
Gossypol Treatment Restores Insufficient Apoptotic Function of DFF40/CAD in Human Glioblastoma Cells

 , , , , , , and
, , , , , , and 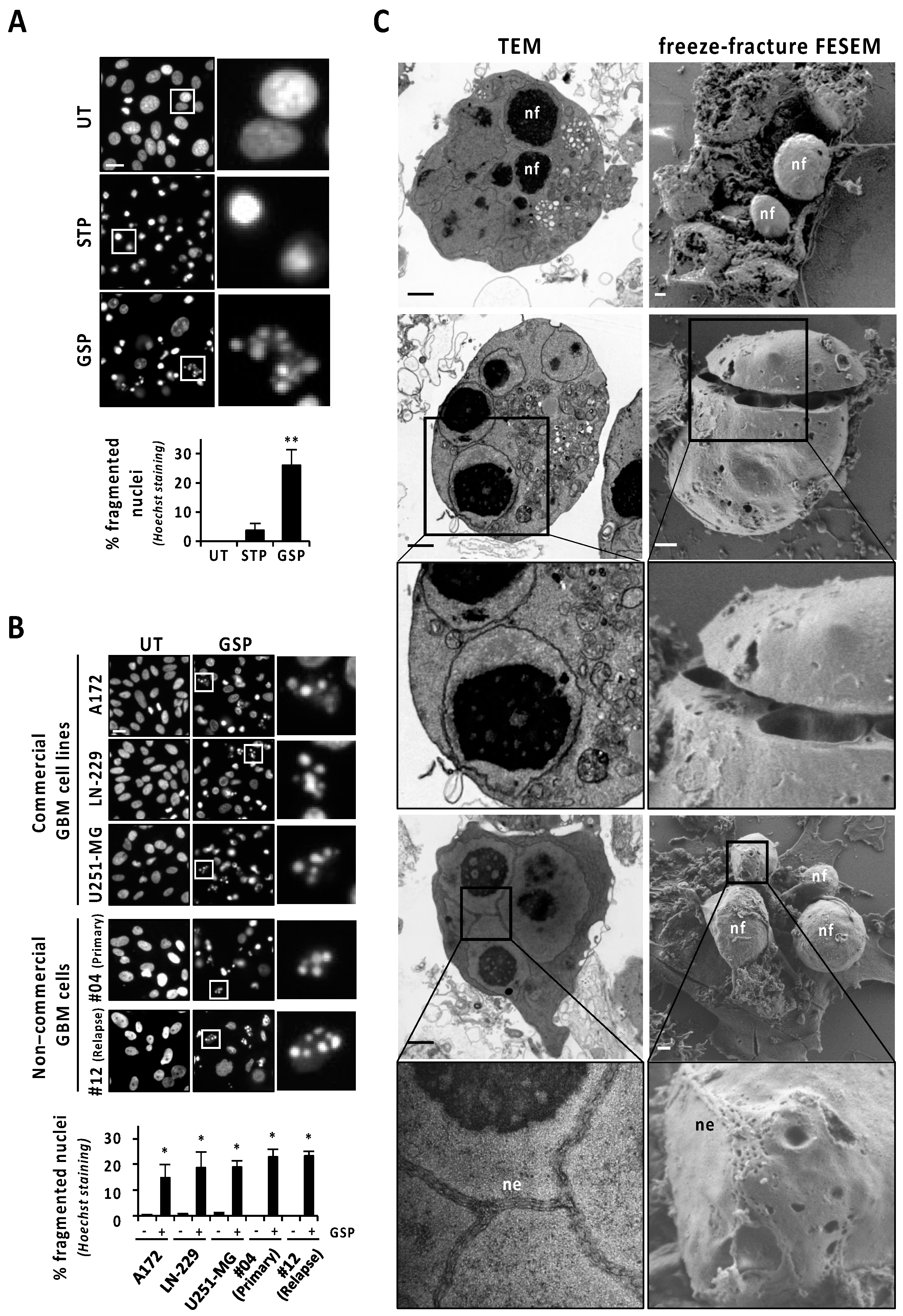{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Cell Lines
2.2. Patient-Derived Non-Commercial GBM Cell Cultures
2.3. DEVD-Directed Activity
2.4. Immunofluorescence
2.5. Lysosomes Labeling
2.6. Protein Extractions and Western Blotting
2.7. Transmission and Field Emission Scanning Electron Microscopy
2.7.1. Transmission Electron Microscope (TEM)
2.7.2. Field Emission-Scanning Electron Microscopy (FE-SEM)
2.8. Nuclear Morphology Analysis by Chromatin Staining
2.9. DFF40/CAD Overexpression and Silencing
2.10. TUNEL Assay
2.11. Statistical Analysis
3. Results
3.1. Human Glioblastoma Cells Show Nuclear Alterations Compatible with Apoptotic Nuclear Morphologies When Challenged with Gossypol
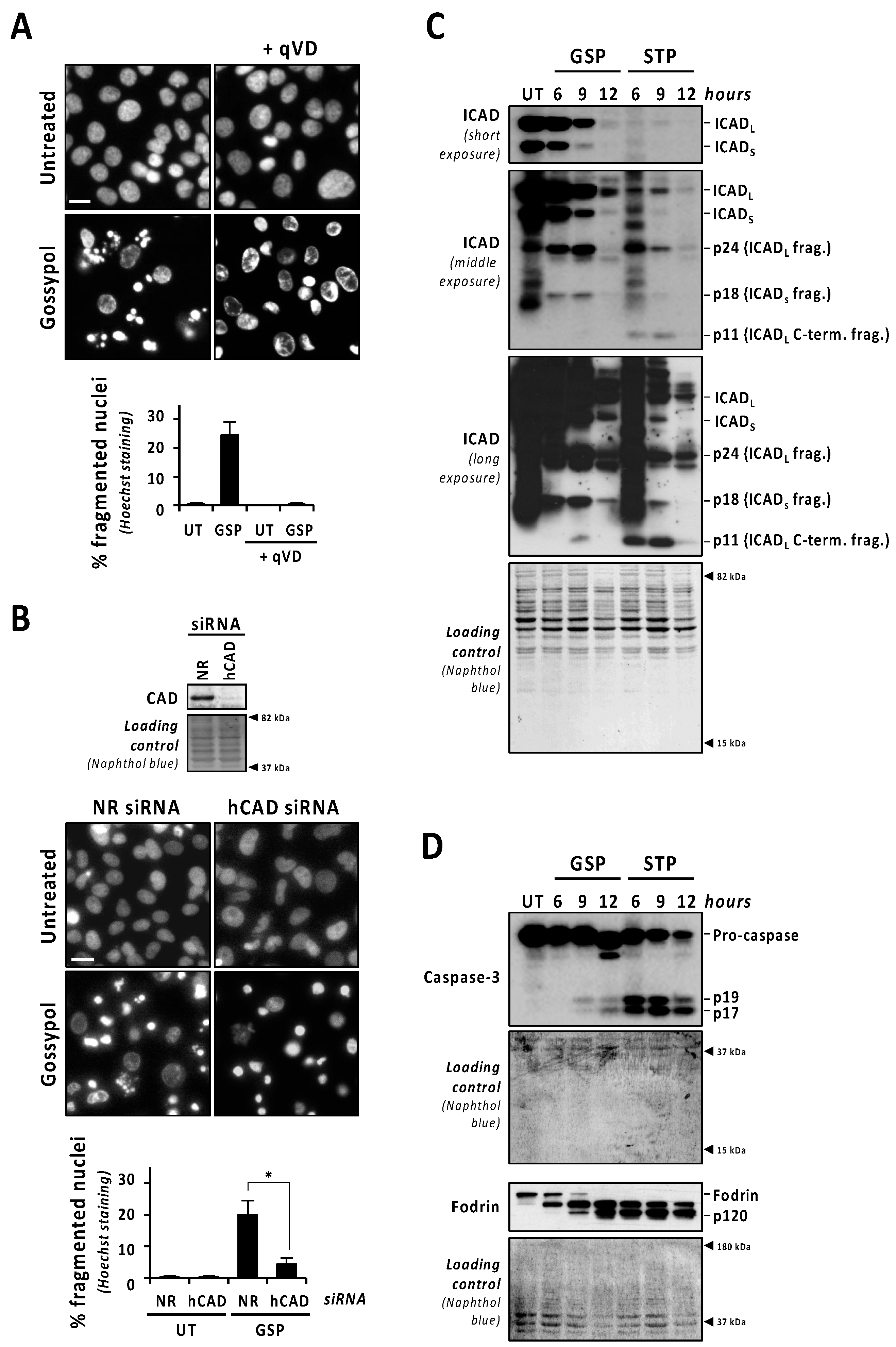
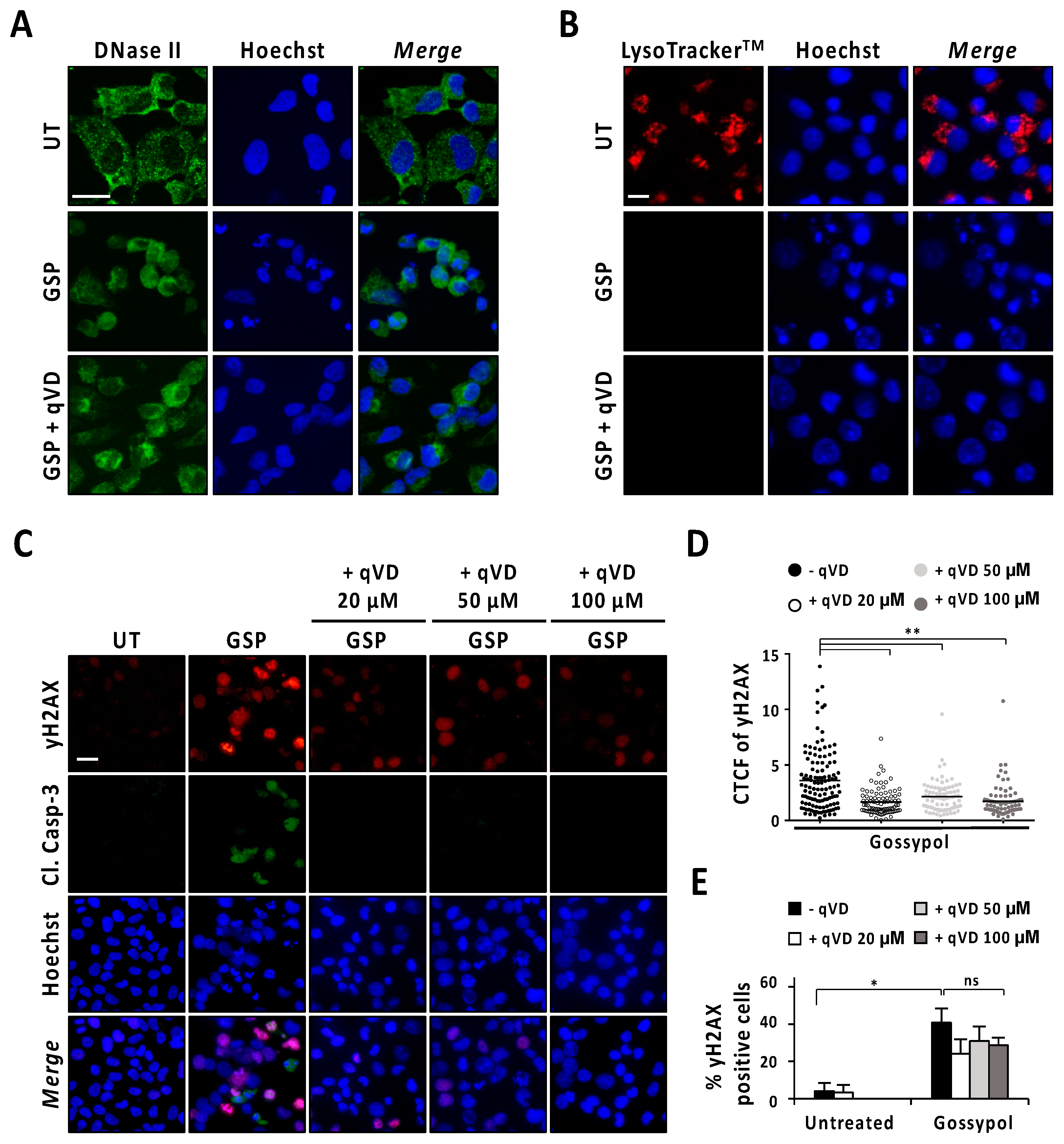
3.2. Caspases and DFF40/CAD Are Required for Gossypol-Induced Apoptotic Nuclear Morphologies
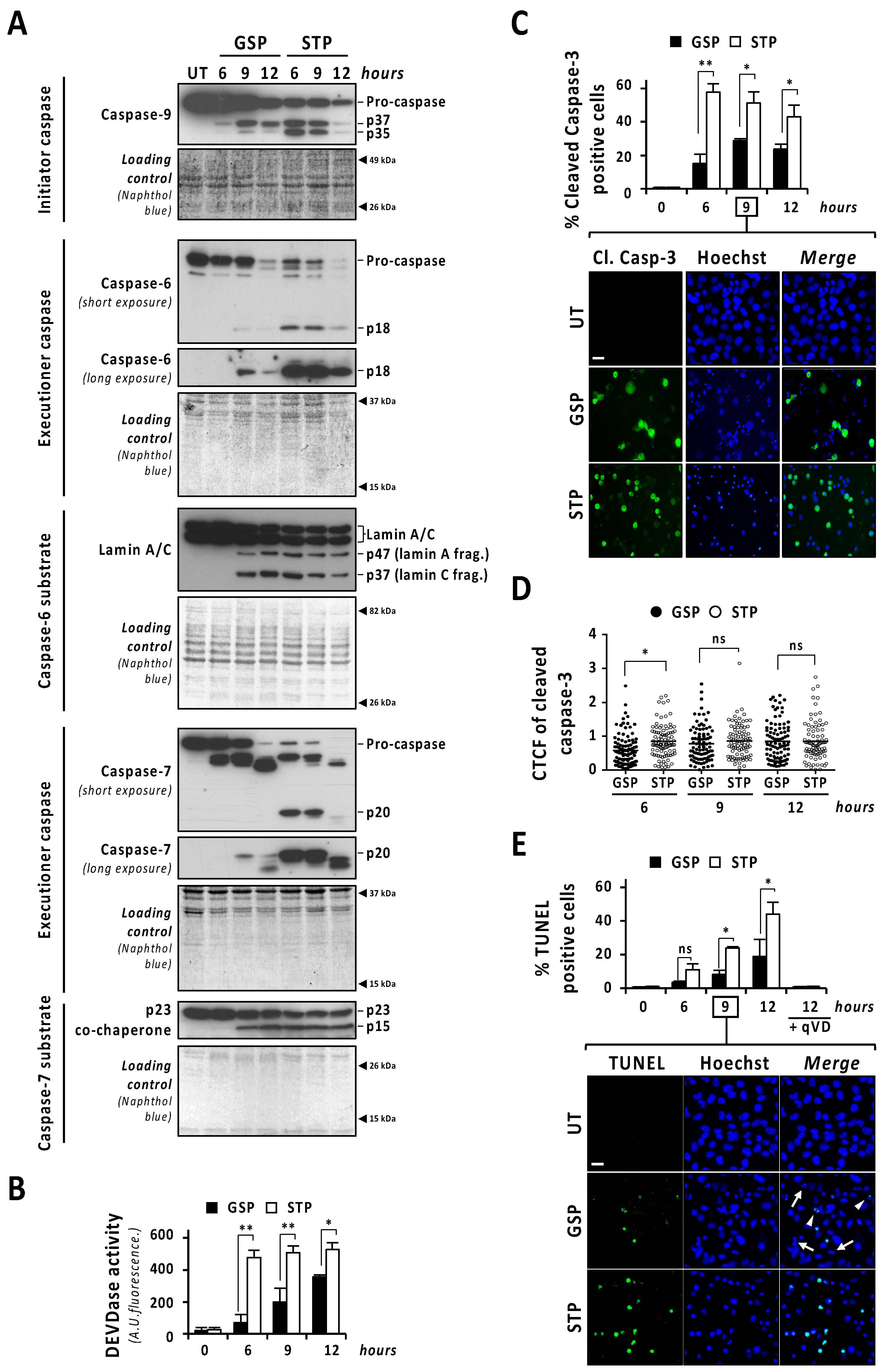
3.3. The Activation of Caspases and DFF40/CAD Are Necessary but Not Sufficient to Trigger Apoptotic Nuclear Disassembling after Gossypol Treatment
3.4. Gossypol Induces Both Caspase-Dependent and Caspase-Independent DNA Damage
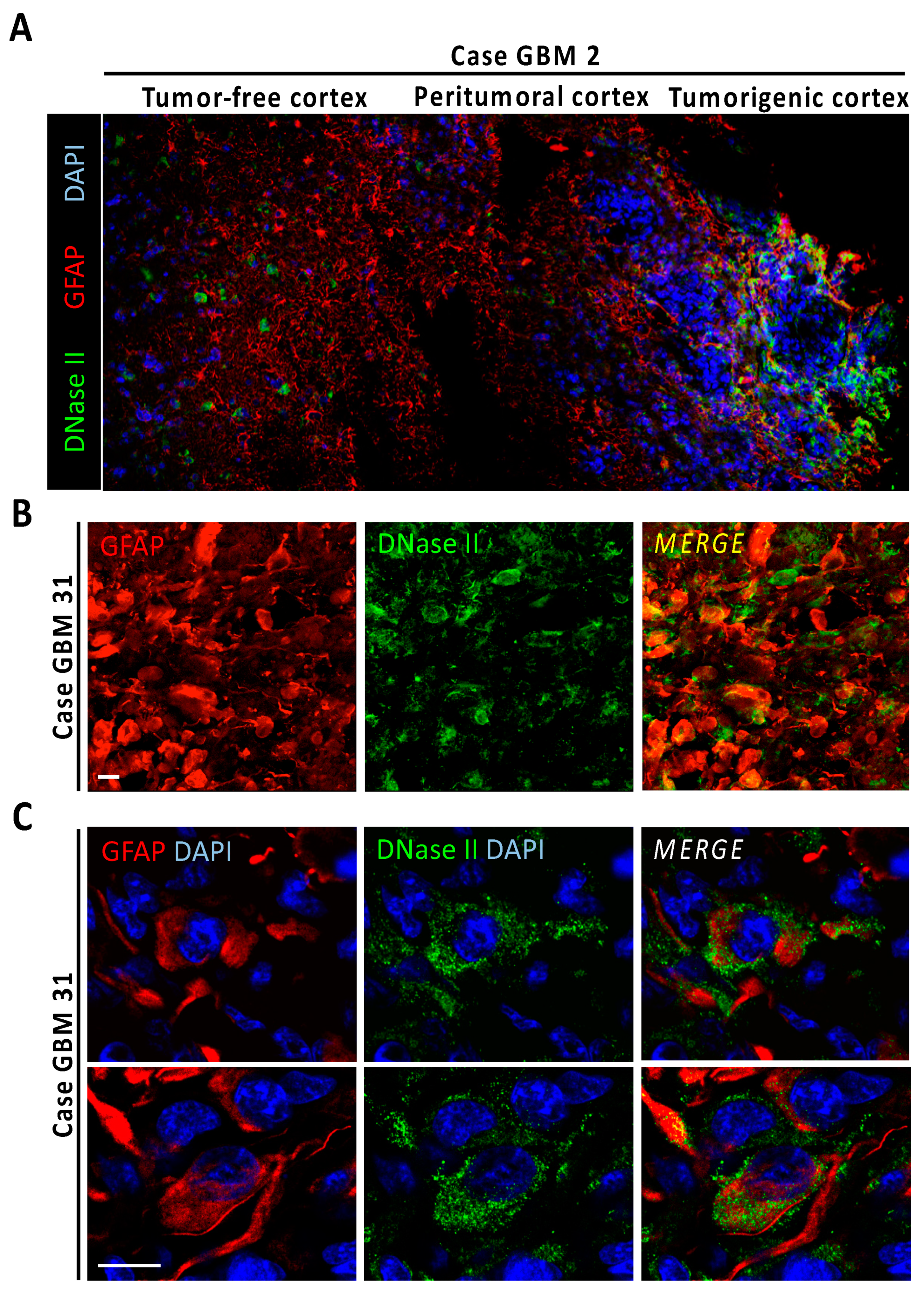
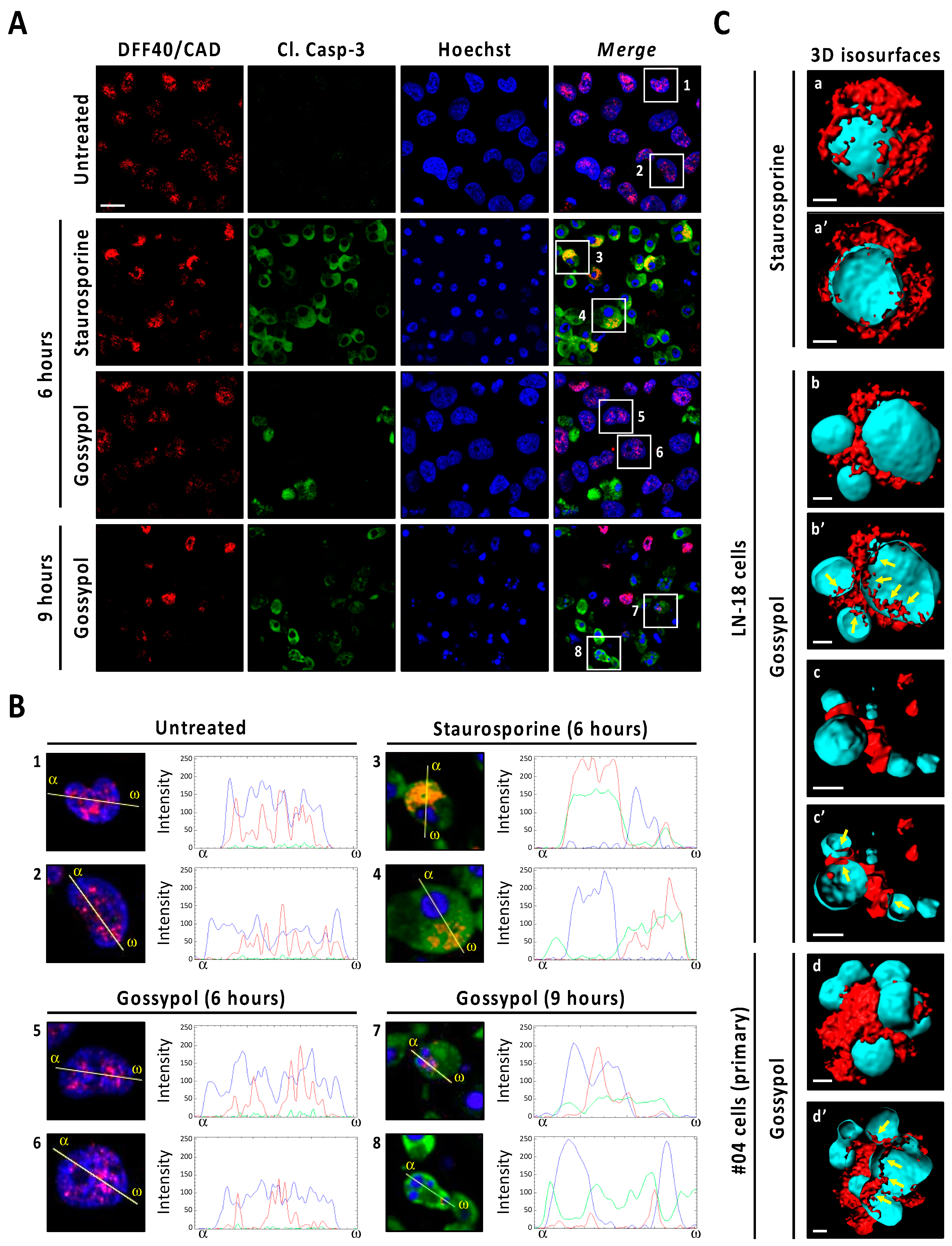
3.5. Gossypol Facilitates Nuclear DFF40/CAD Assembly into High-Order Structures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wen, P.Y.; Weller, M.; Lee, E.Q.; Alexander, B.M.; Barnholtz-Sloan, J.S.; Barthel, F.P.; Batchelor, T.T.; Bindra, R.S.; Chang, S.M.; Chiocca, E.A.; et al. Glioblastoma in adults: A Society for Neuro-Oncology (SNO) and European Society of Neuro-Oncology (EANO) consensus review on current management and future directions. Neuro-Oncology 2020, 22, 1073–1113. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E. Glioblastoma Treatments: An Account of Recent Industrial Developments. Front. Pharmacol. 2018, 9, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kögel, D.; Fulda, S.; Mittelbronn, M. Therapeutic exploitation of apoptosis and autophagy for glioblastoma. Anti-Cancer Agents Med. Chem. 2010, 10, 438–449. [Google Scholar] [CrossRef]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Saraste, A.; Pulkki, K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef]
- Liu, X.; Zou, H.; Slaughter, C.; Wang, X. DFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosis. Cell 1997, 89, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Enari, M.; Sakahira, H.; Yokoyama, H.; Okawa, K.; Iwamatsu, A.; Nagata, S. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature 1998, 391, 43–50. [Google Scholar] [CrossRef]
- Sakahira, H.; Enari, M.; Nagata, S. Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis. Nature 1998, 391, 96–99. [Google Scholar] [CrossRef]
- Sanchez-Osuna, M.; Martinez-Escardo, L.; Granados-Colomina, C.; Martinez-Soler, F.; Pascual-Guiral, S.; Iglesias-Guimarais, V.; Velasco, R.; Plans, G.; Vidal, N.; Tortosa, A.; et al. An intrinsic DFF40/CAD endonuclease deficiency impairs oligonucleosomal DNA hydrolysis during caspase-dependent cell death: A common trait in human glioblastoma cells. Neuro-Oncology 2016, 18, 950–961. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Osuna, M.; Garcia-Belinchon, M.; Iglesias-Guimarais, V.; Gil-Guinon, E.; Casanelles, E.; Yuste, V.J. Caspase-activated DNase Is Necessary and Sufficient for Oligonucleosomal DNA Breakdown, but Not for Chromatin Disassembly during Caspase-dependent Apoptosis of LN-18 Glioblastoma Cells. J. Biol. Chem. 2014, 289, 18752–18769. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Belinchon, M.; Sanchez-Osuna, M.; Martinez-Escardo, L.; Granados-Colomina, C.; Pascual-Guiral, S.; Iglesias-Guimarais, V.; Casanelles, E.; Ribas, J.; Yuste, V.J. An Early and Robust Activation of Caspases Heads Cells for a Regulated Form of Necrotic-like Cell Death. J. Biol. Chem. 2015, 290, 20841–20855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cribaro, G.P.; Saavedra-López, E.; Romarate, L.; Mitxitorena, I.; Díaz, L.R.; Casanova, P.V.; Roig-Martínez, M.; Gallego, J.M.; Perez-Vallés, A.; Barcia, C. Three-dimensional vascular microenvironment landscape in human glioblastoma. Acta Neuropathol. Commun. 2021, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Jakic, B.; Buszko, M.; Cappellano, G.; Wick, G. Elevated sodium leads to the increased expression of HSP60 and induces apoptosis in HUVECs. PLoS ONE 2017, 12, e0179383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozzelino, R.; Sole, C.; Llecha, N.; Segura, M.F.; Moubarak, R.S.; Iglesias-Guimarais, V.; Perez-Garcia, M.J.; Reix, S.; Zhang, J.; Badiola, N.; et al. BCL-XL regulates TNF-alpha-mediated cell death independently of NF-kappaB, FLIP and IAPs. Cell Res. 2008, 18, 1020–1036. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Guimarais, V.; Gil-Guinon, E.; Sanchez-Osuna, M.; Casanelles, E.; Garcia-Belinchon, M.; Comella, J.X.; Yuste, V.J. Chromatin collapse during caspase-dependent apoptotic cell death requires DNA fragmentation factor, 40-kDa subunit-/caspase-activated deoxyribonuclease-mediated 3′-OH single-strand DNA breaks. J. Biol. Chem. 2013, 288, 9200–9215. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Kidd, V.J. Cleavage of DFF-45/ICAD by multiple caspases is essential for its function during apoptosis. J. Biol. Chem. 1998, 273, 28549–28552. [Google Scholar] [CrossRef] [Green Version]
- Ruchaud, S.; Korfali, N.; Villa, P.; Kottke, T.J.; Dingwall, C.; Kaufmann, S.H.; Earnshaw, W.C. Caspase-6 gene disruption reveals a requirement for lamin A cleavage in apoptotic chromatin condensation. EMBO J. 2002, 21, 1967–1977. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.G.; Cullen, S.P.; Sheridan, C.; Luthi, A.U.; Gerner, C.; Martin, S.J. Executioner caspase-3 and caspase-7 are functionally distinct proteases. Proc. Natl. Acad. Sci. USA 2008, 105, 12815–12819. [Google Scholar] [CrossRef] [Green Version]
- Widlak, P.; Li, P.; Wang, X.; Garrard, W.T. Cleavage preferences of the apoptotic endonuclease DFF40 (caspase-activated DNase or nuclease) on naked DNA and chromatin substrates. J. Biol. Chem. 2000, 275, 8226–8232. [Google Scholar] [CrossRef] [Green Version]
- Keyel, P.A. Dnases in health and disease. Dev. Biol. 2017, 429, 1–11. [Google Scholar] [CrossRef]
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell. Proteom. MCP 2005, 4, 1920–1932. [Google Scholar] [CrossRef] [Green Version]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [Green Version]
- Janicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 is required for DNA fragmentation and morphological changes associated with apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar] [CrossRef] [Green Version]
- Yuste, V.J.; Bayascas, J.R.; Llecha, N.; Sanchez-Lopez, I.; Boix, J.; Comella, J.X. The absence of oligonucleosomal DNA fragmentation during apoptosis of IMR-5 neuroblastoma cells: Disappearance of the caspase-activated DNase. J. Biol. Chem. 2001, 276, 22323–22331. [Google Scholar] [CrossRef] [Green Version]
- Iglesias-Guimarais, V.; Gil-Guinon, E.; Gabernet, G.; Garcia-Belinchon, M.; Sanchez-Osuna, M.; Casanelles, E.; Comella, J.X.; Yuste, V.J. Apoptotic DNA degradation into oligonucleosomal fragments, but not apoptotic nuclear morphology, relies on a cytosolic pool of DFF40/CAD endonuclease. J. Biol. Chem. 2012, 287, 7766–7779. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Qiao, Q.; Hong, S.H.; Kim, C.M.; Jeong, J.H.; Kim, Y.G.; Jung, Y.K.; Wu, H.; Park, H.H. CIDE domains form functionally important higher-order assemblies for DNA fragmentation. Proc. Natl. Acad. Sci. USA 2017, 114, 7361–7366. [Google Scholar] [CrossRef] [Green Version]
- Dieker, J.; Iglesias-Guimarais, V.; Decossas, M.; Stevenin, J.; van der Vlag, J.; Yuste, V.J.; Muller, S. Early apoptotic reorganization of spliceosomal proteins involves caspases, CAD and rearrangement of NuMA. Traffic 2012, 13, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Sjakste, N.; Sjakste, T. Possible involvement of DNA strand breaks in regulation of cell differentiation. Eur. J. Histochem. 2007, 51, 81–94. [Google Scholar] [PubMed]
- Lechardeur, D.; Xu, M.; Lukacs, G.L. Contrasting nuclear dynamics of the caspase-activated DNase (CAD) in dividing and apoptotic cells. J. Cell Biol. 2004, 167, 851–862. [Google Scholar] [CrossRef] [Green Version]
- Aitchison, J.D.; Rout, M.P. A tense time for the nuclear envelope. Cell 2002, 108, 301–304. [Google Scholar] [CrossRef] [Green Version]
- Amoyel, M.; Bach, E.A. Cell competition: How to eliminate your neighbours. Development 2014, 141, 988–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levayer, R. Cell Competition: How to Take Over the Space Left by Your Neighbours. Curr. Biol. 2018, 28, R741–R744. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Wang, H.; Peng, Y.; Hu, Y.; Wang, H.; Zhang, X.; Chen, Q.; Bedford, J.S.; Dewhirst, M.W.; Li, C.Y. A unique role of the DNA fragmentation factor in maintaining genomic stability. Proc. Natl. Acad. Sci. USA 2006, 103, 1504–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, B.; Wang, H.; Wang, H.; Zhuo, D.; Li, F.; Kon, T.; Dewhirst, M.; Li, C.Y. Apoptotic DNA fragmentation factor maintains chromosome stability in a P53-independent manner. Oncogene 2006, 25, 5370–5376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, B.D.; Sørensen, C.S. The caspase-activated DNase: Apoptosis and beyond. FEBS J. 2017, 284, 1160–1170. [Google Scholar] [CrossRef]
- Kulbay, M.; Bernier-Parker, N.; Bernier, J. The role of the DFF40/CAD endonuclease in genomic stability. Apoptosis Int. J. Program. Cell Death 2021, 26, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Lovric, M.M.; Hawkins, C.J. TRAIL treatment provokes mutations in surviving cells. Oncogene 2010, 29, 5048–5060. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; He, Y.; Li, F.; Huang, Q.; Kato, T.A.; Hall, R.P.; Li, C.Y. Caspase-3 promotes genetic instability and carcinogenesis. Mol. Cell 2015, 58, 284–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, M.A.; Hawkins, C.J. Executioner caspases and CAD are essential for mutagenesis induced by TRAIL or vincristine. Cell Death Dis. 2017, 8, e3062. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Escardó, L.; Alemany, M.; Sánchez-Osuna, M.; Sánchez-Chardi, A.; Roig-Martínez, M.; Suárez-García, S.; Ruiz-Molina, D.; Vidal, N.; Plans, G.; Majós, C.; et al. Gossypol Treatment Restores Insufficient Apoptotic Function of DFF40/CAD in Human Glioblastoma Cells. Cancers 2021, 13, 5579. https://doi.org/10.3390/cancers13215579
Martínez-Escardó L, Alemany M, Sánchez-Osuna M, Sánchez-Chardi A, Roig-Martínez M, Suárez-García S, Ruiz-Molina D, Vidal N, Plans G, Majós C, et al. Gossypol Treatment Restores Insufficient Apoptotic Function of DFF40/CAD in Human Glioblastoma Cells. Cancers. 2021; 13(21):5579. https://doi.org/10.3390/cancers13215579
Chicago/Turabian StyleMartínez-Escardó, Laura, Montse Alemany, María Sánchez-Osuna, Alejandro Sánchez-Chardi, Meritxell Roig-Martínez, Salvio Suárez-García, Daniel Ruiz-Molina, Noemi Vidal, Gerard Plans, Carles Majós, and et al. 2021. "Gossypol Treatment Restores Insufficient Apoptotic Function of DFF40/CAD in Human Glioblastoma Cells" Cancers 13, no. 21: 5579. https://doi.org/10.3390/cancers13215579