
FNTB Promoter Polymorphisms Are Independent Predictors of Survival in Patients with Triple Negative Breast Cancer

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Characteristics
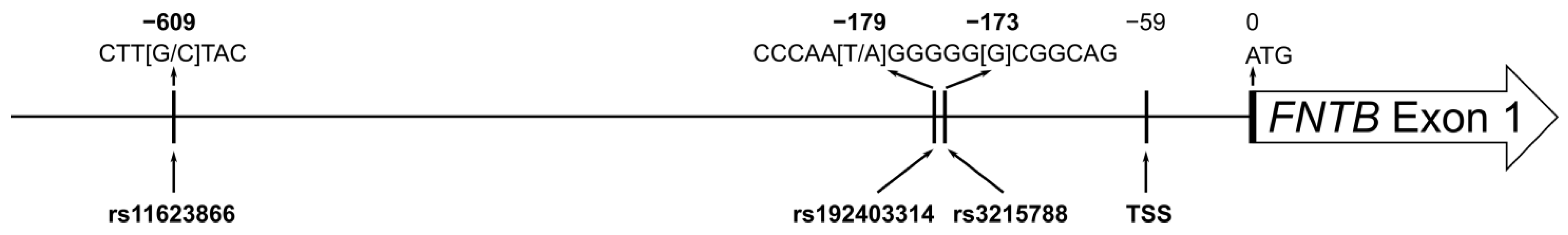
2.2. Polymorphism Retrieval and Bioinformatic Analyses
2.3. DNA Preparation and Determination of FNTB Genotypes
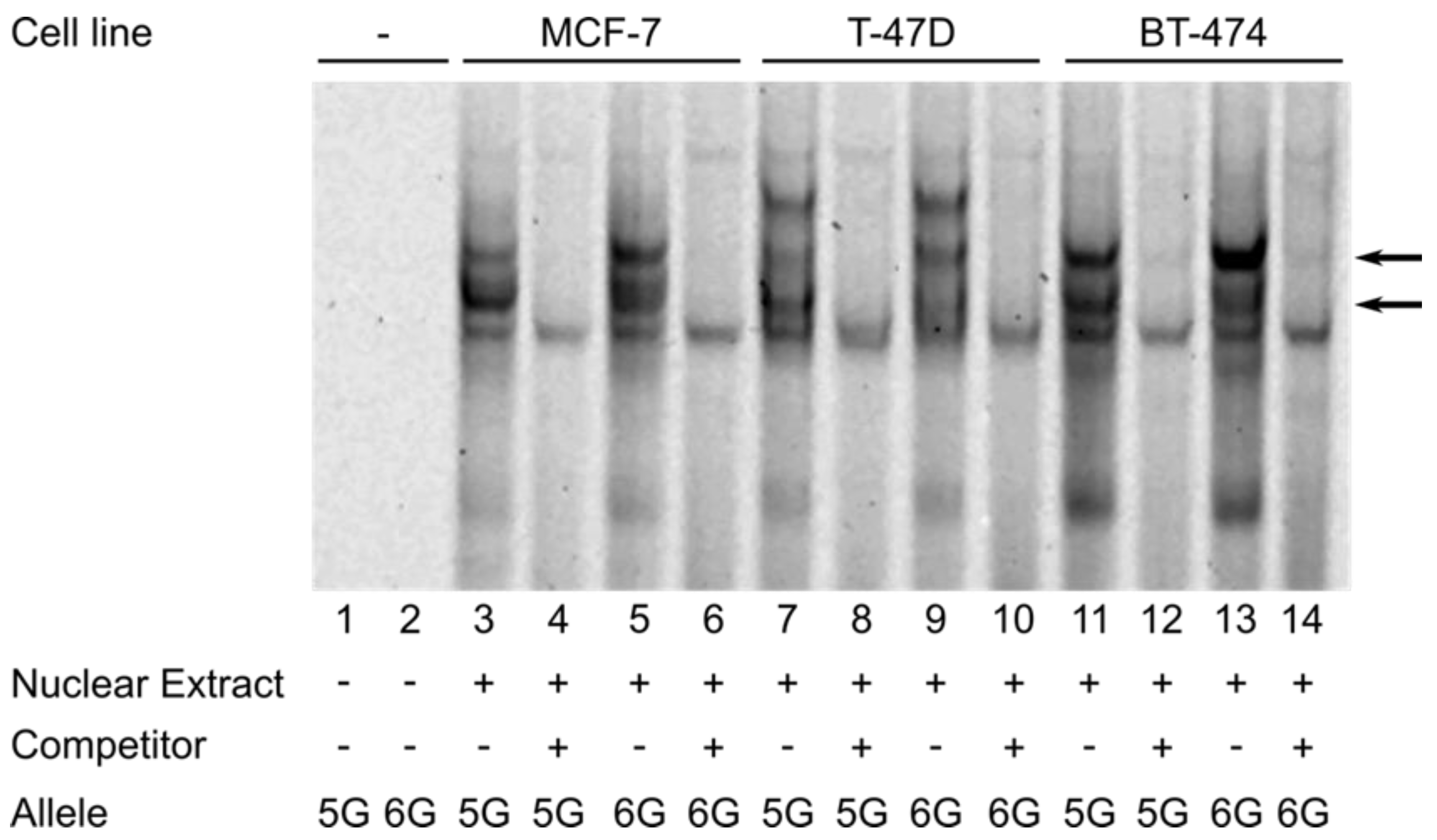
2.4. Electrophoretic Mobility Shift Assay
2.5. Statistical Analysis
3. Results
3.1. Association of FNTB Promoter Polymorphisms with Clinical and Pathological Parameters of Breast Cancer
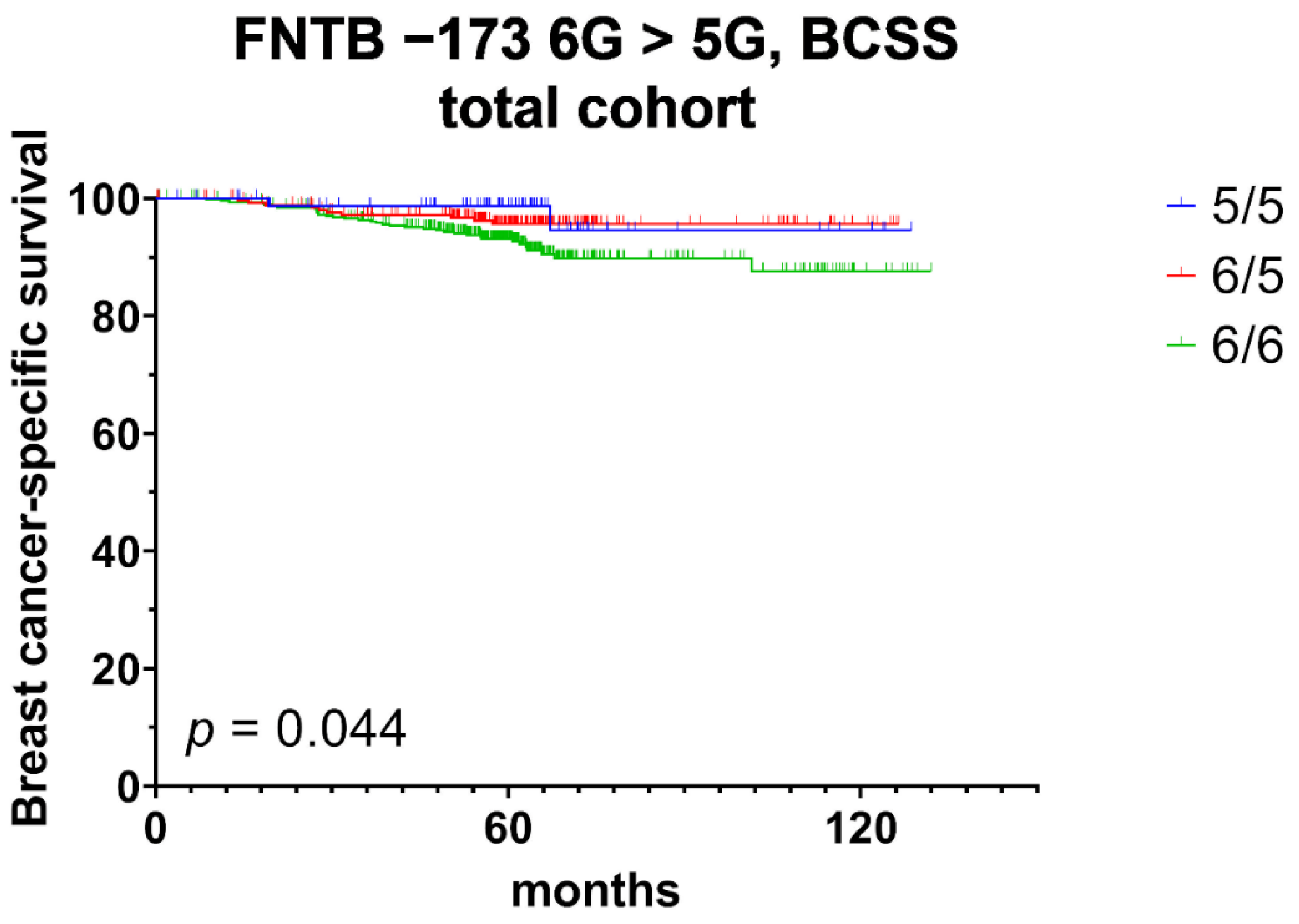
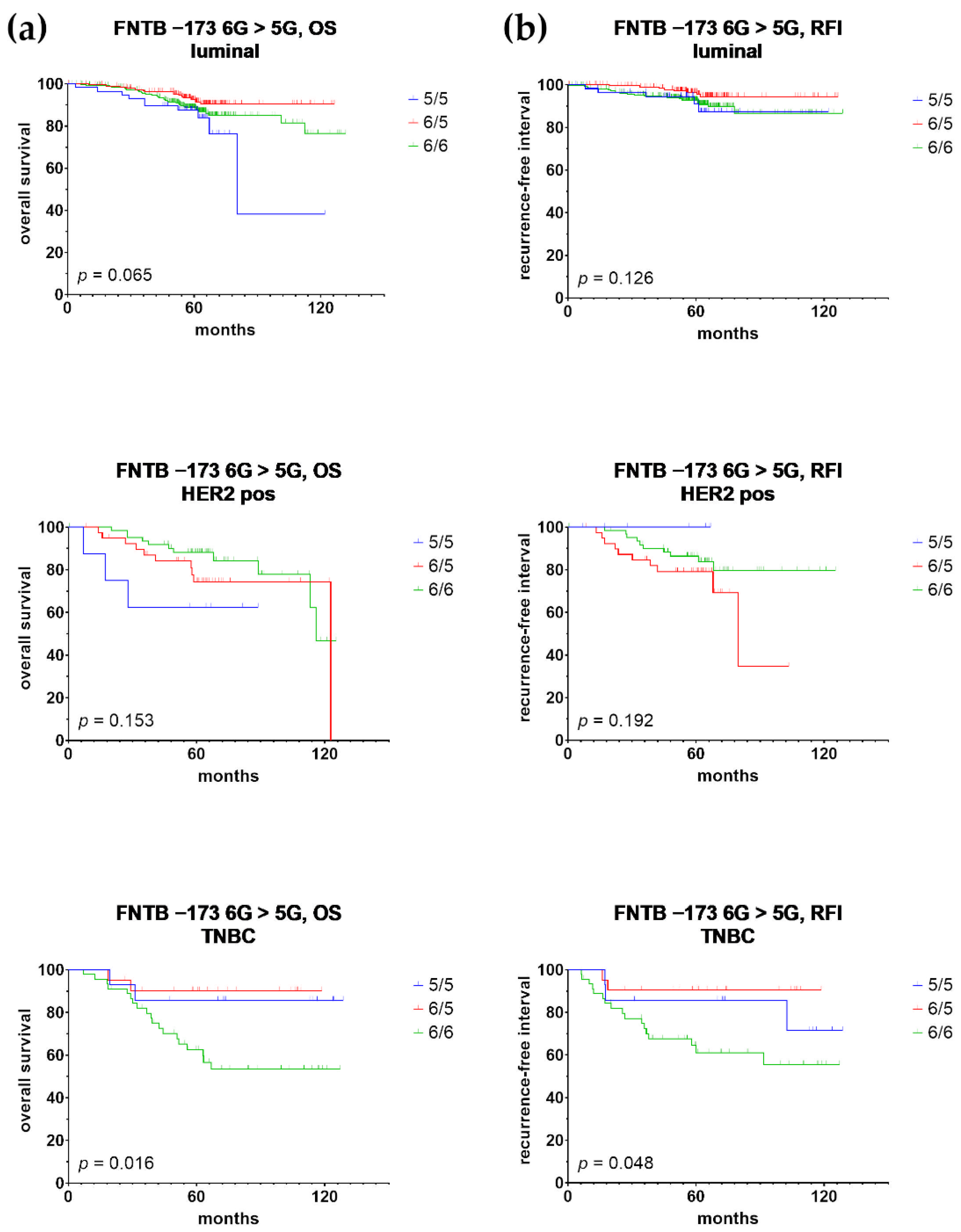
3.2. Univariate Prognostic Relevance of FNTB Promoter Polymorphisms and Their Association with Triple Negative Breast Cancer
3.3. Multivariate Prognostic Relevance of FNTB Promoter Polymorphisms and Their Association with Triple Negative Breast Cancer
3.4. The FNTB-173 6G > 5G Polymorphism Confers Genotype-Specific Binding of Breast Cancer Cell Line-Derived Nuclear Protein to the FNTB Promoter Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muller, V.; Fehm, T.; Janni, W.; Gebauer, G.; Solomayer, E.; Pantel, K. Clinical Relevance of Disseminated Tumor Cells in the Bone Marrow and Circulating Tumor Cells in the Blood of Breast Cancer Patients. Breast Care 2009, 4, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Riethdorf, S.; Pantel, K. Disseminated tumor cells in bone marrow and circulating tumor cells in blood of breast cancer patients: Current state of detection and characterization. Pathobiology 2008, 75, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Pantel, K.; Brakenhoff, R.H. Dissecting the metastatic cascade. Nat. Rev. Cancer 2004, 4, 448–456. [Google Scholar] [CrossRef]
- Early Breast Cancer Trialists’ Collaborative Group. Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: An overview of the randomised trials. Lancet 2005, 365, 1687–1717. [Google Scholar] [CrossRef]
- Bahrami, A.; Hassanian, S.M.; ShahidSales, S.; Farjami, Z.; Hasanzadeh, M.; Anvari, K.; Aledavood, A.; Maftouh, M.; Ferns, G.A.; Khazaei, M.; et al. Targeting RAS signaling pathway as a potential therapeutic target in the treatment of colorectal cancer. J. Cell. Physiol. 2018, 233, 2058–2066. [Google Scholar] [CrossRef]
- Sebti, S.M. Protein farnesylation: Implications for normal physiology, malignant transformation, and cancer therapy. Cancer Cell 2005, 7, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, K.T.; Beese, L.S. Thematic review series: Lipid posttranslational modifications. Structural biology of protein farnesyltransferase and geranylgeranyltransferase type I. J. Lipid Res. 2006, 47, 681–699. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Medarde, A.; Santos, E. Ras in cancer and developmental diseases. Genes Cancer 2011, 2, 344–358. [Google Scholar] [CrossRef] [Green Version]
- Clark, G.J.; Der, C.J. Aberrant function of the Ras signal transduction pathway in human breast cancer. Breast Cancer Res. Treat. 1995, 35, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.A.; Palmer, D.H.; Spooner, D.; Rea, D.W. Molecularly targeted therapeutics for breast cancer. BioDrugs 2007, 21, 215–224. [Google Scholar] [CrossRef]
- Izbicka, E.; Campos, D.; Carrizales, G.; Patnaik, A. Biomarkers of anticancer activity of R115777 (Tipifarnib, Zarnestra) in human breast cancer models in vitro. Anticancer Res. 2005, 25, 3215–3223. [Google Scholar] [PubMed]
- Warnberg, F.; White, D.; Anderson, E.; Knox, F.; Clarke, R.B.; Morris, J.; Bundred, N.J. Effect of a farnesyl transferase inhibitor (R115777) on ductal carcinoma in situ of the breast in a human xenograft model and on breast and ovarian cancer cell growth in vitro and in vivo. Breast Cancer Res. 2006, 8, R21. [Google Scholar] [CrossRef] [Green Version]
- Lubet, R.A.; Christov, K.; You, M.; Yao, R.; Steele, V.E.; End, D.W.; Juliana, M.M.; Grubbs, C.J. Effects of the farnesyl transferase inhibitor R115777 (Zarnestra) on mammary carcinogenesis: Prevention, therapy, and role of HaRas mutations. Mol. Cancer Ther. 2006, 5, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, S.R.; Semiglazov, V.F.; Manikhas, G.M.; Spaeth, D.; Romieu, G.; Dodwell, D.J.; Wardley, A.M.; Neven, P.; Bessems, A.; Park, Y.C.; et al. A phase II, randomized, blinded study of the farnesyltransferase inhibitor tipifarnib combined with letrozole in the treatment of advanced breast cancer after antiestrogen therapy. Breast Cancer Res. Treat. 2008, 110, 327–335. [Google Scholar] [CrossRef]
- Li, T.; Guo, M.; Gradishar, W.J.; Sparano, J.A.; Perez, E.A.; Wang, M.; Sledge, G.W. A phase II trial of capecitabine in combination with the farnesyltransferase inhibitor tipifarnib in patients with anthracycline-treated and taxane-resistant metastatic breast cancer: An Eastern Cooperative Oncology Group Study (E1103). Breast Cancer Res. Treat. 2012, 134, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Sparano, J.A.; Moulder, S.; Kazi, A.; Coppola, D.; Negassa, A.; Vahdat, L.; Li, T.; Pellegrino, C.; Fineberg, S.; Munster, P.; et al. Phase II trial of tipifarnib plus neoadjuvant doxorubicin-cyclophosphamide in patients with clinical stage IIB-IIIC breast cancer. Clin. Cancer Res. 2009, 15, 2942–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparano, J.A.; Moulder, S.; Kazi, A.; Vahdat, L.; Li, T.; Pellegrino, C.; Munster, P.; Malafa, M.; Lee, D.; Hoschander, S.; et al. Targeted inhibition of farnesyltransferase in locally advanced breast cancer: A phase I and II trial of tipifarnib plus dose-dense doxorubicin and cyclophosphamide. J. Clin. Oncol. 2006, 24, 3013–3018. [Google Scholar] [CrossRef]
- Raponi, M.; Lancet, J.E.; Fan, H.; Dossey, L.; Lee, G.; Gojo, I.; Feldman, E.J.; Gotlib, J.; Morris, L.E.; Greenberg, P.L.; et al. A 2-gene classifier for predicting response to the farnesyltransferase inhibitor tipifarnib in acute myeloid leukemia. Blood 2008, 111, 2589–2596. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, H.S.; Meier, W.; du Bois, A.; Kimmig, R.; Kuhlmann, J.D.; Siffert, W.; Sehouli, J.; Wollschlaeger, K.; Huober, J.; Hillemanns, P.; et al. The FNTB promoter polymorphism rs11623866 as a potential predictive biomarker for lonafarnib treatment of ovarian cancer patients. Br. J. Clin. Pharmacol. 2015, 80, 1139–1148. [Google Scholar] [CrossRef]
- Sobin, L.H.; Gospodarowicz, M.K.; Wittekind, C. TNM Classification of Malignant Tumours; John Wiley & Sons: New York, NY, USA, 2011. [Google Scholar]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thurlimann, B.; Senn, H.J.; Panel Members. Strategies for subtypes—Dealing with the diversity of breast cancer: Highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef]
- von Minckwitz, G.; Untch, M.; Blohmer, J.U.; Costa, S.D.; Eidtmann, H.; Fasching, P.A.; Gerber, B.; Eiermann, W.; Hilfrich, J.; Huober, J.; et al. Definition and impact of pathologic complete response on prognosis after neoadjuvant chemotherapy in various intrinsic breast cancer subtypes. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2012, 30, 1796–1804. [Google Scholar] [CrossRef] [Green Version]
- Quandt, K.; Frech, K.; Karas, H.; Wingender, E.; Werner, T. MatInd and MatInspector: New fast and versatile tools for detection of consensus matches in nucleotide sequence data. Nucleic Acids Res. 1995, 23, 4878–4884. [Google Scholar] [CrossRef]
- Grabe, N. AliBaba2: Context specific identification of transcription factor binding sites. Silico Biol. 2002, 2, S1–S15. [Google Scholar]
- Bachmann, H.S.; Siffert, W.; Frey, U.H. Successful amplification of extremely GC-rich promoter regions using a novel ‘slowdown PCR’ technique. Pharmacogenetics 2003, 13, 759–766. [Google Scholar] [CrossRef]
- Tolaney, S.M.; Garrett-Mayer, E.; White, J.; Blinder, V.S.; Foster, J.C.; Amiri-Kordestani, L.; Hwang, E.S.; Bliss, J.M.; Rakovitch, E.; Perlmutter, J.; et al. Updated Standardized Definitions for Efficacy End Points (STEEP) in Adjuvant Breast Cancer Clinical Trials: STEEP Version 2.0. J. Clin. Oncol. 2021, 39, 2720–2731. [Google Scholar] [CrossRef]
- Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhlmann, J.D.; Bachmann, H.S.; Link, T.; Wimberger, P.; Krober, E.; Thomssen, C.; Malle, B.; Bethmann, D.; Vetter, M.; Kantelhardt, E.J. Association of caspase 8 polymorphisms-652 6N InsDel and Asp302His with progression-free survival and tumor infiltrating lymphocytes in early breast cancer. Sci. Rep. 2019, 9, 12594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.S.; Kim, Y.S.; ZeRuth, G.; Beak, J.Y.; Gerrish, K.; Kilic, G.; Sosa-Pineda, B.; Jensen, J.; Pierreux, C.E.; Lemaigre, F.P.; et al. Transcription factor Glis3, a novel critical player in the regulation of pancreatic beta-cell development and insulin gene expression. Mol. Cell. Biol. 2009, 29, 6366–6379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, K.; Kumar, D.; Conway, A.E.; Park, K.; Jothi, R.; Jetten, A.M. GLIS3 Transcriptionally Activates WNT Genes to Promote Differentiation of Human Embryonic Stem Cells into Posterior Neural Progenitors. Stem Cells 2019, 37, 202–215. [Google Scholar] [CrossRef] [Green Version]
- Rami, F.; Baradaran, A.; Kahnamooi, M.M.; Salehi, M. Alteration of GLIS3 gene expression pattern in patients with breast cancer. Adv. Biomed. Res. 2016, 5, 44. [Google Scholar] [CrossRef]
- Najafabadi, H.S.; Mnaimneh, S.; Schmitges, F.W.; Garton, M.; Lam, K.N.; Yang, A.; Albu, M.; Weirauch, M.T.; Radovani, E.; Kim, P.M.; et al. C2H2 zinc finger proteins greatly expand the human regulatory lexicon. Nat. Biotechnol. 2015, 33, 555–562. [Google Scholar] [CrossRef]
- Won, K.A.; Spruck, C. Triplenegative breast cancer therapy: Current and future perspectives (Review). Int. J. Oncol. 2020, 57, 1245–1261. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Ikegami, Y.; Nakazawa, H.; Kuriyama, N.; Oki, M.; Hanai, J.; Sukhatme, V.P.; Kaneki, M. Low-Dose Farnesyltransferase Inhibitor Suppresses HIF-1alpha and Snail Expression in Triple-Negative Breast Cancer MDA-MB-231 Cells In Vitro. J. Cell. Physiol. 2017, 232, 192–201. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endpoint | Subgroup | Univariate Cox Regression | Multivariate Cox Regression | ||||||
|---|---|---|---|---|---|---|---|---|---|
| (Patients; Events) | p-Value | HR | 95% CI | p-Value | HR | 95% CI | |||
| RFI | total cohort (n = 797; 83) | 0.056 | 0.607 | 0.364 | 1.013 | 0.031 | 0.568 | 0.339 | 0.949 |
| luminal (n = 606; 41) | 0.056 | 0.465 | 0.212 | 1.021 | - | - | - | - | |
| HER2 positive (n = 111; 20) | 0.170 | 1.857 | 0.768 | 4.494 | 0.854 | 1.094 | 0.420 | 2.853 | |
| TNBC (n = 80; 17) | 0.039 | 0.214 | 0.049 | 0.925 | 0.039 | 0.214 | 0.049 | 0.925 | |
| OS | total cohort (n = 797; 114) | 0.048 | 0.641 | 0.412 | 0.997 | 0.040 | 0.629 | 0.403 | 0.980 |
| luminal (n = 606; 67) | 0.085 | 0.596 | 0.330 | 1.074 | - | - | - | - | |
| HER2 positive (n = 111; 24) | 0.288 | 1.594 | 0.674 | 3.770 | 0.958 | 0.976 | 0.390 | 2.442 | |
| TNBC (n = 80; 23) | 0.031 | 0.201 | 0.047 | 0.865 | 0.031 | 0.201 | 0.047 | 0.865 | |
| BCSS | total cohort (n = 797; 47) | 0.044 | 0.486 | 0.240 | 0.981 | 0.021 | 0.433 | 0.213 | 0.882 |
| luminal (n = 606; 19) | 0.078 | 0.328 | 0.095 | 1.134 | - | - | - | - | |
| HER2 positive (n = 111; 11) | 0.543 | 1.447 | 0.440 | 4.751 | 0.643 | 0.722 | 0.182 | 2.868 | |
| TNBC (n = 80; 17) | 0.089 | 0.276 | 0.063 | 1.217 | 0.089 | 0.276 | 0.063 | 1.217 | |
| Endpoint | Subgroup | Univariate Cox Regression | Multivariate Cox Regression | ||||||
|---|---|---|---|---|---|---|---|---|---|
| (Patients; Events) | p-Value | HR | 95% CI | p-Value | HR | 95% CI | |||
| RFI | total cohort (n = 797; 83) | 0.196 | 0.637 | 0.322 | 1.261 | 0.026 | 0.453 | 0.226 | 0.910 |
| luminal (n = 606; 41) | 0.355 | 0.648 | 0.259 | 1.625 | - | - | - | - | |
| HER2 positive (n = 111; 20) | 0.965 | 0.964 | 0.186 | 4.980 | 0.171 | 0.265 | 0.039 | 1.777 | |
| TNBC (n = 80; 17) | 0.099 | 0.333 | 0.090 | 1.230 | 0.099 | 0.333 | 0.090 | 1.230 | |
| OS | total cohort (n = 797; 114) | 0.645 | 0.883 | 0.520 | 1.498 | - | - | - | - |
| luminal (n = 606; 67) | 0.826 | 0.942 | 0.555 | 1.600 | - | - | - | - | |
| HER2 positive (n = 111; 24) | 0.156 | 2.219 | 0.738 | 6.670 | 0.694 | 0.769 | 0.207 | 2.853 | |
| TNBC (n = 80; 23) | 0.036 | 0.197 | 0.043 | 0.900 | 0.036 | 0.197 | 0.043 | 0.900 | |
| BCSS | total cohort (n = 797; 47) | 0.082 | 0.386 | 0.132 | 1.129 | 0.009 | 0.227 | 0.075 | 0.687 |
| luminal (n = 606; 19) | 0.278 | 0.431 | 0.094 | 1.973 | - | - | - | - | |
| HER2 positive (n = 111; 11) | 0.814 | 0.761 | 0.079 | 7.329 | 0.104 | 0.077 | 0.003 | 1.702 | |
| TNBC (n = 80; 17) | 0.070 | 0.144 | 0.018 | 1.172 | 0.070 | 0.144 | 0.018 | 1.172 | |
| Transcription Factor | Opt. | Strand | Matrix Similarity | Mat. Sim.—Opt. | Sequence | |
|---|---|---|---|---|---|---|
| Major allele only (6G) | BSAP/PAX5 | 0.87 | (+) | 0.869 | −0.001 | cctctgcccAATGgggggcggcagcatct |
| ZIC1 | 0.76 | (+) | 0.742 | −0.018 | ctgcccaatggGGGGcggc | |
| GLIS3 | 0.88 | (−) | 0.885 | 0.005 | gccgCCCCccattgggc | |
| KLF15 | 0.91 | (+) | 0.876 | −0.034 | cccaatggGGGGcggcagc | |
| PATZ1 | 0.89 | (+) | 0.850 | −0.040 | caatggggGGCGgcagcatctcaccagacca | |
| ZBTB14 | 0.89 | (+) | 0.857 | −0.033 | aatggggGGCGgcagca | |
| Minor allele only (5G) | ZNF658 | 0.75 | (−) | 0.702 | −0.048 | cgCCCCcattgggcaga |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachmann, H.S.; Jung, D.; Link, T.; Arnold, A.; Kantelhardt, E.; Thomssen, C.; Wimberger, P.; Vetter, M.; Kuhlmann, J.D. FNTB Promoter Polymorphisms Are Independent Predictors of Survival in Patients with Triple Negative Breast Cancer. Cancers 2022, 14, 468. https://doi.org/10.3390/cancers14030468
Bachmann HS, Jung D, Link T, Arnold A, Kantelhardt E, Thomssen C, Wimberger P, Vetter M, Kuhlmann JD. FNTB Promoter Polymorphisms Are Independent Predictors of Survival in Patients with Triple Negative Breast Cancer. Cancers. 2022; 14(3):468. https://doi.org/10.3390/cancers14030468
Chicago/Turabian StyleBachmann, Hagen Sjard, Dominik Jung, Theresa Link, Anna Arnold, Eva Kantelhardt, Christoph Thomssen, Pauline Wimberger, Martina Vetter, and Jan Dominik Kuhlmann. 2022. "FNTB Promoter Polymorphisms Are Independent Predictors of Survival in Patients with Triple Negative Breast Cancer" Cancers 14, no. 3: 468. https://doi.org/10.3390/cancers14030468