
Cancer Therapy Targeting CD47/SIRPα


1
Department of Internal Medicine, Yale School of Medicine, New Haven, CT 06510, USA
2
Department of Medical Oncology, Dana-Farber Cancer Institute, 450 Brookline Ave, Boston, MA 02215, USA
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(24), 6229; https://doi.org/10.3390/cancers13246229
Submission received: 25 November 2021
/
Accepted: 2 December 2021
/
Published: 11 December 2021
(This article belongs to the Special Issue Targeting Innate Immunity to Treat Cancer)
Abstract
:Simple Summary
The interaction between cluster of differentiation 47 (CD47) on cancer cells and signal regulatory protein alpha (SIRPα) on immune cells, such as macrophages and dendritic cells, generates a “don’t eat me” signal. This is a common mechanism that provides cancer cells an escape from the innate immune system. Several therapeutics directed to CD47 or SIRPα have entered early clinical trials in recent years. In this article, we review the role of CD47/SIRPα axis in cancer, and summarize the literature on the efficacy and safety of therapeutics targeting CD47 or SIRPα. We also discuss the future implementation of these therapeutics in the treatments of various cancer types.
Abstract
In the past decade, the field of cancer immunotherapy has rapidly advanced, establishing a crucial role for immune checkpoint blockers in the treatment of a variety of cancer types. In parallel with these remarkable clinical developments, further efforts have focused on ways of unleashing adaptive immune responses against cancer. CD47, a cell surface molecule overexpressed by several cancer types that facilitates immune escape from macrophages, dendritic cells and natural killer cells, and its ligand SIRPα, have emerged as potential therapeutic targets. A number of agents directed to CD47/SIRPα have been developed and demonstrated preclinical activity. Early phase clinical trials are investigating CD47/SIRPα directed agents with available data, suggesting safety and preliminary activity. Herein, we provide an overview of the mechanistic rationale of targeting CD47/SIRPα axis and associated clinical evidence.
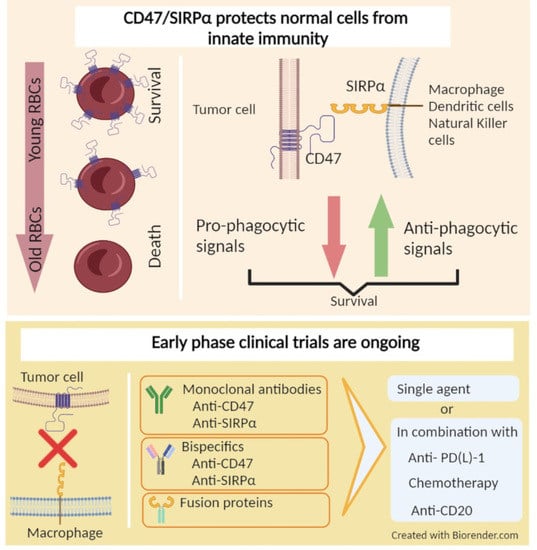
1. Introduction
Immunotherapy with immune checkpoint blockade, cellular therapy and an emerging range of novel approaches to target the immune system are rapidly changing cancer care. The earliest immunotherapies utilized cytokines such as interleukin-2 and interferon, paving the road for subsequent discovery and advancement [1,2]. The revelation that immune checkpoint inhibition targeting cytotoxic T-lymphocyte associated protein-4 (CTLA-4) or programmed cell death protein (PD-1) could prolong survival in a range of malignancies launched a revolution in the immunotherapy space [3,4]. Numerous therapies have emerged and currently are in testing. These include small molecules, antibodies, modified viruses, and cellular therapies including tumor-infiltrating lymphocyte (TIL) therapy and chimeric antigen receptor (CAR) T cell therapies that have led to a profound advancement with substantial efficacy in hematologic malignancies and potential for activity in solid tumors [5,6,7]. Nevertheless, despite these remarkable advancements designed to expand the T cell-mediated immunity, the majority of cancer patients do not respond to or develop resistance to immunotherapy, highlighting the need for additional approaches to expand cancer immunotherapy [8].
Investigations have focused on identifying the processes by which evolving tumor cells overcome innate immunity. Among various pathophysiologic mechanisms fostering immune evasive tumor behavior, cluster of differentiation 47 (CD47), a transmembrane molecule commonly present on non-malignant hematologic cells, including red blood cells and thrombocytes, has emerged as a promising target. Several solid tumor types and hematological malignancies overexpress CD47, enabling immune escape from components of the innate immune system such as macrophages, dendritic cells, and natural killer (NK) cells via binding signal regulatory protein alpha (SIRPα); these effects lead to the disruption of direct tumor killing, and the resultant impairment in antigen presentation and T cell infiltration [9]. Studies of several cancer types have suggested prognostic properties of CD47 overexpression. A number of mechanistically different approaches targeting the CD47/SIRPα axis in order to potentiate the innate immune system have been developed and advanced from preclinical studies to early clinical trials. These include monoclonal antibodies, fusion proteins and bispecific antibodies, along with combination strategies, with most demonstrating encouraging safety and preliminary clinical activity in both hematologic malignancies and solid tumors. Herein, we present a comprehensive review of the rationale behind targeting the CD47/SIRPα axis and CD47/SIRPα-directed therapeutics in clinical development.
2. Innate and Adaptive Immune Systems and Cancer
The majority of immunotherapies in use at this time target adaptive immunity by stimulating and activating T cells to recognize cancer cells [10]. T cell activation begins early in the lymph nodes, when antigen presenting cells (i.e., macrophages and dendritic cells) display tumor antigens in the major histocompatibility complex, which binds the T cell receptor. The activation of T cells via antigen presentation is a highly regulated process, controlled by numerous immune checkpoints requiring the involvement of various co-stimulatory and co-inhibitory molecular interactions. These positive and negative immune checkpoints determine whether the activation will occur [11,12,13]. By its nature, immune checkpoint inhibition works to overcome negative signals designed to protect “self” cells from immune attack. In addition to the currently approved molecular inhibitors of immune checkpoints, several other novel immunotherapy approaches are being investigated in preclinical and clinical settings, including CAR-T cells and TIL therapy, both of which are T cell products targeting specific proteins on the surface of malignant cells, aiming to augment adaptive immunity [7,14].
Innate immunity is the first line of defense against pathogens and other threats [10]. The innate immune system includes macrophages that phagocytose invaders, and are involved in antigen presentation. In cancer, increased infiltration of macrophages within and surrounding the tumor mass correlates with increased tumor invasiveness, growth, and poor prognosis. There is a correlation between tumor-associated macrophages (TAMs) and poor prognosis in breast, prostate, ovarian, and cervical cancers [15,16,17]. Clinical evidence of TAM participation in tumor growth is corroborated in animal models where macrophage signaling is inhibited or knocked out, suggesting that improving the response of the innate immune system to malignancy has potential to build on therapy options for cancer patients [18].
3. CD47/SIRPα Axis
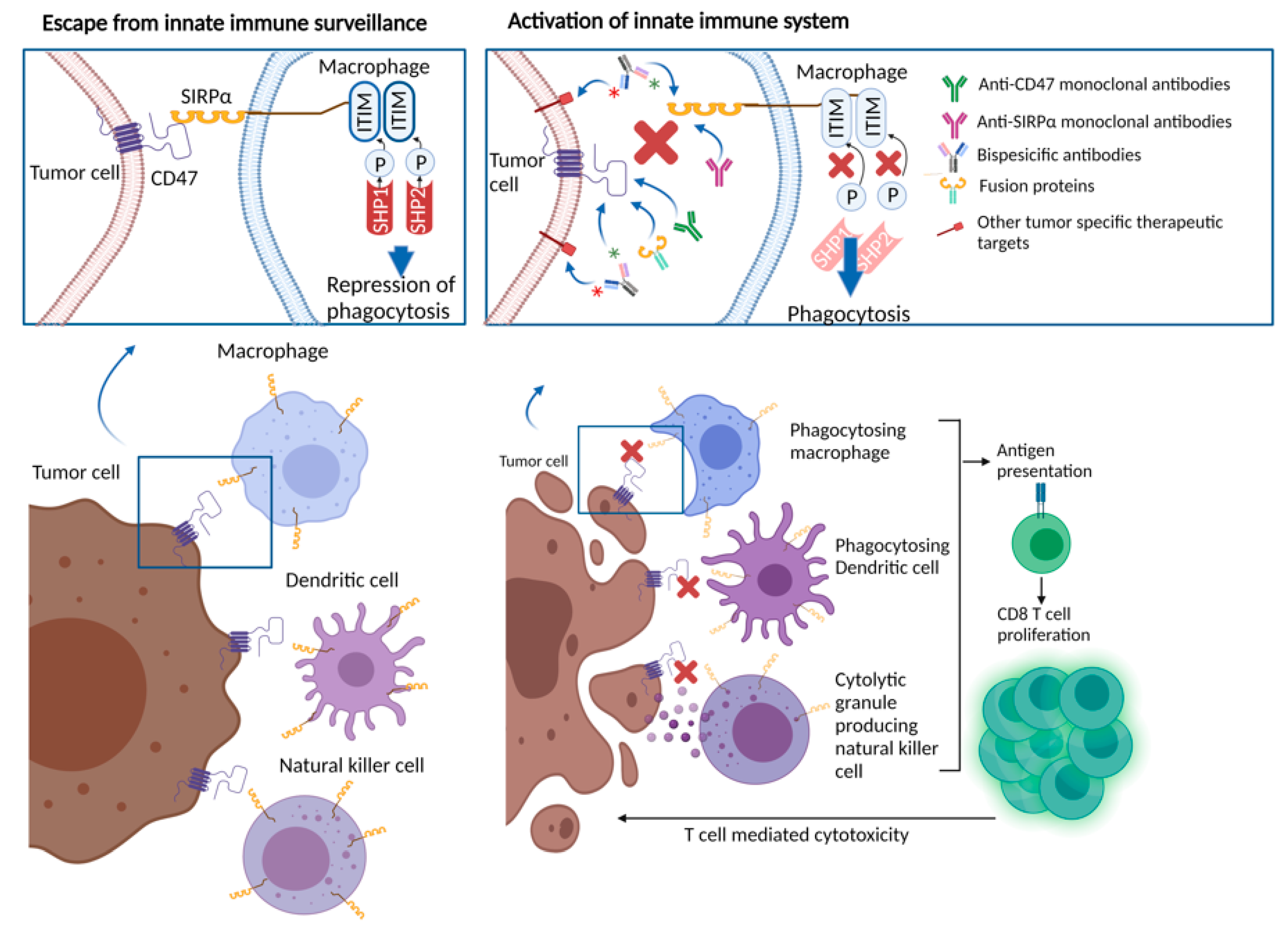
CD47, also known as integrin associated protein, is a transmembrane protein, belonging to the immunoglobulin superfamily broadly expressed on a variety of cell types [19]. CD47 has been identified as a marker on red blood cells where it serves as a “don’t eat me” signal to inhibit red blood cell phagocytosis [20]. As red blood cells age, they lose CD47 signaling, leading to the removal of older cells from circulation by macrophages in the spleen [20,21]. Notably, this overexpression is also seen on several other normal cells, including circulating hematopoietic stem cells and progenitor cells [9,19,20]. The ligands of CD47 include integrins, thrombospondin-1 (TSP-1), and SIRPα, a regulatory membrane glycoprotein expressed on various innate immune system cells, including macrophages and dendritic cells, along with granulocytes, monocytes and neurons [19,22]. SIRPα consists of an extracellular region with three Ig-like domains, and an intracellular region with two immunoreceptor tyrosine-based inhibitory motifs [23]. Upon activation via binding to CD47, the immunoreceptor tyrosine-based inhibitory motif regions become phosphorylated and induce the recruitment and activation of protein tyrosine phosphatases SHP-1 and SHP-2, leading to dephosphorylation of downstream molecules such as myosin IIA and repression of phagocytosis [24]. Overall, under physiologic conditions, CD47 and SIRPα interaction is one of the mechanisms protecting normal cells from macrophage-mediated phagocytosis [22].
4. Role of CD47/SIRPα in Cancer
In the early 1990s, the first oncological studies of CD47 identified it as a potential tumor marker for ovarian cancer [25]. This was followed by investigations on a wide variety of solid and hematological cancer types, including head and neck small-cell carcinoma (HNSCC), breast cancer, acute myeloid leukemia (AML), non-Hodgkin’s lymphoma (NHL), myeloma demonstrating differential overexpression of CD47 between cancer cells and matched normal cells [9,20,21,24,26,27,28,29].
The role of the CD47/SIRPα interaction in providing an escape mechanism for cancer cells from macrophage targeting has been well described. Human-derived xenograft models for several types of malignancies demonstrated sensitivity to CD47-blocking antibodies. In culture, these antibodies induced the macrophage-mediated phagocytosis of tumor cells [30,31,32,33,34,35,36,37]. The impact of the CD47 blockade on macrophage populations within the tumor microenvironment was also studied. In brief, TAMs display different polarization states between M1 macrophages with anti-tumor phenotypes and M2 macrophages with pro-tumor and immunosuppressive phenotypes [38,39,40]. In a human glioblastoma model, anti-CD47 therapy increased M1 macrophages within the tumor. This finding suggests that anti-CD47 therapy may play a role in shifting the phenotype of macrophages toward the anti-tumorigenic M1 subtype [41]. CD47 signaling also participates in macrophage recruitment into tumors. Weiskopf and colleagues showed that phagocytosis, following anti-CD47 treatment, causes systemic and local secretion of chemokines and cytokines that recruit macrophages into tumors in mice engrafted with small-cell lung cancer (SCLC) cell lines [30].
Beyond the activation of macrophage-mediated tumor killing, CD47-SIRPα interruption exerts other multidimensional positive effects on the immune response against cancer cells. For example, CD47-SIRPα blockade augments antibody dependent cellular cytotoxicity (ADCC) via the inhibition of SIRPα, expressed on the surface of NK cells [42,43]. Kim and colleagues demonstrated that impaired NK cell activity present in HNSCC cell lines overexpressing CD47 could be reversed with anti-CD47 antibodies [44]. CD47-SIRPα antagonist agents with an intact or even partially inactive Fc portion embedded in their structure may foster anti-tumor activity via antibody opsonization and destruction of target cells through ADCC or antibody-dependent cellular phagocytosis (ADCP) [45]. In addition, CD47/SIRPα interaction also has roles in tumor cell apoptosis, proliferation and migration [46,47,48]. CD47 inhibition can also negatively impact the function of other CD47 ligands, such as TSP-1 and integrins. These indirect effects may contribute to the anti-tumor and pro-inflammatory activity of CD47 inhibition. Despite contrasting evidence, a growing body of research highlights the role of TSP-1 in cell proliferation, invasion, metastatic potential, and worse survival rates, either through its interaction with CD47 or independently [49,50,51]. Notably, Kamijo and colleagues reported an association between high TSP-1 expression and worse disease-free survival in cutaneous T cell lymphoma patients. TSP-1 was found to be overexpressed in cutaneous T cell lymphoma, and anti-CD47 antibodies led to the inhibition of TSP-1-mediated cell proliferation in vivo [52].
Preclinical work has suggested a synergy between the cytotoxic agents and the CD47 inhibitors, especially when cytotoxic therapies were introduced prior to CD47-directed therapies. Neoantigens and nucleic acid remnants, produced from dying cancer cells and released into the tumor microenvironment after chemotherapy, may potentiate anti-CD47 activity [53]. In the context of hematologic malignancies, in vitro studies showed that azacytidine (a standard of care DNA hypomethylating agent used in the treatment of AML) and myelodysplastic syndrome and venetoclax (a B-cell lymphoma-2 inhibitor used in AML), induces the expression of other pro-phagocytic pathway components such as calreticulin and CD47 [54].
Perhaps more intriguingly, the macrophages involved in phagocytosis function as antigen-presenting cells, linking innate and adaptive immunity [29,53,55]. Thus, targeting the CD47-SIRPα axis, either through the CD47 or SIRPα blockade, may also promote antigen-presenting cell function, and stimulate T cell-mediated anti-cancer immunity (Figure 1) [56,57]. Studies in preclinical models with cancer types including chronic lymphocytic leukemia, colon cancer, melanoma, HNSCC, and glioblastoma, showed the induction of antitumor cytotoxic T cell populations, and reduced regulatory T cell populations in response to anti-CD47 treatment [26,55,58,59,60]. These observations were replicated in ex vivo studies. For example, Tao and colleagues assessed tumor samples from esophageal squamous cell cancer patients, showing an inverse relationship between CD8 T cell density and CD47 expression. In mice models with esophageal squamous cell cancer, treatment with anti-CD47 antibodies led to an increase in PD-1 and CTLA-4 expression. Treatment with the combination of CD47, PD-1 and CTLA-4 inhibitors yielded significantly improved survival in mice, compared with anti-CD47 monotherapy or PD-1 and CTLA-4 inhibitor combination, suggesting a rationale for combinatory therapeutic approaches to obtain synergistic effects [61].
Clinical implications of CD47 overexpression were also studied in various cancer types with the majority showing an inverse relationship between CD47 overexpression and clinical outcomes [62]. Chao et al. used flow cytometry and found that NHL cells had two-fold greater CD47 expression than normal germinal center and peripheral blood B cells. Grouping patient samples based on CD47 mRNA expression levels, investigators showed improved overall survival in patients with CD47 low tumors, especially diffuse large B cell lymphoma (DLBCL), B cell chronic lymphocytic leukemia, and mantle cell lymphoma subsets [63]. Majeti and colleagues, showed high CD47 expression by gene expression arrays and flow cytometry in leukemia stem cells, compared with normal counterparts in a group of 137 AML patients. Compared with those with low CD47 expression, patients with high CD47 expression had significantly worse overall survival rates (22.1 vs 9.1 months, hazard ratio (HR): 2.02) and event free survival (17.1 vs 6.8 months, HR 1.94) [32]. Analyzing immunohistochemistry staining of CD47 in bone marrow biopsy samples from 248 AML patients, Galli et al. detected high CD47 staining in one-fourth of the patient samples. Samples with high CD47 staining had higher median blast count, median bone marrow infiltration, and disease burden. Although there was a trend towards unfavorable progression free survival in patients with high CD47, no statistical difference was observed in median progression-free survival, or overall survival [64]. Melanoma patients with tumors bearing CD47 overexpression were found to have worse overall survival rates and higher rates of distant metastasis [65]. Similarly, head and neck cancer patients with tumors bearing robust CD47 immunohistochemistry staining had diminished overall survival, compared with those with low CD47 staining [26]. A study of ovarian cancer demonstrated that increased CD47 expression is associated with worse prognosis, increased migration and invasion, and the induction of epithelial-mesenchymal transition [66].
5. Therapies Targeting CD47/SIRPα in Cancer
As a result of the promising preclinical data regarding the anti-tumor activity of CD47/SIPRα blockade obtained from in vivo and in vitro studies, several molecules have been developed and are undergoing clinical testing. Functionally, therapeutics under investigation may be classified as (1) CD47 targeting agents, (2) SIRPα targeting agents and (3) bispecific targeting agents. Table 1 provides a comprehensive list of the ongoing clinical trials of the CD47/SIRPα targeting therapeutics at the time of this publication. Although most of those approaches are currently being tested in early-phase clinical trials to assess safety and tolerability, available data from a number of published studies has revealed promising activity and favorable tolerability. In addition to being tested on their own, trials of combinations with other anti-tumor agents are underway. Inspired by the fact that CD47/SIRPα signaling limits the efficacy of tumor-opsonizing antibodies, a number of clinical trials are evaluating agents targeting this axis in combination with agents such as rituximab, cetuximab and trastuzumab [30,63,67]. Histone deacetylase (HDAC) inhibitors have been shown to enhance checkpoint inhibitor therapy by decreasing immune suppressive cells and increasing tumor antigen presentation [68,69]. Given the possible enhancement of tumor immunity, combinations of HDAC inhibitors and CD47 targeting therapies are underway. Other strategies employ a combination of CD47 targeted therapies with immune checkpoint inhibitors and chemotherapies.
Major concerns regarding the use of CD47-targeted agents are driven by the ubiquitous expression of CD47, which leads to rapid drug elimination, “antigen sink” and hematologic toxicity, such as anemia and thrombocytopenia [70]. The impact on hematopoietic cells, particularly red blood cells, presents a substantial issue with CD47/SIRPα targeted drugs. Given that older red blood cells are more sensitive to phagocytosis, red blood cell destruction remains a limiting toxicity with these drugs, and may influence the age of patients that can be treated with these agents [71]. Notably, as opposed to red blood cells, other normal functioning cells are less vulnerable to macrophage-mediated immune destruction with anti-CD47 therapies, as further activation of prophagocytic signals and involvement of calreticulin are suggested to be necessary steps to generate immune-related adverse events [72,73]. Furthermore, the use of CD47 or SIPRα-directed antibodies structured with an intact Fc portion raises similar concerns, due to the widespread expression of Fc receptors on normal cells, and the risks for potentially causing immune destruction of “self” cells [45]. From the drug development standpoint, the efficacy of the CD47/SIRPα blockade can vary based upon drug delivery method and compartmentalization of the drug. In addition, tumor type and stage, tumor immune microenvironment and acquired drug resistance, constitute other potential determinants of the efficacy of CD47/SIRPα inhibitors [74].
6. Anti-CD47 Antibodies and CD47-Targeting Recombinant Proteins
CD47-directed monoclonal antibodies and fusion proteins with SIRPα immunoglobulin structure competitively bind CD47 and block the interaction between CD47 and SIRPα. This class of therapeutics constitute the majority of the available in-human data testing CD47/SIRPα inhibition in solid tumors and hematologic malignancies, although data remains limited.
Hu5F9-G4 (5F9, magrolimab) is a humanized antibody with an IgG4 Fc fragment [75]. In a preclinical setting, magrolimab demonstrated anti-tumor activity against AML in-vitro and in vivo. Furthermore, complete disease elimination was observed in human B lymphoblastoid cell-engrafted mice, after treatment with magrolimab in combination with rituximab [75]. Preclinical models testing magrolimab in solid tumors such as colon, liver, ovarian and breast cancers demonstrated promising anti-tumor activity [76]. Another study in which patient-derived NHL xenografted mice were treated with magrolimab/rituximab combination showed an 89% cure rate, defined as over 4 months of disease free survival following the discontinuation of therapy [63]. In-depth analyses suggested that rituximab plays a complementary role in further stimulation of innate immunity, via its active Fc effector function-inducing natural killer cell and macrophage-mediated cellular cytotoxicity. Accordingly, the data from a phase Ib study of 22 patients with relapsed or refractory NHL, 95% of whom were previously treated with rituximab, demonstrated encouraging outcomes with an objective response rate of 50%, and a complete response rate of 36%, with magrolimab and rituximab in combination [77]. Adverse events experienced by patients on trial included chills, anemia and headaches (41% each), all of which occurred only in the first weeks of the trial. There were no significant safety signals in the latter stages of the trial. A simultaneously conducted phase I study of single agent magrolimab in metastatic solid tumors demonstrated a similar safety profile, with transient treatment-related adverse events [78]. Of note, trends in anemia development and transfusion requirements with magrolimab were further examined using the data from the patient population in the phase I dose escalation part of these studies [79]. Patients on escalating doses of magrolimab experienced a median 1.0 g/dL decrease in hemoglobin levels, and subsequent doses were associated with a lesser degree of hemoglobin decline. Red blood cell transfusion yielded appropriate responses in hemoglobin concentration, supporting the evidence regarding the transient nature of anemia after magrolimab administration [79]. A number of clinical trials evaluating magrolimab, either as a single agent or in combination with cytotoxic therapies, targeted therapies or immune checkpoint inhibitors to treat hematologic neoplasms, are ongoing.
Other CD-47-targeting monoclonal antibodies that have entered clinical development include IBI188 (letaplimab), AK117 and SRF231. A phase I study of letaplimab in patients with advanced solid tumors and lymphomas was recently completed. Letaplimab demonstrated a favorable toxicity profile, with no dose-limiting toxicities. The majority of the treatment-related adverse events were grade 1–2. The rate of anemia was 15%, and only one patient developed grade 3 anemia. Notably, infusion related reactions were seen in 65% of the patient population, but all were grade 1–2 and manageable with a standard infusion-related reaction treatment algorithm [80]. AK117 monotherapy in patients with metastatic solid tumors demonstrated safety with no dose-limiting toxicities, no infusion-related reactions, or grade ≥ 3 treatment-related adverse events observed with up to 20 mg/kg dosing. Further dose escalation is underway with 30 mg/kg dosing [81]. For SRF231, further exploration in clinical trials was held by the pharmaceutical company.
SIRPα-Fc fusion proteins are produced via combining the CD47 binding domain of SIRPα and the Fc region of human IgG1 or IgG4. These comprise a mechanistically different group of novel therapeutics, that are targeting CD47. SIRPα-Fc fusion proteins undergoing phase I clinical trials were engineered using different Fc structures to balance benefit and toxicity. Therefore, the extent of the contribution from the Fc portion of the fusion proteins on Fc receptor engagement, ADCC, ADCP and overall anti-tumor activity, may vary across strategies [45]. A SIRPα-Fc fusion protein, TTI-621 which is engineered using the Fc region of IgG1, competes with endogenous SIRPα in immune cells [82]. This agent can trigger phagocytosis of tumor cells by subsets of isolated TAMs. In the phase I study of TTI-621, the maximum tolerated dose was safely determined in 18 patients, based on transient grade 4 thrombocytopenia; importantly, however, no clinically significant anemia events were observed [82]. TTI-621 was administered to 146 patients in the dose expansion phase as a single agent in patients with T cell non-Hodgkin lymphoma, and in combination with rituximab in relapsed or treatment refractory patients with B-cell NHL, or with nivolumab in patients with Hodgkin lymphoma [83]. Notable adverse events included infusion reactions and thrombocytopenia (in 20% of the population)—although none were dose-limiting. Encouraging objective response rates were observed with monotherapy in DLBCL (29%), T cell NHL (25%) and with TTI-621 and rituximab combination in DLBCL (21%). In patients with relapsed/refractory mycosis fungoides and Sézary Syndrome, the intratumoral injection of TTI-621 showed appreciable improvement in both injected and non-injected adjacent lesions in 89% of the patients, as well as a reduction in lesion severity in one of two patients on maintenance intralesional therapy, suggesting a potential for abscopal or systemic effect [84]. TTI-622 is a new SIRPα-Fc fusion protein variant structured using the Fc region of human IgG4. This structure was designed to augment prophagocytic function of CD47 inhibition, by enabling higher levels of medication due to lower Fc receptor interactions, and associated toxicities expected with IgG4 compared with IgG1. This agent is currently being tested in a phase I setting in NHL patients. Initial results from this study showed that TTI-622 was well tolerated with no dose-limiting toxicities and grade ≥3 thrombocytopenia or anemia [85]. Another engineered protein, CV1-hIgG4, demonstrated activity but resulted in notable toxicity in vivo [86].
ALX148 was constructed by combining the high affinity D1 domain of SIRPα with the inactive human IgG1 Fc domain to avoid toxicity [87,88]. Indeed, a phase I clinical trial of this agent as monotherapy confirmed its safety, with no dose-limiting hematologic or non-hematologic toxicities observed. Investigators proceeded with testing two 10 mg/kg and 15 mg/kg weekly doses of ALX148, in combination with rituximab in relapsed/recurrent NHL patients, yielding an objective response rate of 40% and 55%, respectively, and a favorable toxicity profile, with the only grade 3 toxicities being a decreased neutrophil count (6%) and anemia (3%) [89]. A more extensive investigation of this agent was recently completed in combination with trastuzumab, with or without chemotherapy in metastatic gastric/gastroesophageal junction cancers, and in combination with pembrolizumab, with or without chemotherapy in metastatic head and neck squamous cell carcinoma patients [90]. The safety profile was favorable, with low rates of cytopenias in all cohorts. Encouraging activity signals have emerged with ALX148, in combination with pembrolizumab in HNSCC patients, who were previously treated with ≥2 lines of therapy with an objective response rate of 40%, and the ALX148 and trastuzumab combination in gastric cancer patients who previously progressed on HER2-targeted therapies with an objective response rate of 21%. Accordingly, various combination strategies with ALX148 are being investigated in phase II settings, in patients with HNSCC and gastric/gastroesophageal junction cancer. This type of approach attempts to limit the toxicity on normal cells; however, the anti-tumor efficacy may be compromised by losing the ADCC and ADCP effect [88].
7. SIRPα Targeting Agents
Agents targeting SIRPα include anti-SIRPα antibodies and modified CD47 proteins targeting SIRPα. Given that SIRPα expression is limited to myeloid cells and neurons, this approach is attractive because it avoids broader binding and undesirable effects observed with agents targeting CD47 on normal cells. However, the use of SIRPα-directed therapies may be limited by binding to neurons expressing SIRPα [91,92]. In addition, there is concern about cross reactivity with other SIRP family members such as SIRPβ which is, as opposed to SIRPα, proinflammatory in nature and induces neutrophil migration and macrophage phagocytosis, and SIRPγ which has previously been shown to induce T cell migration [93,94,95,96].
Two anti-SIRPα monoclonal antibodies have recently entered early clinical trial testing: BI 765063 and GS-0189 (also known as FIS-189). Both agents were designed to bear high affinity to SIRPα without binding to SIRPγ [97,98,99]. The dose escalation part of the phase I study of BI 765063 monotherapy in metastatic tumors was recently published and showed an impressive safety profile with no dose-limiting toxicities. As expected, no anemia or thrombocytopenia were observed. The most common treatment-related adverse events were infusion-related reaction (46%) and fatigue (12%) [99]. The phase I study of BI 765063 is currently recruiting patients with advanced solid tumors for the dose expansion phase, where patients are randomized into either BI 765063 monotherapy or in combination with the PD-1 inhibitor. GS-0189 is being tested in the phase I setting as a single agent, or in combination with rituximab in relapsed or refractory NHL patients.
Several other SIRPα-directed agents are currently in preclinical development [100,101,102,103]. For example, ADU-1805, an anti-SIRPα monoclonal antibody, generated to block all known SIRPα alleles to excel anti-tumor activity across SIRPα variants, demonstrated in vitro and in vivo activity [100]. KWAR23 is an anti-SIRPα antibody, which has been demonstrated to synergize with rituximab in mouse models of lymphoma [36]. While the effect was limited when KWAR23 was used on its own, the combination augmented myeloid cell-dependent killing. There has also been some synergy observed with KWAR23 and cetuximab or panitumumab in colorectal adenocarcinoma cell lines [36]. A novel high affinity CD47 variant, velcro-CD47, was developed with augmentation of the existing contact interference via N-terminal peptide extension leading to enhanced SIRPα binding. The velcro-CD47 demonstrated anti-tumor activity and synergy with tumor-specific monoclonal antibodies (trastuzumab or cetuximab) in tumor cell lines [101].
8. Bispecific Agents Targeting CD47 and Another Molecule
Bispecific agents comprise a novel group of recombinant antibodies designed to target two different cell surface molecules simultaneously. In addition to CD47/SIRPα blockade, bispecific agents currently under investigation were designed to target either another cancer specific cell surface molecule concurrently expressed on the target tumor cells (i.e., CD19, CD20, PD-L1) or a cell surface molecule expressed by T cells (i.e., PD-1). While the first strategy aims to improve specificity of tumor targeting and prevent off-target effects, the latter promises synergistic activity with stimulation of both innate and adaptive immune systems [104]. The primary nuances that require attention in developing the dual targeting CD47/SIRPα axis-directed bispecific agents include the degree of the CD47 binding capacity and the presence of the Fc portion. The intact Fc portion fosters immune effector cell functions, and provides a longer half-life to the engineered antibody model; however, this moiety may be associated with unwanted side effects, due to the expression of Fc receptors on normal cells resulting in a similar phenomenon, as observed with CD47 [105].
HX009, a PD-1 and CD47 dual blocker (via IgG4-Fc of anti-PD-1 antibody and extracellular domain of SIRPα) was assessed in a phase I setting, and demonstrated to be safe by achieving a maximum tolerated dose, without dose-limiting toxicities or hematologic adverse events in patients with metastatic solid tumors. Clinical benefit signals were observed in this heavily pretreated patient population, with a median of three previous treatment lines. Of 18 patients with follow-up imaging studies available, three patients achieved partial response, and seven patients achieved stable disease. A phase II study of HX009 is currently recruiting metastatic solid cancer patients [106].
IBI322, an anti-CD47/PD-L1 bispecific monoclonal antibody, was designed with a monovalent CD47-binding domain and bivalent PD-L1-binding domain, to enable selective tumor targeting and spare red blood cells [107,108]. As a result, IBI322 exerts 14-fold lower affinity to red blood cells, compared with anti-CD47 monoclonal antibodies, while enabling equivalent affinity to PD-L1 as anti-PD-L1 agents [109]. This promising agent is being investigated in large early phase clinical trials, in patients with metastatic solid tumors and hematologic malignancies separately.
SL-172154 is a bifunctional fusion protein, developed via a mechanistically novel technology; it consists of human SIRPα and CD40L parts, that are connected via a human Fc. Whereas SIRPα inhibits CD47, CD40L binds and activates CD40, a costimulatory molecule present on B cells, macrophages and dendritic cells [110]. CD40/CD40L interaction partakes in both innate and adaptive anti-tumor immunity via stimulating pro-inflammatory cytokine production, including IL12 and TNF- α, the induction of ADCC, and T cell-mediated immunity and antibody production. It is also currently an active area of investigation for drug development in immune-oncology [111,112,113]. In vivo studies of SL-172154 have demonstrated superior anti-tumor activity with the fusion protein over either CD47 antagonist, CD40 agonist antibody as monotherapy or the combination of the two, suggesting a synergistic role [110]. Moreover, experiments combining SL-172154 and CTLA-4 or PD-1 blockage yielded enhanced the survival benefit in mice. Intravenous and intratumoral formulations of SL-172154 are being studied in phase I clinical trials in patients with ovarian cancer and HNSCC, respectively.
Several other dual blockers were generated with the goal of co-engaging two concurrently expressed tumor cell surface molecules. In light of the synergy observed with anti-CD47 and anti-CD20 combination in vivo, scientists generated bispecific antibodies (i.e., IMM0306 and CPO107) which target cells with CD47 and CD20 co-expression [63,105,114]. A similar strategy was employed for the development of TG-1801, but targeting CD19 instead of CD20. This design not only addresses the challenges raised by non-specific CD47 targeting and the antigen sink, observed with solely CD47 targeting agents, but also exploits the pro-phagocytic activity of engaged Fc receptors via CD19 and CD20 inhibition [105,114]. Promising response and survival outcomes were observed with IMM0306 in animal models with NHL. TG-1801 demonstrated pre-clinical efficacy in cell lines and mouse models of numerous B-cell malignancies. Co-administration with rituximab led to significant tumor-growth inhibition and regression [115].
9. Other Approaches Targeting CD47/SIRPα Axis
Several other mechanistically unique CD47/SIRPα-directed strategies are currently in the preclinical setting. A revolutionary advancement in immune-oncology research, CAR-T cell treatments were applied in the CD47 blockade arena, with studies showing tumor killing against lung, ovarian and pancreatic cell lines, as well as the blocking of pancreatic and ovarian cancer xenograft growth in vivo [116,117,118].
Novel drug delivery systems, such as nanomedicine and synthetic biology technologies, have entered preclinical development of anti-CD47 therapeutics as well. HuNb1-IgG4, a nanobody generated via fusion of heavy-chain IgG4 and anti-CD47 antibodies, offers promise with its small and stable molecular structure, high affinity to CD47 resulting in in vivo and in vitro activity against ovarian and lymphoma cancer, and lack of hematopoietic cytotoxicity [119]. Moreover, nanoparticles made up of multi-functionalized iron oxide nanoparticles that include the anti-CD47 antibody and gemcitabine have been developed for use in pancreatic cancer therapy. Chen et al. developed a bio-responsive fibrin gel solution, containing CD47-conjugated nanoparticles as a post-surgical tool to induce the local phagocytosis of cancer cells and modulate an innate and adaptive immune response [120]. Quorum-sensing bacteria that delivers single chain antibody fragments targeting CD47 has also been tested with increased anti-tumor immunity and reduced progression [121].
10. Conclusions
Immunotherapies to date have mainly targeted adaptive immunity. However, innate immunity plays an important role in controlling tumor development and growth. Macrophages have demonstrated a correlation between poor prognosis and the degree of anti-tumor immune activity. Modifying macrophages and other innate immune activity via the blockade of the CD47/SIRPα axis has emerged as an attractive approach in the field of hematology-oncology. Many mechanistically and functionally unique compounds have been developed and are currently undergoing clinical testing. Although only a small number of clinical trials have been published to date, available data suggests overall safety, along with considerable activity signals. Potential limitations of targeting the CD47/SIRPα axis includes the ability to target agents into the tumor and toxicity such as cytopenias, which may potentially be overcome by novel therapeutic strategies. Questions remain on the cancer-type specific efficacy of this approach, and the synergistic potential with chemotherapy, targeted therapy and immune checkpoint blockade. Promising early signals in combination with tumor-opsonizing antibodies suggest that this may be a way to expand the effectiveness of these drugs. In addition, there remains interest in the combined targeting of the innate and adaptive immune systems. Numerous ongoing clinical trials and further clinical investigations will guide the tumor type, optimal combination therapy and timing of the therapy targeting the CD47/SIRPα axis.
Author Contributions
Conceptualization, E.I.B.; resources, E.I.B.; data curation, E.I.B. and N.D.; writing—original draft preparation, E.I.B. and N.D.; writing—review and editing, E.I.B. and N.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Atkins, M.B.; Lotze, M.T.; Dutcher, J.P.; Fisher, R.I.; Weiss, G.; Margolin, K.; Abrams, J.; Sznol, M.; Parkinson, D.; Hawkins, M.; et al. High-Dose Recombinant Interleukin 2 Therapy for Patients with Metastatic Melanoma: Analysis of 270 Patients Treated between 1985 and 1993. J. Clin. Oncol. 1999, 17, 2105–2116. [Google Scholar] [CrossRef]
- Kirkwood, J.M.; Strawderman, M.H.; Ernstoff, M.S.; Smith, T.J.; Borden, E.C.; Blum, R.H. Interferon Alfa-2b Adjuvant Therapy of High-Risk Resected Cutaneous Melanoma: The Eastern Cooperative Oncology Group Trial EST 1684. J. Clin. Oncol. 1996, 14, 7–17. [Google Scholar] [CrossRef]
- Hodi, F.S.; Oble, D.A.; Drappatz, J.; Velazquez, E.F.; Ramaiya, N.; Ramakrishna, N.; Day, A.L.; Kruse, A.; Mac Rae, S.; Hoos, A.; et al. CTLA-4 Blockade with Ipilimumab Induces Significant Clinical Benefit in a Female with Melanoma Metastases to the CNS. Nat. Clin. Pract. Oncol. 2008, 5, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Sznol, M.; McDermott, D.F.; Kluger, H.M.; Carvajal, R.D.; Sharfman, W.H.; Brahmer, J.R.; Lawrence, D.P.; Atkins, M.B.; Powderly, J.D.; et al. Survival, Durable Tumor Remission, and Long-Term Safety in Patients with Advanced Melanoma Receiving Nivolumab. J. Clin. Oncol. 2014, 32, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Schuster, S.J.; Svoboda, J.; Chong, E.A.; Nasta, S.D.; Mato, A.R.; Anak, Ö.; Brogdon, J.L.; Pruteanu-Malinici, I.; Bhoj, V.; Landsburg, D.; et al. Chimeric Antigen Receptor T Cells in Refractory B-Cell Lymphomas. N. Engl. J. Med. 2017, 377, 2545–2554. [Google Scholar] [CrossRef]
- Sarnaik, A.A.; Hamid, O.; Khushalani, N.I.; Lewis, K.D.; Medina, T.; Kluger, H.M.; Thomas, S.S.; Domingo-Musibay, E.; Pavlick, A.C.; Whitman, E.D.; et al. Lifileucel, a Tumor-Infiltrating Lymphocyte Therapy, in Metastatic Melanoma. J. Clin. Oncol. 2021, 39, 2656–2666. [Google Scholar] [CrossRef]
- Hou, A.J.; Chen, L.C.; Chen, Y.Y. Navigating CAR-T Cells through the Solid-Tumour Microenvironment. Nat. Rev. Drug Discov. 2021, 20, 531–550. [Google Scholar] [CrossRef]
- Poschke, I.; Mougiakakos, D.; Kiessling, R. Camouflage and Sabotage: Tumor Escape from the Immune System. Cancer Immunol. Immunother. 2011, 60, 1161–1171. [Google Scholar] [CrossRef]
- Jaiswal, S.; Jamieson, C.H.M.; Pang, W.W.; Park, C.Y.; Chao, M.P.; Majeti, R.; Traver, D.; van Rooijen, N.; Weissman, I.L. CD47 Is Upregulated on Circulating Hematopoietic Stem Cells and Leukemia Cells to Avoid Phagocytosis. Cell 2009, 138, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Hiam-Galvez, K.J.; Allen, B.M.; Spitzer, M.H. Systemic Immunity in Cancer. Nat. Rev. Cancer 2021, 21, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Xu, L.; Yi, M.; Yu, S.; Wu, K.; Luo, S. Novel Immune Checkpoint Targets: Moving beyond PD-1 and CTLA-4. Mol. Cancer 2019, 18, 155. [Google Scholar] [CrossRef]
- Goldrath, A.W.; Bevan, M.J. Selecting and Maintaining a Diverse T-Cell Repertoire. Nature 1999, 402, 255–262. [Google Scholar] [CrossRef]
- Fife, B.T.; Bluestone, J.A. Control of Peripheral T-Cell Tolerance and Autoimmunity via the CTLA-4 and PD-1 Pathways. Immunol. Rev. 2008, 224, 166–182. [Google Scholar] [CrossRef] [PubMed]
- Morotti, M.; Albukhari, A.; Alsaadi, A.; Artibani, M.; Brenton, J.D.; Curbishley, S.M.; Dong, T.; Dustin, M.L.; Hu, Z.; McGranahan, N.; et al. Promises and Challenges of Adoptive T-Cell Therapies for Solid Tumours. Br. J. Cancer 2021, 124, 1759–1776. [Google Scholar] [CrossRef]
- Nowicki, A.; Szenajch, J.; Ostrowska, G.; Wojtowicz, A.; Wojtowicz, K.; Kruszewski, A.A.; Maruszynski, M.; Aukerman, S.L.; Wiktor-Jedrzejczak, W. Impaired Tumor Growth in Colony-Stimulating Factor 1 (CSF-1)-Deficient, Macrophage-Deficient Op/Op Mouse: Evidence for a Role of CSF-1-Dependent Macrophages in Formation of Tumor Stroma. Int. J. Cancer 1996, 65, 112–119. [Google Scholar] [CrossRef]
- Lewis, C.E.; Pollard, J.W. Distinct Role of Macrophages in Different Tumor Microenvironments. Cancer Res. 2006, 66, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Bingle, L.; Brown, N.J.; Lewis, C.E. The Role of Tumour-Associated Macrophages in Tumour Progression: Implications for New Anticancer Therapies. J. Pathol. 2002, 196, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.Y.; Nguyen, A.V.; Russell, R.G.; Pollard, J.W. Colony-Stimulating Factor 1 Promotes Progression of Mammary Tumors to Malignancy. J. Exp. Med. 2001, 193, 727–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, E.J.; Frazier, W.A. Integrin-Associated Protein (CD47) and Its Ligands. Trends Cell Biol. 2001, 11, 130–135. [Google Scholar] [CrossRef]
- Oldenborg, P.A.; Zheleznyak, A.; Fang, Y.F.; Lagenaur, C.F.; Gresham, H.D.; Lindberg, F.P. Role of CD47 as a Marker of Self on Red Blood Cells. Science 2000, 288, 2051–2054. [Google Scholar] [CrossRef] [PubMed]
- Burger, P.; Hilarius-Stokman, P.; de Korte, D.; van den Berg, T.K.; van Bruggen, R. CD47 Functions as a Molecular Switch for Erythrocyte Phagocytosis. Blood 2012, 119, 5512–5521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barclay, A.N.; Brown, M.H. The SIRP Family of Receptors and Immune Regulation. Nat. Rev. Immunol. 2006, 6, 457–464. [Google Scholar] [CrossRef]
- Kurihara, H.; Harita, Y.; Ichimura, K.; Hattori, S.; Sakai, T. SIRP-Alpha-CD47 System Functions as an Intercellular Signal in the Renal Glomerulus. Am. J. Physiol. Renal Physiol. 2010, 299, F517–F527. [Google Scholar] [CrossRef] [PubMed]
- Matlung, H.L.; Szilagyi, K.; Barclay, N.A.; van den Berg, T.K. The CD47-SIRPα Signaling Axis as an Innate Immune Checkpoint in Cancer. Immunol. Rev. 2017, 276, 145–164. [Google Scholar] [CrossRef]
- Mawby, W.J.; Holmes, C.H.; Anstee, D.J.; Spring, F.A.; Tanner, M.J.A. Isolation and Characterization of CD47 Glycoprotein: A Multispanning Membrane Protein Which Is the Same as Integrin-Associated Protein (IAP) and the Ovarian Tumour Marker OA3. Biochem. J. 1994, 304, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Yu, G.-T.; Deng, W.-W.; Mao, L.; Yang, L.-L.; Ma, S.-R.; Bu, L.-L.; Kulkarni, A.B.; Zhang, W.-F.; Zhang, L.; et al. Anti-CD47 Treatment Enhances Anti-Tumor T-Cell Immunity and Improves Immunosuppressive Environment in Head and Neck Squamous Cell Carcinoma. OncoImmunology 2018, 7, e1397248. [Google Scholar] [CrossRef]
- Chao, M.P.; Tang, C.; Pachynski, R.K.; Chin, R.; Majeti, R.; Weissman, I.L. Extranodal Dissemination of Non-Hodgkin Lymphoma Requires CD47 and Is Inhibited by Anti-CD47 Antibody Therapy. Blood 2011, 118, 4890–4901. [Google Scholar] [CrossRef]
- Willingham, S.B.; Volkmer, J.-P.; Gentles, A.J.; Sahoo, D.; Dalerba, P.; Mitra, S.S.; Wang, J.; Contreras-Trujillo, H.; Martin, R.; Cohen, J.D.; et al. The CD47-Signal Regulatory Protein Alpha (SIRPa) Interaction Is a Therapeutic Target for Human Solid Tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 6662–6667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molecular Pathways: Activating T Cells after Cancer Cell Phagocytosis from Blockade of CD47 “Don’t Eat Me” Signals|Clinical Cancer Research. Available online: https://clincancerres.aacrjournals.org/content/21/16/3597 (accessed on 24 August 2021).
- Weiskopf, K.; Jahchan, N.S.; Schnorr, P.J.; Cristea, S.; Ring, A.M.; Maute, R.L.; Volkmer, A.K.; Volkmer, J.-P.; Liu, J.; Lim, J.S.; et al. CD47-Blocking Immunotherapies Stimulate Macrophage-Mediated Destruction of Small-Cell Lung Cancer. J. Clin. Investig. 2016, 126, 2610–2620. [Google Scholar] [CrossRef]
- Edris, B.; Weiskopf, K.; Volkmer, A.K.; Volkmer, J.-P.; Willingham, S.B.; Contreras-Trujillo, H.; Liu, J.; Majeti, R.; West, R.B.; Fletcher, J.A.; et al. Antibody Therapy Targeting the CD47 Protein Is Effective in a Model of Aggressive Metastatic Leiomyosarcoma. Proc. Natl. Acad. Sci. USA 2012, 109, 6656–6661. [Google Scholar] [CrossRef] [Green Version]
- Majeti, R.; Chao, M.P.; Alizadeh, A.A.; Pang, W.W.; Jaiswal, S.; Gibbs, K.D.; van Rooijen, N.; Weissman, I.L. CD47 Is an Adverse Prognostic Factor and Therapeutic Antibody Target on Human Acute Myeloid Leukemia Stem Cells. Cell 2009, 138, 286–299. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wang, S.; Li, J.; Li, B. CD47/SIRPα Blocking Enhances CD19/CD3-Bispecific T Cell Engager Antibody-Mediated Lysis of B Cell Malignancies. Biochem. Biophys. Res. Commun. 2019, 509, 739–745. [Google Scholar] [CrossRef]
- Yoshida, K.; Tsujimoto, H.; Matsumura, K.; Kinoshita, M.; Takahata, R.; Matsumoto, Y.; Hiraki, S.; Ono, S.; Seki, S.; Yamamoto, J.; et al. CD47 Is an Adverse Prognostic Factor and a Therapeutic Target in Gastric Cancer. Cancer Med. 2015, 4, 1322–1333. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Chung, H.; Banan, B.; Manning, P.T.; Ott, K.C.; Lin, S.; Capoccia, B.J.; Subramanian, V.; Hiebsch, R.R.; Upadhya, G.A.; et al. Antibody Mediated Therapy Targeting CD47 Inhibits Tumor Progression of Hepatocellular Carcinoma. Cancer Lett. 2015, 360, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ring, N.G.; Herndler-Brandstetter, D.; Weiskopf, K.; Shan, L.; Volkmer, J.-P.; George, B.M.; Lietzenmayer, M.; McKenna, K.M.; Naik, T.J.; McCarty, A.; et al. Anti-SIRPα Antibody Immunotherapy Enhances Neutrophil and Macrophage Antitumor Activity. Proc. Natl. Acad. Sci. USA 2017, 114, E10578–E10585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaeteewoottacharn, K.; Kariya, R.; Pothipan, P.; Fujikawa, S.; Pairojkul, C.; Waraasawapati, S.; Kuwahara, K.; Wongkham, C.; Wongkham, S.; Okada, S. Attenuation of CD47-SIRPα Signal in Cholangiocarcinoma Potentiates Tumor-Associated Macrophage-Mediated Phagocytosis and Suppresses Intrahepatic Metastasis. Transl. Oncol. 2018, 12, 217–225. [Google Scholar] [CrossRef]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-Associated Macrophages (TAM) as Major Players of the Cancer-Related Inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S. Alternative Activation of Macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-Associated Macrophages as Treatment Targets in Oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Hutter, G.; Kahn, S.A.; Azad, T.D.; Gholamin, S.; Xu, C.Y.; Liu, J.; Achrol, A.S.; Richard, C.; Sommerkamp, P.; et al. Anti-CD47 Treatment Stimulates Phagocytosis of Glioblastoma by M1 and M2 Polarized Macrophages and Promotes M1 Polarized Macrophages In Vivo. PLoS ONE 2016, 11, e0153550. [Google Scholar] [CrossRef]
- Nath, P.R.; Pal-Nath, D.; Mandal, A.; Cam, M.C.; Schwartz, A.L.; Roberts, D.D. Natural Killer Cell Recruitment and Activation Are Regulated by CD47 Expression in the Tumor Microenvironment. Cancer Immunol. Res. 2019, 7, 1547–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deuse, T.; Hu, X.; Agbor-Enoh, S.; Jang, M.K.; Alawi, M.; Saygi, C.; Gravina, A.; Tediashvili, G.; Nguyen, V.Q.; Liu, Y.; et al. The SIRPα–CD47 Immune Checkpoint in NK Cells. J. Exp. Med. 2021, 218, e20200839. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, J.-C.; Lee, J.-J.; Kim, S.; Lee, S.G.; Park, S.-W.; Sung, M.W.; Heo, D.S. Association of CD47 with Natural Killer Cell-Mediated Cytotoxicity of Head-and-Neck Squamous Cell Carcinoma Lines. Tumor Biol. 2008, 29, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Veillette, A.; Chen, J. SIRPα–CD47 Immune Checkpoint Blockade in Anticancer Therapy. Trends Immunol. 2018, 39, 173–184. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Uno, S.; Kinoshita, Y.; Yoshimura, Y.; Iida, S.-I.; Wakahara, Y.; Tsuchiya, M.; Yamada-Okabe, H.; Fukushima, N. Apoptosis Inducing Bivalent Single-Chain Antibody Fragments against CD47 Showed Antitumor Potency for Multiple Myeloma. Leuk Res. 2005, 29, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Boukhari, A.; Alhosin, M.; Bronner, C.; Sagini, K.; Truchot, C.; Sick, E.; Schini-Kerth, V.B.; André, P.; Mély, Y.; Mousli, M.; et al. CD47 Activation-Induced UHRF1 over-Expression Is Associated with Silencing of Tumor Suppressor Gene P16INK4A in Glioblastoma Cells. Anticancer Res. 2015, 35, 149–157. [Google Scholar]
- Uluçkan, O.; Becker, S.N.; Deng, H.; Zou, W.; Prior, J.L.; Piwnica-Worms, D.; Frazier, W.A.; Weilbaecher, K.N. CD47 Regulates Bone Mass and Tumor Metastasis to Bone. Cancer Res. 2009, 69, 3196–3204. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Sun, L.; Yuan, X.; Qiu, H. Thrombospondin-1 Is a Multifaceted Player in Tumor Progression. Oncotarget 2017, 8, 84546–84558. [Google Scholar] [CrossRef] [Green Version]
- Byrne, G.J.; Hayden, K.E.; McDowell, G.; Lang, H.; Kirwan, C.C.; Tetlow, L.; Kumar, S.; Bundred, N.J. Angiogenic Characteristics of Circulating and Tumoural Thrombospondin-1 in Breast Cancer. Int. J. Oncol. 2007, 31, 1127–1132. [Google Scholar] [CrossRef] [Green Version]
- Borsotti, P.; Ghilardi, C.; Ostano, P.; Silini, A.; Dossi, R.; Pinessi, D.; Foglieni, C.; Scatolini, M.; Lacal, P.M.; Ferrari, R.; et al. Thrombospondin-1 Is Part of a Slug-Independent Motility and Metastatic Program in Cutaneous Melanoma, in Association with VEGFR-1 and FGF-2. Pigment. Cell Melanoma Res. 2015, 28, 73–81. [Google Scholar] [CrossRef]
- Kamijo, H.; Miyagaki, T.; Takahashi-Shishido, N.; Nakajima, R.; Oka, T.; Suga, H.; Sugaya, M.; Sato, S. Thrombospondin-1 Promotes Tumor Progression in Cutaneous T-Cell Lymphoma via CD47. Leukemia 2020, 34, 845–856. [Google Scholar] [CrossRef]
- Liu, X.; Pu, Y.; Cron, K.; Deng, L.; Kline, J.; Frazier, W.A.; Xu, H.; Peng, H.; Fu, Y.-X.; Xu, M.M. CD47 Blockade Triggers T Cell-Mediated Destruction of Immunogenic Tumors. Nat. Med. 2015, 21, 1209–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.; Harrabi, O.; Fong, A.P.; Ruffner, K.L.; Forgie, A.J.; Sim, J.; Randolph, S.S.; Wan, H.; Pons, J.; Kuo, T.C. ALX148 Enhances the Depth and Durability of Response to Multiple AML Therapies. Blood 2020, 136, 15–16. [Google Scholar] [CrossRef]
- Tseng, D.; Volkmer, J.-P.; Willingham, S.B.; Contreras-Trujillo, H.; Fathman, J.W.; Fernhoff, N.B.; Seita, J.; Inlay, M.A.; Weiskopf, K.; Miyanishi, M.; et al. Anti-CD47 Antibody-Mediated Phagocytosis of Cancer by Macrophages Primes an Effective Antitumor T-Cell Response. Proc. Natl. Acad. Sci. USA 2013, 110, 11103–11108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Shao, R.; Huang, H.; Wang, X.; Rong, Z.; Lin, Y. Engineering Macrophages to Phagocytose Cancer Cells by Blocking the CD47/SIRPɑ Axis. Cancer Med. 2019, 8, 4245–4253. [Google Scholar] [CrossRef] [Green Version]
- Gauttier, V.; Pengam, S.; Durand, J.; Biteau, K.; Mary, C.; Morello, A.; Néel, M.; Porto, G.; Teppaz, G.; Thepenier, V.; et al. Selective SIRPα Blockade Reverses Tumor T Cell Exclusion and Overcomes Cancer Immunotherapy Resistance. J. Clin. Investig. 2020, 130, 6109–6123. [Google Scholar] [CrossRef] [PubMed]
- Soto-Pantoja, D.R.; Terabe, M.; Ghosh, A.; Ridnour, L.A.; DeGraff, W.G.; Wink, D.A.; Berzofsky, J.A.; Roberts, D.D. CD47 in the Tumor Microenvironment Limits Cooperation between Antitumor T-Cell Immunity and Radiotherapy. Cancer Res. 2014, 74, 6771–6783. [Google Scholar] [CrossRef] [Green Version]
- Von Roemeling, C.A.; Wang, Y.; Qie, Y.; Yuan, H.; Zhao, H.; Liu, X.; Yang, Z.; Yang, M.; Deng, W.; Bruno, K.A.; et al. Therapeutic Modulation of Phagocytosis in Glioblastoma Can Activate Both Innate and Adaptive Antitumour Immunity. Nat. Commun. 2020, 11, 1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Torres, A.-C.; Quiney, C.; Attout, T.; Boullet, H.; Herbi, L.; Vela, L.; Barbier, S.; Chateau, D.; Chapiro, E.; Nguyen-Khac, F.; et al. CD47 Agonist Peptides Induce Programmed Cell Death in Refractory Chronic Lymphocytic Leukemia B Cells via PLCγ1 Activation: Evidence from Mice and Humans. PLoS Med. 2015, 12, e1001796. [Google Scholar] [CrossRef]
- Tao, H.; Qian, P.; Wang, F.; Yu, H.; Guo, Y. Targeting CD47 Enhances the Efficacy of Anti-PD-1 and CTLA-4 in an Esophageal Squamous Cell Cancer Preclinical Model. Oncol. Res. 2017, 25, 1579–1587. [Google Scholar] [CrossRef]
- Nagahara, M.; Mimori, K.; Kataoka, A.; Ishii, H.; Tanaka, F.; Nakagawa, T.; Sato, T.; Ono, S.; Sugihara, K.; Mori, M. Correlated Expression of CD47 and SIRPA in Bone Marrow and in Peripheral Blood Predicts Recurrence in Breast Cancer Patients. Clin. Cancer Res. 2010, 16, 4625–4635. [Google Scholar] [CrossRef] [Green Version]
- Chao, M.P.; Alizadeh, A.A.; Tang, C.; Myklebust, J.H.; Varghese, B.; Gill, S.; Jan, M.; Cha, A.C.; Chan, C.K.; Tan, B.T.; et al. Anti-CD47 Antibody Synergizes with Rituximab to Promote Phagocytosis and Eradicate Non-Hodgkin Lymphoma. Cell 2010, 142, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Galli, S.; Zlobec, I.; Schürch, C.; Perren, A.; Ochsenbein, A.F.; Banz, Y. CD47 Protein Expression in Acute Myeloid Leukemia: A Tissue Microarray-Based Analysis. Leuk Res. 2015, 39, 749–756. [Google Scholar] [CrossRef]
- Fu, W.; Li, J.; Zhang, W.; Li, P. High Expression of CD47 Predicts Adverse Prognosis in Chinese Patients and Suppresses Immune Response in Melanoma. Biomed. Pharmacother. 2017, 93, 1190–1196. [Google Scholar] [CrossRef]
- Overexpression of CD47 Predicts Poor Prognosis and Promotes Cancer Cell Invasion in High-Grade Serous Ovarian Carcinoma. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5489890/ (accessed on 24 August 2021).
- Upton, R.; Banuelos, A.; Feng, D.; Biswas, T.; Kao, K.; McKenna, K.; Willingham, S.; Ho, P.Y.; Rosental, B.; Tal, M.C.; et al. Combining CD47 Blockade with Trastuzumab Eliminates HER2-Positive Breast Cancer Cells and Overcomes Trastuzumab Tolerance. Proc. Natl. Acad. Sci. USA 2021, 118, e2026849118. [Google Scholar] [CrossRef]
- Briere, D.; Sudhakar, N.; Woods, D.M.; Hallin, J.; Engstrom, L.D.; Aranda, R.; Chiang, H.; Sodré, A.L.; Olson, P.; Weber, J.S.; et al. The Class I/IV HDAC Inhibitor Mocetinostat Increases Tumor Antigen Presentation, Decreases Immune Suppressive Cell Types and Augments Checkpoint Inhibitor Therapy. Cancer Immunol. Immunother. 2018, 67, 381–392. [Google Scholar] [CrossRef]
- Orillion, A.; Hashimoto, A.; Damayanti, N.; Shen, L.; Adelaiye-Ogala, R.; Arisa, S.; Chintala, S.; Ordentlich, P.; Kao, C.; Elzey, B.; et al. Entinostat Neutralizes Myeloid-Derived Suppressor Cells and Enhances the Antitumor Effect of PD-1 Inhibition in Murine Models of Lung and Renal Cell Carcinoma. Clin. Cancer Res. 2017, 23, 5187–5201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catani, L.; Sollazzo, D.; Ricci, F.; Polverelli, N.; Palandri, F.; Baccarani, M.; Vianelli, N.; Lemoli, R.M. The CD47 Pathway Is Deregulated in Human Immune Thrombocytopenia. Exp. Hematol. 2011, 39, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, S.; van Rooijen, N.; Saxena, R.K. Reduced Expression of CD47 during Murine Red Blood Cell (RBC) Senescence and Its Role in RBC Clearance from the Circulation. Transfusion 2007, 47, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, C.H.; Chao, M.P.; Gibbs, C.; McCamish, M.A.; Liu, J.; Chen, J.Y.; Majeti, R.; Weissman, I.L. The Macrophage ‘Do Not Eat Me’ Signal, CD47, Is a Clinically Validated Cancer Immunotherapy Target. Ann. Oncol. 2019, 30, 486–489. [Google Scholar] [CrossRef]
- Chao, M.P.; Majeti, R.; Weissman, I.L. Programmed Cell Removal: A New Obstacle in the Road to Developing Cancer. Nat. Rev. Cancer 2012, 12, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, Q.; Xiao, W.; Zhao, Y.; Pi, J.; Xu, H.; Zhao, H.; Xu, J.; Evans, C.E.; Jin, H. Advances in Anti-Tumor Treatments Targeting the CD47/SIRPα Axis. Front. Immunol. 2020, 11, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, L.; Zhao, F.; Tseng, S.; Narayanan, C.; Shura, L.; Willingham, S.; Howard, M.; Prohaska, S.; Volkmer, J.; et al. Pre-Clinical Development of a Humanized Anti-CD47 Antibody with Anti-Cancer Therapeutic Potential. PLoS ONE 2015, 10, e0137345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gholamin, S.; Mitra, S.S.; Feroze, A.H.; Liu, J.; Kahn, S.A.; Zhang, M.; Esparza, R.; Richard, C.; Ramaswamy, V.; Remke, M.; et al. Disrupting the CD47-SIRPα Anti-Phagocytic Axis by a Humanized Anti-CD47 Antibody Is an Efficacious Treatment for Malignant Pediatric Brain Tumors. Sci. Transl. Med. 2017, 9, eaaf2968. [Google Scholar] [CrossRef] [Green Version]
- Advani, R.; Flinn, I.; Popplewell, L.; Forero, A.; Bartlett, N.L.; Ghosh, N.; Kline, J.; Roschewski, M.; LaCasce, A.; Collins, G.P.; et al. CD47 Blockade by Hu5F9-G4 and Rituximab in Non-Hodgkin’s Lymphoma. N. Engl. J. Med. 2018, 379, 1711–1721. [Google Scholar] [CrossRef]
- Sikic, B.I.; Lakhani, N.; Patnaik, A.; Shah, S.A.; Chandana, S.R.; Rasco, D.; Colevas, A.D.; O’Rourke, T.; Narayanan, S.; Papadopoulos, K.; et al. First-in-Human, First-in-Class Phase I Trial of the Anti-CD47 Antibody Hu5F9-G4 in Patients with Advanced Cancers. J. Clin. Oncol. 2019, 37, 946–953. [Google Scholar] [CrossRef]
- Brierley, C.K.; Staves, J.; Roberts, C.; Johnson, H.; Vyas, P.; Goodnough, L.T.; Murphy, M.F. The Effects of Monoclonal Anti-CD47 on RBCs, Compatibility Testing, and Transfusion Requirements in Refractory Acute Myeloid Leukemia. Transfusion 2019, 59, 2248–2254. [Google Scholar] [CrossRef] [PubMed]
- Lakhani, N.; Orloff, M.; Fu, S.; Liu, Y.; Wang, Y.; Zhou, H.; Lin, K.; Liu, F.; Yan, S.; Patnaik, A. 295 First-in-Human Phase I Trial of IBI188, an Anti-CD47 Targeting Monoclonal Antibody, in Patients with Advanced Solid Tumors and Lymphomas. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef]
- Gan, H.K.; Coward, J.; Mislang, A.R.A.; Cosman, R.; Nagrial, A.; Jin, X.; Li, B.; Wang, Z.M.; Kwek, K.Y.; Xia, D.; et al. Safety of AK117, an Anti-CD47 Monoclonal Antibody, in Patients with Advanced or Metastatic Solid Tumors in a Phase I Study. J. Clin. Oncol. 2021, 39, 2630. [Google Scholar] [CrossRef]
- Ansell, S.M.; Flinn, I.W.; Maris, M.B.; O’Connor, O.A.; Lesokhin, A.; Advani, A.S.; Minden, M.D.; Percival, M.B.M.; Johnson, L.D.; Catalano, T.; et al. TTI-621 (SIRPαFc), an Immune Checkpoint Inhibitor Blocking the CD47 “Do Not Eat” Signal, Induces Objective Responses in Patients with Advanced, Relapsed/Refractory Diffuse Large B-Cell Lymphoma (DLBCL). Blood 2017, 130, 4116. [Google Scholar] [CrossRef]
- Ansell, S.M.; Maris, M.B.; Lesokhin, A.M.; Chen, R.W.; Flinn, I.W.; Sawas, A.; Minden, M.D.; Villa, D.; Percival, M.-E.M.; Advani, A.S.; et al. Phase I Study of the CD47 Blocker TTI-621 in Patients with Relapsed or Refractory Hematologic Malignancies. Clin. Cancer Res. 2021, 27, 2190–2199. [Google Scholar] [CrossRef]
- Querfeld, C.; Thompson, J.A.; Taylor, M.; Pillai, R.; Johnson, L.D.S.; Catalano, T.; Petrova, P.S.; Uger, B.A.; Irwin, M.; Thompson, T.; et al. Intralesional Injection of the CD47-Blocking Immune Checkpoint Inhibitor TTI-621 (SIRPaFc) Induces Antitumor Activity in Patients with Relapsed/Refractory Mycosis Fungoides and Sézary Syndrome: Interim Results of a Multicenter Phase 1 Trial. Eur. J. Cancer 2018, 101, S34. [Google Scholar] [CrossRef]
- Patel, K. Investigational CD47-Blocker TTI-622 Shows Single-Agent Activity in Patients with Advanced Relapsed or Refractory Lymphoma: Update from the Ongoing First-in-Human Dose Escalation Study. Blood 2020, 136, 46–47. [Google Scholar] [CrossRef]
- Weiskopf, K.; Ring, A.M.; Ho, C.C.M.; Volkmer, J.-P.; Levin, A.M.; Volkmer, A.K.; Ozkan, E.; Fernhoff, N.B.; van de Rijn, M.; Weissman, I.L.; et al. Engineered SIRPα Variants as Immunotherapeutic Adjuvants to Anticancer Antibodies. Science 2013, 341, 88–91. [Google Scholar] [CrossRef] [Green Version]
- Lakhani, N.J.; LoRusso, P.; Hafez, N.; Krishnamurthy, A.; O’Rourke, T.J.; Kamdar, M.K.; Fanning, P.; Zhao, Y.; Jin, F.; Wan, H.; et al. A Phase 1 Study of ALX148, a CD47 Blocker, Alone and in Combination with Established Anticancer Antibodies in Patients with Advanced Malignancy and Non-Hodgkin Lymphoma. J. Clin. Oncol. 2018, 36, 3068. [Google Scholar] [CrossRef]
- Kauder, S.E.; Kuo, T.C.; Harrabi, O.; Chen, A.; Sangalang, E.; Doyle, L.; Rocha, S.S.; Bollini, S.; Han, B.; Sim, J.; et al. ALX148 Blocks CD47 and Enhances Innate and Adaptive Antitumor Immunity with a Favorable Safety Profile. PLoS ONE 2018, 13, e0201832. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.M.; Laknahi, N.; Gainor, J.; Kamdar, M.; Fanning, P.; Squifflet, P.; Jin, F.; Forgie, A.J.; Wan, H.; Pons, J.; et al. ALX148, a CD47 Blocker, in Combination with Rituximab in Patients with Non-Hodgkin Lymphoma. Blood 2020, 136, 13–14. [Google Scholar] [CrossRef]
- Chow, L.Q.M.; Gainor, J.F.; Lakhani, N.J.; Lee, K.W.; Chung, H.C.; Lee, J.; LoRusso, P.; Bang, Y.-J.; Hodi, F.S.; Santana-Davila, R.; et al. A Phase I Study of ALX148, a CD47 Blocker, in Combination with Standard Anticancer Antibodies and Chemotherapy Regimens in Patients with Advanced Malignancy. J. Clin. Oncol. 2020, 38, 3056. [Google Scholar] [CrossRef]
- Adams, K.F.; Leitzmann, M.F.; Albanes, D.; Kipnis, V.; Moore, S.C.; Schatzkin, A.; Chow, W.-H. Body Size and Renal Cell Cancer Incidence in a Large US Cohort Study. Am. J. Epidemiol. 2008, 168, 268–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, F.; Yang, Y.; Chen, J.; Hu, X. SIRP/CD47 Signaling in Neurological Disorders. Brain Res. 2015, 1623, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanidakis, M.; Newton, G.; Lee, W.Y.; Parkos, C.A.; Luscinskas, F.W. Endothelial CD47 Interaction with SIRPgamma Is Required for Human T-Cell Transendothelial Migration under Shear Flow Conditions in Vitro. Blood 2008, 112, 1280–1289. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, A.; Ohnishi, H.; Okazawa, H.; Nakazawa, S.; Ikeda, H.; Motegi, S.; Aoki, N.; Kimura, S.; Mikuni, M.; Matozaki, T. Positive Regulation of Phagocytosis by SIRPbeta and Its Signaling Mechanism in Macrophages. J. Biol. Chem. 2004, 279, 29450–29460. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Soto, I.; Tong, Q.; Chin, A.; Bühring, H.-J.; Wu, T.; Zen, K.; Parkos, C.A. SIRPbeta1 Is Expressed as a Disulfide-Linked Homodimer in Leukocytes and Positively Regulates Neutrophil Transepithelial Migration. J. Biol. Chem. 2005, 280, 36132–36140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccio, L.; Vermi, W.; Boles, K.S.; Fuchs, A.; Strader, C.A.; Facchetti, F.; Cella, M.; Colonna, M. Adhesion of Human T Cells to Antigen-Presenting Cells through SIRPβ2-CD47 Interaction Costimulates T-Cell Proliferation. Blood 2005, 105, 2421–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forty Seven to Gilead: “Eat Me”. Nat. Biotechnol. 2020, 38, 389. [CrossRef] [PubMed] [Green Version]
- Delord, J.-P.; Kotecki, N.; Marabelle, A.; Vinceneux, A.; Korakis, I.; Jungels, C.; Champiat, S.; Huhn, R.D.; Poirier, N.; Costantini, D.; et al. A Phase 1 Study Evaluating BI 765063, a First in Class Selective Myeloid Sirpa Inhibitor, As Stand-Alone and in Combination with BI 754091, a Programmed Death-1 (PD-1) Inhibitor, in Patients with Advanced Solid Tumours. Blood 2019, 134, 1040. [Google Scholar] [CrossRef]
- Champiat, S.; Cassier, P.A.; Kotecki, N.; Korakis, I.; Vinceneux, A.; Jungels, C.; Blatchford, J.; Elgadi, M.M.; Clarke, N.; Fromond, C.; et al. Safety, Pharmacokinetics, Efficacy, and Preliminary Biomarker Data of First-in-Class BI 765063, a Selective SIRPα Inhibitor: Results of Monotherapy Dose Escalation in Phase 1 Study in Patients with Advanced Solid Tumors. J. Clin. Oncol. 2021, 39, 2623. [Google Scholar] [CrossRef]
- Voets, E.; Paradé, M.; Lutje Hulsik, D.; Spijkers, S.; Janssen, W.; Rens, J.; Reinieren-Beeren, I.; van den Tillaart, G.; van Duijnhoven, S.; Driessen, L.; et al. Functional Characterization of the Selective Pan-Allele Anti-SIRPα Antibody ADU-1805 That Blocks the SIRPα-CD47 Innate Immune Checkpoint. J. Immunother. Cancer 2019, 7, 340. [Google Scholar] [CrossRef] [PubMed]
- Ho CC, M.; Guo, N.; Sockolosky, J.T.; Ring, A.M.; Weiskopf, K.; Özkan, E.; Mori, Y.; Weissman, I.L.; Garcia, K.C. “Velcro” Engineering of High Affinity CD47 Ectodomain as Signal Regulatory Protein α (SIRPα) Antagonists That Enhance Antibody-Dependent Cellular Phagocytosis. J. Biol. Chem. 2015, 290, 12650–12663. [Google Scholar]
- Andrejeva, G.; Capoccia, B.J.; Hiebsch, R.R.; Donio, M.J.; Darwech, I.M.; Puro, R.J.; Pereira, D.S. Novel SIRPα Antibodies That Induce Single-Agent Phagocytosis of Tumor Cells While Preserving T Cells. J. Immunol. 2021, 206, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Sim, J.; Sockolosky, J.T.; Sangalang, E.; Izquierdo, S.; Pedersen, D.; Harriman, W.; Wibowo, A.S.; Carter, J.; Madan, A.; Doyle, L.; et al. Discovery of High Affinity, Pan-Allelic, and Pan-Mammalian Reactive Antibodies against the Myeloid Checkpoint Receptor SIRPα. MAbs 2019, 11, 1036–1052. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Liu, L.; Ren, Z.; Yang, K.; Xu, H.; Luan, Y.; Fu, K.; Guo, J.; Peng, H.; Zhu, M.; et al. Dual Targeting of Innate and Adaptive Checkpoints on Tumor Cells Limits Immune Evasion. Cell Rep. 2018, 24, 2101–2111. [Google Scholar] [CrossRef] [Green Version]
- Piccione, E.C.; Juarez, S.; Liu, J.; Tseng, S.; Ryan, C.E.; Narayanan, C.; Wang, L.; Weiskopf, K.; Majeti, R. A Bispecific Antibody Targeting CD47 and CD20 Selectively Binds and Eliminates Dual Antigen Expressing Lymphoma Cells. MAbs 2015, 7, 946–956. [Google Scholar] [CrossRef] [Green Version]
- Roohullah, A.; Ganju, V.; Zhang, F.; Zhang, L.; Yu, T.; Wilkinson, K.; Cooper, A.; de Souza, P. First-in-Human Phase 1 Dose Escalation Study of HX009, a Novel Recombinant Humanized Anti-PD-1 and CD47 Bispecific Antibody, in Patients with Advanced Malignancies. J. Clin. Oncol. 2021, 39, 2517. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, M.; Lin, P.; Liu, J.; Liu, C.; Strohl, W.R.; Wang, S.; Ho, M. Frontiers and Opportunities: Highlights of the 2nd Annual Conference of the Chinese Antibody Society. Antib. Ther. 2018, 1, 27–36. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, D.; Huang, C.; Chen, B.; Li, M.; Zhou, S.; Wang, L.; Wu, M.; Wang, X.; Bian, Y.; et al. Dose Escalation PET Imaging for Safety and Effective Therapy Dose Optimization of a Bispecific Antibody. MAbs 2020, 12, 1748322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ni, H.; Zhou, S.; He, K.; Gao, Y.; Wu, W.; Wu, M.; Wu, Z.; Qiu, X.; Zhou, Y.; et al. Tumor-Selective Blockade of CD47 Signaling with a CD47/PD-L1 Bispecific Antibody for Enhanced Anti-Tumor Activity and Limited Toxicity. Cancer Immunol. Immunother. 2021, 70, 365–376. [Google Scholar] [CrossRef] [PubMed]
- 33rd Annual Meeting & Pre-Conference Programs of the Society for Immunotherapy of Cancer (SITC 2018). J. Immunother. Cancer 2018, 6, 115. [CrossRef] [Green Version]
- Djureinovic, D.; Wang, M.; Kluger, H.M. Agonistic CD40 Antibodies in Cancer Treatment. Cancers 2021, 13, 1302. [Google Scholar] [CrossRef]
- De Silva, S.; Fromm, G.; Shuptrine, C.W.; Johannes, K.; Patel, A.; Yoo, K.J.; Huang, K.; Schreiber, T.H. CD40 Enhances Type I Interferon Responses Downstream of CD47 Blockade, Bridging Innate and Adaptive Immunity. Cancer Immunol. Res. 2020, 8, 230–245. [Google Scholar] [CrossRef] [Green Version]
- Elgueta, R.; Benson, M.J.; de Vries, V.C.; Wasiuk, A.; Guo, Y.; Noelle, R.J. Molecular Mechanism and Function of CD40/CD40L Engagement in the Immune System. Immunol. Rev. 2009, 229, 152–172. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Li, S.; Chen, D.; Liang, G.; Zhang, L.; Zhang, W.; Tu, X.; Peng, L.; Weng, J.; Zhao, G.; et al. Abstract 545: Preclinical Development of a Bispecific Antibody-Trap Selectively Targeting CD47 and CD20 for the Treatment of B Cell Lineage Cancer. Cancer Res. 2019, 79, 545. [Google Scholar] [CrossRef]
- Buatois, V.; Johnson, Z.; Salgado-Pires, S.; Papaioannou, A.; Hatterer, E.; Chauchet, X.; Richard, F.; Barba, L.; Daubeuf, B.; Cons, L.; et al. Preclinical Development of a Bispecific Antibody That Safely and Effectively Targets CD19 and CD47 for the Treatment of B-Cell Lymphoma and Leukemia. Mol. Cancer Ther. 2018, 17, 1739–1751. [Google Scholar] [CrossRef] [Green Version]
- Golubovskaya, V.; Berahovich, R.; Zhou, H.; Xu, S.; Harto, H.; Li, L.; Chao, C.-C.; Mao, M.M.; Wu, L. CD47-CAR-T Cells Effectively Kill Target Cancer Cells and Block Pancreatic Tumor Growth. Cancers 2017, 9, 139. [Google Scholar] [CrossRef] [Green Version]
- La, H.T.; Tran, D.B.T.; Tran, H.M.; Nguyen, L.T. Third-Generation Anti-CD47-Specific CAR-T Cells Effectively Kill Cancer Cells and Reduce the Genes Expression in Lung Cancer Cell Metastasis. J. Immunol. Res. 2021, 2021, e5575260. [Google Scholar] [CrossRef]
- Shu, R.; Evtimov, V.J.; Hammett, M.V.; Nguyen, N.-Y.N.; Zhuang, J.; Hudson, P.J.; Howard, M.C.; Pupovac, A.; Trounson, A.O.; Boyd, R.L. Engineered CAR-T Cells Targeting TAG-72 and CD47 in Ovarian Cancer. Mol. Ther. Oncolytics 2021, 20, 325–341. [Google Scholar] [CrossRef]
- Ma, L.; Zhu, M.; Gai, J.; Li, G.; Chang, Q.; Qiao, P.; Cao, L.; Chen, W.; Zhang, S.; Wan, Y. Preclinical Development of a Novel CD47 Nanobody with Less Toxicity and Enhanced Anti-Cancer Therapeutic Potential. J. Nanobiotechnol. 2020, 18, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Wang, C.; Zhang, X.; Chen, G.; Hu, Q.; Li, H.; Wang, J.; Wen, D.; Zhang, Y.; Lu, Y.; et al. In Situ Sprayed Bioresponsive Immunotherapeutic Gel for Post-Surgical Cancer Treatment. Nat. Nanotechnol. 2019, 14, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Castro, S.; Coker, C.; Hinchliffe, T.E.; Arpaia, N.; Danino, T. Programmable Bacteria Induce Durable Tumor Regression and Systemic Antitumor Immunity. Nat. Med. 2019, 25, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
CD47/SIRPα interaction leading to repression of phagocytosis and therapeutic approaches blocking CD47/SIRPα axis.
Figure 1.
CD47/SIRPα interaction leading to repression of phagocytosis and therapeutic approaches blocking CD47/SIRPα axis.


{kind=link}
{kind=link}
Table 1.
Clinical trials testing agents targeting CD47/SIRPα axis.
| Agent | Therapeutic Target | Design | Phase | Disease Site | Accrual Goal | Identifier |
|---|---|---|---|---|---|---|
| Monoclonal Antibodies | ||||||
| IBI188 (Letaplimab) | CD47 | IBI188 +/− rituximab | I | Metastatic solid tumors or lymphoma | 92 | NCT03717103 |
| IBI188 +/− azacitidine | I | Myelodysplastic syndrome | 12 | NCT04485065 | ||
| Hu5F9-G4 (Magrolimab) | CD47 | Hu5F9-G4 (Magrolimab) + Pembrolizumab | II | Hodgkin’s lymphoma | 24 | NCT04788043 |
| Hu5F9-G4 (Magrolimab) | I | Hematologic malignancies | 20 | NCT02678338 | ||
| Hu5F9-G4 (Magrolimab) + acalabrutinib + rituximab or other combinations without Hu5F9-G4 (Magrolimab) | I | Non-Hodgkin’s Lymphoma | 30 | NCT03527147 | ||
| Hu5F9-G4 (Magrolimab) + Obinutuzumab + venetoclax | I | Non-Hodgkin’s Lymphoma | 76 | NCT04599634 | ||
| ZL-1201 | CD47 | ZL-1201 | I | Metastatic solid tumors or refractory lymphomas | 66 | NCT04257617 |
| STI-6643 | CD47 | STI-6643 | I | Metastatic solid tumors | 24 | NCT04900519 |
| CC-9002 | CD47 | CC-90002 +/−rituximab | Part A: Metastatic solid tumors, multiple Myeloma or non-Hodgkin’s lymphoma Part B, relapsed and/or refractory CD20-positive NHL | 60 | NCT02367196 | |
| AK117 | CD47 | AK117 | I | Metastatic solid tumors or lymphoma | 162 | NCT04728334 |
| AK117 + azacitidine | I/II | Myelodysplastic syndrome | 190 | NCT04900350 | ||
| AO-176 | CD47 | AO-176 +/− paclitaxel | I/II | Metastatic solid tumors | 132 | NCT03834948 |
| AO-176 +/− dexamethasone or dexhamethasone + bortezomide | I | Multiple myeloma | 102 | NCT04445701 | ||
| IMC-002 | CD47 | IMC-002 | I | Metastatic solid tumors or lymphoma | 24 | NCT04306224 |
| TQB2928 | CD47 | TQB2928 | I | Metastatic solid tumors or hematologic malignancies | 20 | NCT04854681 |
| FSI-189 | SIRPα | FSI-189 +/− rituximab | I | Non-Hodgkin’s lymphoma (B-cell) | 63 | NCT04502706 |
| BI 765063 | SIRPα | BI 765063 +/− PD-1 inhibitor | I | Metastatic solid tumors with SIRPα polymorphism | 116 | NCT03990233 |
| Bispecific antibodies | ||||||
| HX009 | CD47 and PD-1 | HX009 | II | Metastatic solid tumors | 210 | NCT04886271 |
| PF-07257876 | CD47 and PD-L1 | PF-07257876 | I | Non small-cell lung cancer, head and neck squamous cell carcinoma, ovarian cancer | 90 | NCT04881045 |
| CPO107 (JMP601) | CD47 and CD20 | CPO107 (JMP601) | I | Non-Hodgkin’s lymphoma (CD-20 positive) | 75 | NCT04853329 |
| IBI322 | CD47 and PD-L1 | IBI322 | I | Hematologic malignancies | 182 | NCT04795128 |
| IBI322 | Ia | Metastatic solid tumors | 45 | NCT04338659 | ||
| IBI322 | Ia/Ib | Metastatic solid tumors | 218 | NCT04328831 | ||
| SL-172154 | SIRPα and CD40L | SL-172154 (intravenous) | I | Ovarian cancer | 40 | NCT04406623 |
| SL-172154 (intratumoral) | I | Head and neck or cutaneous squamous cell carcinoma | 18 | NCT04502888 | ||
| TG-1801 | CD47 and CD19 | TG-1801 +/− ubitixumab | Ib | Hematologic malignancies | 60 | NCT04806035 |
| IMM0306 | CD47 and CD20 | IMM0306 | I | Refractory or Relapsed CD20-positive B cell Non-Hodgkin’s Lymphoma | 131 | NCT04746131 |
| Fusion proteins | ||||||
| TTI-622 | CD47 via SIRPαFc (IgG4) structure | TTI-622 + rituximab, PD-1 inhibitor, Proteasome inhibitor regimen or rituximab | Ia/Ib | Lymphoma or myeloma | 156 | NCT03530683 |
| ALX148 | CD47 via SIRPαFc (IgG1) structure | ALX148 + azacitidine | I/II | Myelodysplastic syndrome | 173 | NCT04417517 |
| ALX148 + venetoclax or azacitidine | I/II | Acute myleoid leukemia | 97 | NCT04755244 | ||
| ALX148 | II | Head and neck squamous cell carcinoma | 112 | NCT04675333 | ||
| ALX148 + pembrolizumab | II | Head and neck squamous cell carcinoma | 111 | NCT04675294 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dizman, N.; Buchbinder, E.I. Cancer Therapy Targeting CD47/SIRPα. Cancers 2021, 13, 6229. https://doi.org/10.3390/cancers13246229
AMA Style
Dizman N, Buchbinder EI. Cancer Therapy Targeting CD47/SIRPα. Cancers. 2021; 13(24):6229. https://doi.org/10.3390/cancers13246229
Chicago/Turabian StyleDizman, Nazli, and Elizabeth I. Buchbinder. 2021. "Cancer Therapy Targeting CD47/SIRPα" Cancers 13, no. 24: 6229. https://doi.org/10.3390/cancers13246229
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.