nanoparticles to leave systemic circulation and concentrate in tumor cells. One of the major advantages of using NK cell-derived exosomes is that these exosomes might have tumor-specific accumulation and may not be cytotoxic to normal tissues. It was demonstrated that when mice bearing CHLA-255-luc-induced tumors were treated with this cocktail, tumor growth inhibition was observed, which involved synergistic activity between exosomes and miRNA-loaded nanoparticles [95]. Another NK cell-derived exosome was utilized by Neviani et al., where they conjugated NK cell-derived exosomes with miRNA-186. These exosomes exhibited significant cytotoxicity against MYCN gene-amplified neuroblastoma cell lines. Moreover, the cytotoxicity was dependent on the expression of miRNA-186. In vitro studies revealed that these exosomes induced downregulation of TGF-β, which is involved in immune escape. These results suggested that NK cell-derived exosomes loaded with miRNA-186 are a promising therapeutic to promote cytotoxicity of NK cells and block immune escape by tumor cells [146].
2.4. Nanoparticles for Genetic Modification of NK Cells
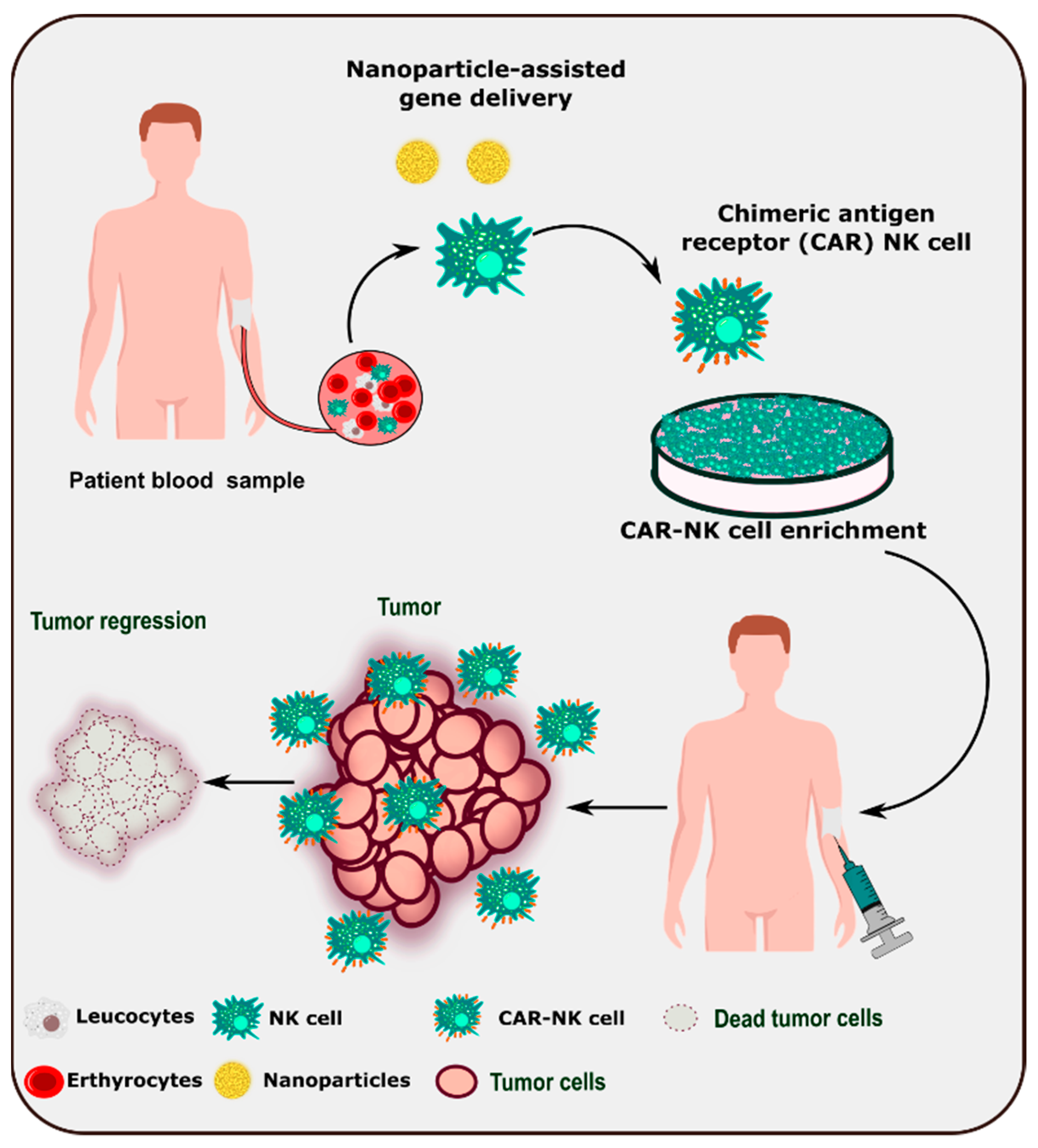
Due to the dynamic role of NK cells in tumor identification and surveillance, adoptive immunotherapy is being developed as a next-generation therapeutic tool. Various strategies have been employed to improve the efficiency and number of NK cells in the tumor microenvironment (Table 5). Understanding NK cell biology and its interactions with the tumor microenvironment enable the modification of NK cells for better and relevant NK cell immunotherapy [147]. Nanoparticle-based delivery of chimeric antigen receptor genes to patient-derived NK cells could lead to the development of CAR-NK cell-based therapy (Figure 4).
Chitosan nanoparticles loaded with IL-2 and NKG2D genes activated NK cells and cytotoxic T cells in vitro [71]. They also showed enhanced tumor accumulation due to the enhanced permeability and retention (EPR) effect and the gathering of lymphocytes in the tumor microenvironment [71]. Consistent with the in vitro results, reduced tumor volumes and improved survival time was observed in CT-26 tumor-bearing mice. These results suggested that nanoparticle-mediated delivery of IL-21 stimulated the antitumor effects of NK cells efficiently, which led to enhanced antitumor activity [71]. In another independent study, chitosan nanoparticles successfully delivered IL-15 and NKG2D genes into cancer cells [148]. These nanoparticles, containing genes for the NKG2D-IL-15 fusion protein, bind to the NKG2D receptor of cytotoxic T cells and NK cells and activate them. IL-15 and NKG2D fusion proteins enhanced the antitumor immune response in B16BL6 melanoma cells by activating >5% of cytotoxic T cells and >50% of NK cells. Moreover, they also showed reduced tumor volume and enhanced survival time in B16BL6 tumor-bearing mice. This suggested that these nanoparticles can be used as a fusion gene vaccine for immunomodulation and tumor growth suppression [148].
An effective method to destroy tumor cells is to use engineered NK cells that consist of dendrimer-entrapped gold nanoparticles containing human ferritin heavy chain (hFTH1) gene-transfected NK cells [149]. These PEG-modified dendrimer-entrapped gold nanoparticles efficiently provided high-quality imaging of transfected NK cells. This system has hFTH1 transfected effectively at a ratio of 5:1 to allow magnetic resonance imaging (MRI) of NK-92 cells and breast cancer cells. Furthermore, these nanoparticles guided NK cells toward the tumor environment for efficient gene therapy in 4T1 tumor-bearing mice. It was suggested that this system could be an efficient vector for genes and also for real-time monitoring with MRI [149]. Similarly, a multi-kinase inhibitor TUS2 (Tumor suppressor candidate 2) gene was delivered using nanovesicles [150]. TUS2 contributes to significant tumor growth reduction in a Kras-mutant syngeneic mouse lung cancer model. Furthermore, it was experimentally found that it increases the levels of circulating and splenic NK cells and CD8+ T cells, decreases the action of Treg cells and MDSCs, and reduces some of the checkpoint receptors such as PD-1, CTLA-4, and TIM-3 [150]. Later in the same study, the anti-PD-1 antibody and the TUS2 plasmid were loaded in the nanovesicle, which then revealed synergistic action against tumor cells via enhanced cytokine-based NK cell activation. A multifunctional magnetic nanoparticle system was synthesized, and it has been shown to be capable of enhancing NK cell function and tracking nanoparticles using magnetic resonance and fluorescence imaging. These multifunctional nanoparticles were designed by applying cationic polydopamine (PDA) coating and plasmid DNA to the surface of magnetic nanoparticles. The magnetic core enabled MRI of NK cells, and the cationic layer enabled them to serve as plasmid DNA carriers. This system showed better cytocompatibility and induced the expression of EGFR targeting chimeric antigen receptors. In vivo results showed that tumor volumes were reduced when treated with these multifunctional nanoparticles in the MDA-MB-231 xenograft mice model. This shows the excellent ability of these cytocompatible multifunctional nanoparticles to potentiate NK cell-mediated antitumor activity and to enable in vivo monitoring [151].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 5.
Comprehensive information on nanoparticle-based genetic modification on NK cells.
| Sr No. | Nanoparticles | Target NK Cell | Ligand | Mechanism of Activation | Effect on Tumor Cells | Ref. |
|---|---|---|---|---|---|---|
| 1. | Chitosan nanoparticles comprised of extracellular NKG2D gene domains and IL-21 gene | - | - |
|
| [71] |
| 2. | Chitosan nanoparticles comprised of extracellular NKG2D gene domains and IL-15 gene | - | - |
|
| [148] |
| 3. | Dendrimer-entrapped gold nanoparticles containing human ferritin heavy chain (hFTH1) gene | - | - |
|
| [149] |
| 4. | Magnetic nanoparticle coated with polydopamine containing plasmid DNA for targeting EGFR chimeric antigen receptor | EGFR | EGFR targeting CAR-NKs |
|
| [151] |
2.5. Nanoparticles Activating NKG2D Receptor
NKG2D is a C-type lectin-like activating receptor expressed on NK cells, NKT cells, and cytotoxic T cells. The ligands of the NKG2D receptor-retinoic acid early induced transcript-1 (RAE-1), H60, UL-16 binding protein like transcript-1, MHC-I chain-related protein A, and ULBPs. These ligands could initiate the NKG2D signaling pathway, which results in the activation, proliferation, and expansion of immune cells [71]. Many researchers have explored these ligands and other novel methods to activate NK cells.
NKG2D receptors initiate ITAM (tyrosine-based activation motif) signaling. Once the signal is triggered, co-stimulatory molecules CD28 and ICOS are triggered and lead to a cascade of reactions that culminate in the activation of PI3K, including reactions involving transcription factors [152]. Activation of these pathways requires targeted delivery of NKG2D ligands or certain cytokines, and nanoparticles mediate the delivery of these ligands. For instance, nanoemulsion of a TGF-β inhibitor and selenocysteine increases the lytic capability of NK cells by sensitizing NKG2D ligands [153]. The TGF-β inhibitor effectively restricted the TGF-β/TGF-β RI/Smad2/3 signaling pathway, which increased the concentration of NKG2D ligands on the tumor cell surface. Selenocysteine supports the expression of NKG2D receptors and suppresses PD-1 expression in γδ T cells [153]. Thus, these combinations enhance the activity of NK cells and act as an appropriate example of adoptive immunotherapy. Some of the other strategies involved are the use of zinc-doped vascular endothelial growth factor (VEGF) receptor-targeted super magnetic nanoparticles, which not only activate NK cells but also trigger magnetic hyperthermia [154]. Further methods for the activation of NK cells are mentioned in Table 6.
3. Limitations of Nanotechnology-Based NK Cell Therapy
As in any immunotherapy, nanotechnology-based NK cell therapy has several limitations. It has been reported that metallic nanoparticles (e.g., Cu) could impose hepatoxicity [157]. The disintegration of chemotherapeutic drugs is mostly dependent on the kidney, whereas degradation of nanoparticles is mostly dependent on the liver. In the process of liver detoxification, many of these nanoparticles accumulate in nearby tissues. This accumulation interrupts various other enzymatic degradation processes [158]. For this reason, it is highly recommended that during nanoparticle evaluation, potential hepatotoxicity is carefully assessed [159]. Similarly, the accumulation of Fe from nanoparticles could potentially cause oxidative stress and ferroptosis, which can potentially affect immune cells and therapy [160].
Although NK cell-based immunotherapies are promising, there are some limitations or challenges in utilizing this therapy. One of the most crucial issues is the uninterrupted supply of NK cells for treatment, which could make treatment very expensive. Clinical trials utilizing NK cells require large numbers, typically 5 × 106 to 5 × 107 cells per kilogram [161]. Sometimes it is difficult to obtain autologous NK cells, which may be due to the health condition of the patients [162]. NK92 cells are generally used in clinical trials and even for the development of CAR-NK cells. NK92 cells are transformed cell lines and thus could be easily enriched in laboratory conditions. However, there are changes in the tumorigenic conversion of these cells, which is a threat during the treatment regimen [163]. However, the use of induced pluripotent stem cells (iPSCs) in the generation of NK cells might be a future strategy to resolve these issues and to obtain large-scale production of NK cells for therapies [164,165]. Another problem is the identification of an appropriate tumor antigen for developing CAR-NK cells that target resistant tumor cells [162].
4. Future Prospects
Our immune system protects the body from a host of diseases, including cancer. The advent of immunotherapy offers the promise of cancer prevention and cure based on activation and control of the immune system. So far, however, the clinical results of immunotherapies have been mixed and work only for a fraction of people as monotherapies. Therefore, a combination approach is being attempted, which has seen incremental therapeutic success. It is becoming more apparent that there are potentially many reasons for immunotherapy failure, including T cell exhaustion, defects in homing molecules on the cell surface, increased MDSC cells in circulation, CAR-T damaging normal tissues, and the release of immunosuppressive cytokines and enzymes that impair T cells, NK cells, and other immune cells.
The idea of using nanoparticles is to force the TME to interact artificially (e.g., blocking CTLA-4) if conventional antibodies fail to elicit a response. Furthermore, nanoparticle-based drugs can also act as multiple lines of defenses by design. Nanoparticles, due to their size, can be designed into multiple different types: nanoparticles to release antigens by necroptotic killing of tumor cells to drive dendritic cell activation, better homing to the TME compared with immune cells, anti-inflammatory nanoparticles to minimize inflammation in the tumor environment in the absence of an immune response, and nanoparticles to aid the proliferation of NK cells. Many of these approaches must be combined for the complete eradication of the TME.
For example, necroptotic cancer cell mimetic nanovaccines have been demonstrated to contain those artificial tumor antigens to induce NKG2D+ NK cells and IFN-γ-expressing CD8+ T cells [166]. Similarly, synthetic vaccine nanoparticles using poly (γ-glutamic acid) tumor antigens and TLR-3 agonists have been demonstrated [167]. These vaccines were able to successfully generate the expansion and activation of NK cells and cytotoxic T cells [167]. It has been observed that certain cytokines such as IL-18, IL-12, IL-15, CD16, CD25, and NKG2A receptors can induce NK cell memories [168]. Nanoparticles inducing these receptors and cytokines could induce cytotoxicity, a higher tumor response, and could trigger memory-like NK cells [168]. Similarly, haptens can also induce memory-like NK cells, and hapten-decorated nanoparticles could be designed for NK cell-based immunotherapies [169]. Conversion of a cold tumor environment to a hot tumor environment is also one of the strategies employed in nano-immunotherapy. Cold tumor environment conversion sometimes includes increased infiltration of immune cells and increased in expression of MHC class I and II [170]. Thus, the synergistic role of nanoparticles could include the possibility of future on-immunotherapies.
5. Conclusions
In conclusion, the disadvantages of T cell-based therapies (namely T cell exhaustion resulting in an inadequate or failed therapeutic effect, CAR-T cell-associated toxicities, the limited response of T cell-based therapies in solid tumors such as breast cancer, and MHC class I-deficient cells escaping and resulting in metastasis) are the reasons for NK cell-based therapy development. NK cells are more sophisticated than T cells, as they can recognize “self” and “missing self”. Recognition of the “missing self” triggers a response in which NK cells kill the target cell using a synapse and deliver cytotoxic cargo. Conventional limitations of NK cells are the reduced lifespan of NK cells in the blood, low activity of NK cells in the suppressive TME, inadequate homing of NK cells, and limited interaction frequency of NK cells with tumor cells [171].
Nanoparticles have proved to possess great potential in the various preclinical studies that have assessed their ability to assist NK cell effectiveness. Nanoparticles could assist NK cells in many ways, including their activation, proliferation, the release of tumor antigens, and removing the chronic inflammatory environment of the tumor. Nanoparticles that assist immunotherapy are attractive due to their small size and diverse choice of nanoparticles. One could potentially influence several receptors on NK cells through nanoparticle–antibody homing, resulting in cytolytic activity. While much is to be understood about the interactions of nanoparticles with NK cells, the present research shows much promise in developing nanoparticle-based immunomodulatory drugs to further increase the effectiveness of immunotherapy.
Author Contributions
Conceptualization, D.M., L.R. and B.P.; writing—original draft preparation, D.M. and V.M.; writing—review and editing, B.P. and L.R.; and supervision, B.P. and L.R. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support from the Department of Biotechnology, New Delhi, for the DBT Ramalingaswami Re-entry Fellowship project (BT/RLF/Re-entry/44/2018) and Science and Engineering Research Board (SERB), New Delhi, for a Core Research Grant (CRG) (CRG/2020/001213), Board of Research in Nuclear Sciences (BRNS) 54/14/03/2022-BRNS/10207 and VIT SEED GRANT 2020-21, 2021-2022 are kindly acknowledged. B.P. acknowledges funding from the United States-India Educational Foundation through the Fulbright-Nehru Academic and Professional Excellence Award on Breast Cancer, Grant Number: 2021/APE-R/63.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Parkin, J.; Cohen, B. An overview of the immune system. Lancet 2001, 357, 1777–1789. [Google Scholar] [CrossRef]
- Delves, P.J.; Roitt, I.M. The Immune System. N. Engl. J. Med. 2000, 343, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The Immunobiology of Cancer Immunosurveillance and Immunoediting. Immunity 2004, 21, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, A.D.; Dudek, A.M.; Agostinis, P. Cancer immunogenicity, danger signals, and DAMPs: What, when, and how? BioFactors 2013, 39, 355–367. [Google Scholar] [CrossRef]
- Hobohm, U.; Stanford, J.L.; Grange, J.M. Pathogen-Associated Molecular Pattern in Cancer Immunotherapy. Crit. Rev. Immunol. 2008, 28, 95–107. [Google Scholar] [CrossRef]
- Locy, H.; DE Mey, S.L.; De Mey, W.; De Ridder, M.; Thielemans, K.; Maenhout, S.K. Immunomodulation of the Tumor Microenvironment: Turn Foe into Friend. Front. Immunol. 2018, 9, 2909. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-Like Receptor Signaling Pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Chalifour, A.; Jeannin, P.; Gauchat, J.-F.; Blaecke, A.; Malissard, M.; N’Guyen, T.; Thieblemont, N.; Delneste, Y. Direct bacterial protein PAMP recognition by human NK cells involves TLRs and triggers α-defensin production. Blood 2004, 104, 1778–1783. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, M.E. DAMPs, PAMPs and alarmins: All we need to know about danger. J. Leukoc. Biol. 2006, 81, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, H.M.C.S.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin. Cancer Biol. 2015, 35, S185–S198. [Google Scholar] [CrossRef]
- Li, Y.; Sun, R. Tumor immunotherapy: New aspects of natural killer cells. Chin. J. Cancer Res. 2018, 30, 173–196. [Google Scholar] [CrossRef] [PubMed]
- Harjunpää, H.; Asens, M.L.; Guenther, C.; Fagerholm, S.C. Cell Adhesion Molecules and Their Roles and Regulation in the Immune and Tumor Microenvironment. Front. Immunol. 2019, 10, 1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petty, A.J.; Yang, Y. Tumor-associated macrophages: Implications in cancer immunotherapy. Immunotherapy 2017, 9, 289–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, Y.; Ji, Q. Tumor-Associated Macrophages Regulate PD-1/PD-L1 Immunosuppression. Front. Immunol. 2022, 13, 874589. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, S.; Zhang, B.; Qiao, L.; Zhang, Y. T Cell Dysfunction and Exhaustion in Cancer. Front. Cell Dev. Biol. 2020, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.-L.; Zhou, W.-J.; Chang, K.-K.; Mei, J.; Huang, L.-Q.; Wang, M.-Y.; Meng, Y.; Ha, S.-Y.; Li, D.-J.; Li, M.-Q. The crosstalk between endometrial stromal cells and macrophages impairs cytotoxicity of NK cells in endometriosis by secreting IL-10 and TGF-β. Reproduction 2017, 154, 815–825. [Google Scholar] [CrossRef] [Green Version]
- Mellor, A.L.; Keskin, D.B.; Johnson, T.; Chandler, P.; Munn, D. Cells Expressing Indoleamine 2,3-Dioxygenase Inhibit T Cell Responses. J. Immunol. 2002, 168, 3771–3776. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Chen, L. Inhibitory B7-family molecules in the tumour microenvironment. Nat. Rev. Immunol. 2008, 8, 467–477. [Google Scholar] [CrossRef]
- Hou, D.-Y.; Muller, A.J.; Sharma, M.D.; DuHadaway, J.; Banerjee, T.; Johnson, M.; Mellor, A.L.; Prendergast, G.C.; Munn, D.H. Inhibition of Indoleamine 2,3-Dioxygenase in Dendritic Cells by Stereoisomers of 1-Methyl-Tryptophan Correlates with Antitumor Responses. Cancer Res. 2007, 67, 792–801. [Google Scholar] [CrossRef]
- Rodríguez, P.C.; Ochoa, A.C. Arginine regulation by myeloid derived suppressor cells and tolerance in cancer: Mechanisms and therapeutic perspectives. Immunol. Rev. 2008, 222, 180–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, P.C.; Ernstoff, M.S.; Hernandez, C.; Atkins, M.; Zabaleta, J.; Sierra, R.; Ochoa, A.C. Arginase I–Producing Myeloid-Derived Suppressor Cells in Renal Cell Carcinoma Are a Subpopulation of Activated Granulocytes. Cancer Res. 2009, 69, 1553–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The Three Es of Cancer Immunoediting. Annu. Rev. Immunol. 2004, 22, 329–360. [Google Scholar] [CrossRef] [PubMed]
- Mittal, D.; Gubin, M.M.; Schreiber, R.D.; Smyth, M.J. New insights into cancer immunoediting and its three component phases—Elimination, equilibrium and escape. Curr. Opin. Immunol. 2014, 27, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubin, M.M.; Vesely, M.D. Cancer Immunoediting in the Era of Immuno-oncology. Clin. Cancer Res. 2022, 28, 3917–3928. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, S.R.; Jammeh, M.L.; Wattenberg, M.M.; Tsang, K.Y.; Ferrone, S.; Hodge, J.W. Radiation-induced immunogenic modulation of tumor enhances antigen processing and calreticulin exposure, resulting in enhanced T-cell killing. Oncotarget 2014, 5, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Fucikova, J.; Kline, J.P.; Galluzzi, L.; Spisek, R. Calreticulin arms NK cells against leukemia. OncoImmunology 2019, 9, 1671763. [Google Scholar] [CrossRef] [Green Version]
- Porcellini, S.; Traggiai, E.; Schenk, U.; Ferrera, D.; Matteoli, M.; Lanzavecchia, A.; Michalak, M.; Grassi, F. Regulation of peripheral T cell activation by calreticulin. J. Exp. Med. 2006, 203, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Zitvogel, L.; Kepp, O.; Senovilla, L.; Menger, L.; Chaput, N.; Kroemer, G. Immunogenic Tumor Cell Death for Optimal Anticancer Therapy: The Calreticulin Exposure Pathway. Clin. Cancer Res. 2010, 16, 3100–3104. [Google Scholar] [CrossRef] [Green Version]
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef]
- O’Donnell, J.S.; Teng, M.W.L.; Smyth, M.J. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat. Rev. Clin. Oncol. 2019, 16, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, P.; Lasagni, L.; Annunziato, F.; Serio, M.; Romagnani, S. CXC chemokines: The regulatory link between inflammation and angiogenesis. Trends Immunol. 2004, 25, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Yanofsky, V.R.; Mitsui, H.; Felsen, D.; Carucci, J.A. Understanding Dendritic Cells and Their Role in Cutaneous Carcinoma and Cancer Immunotherapy. Clin. Dev. Immunol. 2013, 2013, 624123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, K.C.; Lugli, E.; Welles, H.C.; Rosenberg, S.A.; Fojo, A.T.; Morris, J.C.; Fleisher, T.A.; Dubois, S.P.; Perera, L.P.; Stewart, D.M.; et al. Redistribution, Hyperproliferation, Activation of Natural Killer Cells and CD8 T Cells, and Cytokine Production During First-in-Human Clinical Trial of Recombinant Human Interleukin-15 in Patients with Cancer. J. Clin. Oncol. 2015, 33, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, E.; Rossi, M.; Ratzinger, G.; De Cos, M.-A.; Chung, D.J.; Panageas, K.S.; Wolchock, J.D.; Houghton, A.N.; Chapman, P.B.; Heller, G.; et al. Peptide-Loaded Langerhans Cells, Despite Increased IL15 Secretion and T-Cell Activation In Vitro, Elicit Antitumor T-Cell Responses Comparable to Peptide-Loaded Monocyte-Derived Dendritic Cells In Vivo. Clin. Cancer Res. 2011, 17, 1984–1997. [Google Scholar] [CrossRef] [Green Version]
- Padoan, A.; Plebani, M.; Basso, D. Inflammation and Pancreatic Cancer: Focus on Metabolism, Cytokines, and Immunity. Int. J. Mol. Sci. 2019, 20, 676. [Google Scholar] [CrossRef] [Green Version]
- Olson, B.M.; McNeel, D. Antigen loss and tumor-mediated immunosuppression facilitate tumor recurrence. Expert Rev. Vaccines 2012, 11, 1315–1317. [Google Scholar] [CrossRef]
- Saleh, R.; Elkord, E. Acquired resistance to cancer immunotherapy: Role of tumor-mediated immunosuppression. Semin. Cancer Biol. 2019, 65, 13–27. [Google Scholar] [CrossRef]
- Pardali, K.; Moustakas, A. Actions of TGF-β as tumor suppressor and pro-metastatic factor in human cancer. Biochim. Biophys. Acta Rev. Cancer. 2007, 1775, 21–62. [Google Scholar] [CrossRef]
- Wojtowicz-Praga, S. Reversal of Tumor-Induced Immunosuppression: A New Approach to Cancer Therapy. J. Immunother. 1997, 20, 165–177. [Google Scholar] [CrossRef]
- Gold, L.I. The role for transforming growth factor-beta (TGF-beta) in human cancer. Crit. Rev. Oncog. 1999, 10, 303–360. [Google Scholar] [PubMed]
- Mittal, S.K.; Cho, K.-J.; Ishido, S.; Roche, P.A. Interleukin 10 (IL-10)-mediated Immunosuppression. J. Biol. Chem. 2015, 290, 27158–27167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamichhane, P.; Karyampudi, L.; Shreeder, B.; Krempski, J.; Bahr, D.; Daum, J.; Kalli, K.R.; Goode, E.L.; Block, M.S.; Cannon, M.J.; et al. IL10 Release upon PD-1 Blockade Sustains Immunosuppression in Ovarian Cancer. Cancer Res. 2017, 77, 6667–6678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frumento, G.; Piazza, T.; Di Carlo, E.; Ferrini, S. Targeting Tumor-Related Immunosuppression for Cancer Immunotherapy. Endocr. Metab. Immune Disord. Drug Targets 2006, 6, 223–237. [Google Scholar] [CrossRef]
- Lapeyre-Prost, A.; Terme, M.; Pernot, S.; Pointet, A.-L.; Voron, T.; Tartour, E.; Taieb, J. Immunomodulatory Activity of VEGF in Cancer. Int. Rev. Cell Mol. Biol. 2017, 330, 295–342. [Google Scholar] [CrossRef]
- Johnson, B.F.; Clay, T.M.; Hobeika, A.C.; Lyerly, H.; Morse, M.A. Vascular endothelial growth factor and immunosuppression in cancer: Current knowledge and potential for new therapy. Expert Opin. Biol. Ther. 2007, 7, 449–460. [Google Scholar] [CrossRef]
- Li, Y.-L.; Zhao, H.; Ren, X.-B. Relationship of VEGF/VEGFR with immune and cancer cells: Staggering or forward? Cancer Biol. Med. 2016, 13, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Bu, L.; Yu, G.; Wu, L.; Mao, L.; Deng, W.; Liu, J.; Kulkarni, A.; Zhang, W.; Zhang, L.; Sun, Z. STAT3 Induces Immunosuppression by Upregulating PD-1/PD-L1 in HNSCC. J. Dent. Res. 2017, 96, 1027–1034. [Google Scholar] [CrossRef]
- Lee, H.; Pal, S.K.; Reckamp, K.; Figlin, R.A.; Yu, H. STAT3: A Target to Enhance Antitumor Immune Response. Curr. Top Microbiol. Immunol. 2010, 344, 41–59. [Google Scholar] [CrossRef] [Green Version]
- Roychoudhuri, R.; Eil, R.L.; Clever, D.; Klebanoff, C.A.; Sukumar, M.; Grant, F.M.; Yu, Z.; Mehta, G.; Liu, H.; Jin, P.; et al. The transcription factor BACH2 promotes tumor immunosuppression. J. Clin. Investig. 2016, 126, 599–604. [Google Scholar] [CrossRef]
- Grant, F.M.; Yang, J.; Nasrallah, R.; Clarke, J.; Sadiyah, F.; Whiteside, S.K.; Imianowski, C.J.; Kuo, P.; Vardaka, P.; Todorov, T.; et al. BACH2 drives quiescence and maintenance of resting Treg cells to promote homeostasis and cancer immunosuppression. J. Exp. Med. 2020, 217, e20190711. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Agata, Y.; Shibahara, K.; Honjo, T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. 1992, 11, 3887–3895. [Google Scholar] [CrossRef] [PubMed]
- Leach, D.R.; Krummel, M.F.; Allison, J.P. Enhancement of Antitumor Immunity by CTLA-4 Blockade. Science 1996, 271, 1734–1736. [Google Scholar] [CrossRef] [Green Version]
- Hodi, F.S.; O’Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved Survival with Ipilimumab in Patients with Metastatic Melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Fellner, C. Ipilimumab (yervoy) prolongs survival in advanced melanoma: Serious side effects and a hefty price tag may limit its use. Pharm. Ther. 2012, 37, 503–530. [Google Scholar]
- Haslam, A.; Prasad, V. Estimation of the Percentage of US Patients with Cancer Who Are Eligible for and Respond to Checkpoint Inhibitor Immunotherapy Drugs. JAMA Netw. Open 2019, 2, e192535. [Google Scholar] [CrossRef] [Green Version]
- Freud, A.G.; Yu, J.; Caligiuri, M.A. Human natural killer cell development in secondary lymphoid tissues. Semin. Immunol. 2014, 26, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Töpfer, K.; Kempe, S.; Müller, N.; Schmitz, M.; Bachmann, M.; Cartellieri, M.; Schackert, G.; Temme, A. Tumor Evasion from T Cell Surveillance. J. Biomed. Biotechnol. 2011, 2011, 918471. [Google Scholar] [CrossRef] [Green Version]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Yawata, M.; Yawata, N.; Draghi, M.; Partheniou, F.; Little, A.-M.; Parham, P. MHC class I–specific inhibitory receptors and their ligands structure diverse human NK-cell repertoires toward a balance of missing self-response. Blood 2008, 112, 2369–2380. [Google Scholar] [CrossRef] [Green Version]
- Long, E.O. Tumor cell recognition by natural killer cells. Semin. Cancer Biol. 2002, 12, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Molfetta, R.; Quatrini, L.; Santoni, A.; Paolini, R. Regulation of NKG2D-Dependent NK Cell Functions: The Yin and the Yang of Receptor Endocytosis. Int. J. Mol. Sci. 2017, 18, 1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauken, K.E.; Wherry, E.J. Overcoming T cell exhaustion in infection and cancer. Trends Immunol. 2015, 36, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, J.S.; Cox, M.A.; Zajac, A.J. T-Cell Exhaustion: Characteristics, Causes and Conversion. Immunology 2010, 129, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Brudno, J.N.; Kochenderfer, J.N. Toxicities of chimeric antigen receptor T cells: Recognition and management. Blood 2016, 127, 3321–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Zou, X.; Zheng, S.; Tang, H.; Zhang, L.; Liu, P.; Xie, X. Efficacy and predictive factors of immune checkpoint inhibitors in metastatic breast cancer: A systematic review and meta-analysis. Ther. Adv. Med. Oncol. 2020, 12, 175883592094092. [Google Scholar] [CrossRef]
- Garrido, F.; Aptsiauri, N.; Doorduijn, E.M.; Lora, A.M.G.; van Hall, T. The urgent need to recover MHC class I in cancers for effective immunotherapy. Curr. Opin. Immunol. 2016, 39, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.R.; Kim, T.-D.; Choi, I. Understanding of molecular mechanisms in natural killer cell therapy. Exp. Mol. Med. 2015, 47, e141. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wang, X.; Jin, T.; Tian, Y.; Dai, C.; Widarma, C.; Song, R.; Xu, F. Immune checkpoint molecules in natural killer cells as potential targets for cancer immunotherapy. Signal Transduct. Target. Ther. 2020, 5, 250. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, Y. Targeting NK Cell Checkpoint Receptors or Molecules for Cancer Immunotherapy. Front. Immunol. 2020, 11, 1295. [Google Scholar] [CrossRef]
- Tan, L.; Han, S.; Ding, S.; Xiao, W.; Ding, Y.; Qian, L.; Wang, C.; Gong, W. Chitosan nanoparticle-based delivery of fused NKG2D-IL-21 gene suppresses colon cancer growth in mice. Int. J. Nanomed. 2017, 12, 3095–3107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, V.; Wu, N.; Lu, Y.; Davidson, D.; Colonna, M.; Veillette, A. DNAM-1 controls NK cell activation via an ITT-like motif. J. Exp. Med. 2015, 212, 2165–2182. [Google Scholar] [CrossRef] [PubMed]
- Konjević, G.; Vuletić, A.; Martinović, K.M.; Džodić, K.M.M.a.R. The Role of Activating and Inhibitory NK Cell Receptors in Antitumor Immune Response. In Natural Killer Cells; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Watzl, C. Chapter Five—How to Trigger a Killer: Modulation of Natural Killer Cell Reactivity on Many Levels. Adv. Immunol. 2014, 124, 137–170. [Google Scholar] [CrossRef] [PubMed]
- Van Acker, H.H.; Capsomidis, A.; Smits, E.L.; Van Tendeloo, V.F. CD56 in the Immune System: More Than a Marker for Cytotoxicity? Front. Immunol. 2017, 8, 892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, A.; Cella, M.; Giurisato, E.; Shaw, A.S.; Colonna, M. Cutting Edge: CD96 (Tactile) Promotes NK Cell-Target Cell Adhesion by Interacting with the Poliovirus Receptor (CD155). J. Immunol. 2004, 172, 3994–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, Y.Y.; Tay, J.C.; Wang, S. CXCR1 Expression to Improve Anti-Cancer Efficacy of Intravenously Injected CAR-NK Cells in Mice with Peritoneal Xenografts. Mol. Ther. Oncolytics 2019, 16, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas-Fuentes, S.; Salgado-Aguayo, A.; Arratia-Quijada, J.; Gorocica-Rosete, P. Regulation and biological functions of the CX3CL1-CX3CR1 axis and its relevance in solid cancer: A mini-review. J. Cancer 2021, 12, 571–583. [Google Scholar] [CrossRef]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef] [Green Version]
- Somanchi, S.S.; Somanchi, A.; Cooper, L.J.N.; Lee, D.A. Engineering lymph node homing of ex vivo–expanded human natural killer cells via trogocytosis of the chemokine receptor CCR7. Blood 2012, 119, 5164–5172. Available online: https://ashpublications.org/blood/article/119/22/5164/105509/Engineering-lymph-node-homing-of-ex-vivo-expanded (accessed on 27 September 2022). [CrossRef]
- Borrego, F.; Masilamani, M.; Kabat, J.; Sanni, T.B.; Coligan, J.E. The cell biology of the human natural killer cell CD94/NKG2A inhibitory receptor. Mol. Immunol. 2005, 42, 485–488. [Google Scholar] [CrossRef]
- Tang, R.; Rangachari, M.; Kuchroo, V.K. Tim-3: A co-receptor with diverse roles in T cell exhaustion and tolerance. Semin. Immunol. 2019, 42, 101302. [Google Scholar] [CrossRef] [PubMed]
- Stanietsky, N.; Simic, H.; Arapovic, J.; Toporik, A.; Levy, O.; Novik, A.; Levine, Z.; Beiman, M.; Dassa, L.; Achdout, H.; et al. The interaction of TIGIT with PVR and PVRL2 inhibits human NK cell cytotoxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 17858–17863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pende, D.; Falco, M.; Vitale, M.; Cantoni, C.; Vitale, C.; Munari, E.; Bertaina, A.; Moretta, F.; Del Zotto, G.; Pietra, G.; et al. Killer Ig-Like Receptors (KIRs): Their Role in NK Cell Modulation and Developments Leading to Their Clinical Exploitation. Front. Immunol. 2019, 10, 1179. Available online: https://www.frontiersin.org/articles/10.3389/fimmu.2019.01179 (accessed on 27 September 2022). [CrossRef] [PubMed] [Green Version]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and TIGIT: Co-inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, C.; Li, M.; Zhu, S.; Chen, Y.; Zhou, L.; Xu, D.; Xu, J.; Li, Z.; Li, W.; Cui, J. PD-1-positive Natural Killer Cells have a weaker antitumor function than that of PD-1-negative Natural Killer Cells in Lung Cancer. Int. J. Med. Sci. 2020, 17, 1964–1973. [Google Scholar] [CrossRef] [PubMed]
- Press Release|CytoSen Therapeutics and KBI Biopharma Enter into Strategic Partnership to Manufacture NK Cells and Nanoparticles. Available online: https://www.kbibiopharma.com/news/cytosen-therapeutics-and-kbi-biopharma-enter-into-strategic-partnership-to-manufacture-nk-cells-and-nanoparticles (accessed on 30 October 2022).
- Dolen, Y.; Kreutz, M.; Gileadi, U.; Tel, J.; Vasaturo, A.; Van Dinther, E.A.W.; Van Hout-Kuijer, M.A.; Cerundolo, V.; Figdor, C.G. Co-delivery of PLGA encapsulated invariant NKT cell agonist with antigenic protein induce strong T cell-mediated antitumor immune responses. OncoImmunology 2015, 5, e1068493. [Google Scholar] [CrossRef] [Green Version]
- Burga, R.A.; Khan, D.H.; Agrawal, N.; Bollard, C.M.; Fernandes, R. Designing Magnetically Responsive Biohybrids Composed of Cord Blood-Derived Natural Killer Cells and Iron Oxide Nanoparticles. Bioconjug. Chem. 2019, 30, 552–560. [Google Scholar] [CrossRef]
- Nayyar, G.; Chu, Y.; Cairo, M.S. Overcoming Resistance to Natural Killer Cell Based Immunotherapies for Solid Tumors. Front. Oncol. 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Jonjic, S. Manipulation of NKG2D ligands by cytomegaloviruses: Impact on innate and adaptive immune response. Front. Immunol. 2011, 2, 85. [Google Scholar] [CrossRef] [Green Version]
- Jindal, A.; Sarkar, S.; Alam, A. Nanomaterials-Mediated Immunomodulation for Cancer Therapeutics. Front. Chem. 2021, 9, 629635. [Google Scholar] [CrossRef]
- Mikelez-Alonso, I.; Magadán, S.; González-Fernández, F.; Borrego, F. Natural killer (NK) cell-based immunotherapies and the many faces of NK cell memory: A look into how nanoparticles enhance NK cell activity. Adv. Drug Deliv. Rev. 2021, 176, 113860. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Hou, W.; Liu, Y.; Wang, W.; Han, Y.; Yang, M.; Zhi, X.; Li, C.; Qi, D.; Li, T.; et al. Cytokine induced killer cells-assisted delivery of chlorin e6 mediated self-assembled gold nanoclusters to tumors for imaging and immuno-photodynamic therapy. Biomaterials 2018, 170, 1–11. [Google Scholar] [CrossRef]
- Wang, G.; Hu, W.; Chen, H.; Shou, X.; Ye, T.; Xu, Y. Cocktail Strategy Based on NK Cell-Derived Exosomes and Their Biomimetic Nanoparticles for Dual Tumor Therapy. Cancers 2019, 11, 1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshy, S.T.; Cheung, A.S.; Gu, L.; Graveline, A.R.; Mooney, D.J. Liposomal Delivery Enhances Immune Activation by STING Agonists for Cancer Immunotherapy. Adv. Biosyst. 2017, 1, 1600013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Fong, C.I.; Xu, M.; Han, B.-N.; Yuan, Z.; Zhao, Q. Nano-loaded natural killer cells as carriers of indocyanine green for synergetic cancer immunotherapy and phototherapy. J. Innov. Opt. Health Sci. 2019, 12, 19410025. [Google Scholar] [CrossRef] [Green Version]
- Pitchaimani, A.; Nguyen, T.D.T.; Aryal, S. Natural killer cell membrane infused biomimetic liposomes for targeted tumor therapy. Biomaterials 2018, 160, 124–137. [Google Scholar] [CrossRef]
- Liu, Z.; Jiang, W.; Nam, J.; Moon, J.J.; Kim, B.Y.S. Immunomodulating Nanomedicine for Cancer Therapy. Nano Lett. 2018, 18, 6655–6659. [Google Scholar] [CrossRef] [Green Version]
- Christian, D.A.; Hunter, C.A. Particle-mediated delivery of cytokines for immunotherapy. Immunotherapy 2012, 4, 425–441. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Wrzesinski, S.H.; Stern, E.; Look, M.; Criscione, J.M.; Ragheb, R.; Jay, S.; Demento, S.L.; Agawu, A.; Limon, P.L.; et al. Combination delivery of TGF-β inhibitor and IL-2 by nanoscale liposomal polymeric gels enhances tumour immunotherapy. Nat. Mater. 2012, 11, 895–905. [Google Scholar] [CrossRef]
- McHugh, M.D.; Park, J.; Uhrich, R.; Gao, W.; Horwitz, D.A.; Fahmy, T.M. Paracrine co-delivery of TGF-β and IL-2 using CD4-targeted nanoparticles for induction and maintenance of regulatory T cells. Biomaterials 2015, 59, 172–181. [Google Scholar] [CrossRef]
- Gao, S.; Li, T.; Guo, Y.; Sun, C.; Xianyu, B.; Xu, H. Selenium-Containing Nanoparticles Combine the NK Cells Mediated Immunotherapy with Radiotherapy and Chemotherapy. Adv. Mater. 2020, 32, e1907568. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, M.H.; Mahdavi, M.; Setayesh, N.; Esfandyar, M.; Shahverdi, A.R. Selenium nanoparticle-enriched Lactobacillus brevis causes more efficient immune responses in vivo and reduces the liver metastasis in metastatic form of mouse breast cancer. DARU J. Pharm. Sci. 2013, 21, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferro, C.; Florindo, H.F.; Santos, H.A. Selenium Nanoparticles for Biomedical Applications: From Development and Characterization to Therapeutics. Adv. Health Mater. 2021, 10, e2100598. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wen, Y.; Liu, Y.; Tan, X.; Chen, X.; Zhu, X.; Wei, C.; Chen, L.; Wang, Z.; Liu, J.; et al. Hollow mesoporous ruthenium nanoparticles conjugated bispecific antibody for targeted anti-colorectal cancer response of combination therapy. Nanoscale 2019, 11, 9661–9678. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; He, L.; Li, X.; Xu, L.; Chen, T. Ruthenium complexes boost NK cell immunotherapy via sensitizing triple-negative breast cancer and shaping immuno-microenvironment. Biomaterials 2022, 281, 121371. [Google Scholar] [CrossRef]
- Li, T.; Pan, S.; Gao, S.; Xiang, W.; Sun, C.; Cao, W.; Xu, H. Diselenide–Pemetrexed Assemblies for Combined Cancer Immuno-, Radio-, and Chemotherapies. Angew. Chem. 2019, 132, 2722–2726. [Google Scholar] [CrossRef]
- Chen, H.; Liu, L.; Ma, A.; Yin, T.; Chen, Z.; Liang, R.; Qiu, Y.; Zheng, M.; Cai, L. Noninvasively immunogenic sonodynamic therapy with manganese protoporphyrin liposomes against triple-negative breast cancer. Biomaterials 2021, 269, 120639. [Google Scholar] [CrossRef]
- Liu, B.; Cao, W.; Cheng, E.A.J.; Fan, S.; Pan, S.; Wang, L.; Niu, J.; Pan, Y.; Liu, Y.; Sun, X.; et al. Human natural killer cells for targeting delivery of gold nanostars and bimodal imaging directed photothermal/photodynamic therapy and immunotherapy. Cancer Biol. Med. 2019, 16, 756–770. [Google Scholar] [CrossRef]
- Yang, X.; Lai, C.; Liu, A.; Hou, X.; Tang, Z.; Mo, F.; Yin, S.; Lu, X. Anti-Tumor Activity of Mannose-CpG-Oligodeoxynucleotides-Conjugated and Hepatoma Lysate-Loaded Nanoliposomes for Targeting Dendritic Cells In Vivo. J. Biomed. Nanotechnol. 2019, 15, 1018–1032. [Google Scholar] [CrossRef]
- Kim, H.; Sehgal, D.; Kucaba, T.A.; Ferguson, D.M.; Griffith, T.S.; Panyam, J. Acidic pH-responsive polymer nanoparticles as a TLR7/8 agonist delivery platform for cancer immunotherapy. Nanoscale 2018, 10, 20851–20862. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Wang, Y.-H.; Lei, C.-S.; Changou, C.A.; Davis, M.E.; Yen, Y. Host immune response to anti-cancer camptothecin conjugated cyclodextrin-based polymers. J. Biomed. Sci. 2019, 26, 85. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-S.; Han, J.-H.; Choi, S.H.; Jung, H.-Y.; Park, J.D.; An, H.-J.; Kim, S.-E.; Kim, D.-H.; Doh, J.; Han, D.K.; et al. Cationic Nanoparticle-Mediated Activation of Natural Killer Cells for Effective Cancer Immunotherapy. ACS Appl. Mater. Interfaces 2020, 12, 56731–56740. [Google Scholar] [CrossRef] [PubMed]
- Loftus, C.; Saeed, M.; Davis, D.M.; Dunlop, I.E. Activation of Human Natural Killer Cells by Graphene Oxide-Templated Antibody Nanoclusters. Nano Lett. 2018, 18, 3282–3289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegler, E.L.; Kim, Y.J.; Chen, X.; Siriwon, N.; Mac, J.; Rohrs, J.A.; Bryson, P.D.; Wang, P. Combination Cancer Therapy Using Chimeric Antigen Receptor-Engineered Natural Killer Cells as Drug Carriers. Mol. Ther. 2017, 25, 2607–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Qi, M.; Ding, F.; Lv, Y.; Ma, J.; Zhu, Y. Tumour targetable and microenvironment-responsive nanoparticles simultaneously disrupt the PD-1/PD-L1 pathway and MAPK/ERK/JNK pathway for efficient treatment of colorectal cancer. J. Drug Target. 2020, 29, 454–465. [Google Scholar] [CrossRef]
- Lee, M.-H.; Liu, K.-H.; Thomas, J.L.; Chen, J.-R.; Lin, H.-Y. Immunotherapy of Hepatocellular Carcinoma with Magnetic PD-1 Peptide-Imprinted Polymer Nanocomposite and Natural Killer Cells. Biomolecules 2019, 9, 651. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, N.; Suh, H.; Irvine, D.J. Nanoparticle anchoring targets immune agonists to tumors enabling anti-cancer immunity without systemic toxicity. Nat. Commun. 2018, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Au, K.M.; Park, S.I.; Wang, A.Z. Trispecific natural killer cell nanoengagers for targeted chemoimmunotherapy. Sci. Adv. 2020, 6, eaba8564. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; Chan, M.F.; Li, J.; King, M.R. Super natural killer cells that target metastases in the tumor draining lymph nodes. Biomaterials 2015, 77, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Atukorale, P.U.; Raghunathan, S.P.; Raguveer, V.; Moon, T.J.; Zheng, C.; Bielecki, P.A.; Wiese, M.L.; Goldberg, A.L.; Covarrubias, G.; Hoimes, C.J.; et al. Nanoparticle Encapsulation of Synergistic Immune Agonists Enables Systemic Codelivery to Tumor Sites and IFNβ-Driven Antitumor Immunity. Cancer Res. 2019, 79, 5394–5406. [Google Scholar] [CrossRef]
- Murphy, D.A.; Cheng, H.; Yang, T.; Yan, X.; Adjei, I.M. Reversing Hypoxia with PLGA-Encapsulated Manganese Dioxide Nanoparticles Improves Natural Killer Cell Response to Tumor Spheroids. Mol. Pharm. 2021, 18, 2935–2946. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Sato, T.; Endo, R.; Sasaki, S.; Takahashi, N.; Sato, Y.; Hyodo, M.; Hayakawa, Y.; Harashima, H. STING agonist loaded lipid nanoparticles overcome anti-PD-1 resistance in melanoma lung metastasis via NK cell activation. J. Immunother. Cancer 2021, 9, e002852. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Li, T.; Tan, Y.; Xu, H. Selenium-containing nanoparticles synergistically enhance Pemetrexed&NK cell-based chemoimmunotherapy. Biomaterials 2021, 280, 121321. [Google Scholar] [CrossRef]
- Wei, Z.; Yi, Y.; Luo, Z.; Gong, X.; Jiang, Y.; Hou, D.; Zhang, L.; Liu, Z.; Wang, M.; Wang, J.; et al. Selenopeptide Nanomedicine Activates Natural Killer Cells for Enhanced Tumor Chemoimmunotherapy. Adv. Mater. 2022, 34, 2108167. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.; Sun, Z.; Li, S.; Peng, X.; Li, W.; Zhou, L.; Ma, Y.; Gong, P.; Cai, L. Cell-Membrane Immunotherapy Based on Natural Killer Cell Membrane Coated Nanoparticles for the Effective Inhibition of Primary and Abscopal Tumor Growth. ACS Nano 2018, 12, 12096–12108. [Google Scholar] [CrossRef]
- Kwak, M.; Erdag, G.; Leick, K.M.; Bekiranov, S.; Engelhard, V.H.; Slingluff, C.L. Associations of immune cell homing gene signatures and infiltrates of lymphocyte subsets in human melanomas: Discordance with CD163+ myeloid cell infiltrates. J. Transl. Med. 2021, 19, 371. [Google Scholar] [CrossRef]
- Zhang, S.-C.; Hu, Z.-Q.; Long, J.-H.; Zhu, G.-M.; Wang, Y.; Jia, Y.; Zhou, J.; Ouyang, Y.; Zeng, Z. Clinical Implications of Tumor-Infiltrating Immune Cells in Breast Cancer. J. Cancer 2019, 10, 6175–6184. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.; Xue, H.; Sun, Y.; Zhang, C.; Song, Y.; Qi, Y. The Role of Tumor Inflammatory Microenvironment in Lung Cancer. Front. Pharmacol. 2021, 12, 1168. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, X.; Wang, L.; Zhang, C. Interplay between inflammatory tumor microenvironment and cancer stem cells (Review). Oncol. Lett. 2018, 16, 679–686. [Google Scholar] [CrossRef]
- Gun, S.Y.; Lee, S.W.L.; Sieow, J.L.; Wong, S.C. Targeting immune cells for cancer therapy. Redox Biol. 2019, 25, 101174. [Google Scholar] [CrossRef]
- Melero, I.; Rouzaut, A.; Motz, G.T.; Coukos, G. T-Cell and NK-Cell Infiltration into Solid Tumors: A Key Limiting Factor for Efficacious Cancer Immunotherapy. Cancer Discov. 2014, 4, 522–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, G.H.; Lin, Y.Q.; Tian, L.; Zhang, T.; Yan, D.M.; Yu, J.H.; Deng, Y.C. Natural killer cell homing and trafficking in tissues and tumors: From biology to application. Signal Transduct. Target. Ther. 2022, 7, 205. [Google Scholar] [CrossRef] [PubMed]
- Cifaldi, L.; Doria, M.; Cotugno, N.; Zicari, S.; Cancrini, C.; Palma, P.; Rossi, P. DNAM-1 Activating Receptor and Its Ligands: How Do Viruses Affect the NK Cell-Mediated Immune Surveillance during the Various Phases of Infection? Int. J. Mol. Sci. 2019, 20, 3715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, W.; Gordon, A.C.; Cho, S.; Huang, X.; Harris, K.R.; Larson, A.C.; Kim, D.-H. Immunomodulatory Magnetic Microspheres for Augmenting Tumor-Specific Infiltration of Natural Killer (NK) Cells. ACS Appl. Mater. Interfaces 2017, 9, 13819–13824. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Zhang, F.; Wei, Z.; Li, X.; Zhao, H.; Lv, H.; Ge, R.; Ma, H.; Zhang, H.; Yang, B.; et al. Magnetic delivery of Fe3O4@polydopamine nanoparticle-loaded natural killer cells suggest a promising anticancer treatment. Biomater. Sci. 2018, 6, 2714–2725. [Google Scholar] [CrossRef]
- Shevtsov, M.; Stangl, S.; Nikolaev, B.; Yakovleva, L.; Marchenko, Y.; Tagaeva, R.; Sievert, W.; Pitkin, E.; Mazur, A.; Tolstoy, P.; et al. Granzyme B Functionalized Nanoparticles Targeting Membrane Hsp70-Positive Tumors for Multimodal Cancer Theranostics. Small 2019, 15, e1900205. [Google Scholar] [CrossRef]
- Gasparri, A.M.; Sacchi, A.; Basso, V.; Cortesi, F.; Freschi, M.; Rrapaj, E.; Bellone, M.; Casorati, G.; Dellabona, P.; Mondino, A.; et al. Boosting Interleukin-12 Antitumor Activity and Synergism with Immunotherapy by Targeted Delivery with isoDGR-Tagged Nanogold. Small 2019, 15, e1903462. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Guo, L.; Gu, X.; Zhang, B.; Hu, X.; Zhang, J.; Chen, J.; Wang, Y.; Chen, C.; Gao, B.; et al. Prevention of colorectal cancer liver metastasis by exploiting liver immunity via chitosan-TPP/nanoparticles formulated with IL-12. Biomaterials 2012, 33, 3909–3918. [Google Scholar] [CrossRef]
- Jiao, P.; Otto, M.; Geng, Q.; Li, C.; Li, F.; Butch, E.R.; Snyder, S.E.; Zhou, H.; Yan, B. Enhancing both CT imaging and natural killer cell-mediated cancer cell killing by a GD2-targeting nanoconstruct. J. Mater. Chem. B 2015, 4, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Monty, M.A.; Islam, A.; Nan, X.; Tan, J.; Tuhin, I.J.; Tang, X.; Miao, M.; Wu, D.; Yu, L. Emerging role of RNA interference in immune cells engineering and its therapeutic synergism in immunotherapy. Br. J. Pharmacol. 2021, 178, 1741–1755. [Google Scholar] [CrossRef]
- Adjei, I.M.; Jordan, J.; Tu, N.; Le Trinh, T.; Kandell, W.; Wei, S.; Sharma, B. Functional recovery of natural killer cell activity by nanoparticle-mediated delivery of transforming growth factor beta 2 small interfering RNA. J. Interdiscip. Nanomed. 2019, 4, 98–112. [Google Scholar] [CrossRef] [Green Version]
- Lian, S.; Xie, R.; Ye, Y.; Xie, X.; Li, S.; Lu, Y.; Li, B.; Cheng, Y.; Katanaev, V.; Jia, L. Simultaneous blocking of CD47 and PD-L1 increases innate and adaptive cancer immune responses and cytokine release. eBioMedicine 2019, 42, 281–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Luo, Y.-L.; Chen, Y.-F.; Lu, Z.-D.; Wang, Y.; Czarna, A.; Shen, S.; Xu, C.-F.; Wang, J. Dually regulating the proliferation and the immune microenvironment of melanoma via nanoparticle-delivered siRNA targeting onco-immunologic CD155. Biomater. Sci. 2020, 8, 6683–6694. [Google Scholar] [CrossRef] [PubMed]
- Neviani, P.; Wise, P.M.; Murtadha, M.; Liu, C.W.; Wu, C.-H.; Jong, A.Y.; Seeger, R.C.; Fabbri, M. Natural killer–derived exosomal miR-186 inhibits neuroblastoma growth and immune escape mechanisms. Cancer Res. 2019, 79, 1151–1164. [Google Scholar] [CrossRef]
- Islam, R.; Pupovac, A.; Evtimov, V.; Boyd, N.; Shu, R.; Boyd, R.; Trounson, A. Enhancing a Natural Killer: Modification of NK Cells for Cancer Immunotherapy. Cells 2021, 10, 1058. [Google Scholar] [CrossRef]
- Yan, C.; Jie, L.; Yongqi, W.; Weiming, X.; Juqun, X.; Yanbing, D.; Li, Q.; Xingyuan, P.; Mingchun, J.; Weijuan, G. Delivery of human NKG2D-IL-15 fusion gene by chitosan nanoparticles to enhance antitumor immunity. Biochem. Biophys. Res. Commun. 2015, 463, 336–343. [Google Scholar] [CrossRef]
- Zhuo, Y.; Chen, F.; Kong, L.; Li, T.; Lu, L.; Yang, J.; Yu, T.; Shi, X.; Li, K. Magnetic Resonance Imaging of the Human Ferritin Heavy Chain Reporter Gene Carried by Dendrimer-Entrapped Gold Nanoparticles. J. Biomed. Nanotechnol. 2019, 15, 518–530. [Google Scholar] [CrossRef]
- Meraz, I.M.; Majidi, M.; Cao, X.; Lin, H.; Li, L.; Wang, J.; Baladandayuthapani, V.; Rice, D.; Sepesi, B.; Ji, L.; et al. TUSC2 Immunogene Therapy Synergizes with Anti–PD-1 through Enhanced Proliferation and Infiltration of Natural Killer Cells in Syngeneic Kras-Mutant Mouse Lung Cancer Models. Cancer Immunol. Res. 2018, 6, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-S.; Han, J.-H.; Park, J.-H.; Kim, H.-K.; Choi, S.H.; Kim, G.R.; Song, H.; An, H.J.; Han, D.K.; Park, W.; et al. Multifunctional nanoparticles for genetic engineering and bioimaging of natural killer (NK) cell therapeutics. Biomaterials 2019, 221, 119418. [Google Scholar] [CrossRef]
- Spear, P.; Wu, M.R.; Sentman, M.L.; Sentman, C.L. NKG2D ligands as therapeutic targets. Cancer Immun. 2013, 13, 8. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3700746/ (accessed on 4 October 2022).
- Liu, C.; Lai, H.; Chen, T. Boosting Natural Killer Cell-Based Cancer Immunotherapy with Selenocystine/Transforming Growth Factor-Beta Inhibitor-Encapsulated Nanoemulsion. ACS Nano 2020, 14, 11067–11082. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Xu, Y.; Wu, Q.; Hu, P.; Shi, J. Mild Magnetic Hyperthermia-Activated Innate Immunity for Liver Cancer Therapy. J. Am. Chem. Soc. 2021, 143, 8116–8128. [Google Scholar] [CrossRef] [PubMed]
- Viaud, S.; Terme, M.; Flament, C.; Taieb, J.; Andre, F.; Novault, S.; Escudier, B.; Robert, C.; Caillat-Zucman, S.; Tursz, T.; et al. Dendritic Cell-Derived Exosomes Promote Natural Killer Cell Activation and Proliferation: A Role for NKG2D Ligands and IL-15Rα. PLoS ONE 2009, 4, e4942. [Google Scholar] [CrossRef] [PubMed]
- Shoae-Hassani, A.; Hamidieh, A.A.; Behfar, M.; Mohseni, R.; Mortazavi-Tabatabaei, S.A.; Asgharzadeh, S. NK Cell–derived Exosomes from NK Cells Previously Exposed to Neuroblastoma Cells Augment the Antitumor Activity of Cytokine-activated NK Cells. J. Immunother. 2017, 40, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Xu, M.; Luo, J.; Zhao, L.; Ye, G.; Shi, F.; Lv, C.; Chen, H.; Wang, Y.; Li, Y. Liver toxicity assessments in rats following sub-chronic oral exposure to copper nanoparticles. Environ. Sci. Eur. 2019, 31, 30. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Zang, Y.; Qu, J.; Tang, M.; Zhang, T. The Toxicity of Metallic Nanoparticles on Liver: The Subcellular Damages, Mechanisms, and Outcomes. Int. J. Nanomed. 2019, 14, 8787–8804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kermanizadeh, A.; Powell, L.G.; Stone, V. A review of hepatic nanotoxicology—Summation of recent findings and considerations for the next generation of study designs. J. Toxicol. Environ. Health Part B 2020, 23, 137–176. [Google Scholar] [CrossRef]
- Wang, P.; Lu, Y.-Q. Ferroptosis: A Critical Moderator in the Life Cycle of Immune Cells. Front. Immunol. 2022, 13, 877634. [Google Scholar] [CrossRef]
- Miller, J.S.; Soignier, Y.; Panoskaltsis-Mortari, A.; McNearney, S.A.; Yun, G.H.; Fautsch, S.K.; McKenna, D.; Le, C.; DeFor, T.E.; Burns, L.J.; et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood 2005, 105, 3051–3057. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.H.; Kim, J.; Lim, S.A.; Kim, J.; Kim, S.-J.; Lee, K.-M. NK Cell-Based Immunotherapies in Cancer. Immune Netw. 2020, 20, e14. [Google Scholar] [CrossRef]
- Liu, S.; Galat, V.; Galat, Y.; Lee, Y.K.A.; Wainwright, D.; Wu, J. NK cell-based cancer immunotherapy: From basic biology to clinical development. J. Hematol. Oncol. 2021, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, S. Pluripotent Stem Cell-Based Cell Therapy—Promise and Challenges. Cell Stem Cell 2020, 27, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Langenberg, K.; Kamphorst, J.; Bonhomme, C.; D’Amico, E.; Dublin, S.; Braam, S.; Reijerkerk, A. Controlled stirred tank bioreactors for large-scale manufacture of human iPSC models for cell therapy. Cytotherapy 2020, 22, S43. [Google Scholar] [CrossRef]
- Kang, T.; Huang, Y.; Zhu, Q.; Cheng, H.; Pei, Y.; Feng, J.; Xu, M.; Jiang, G.; Song, Q.; Jiang, T.; et al. Necroptotic cancer cells-mimicry nanovaccine boosts anti-tumor immunity with tailored immune-stimulatory modality. Biomaterials 2018, 164, 80–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Noh, Y.-W.; Kang, T.H.; Kim, J.-E.; Kim, S.; Um, S.H.; Oh, D.-B.; Park, Y.-M.; Lim, Y.T. Synthetic vaccine nanoparticles target to lymph node triggering enhanced innate and adaptive antitumor immunity. Biomaterials 2017, 130, 56–66. [Google Scholar] [CrossRef]
- Gang, M.; Wong, P.; Berrien-Elliott, M.M.; Fehniger, T.A. Memory-like natural killer cells for cancer immunotherapy. Semin. Hematol. 2020, 57, 185–193. [Google Scholar] [CrossRef]
- Fehniger, T.A.; Cooper, M.A. Harnessing NK Cell Memory for Cancer Immunotherapy. Trends Immunol. 2016, 37, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Giustarini, G.; Pavesi, A.; Adriani, G. Nanoparticle-Based Therapies for Turning Cold Tumors Hot: How to Treat an Immunosuppressive Tumor Microenvironment. Front. Bioeng. Biotechnol. 2021, 9, 458. [Google Scholar] [CrossRef]
- Hu, W.; Wang, G.; Huang, D.; Sui, M.; Xu, Y. Cancer Immunotherapy Based on Natural Killer Cells: Current Progress and New Opportunities. Front. Immunol. 2019, 10, 1205. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the phases of immunoediting, namely elimination, equilibrium, and escape.
Figure 1.
Schematic representation of the phases of immunoediting, namely elimination, equilibrium, and escape.

Figure 3.
Schematic representation of various strategies for NK cell-based nano-immunotherapies. Adapted from [89,90,91].

Figure 4.
Schematic representation of nanoparticle-assisted development of CAR-NK cell-based therapy.
Figure 4.
Schematic representation of nanoparticle-assisted development of CAR-NK cell-based therapy.

Table 1.
Comprehensive information on potential NK cell receptors for targeting NK cell-based immunotherapy.
Table 1.
Comprehensive information on potential NK cell receptors for targeting NK cell-based immunotherapy.
| Sr No. | Type of Receptors | Potential Targets | Ligands | Function | Ref. |
|---|---|---|---|---|---|
| 1. | Activating receptors | NKG2D |
| Activation, proliferation, and expansion of the immune cells | [71] |
| 2. | DNAM-1 |
| Enhances the cytotoxic activity | [72] | |
| 3. | NCR |
| Could stimulate and inhibit NK cell activity based on the ligand | [73] | |
| 4. | CD16 |
| Promotes cytokine production and enhances cytotoxicity | [74] | |
| 5. | NCAM/ CD56 | - | Enhances cytokine production, IFN- γ production and enhances NK cell proliferation | [75] | |
| 6. | CD96 |
| Enhances recognition of tumor cells, increases expression of cytotoxic granules | [76] | |
| 7. | CXCR1 |
| Enhances homing of immune cells | [77] | |
| 8. | CX3CR1 |
| Enhances homing of immune cells | [78] | |
| 9. | CD11 | - | Maturation biomarker for NK cells | [79] | |
| 10. | CCR7 |
| Facilitates homing of NK cells to the lymph nodes | [80] | |
| 11. | Inhibitory receptors | CD94/ NKG2A |
| Inhibits cytotoxic ability | [81] |
| 12. | TIM-3 |
| NK cell dysfunction and induces NK cell exhaustion | [82] | |
| 13. | TIGIT |
| Inhibits cytotoxic ability | [83] | |
| 14. | KIRs |
| Inhibits cytokine release | [84] | |
| 15. | LAG-3 |
| Inhibits immune cell activation and cytokine production | [85] | |
| 16. | PD-1 |
| Functional defects in NK cells, T cells | [86] |
Table 2.
Comprehensive information on nanoparticle-based immunomodulation for enhancing NK cell activity.
Table 2.
Comprehensive information on nanoparticle-based immunomodulation for enhancing NK cell activity.
| Sr No. | Nanoparticles | Target NK Cell | Ligand | Mechanism of Activation | Effect on Tumor Cells | Ref. |
|---|---|---|---|---|---|---|
| 1. | RGD peptide-tagged polyethylene glycol loaded with doxorubicin and diselenium nanoparticle | - | - |
|
| [103] |
| 2. | Lactobacillus brevis enriched selenium nanoparticle |
|
| [104] | ||
| 3. | Pemetrexed and cytosine-containing diselenide-loaded nanoparticles | - | - |
|
| [108] |
| 4. | Folate-containing liposomes, manganese protoporphyrin | Folate receptor | Folate |
|
| [109] |
| 5. | Calcium carbonate-coated gold nanostars with chlorine e6 photosensitizer | - | - |
|
| [110] |
| 6. | Lipid nanoparticles conjugated to CpG oligonucleotides, mannose, and H22 hepatoma lysate | - | - |
|
| [111] |
| 7. | PLGA nanoparticles encapsulated TLR7/8 agonist | TLR 7/8 receptor | TLR7/8 agonist |
|
| [112] |
| 8. | TGF-β and IL-2 loaded nanolipogels | TGF-β receptor | TGF-β |
| - | [101] |
| 9. | Camptothecin-loaded cyclodextrin-based polymers | - | - |
|
| [113] |
| 10. | Magnetic nanoparticles | - | - |
|
| [114] |
| 11. | Graphene oxide nanoclusters loaded with anti-CD16 | CD16 | Anti-CD16 antibody |
| - | [115] |
| 12. | CAR-NK cell loaded with paclitaxel | CD16, HER-2 receptor | Anti-HER-1 antibody, Anti-CD16 antibody |
|
| [116] |
| 13. | PDMAEMA-PTPN6 conjugated atezolizumab | PD-L1 | Atezolizumab |
|
| [117] |
| 14. | Magnetic nanoparticle conjugated with a peptide derived from PD-1 | - | - |
|
| [118] |
| 15. | Immunoliposomes loaded IL-2 and anti-CD137 | CD137 | Anti-CD137 antibody |
|
| [119] |
| 16. | Mesoporous ruthenium nanoparticles conjugated bispecific antibodies (SS-Fc, anti-CD16, and anti-CEA) | CD16 and CEA | Anti-CD16 and anti-CEA |
|
| [106] |
| 17. | Trispecific (antibodies of α-CD16, α-4-1BB, and α-EFGR) nanoengagers | CD16, EGFR | α-CD16, α-4-1BB, and α-EFGR |
|
| [120] |
| 18. | TRAIL and anti-NK1.1 antibody decorated liposomes | NK1.1 receptor, death receptor | An anti-NK1.1 antibody, TRAIL |
|
| [121] |
| 19. | cdGMP and MPLA encapsulated nanoparticle | - | - |
|
| [122] |
| 20. | Manganese dioxide nanoparticles | - | - |
|
| [123] |
| 21. | Lipid nanoparticle-loaded STING agonist | STING receptor | STING agonist |
|
| [124] |
| 22. | Selenium containing nanoparticle loaded pemetrexed | - | - |
|
| [125] |
| 23. | Selenopeptide nanomedicine | - | - |
|
| [126] |
| 24. | 4,4′,4′′,4′′′-(porphine 5,10,15,20-tetrayl) tetrakis (benzoic acid) (TCPP)-loaded nanoparticle | - | - |
|
| [127] |
Table 3.
Comprehensive information on nanoparticle assistance to enhance homing of NK cells.
| Sr No. | Nanoparticles | Target NK Cell | Ligand | Mechanism of Activation | Effect on Tumor Cells | Ref. |
|---|---|---|---|---|---|---|
| 1. | Iron oxide nanoparticles on the surface of primary NK cells | - | - |
|
| [89] |
| 2. | Iron oxide nanoparticles labelled on NK cells | - | - |
|
| [137] |
| 3. | Magnetic PLGA microspheres containing recombinant IFN-γ and iron oxide nanocubes |
|
| [136] | ||
| 4. | Dextran-coated serine protease granzyme-B functionalized superparamagnetic iron oxide nanoparticles | mHsp70 | Granzyme B |
|
| [138] |
| 5. | IL-12 bound gold nanoparticle tagged with homing peptide | αvβ3-integrin receptors | isoAsp-Gly-Arg homing peptide |
|
| [139] |
| 6. | IL-12-loaded chitosan nanoparticles | - | - |
|
| [140] |
| 7. | Anti-GD2 antibody tagged gold nanoparticle | GD-2 receptor | Anti-GD2 antibody |
|
| [141] |
Table 4.
Comprehensive information on nanoparticle-based post-translational gene silencing for activation of NK cells.
Table 4.
Comprehensive information on nanoparticle-based post-translational gene silencing for activation of NK cells.
| Sr No. | Nanoparticles | Target NK Cell | Ligand | Mechanism of Activation | Effect on Tumor Cells | Ref. |
|---|---|---|---|---|---|---|
| 1. | Manganese dioxide nanoparticle containing siRNA-TGFBR2 | TGFBR2 silencing | siRNA |
|
| [143] |
| 2. | EpCAM targeted cationic liposomes containing si-CD47 and si-PD-L1 | EpCAM, CD47, PD-L1 | si-CD47, si-PD-L1 |
|
| [144] |
| 3. | Cationic lipid-assisted nanoparticles encapsulated with siCD155 | CD155 | si-CD155 |
|
| [145] |
| 4. | miRNA-loaded NK cell-derived exosomes | - | let-7a |
|
| [95] |
| 5. | miRNA-186 loaded NK cell-derived exosomes | - | miRNA-186 |
|
| [146] |
Table 6.
Comprehensive information on nanoparticles activating NKG2D receptor.
| Sr No. | Nanoparticles | Target NK Cell | Ligand | Mechanism of Activation | Effect on Tumor Cells | Ref. |
|---|---|---|---|---|---|---|
| 1. | Super magnetic nanoparticle Zn-CoFe2O4@Zn-MnFe2O4 | NKG2D, VEGFR | UL16-binding protein |
|
| [154] |
| 2. | Dendritic cell-derived exosomes | NKG2D | NKG2D ligands |
|
| [155] |
| 3. | Glioblastoma cells pre-exposed NK cell-derived exosomes | NKG2D | - |
|
| [156] |
| 4. | TGF-β inhibitor and selenocysteine-containing nanoemulsion | NKG2D | - |
|
| [153] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Murugan, D.; Murugesan, V.; Panchapakesan, B.; Rangasamy, L. Nanoparticle Enhancement of Natural Killer (NK) Cell-Based Immunotherapy. Cancers 2022, 14, 5438. https://doi.org/10.3390/cancers14215438
AMA Style
Murugan D, Murugesan V, Panchapakesan B, Rangasamy L. Nanoparticle Enhancement of Natural Killer (NK) Cell-Based Immunotherapy. Cancers. 2022; 14(21):5438. https://doi.org/10.3390/cancers14215438
Chicago/Turabian StyleMurugan, Dhanashree, Vasanth Murugesan, Balaji Panchapakesan, and Loganathan Rangasamy. 2022. "Nanoparticle Enhancement of Natural Killer (NK) Cell-Based Immunotherapy" Cancers 14, no. 21: 5438. https://doi.org/10.3390/cancers14215438
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.