
Experimental and Clinical Evidence Supports the Use of Urokinase Plasminogen Activation System Components as Clinically Relevant Biomarkers in Gastroesophageal Adenocarcinoma

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Major Components and Function of the uPAS
3. Regulation of the uPAS
4. The Clinical Relevance of uPAS Expression in GOCs
4.1. Tumour Expression and Association with Clinicopathological Features
4.2. Tumour Expression and Association with Clinical Outcomes
4.3. Intra-Tumoural Heterogeneity
4.4. Expression in Tumour-Associated Stromal Cells
4.5. Interactions of the uPAS with Other Proteolytic Enzymes
5. uPAS Assessment in Blood
5.1. Soluble uPAS Proteins in the Serum
5.2. uPAS Expression on Peripheral Blood Mononuclear Cells in GOC
5.3. Evidence of uPAS on Circulating Tumour Cells
6. Therapeutics and Diagnostics Directed towards the uPAS Pathway
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Enzinger, P.C.; Mayer, R.J. Esophageal cancer. N. Engl. J. Med. 2003, 349, 2241–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riihimaki, M.; Hemminki, A.; Sundquist, K.; Sundquist, J.; Hemminki, K. Metastatic spread in patients with gastric cancer. Oncotarget 2016, 7, 52307–52316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, N.; Howe, J.; Gay, G.; Patel-Parekh, L.; Scott-Conner, C.; Donohue, J. Differences in the pattern of presentation and treatment of proximal and distal gastric cancer: Results of the 2001 gastric patient care evaluation. Ann. Surg. Oncol. 2008, 15, 1644–1650. [Google Scholar] [CrossRef]
- Andreasen, P.A.; Kjøller, L.; Christensen, L.; Duffy, M.J. The urokinase-type plasminogen activator system in cancer metastasis: A review. Int. J. Cancer 1997, 72, 1–22. [Google Scholar] [CrossRef]
- Ranson, M.; Andronicos, N.M. Plasminogen binding and cancer: Promises and pitfalls. Front. Biosci. 2003, 8, s294–s304. [Google Scholar] [CrossRef] [Green Version]
- Laufs, S.; Schumacher, J.; Allgayer, H. Urokinase-receptor (u-PAR): An essential player in multiple games of cancer: A review on its role in tumor progression, invasion, metastasis, proliferation/dormancy, clinical outcome and minimal residual disease. Cell Cycle 2006, 5, 1760–1771. [Google Scholar]
- Duffy, M.J. The urokinase plasminogen activator system: Role in malignancy. Curr. Pharm. Des. 2004, 10, 39–49. [Google Scholar] [CrossRef]
- Dano, K.; Behrendt, N.; Hoyer-Hansen, G.; Johnsen, M.; Lund, L.R.; Ploug, M.; Romer, J. Plasminogen activation and cancer. Thromb. Haemost. 2005, 93, 676–681. [Google Scholar]
- McMahon, B.; Kwaan, H.C. The plasminogen activator system and cancer. Pathophysiol. Haemost. Thromb. 2008, 36, 184–194. [Google Scholar] [CrossRef]
- Croucher, D.R.; Saunders, D.N.; Lobov, S.; Ranson, M. Revisiting the biological roles of PAI2 (SERPINB2) in cancer. Nat. Rev. Cancer 2008, 8, 535–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alpízar-Alpízar, W.; Christensen, I.J.; Santoni-Rugiu, E.; Skarstein, A.; Ovrebo, K.; Illemann, M.; Laerum, O.D. Urokinase plasminogen activator receptor on invasive cancer cells: A prognostic factor in distal gastric adenocarcinoma. Int. J. Cancer 2012, 131, E329–E336. [Google Scholar] [CrossRef] [PubMed]
- Dass, K.; Ahmad, A.; Azmi, A.S.; Sarkar, S.H.; Sarkar, F.H. Evolving role of uPA/uPAR system in human cancers. Cancer Treat. Rev. 2008, 34, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Gårdsvoll, H.; Jacobsen, B.; Kriegbaum, M.C.; Behrendt, N.; Engelholm, L.; Østergaard, S.; Ploug, M. Conformational regulation of urokinase receptor function: Impact of receptor occupancy and epitope-mapped monoclonal antibodies on lamellipodia induction. J. Biol. Chem. 2011, 286, 33544–33556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, N.; Rabbani, S.A. Fibrinolytic System and Cancer: Diagnostic and Therapeutic Applications. Int. J. Mol. Sci. 2021, 22, 4358. [Google Scholar] [CrossRef]
- Scicolone, G.; Sanchez, V.; Vauthay, L.; Fuentes, F.; Scicolone, A.; Scicolone, L.; Rapacioli, M.; Flores, V. Tissue-type plasminogen activator activity in morphologically normal tissues adjacent to gastrointestinal carcinomas is associated with the degree of tumor progression. J. Cancer Res. Clin. Oncol. 2006, 132, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Brungs, D.; Chen, J.; Aghmesheh, M.; Vine, K.L.; Becker, T.M.; Carolan, M.G.; Ranson, M. The urokinase plasminogen activation system in gastroesophageal cancer: A systematic review and meta-analysis. Oncotarget 2017, 8, 23099–23109. [Google Scholar] [CrossRef] [Green Version]
- Laerum, O.D.; Ovrebo, K.; Skarstein, A.; Christensen, I.J.; Alpizar-Alpizar, W.; Helgeland, L.; Dano, K.; Nielsen, B.S.; Illemann, M. Prognosis in adenocarcinomas of lower oesophagus, gastro-oesophageal junction and cardia evaluated by uPAR-immunohistochemistry. Int. J. Cancer 2012, 131, 558–569. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.N.; Miyauchi, M.; Koshikawa, N.; Maruyama, K.; Kubota, T.; Miura, K.; Kurosawa, Y.; Awaya, A.; Kanai, Y. Antigen expression associated with lymph node metastasis in gastric adenocarcinomas. Pathol. Int. 1994, 44, 844–849. [Google Scholar] [CrossRef]
- Hong, S.I.; Park, I.C.; Son, Y.S.; Lee, S.H.; Kim, B.G.; Lee, J.I.; Lee, T.W.; Kook, Y.H.; Min, Y.I.; Hong, W.S. Expression of urokinase-type plasminogen activator, its receptor, and its inhibitor in gastric adenocarcinoma tissues. J. Korean Med. Sci. 1996, 11, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, K.; Hayashi, Y.; Wang, Y.; Suzuki, S.; Morita, Y.; Nakamura, T.; Narita, K.; Doe, W.; Itoh, H.; Kuroda, Y. Expression of urokinase-type plasminogen activator receptor and plasminogen activator inhibitor-1 in gastric cancer. J. Gastroenterol. Hepatol. 1998, 13, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Yonemura, Y.; Fujita, H.; Tsuchihara, K.; Kawamura, T.; Nojima, N.; Fujimura, T.; Nose, H.; Endo, Y.; Sasaki, T. Prognostic relevance of urokinase-type plasminogen activator (uPA) and plasminogen activator inhibitors PAI-1 and PAI-2 in gastric cancer. Virchows Arch. 1996, 427, 487–496. [Google Scholar] [CrossRef]
- Kaneko, T.; Konno, H.; Baba, M.; Tanaka, T.; Nakamura, S. Urokinase-type plasminogen activator expression correlates with tumor angiogenesis and poor outcome in gastric cancer. Cancer Sci. 2003, 94, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekarda, H.; Schlegel, P.; Schmitt, M.; Stark, M.; Mueller, J.D.; Fink, U.; Siewert, J.R. Strong prognostic impact of tumor-associated urokinase-type plasminogen activator in completely resected adenocarcinoma of the esophagus. Clin. Cancer Res. 1998, 4, 1755–1763. [Google Scholar]
- Nekarda, H.; Schmitt, M.; Ulm, K.; Wenninger, A.; Vogelsang, H.; Becker, K.; Roder, J.D.; Fink, U.; Siewert, J.R. Prognostic impact of urokinase-type plasminogen activator and its inhibitor PAI-1 in completely resected gastric cancer. Cancer Res. 1994, 54, 2900–2907. [Google Scholar]
- Beyer, B.C.; Heiss, M.M.; Simon, E.H.; Gruetzner, K.U.; Babic, R.; Jauch, K.W.; Schildberg, F.W.; Allgayer, H. Urokinase system expression in gastric carcinoma: Prognostic impact in an independent patient series and first evidence of predictive value in preoperative biopsy and intestinal metaplasia specimens. Cancer 2006, 106, 1026–1035. [Google Scholar] [CrossRef]
- Boonstra, M.C.; Verbeek, F.P.R.; Mazar, A.P.; Prevoo, H.A.J.M.; Kuppen, P.J.K.; van de Velde, C.J.H.; Vahrmeijer, A.L.; Sier, C.F.M. Expression of uPAR in tumor-associated stromal cells is associated with colorectal cancer patient prognosis: A TMA study. BMC Cancer 2014, 14, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploug, M.; Gardsvoll, H.; Jorgensen, T.J.; Lonborg Hansen, L.; Dano, K. Structural analysis of the interaction between urokinase-type plasminogen activator and its receptor: A potential target for anti-invasive cancer therapy. Biochem. Soc. Trans. 2002, 30, 177–183. [Google Scholar] [CrossRef]
- Deryugina, E.I.; Quigley, J. Cell surface remodeling by plasmin: A new function for an old enzyme. J. Biomed. Biotechnol. 2012, 2012, 564259. [Google Scholar] [CrossRef] [Green Version]
- Stamenkovic, I. Extracellular matrix remodelling: The role of matrix metalloproteinases. J. Pathol. 2003, 200, 448–464. [Google Scholar] [CrossRef]
- Smith, H.W.; Marshall, C.J. Regulation of cell signalling by uPAR. Nat. Rev. Mol. Cell Biol. 2010, 11, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Carriero, M.V.; Franco, P.; Votta, G.; Longanesi-Cattani, I.; Vento, M.T.; Masucci, M.T.; Mancini, A.; Caputi, M.; Iaccarino, I.; Stoppelli, M.P. Regulation of cell migration and invasion by specific modules of uPA: Mechanistic insights and specific inhibitors. Curr. Drug Targets 2011, 12, 1761–1771. [Google Scholar] [CrossRef]
- Ferraris, G.M.S.; Schulte, C.; Buttiglione, V.; De Lorenzi, V.; Piontini, A.; Galluzzi, M.; Podestà, A.; Madsen, C.D.; Sidenius, N. The interaction between uPAR and vitronectin triggers ligand-independent adhesion signalling by integrins. EMBO J. 2014, 33, 2458–2472. [Google Scholar] [CrossRef] [Green Version]
- Thuno, M.; Macho, B.; Eugen-Olsen, J. suPAR: The molecular crystal ball. Dis. Markers 2009, 27, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Fidan, E.; Mentese, A.; Ozdemir, F.; Deger, O.; Kavgaci, H.; Caner Karahan, S.; Aydin, F. Diagnostic and prognostic significance of CA IX and suPAR in gastric cancer. Med. Oncol. 2013, 30, 540. [Google Scholar] [CrossRef]
- Rohrberg, K.S.; Skov, B.G.; Lassen, U.; Christensen, I.J.; Høyer-Hansen, G.; Buysschaert, I.; Pappot, H. Markers of angiogenesis and epidermal growth factor receptor signalling in patients with pancreatic and gastroesophageal junction cancer. Cancer Biomark 2010, 7, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Masucci, M.T.; Pedersen, N.; Blasi, F. A soluble, ligand binding mutant of the human urokinase plasminogen activator receptor. J. Biol. Chem. 1991, 266, 8655–8658. [Google Scholar] [CrossRef]
- Høyer-Hansen, G.; Behrendt, N.; Ploug, M.; Danø, K.; Preissner, K.T. The intact urokinase receptor is required for efficient vitronectin binding: Receptor cleavage prevents ligand interaction. FEBS Lett. 1997, 420, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Ploug, M.; Rahbek-Nielsen, H.; Ellis, V.; Roepstorff, P.; Danø, K. Chemical modification of the urokinase-type plasminogen activator and its receptor using tetranitromethane. Evidence for the involvement of specific tyrosine residues in both molecules during receptor-ligand interaction. Biochemistry 1995, 34, 12524–12534. [Google Scholar] [CrossRef] [PubMed]
- Fazioli, F.; Resnati, M.; Sidenius, N.; Higashimoto, Y.; Appella, E.; Blasi, F. A urokinase-sensitive region of the human urokinase receptor is responsible for its chemotactic activity. EMBO J. 1997, 16, 7279–7286. [Google Scholar] [CrossRef] [Green Version]
- Selleri, C.; Montuori, N.; Ricci, P.; Visconte, V.; Carriero, M.V.; Sidenius, N.; Serio, B.; Blasi, F.; Rotoli, B.; Rossi, G.; et al. Involvement of the urokinase-type plasminogen activator receptor in hematopoietic stem cell mobilization. Blood 2005, 105, 2198–2205. [Google Scholar] [CrossRef] [Green Version]
- Resnati, M.; Pallavicini, I.; Wang, J.M.; Oppenheim, J.; Serhan, C.N.; Romano, M.; Blasi, F. The fibrinolytic receptor for urokinase activates the G protein-coupled chemotactic receptor FPRL1/LXA4R. Proc. Natl. Acad. Sci. USA 2002, 99, 1359–1364. [Google Scholar] [CrossRef] [Green Version]
- Placencio, V.R.; De Clerck, Y.A. Plasminogen Activator Inhibitor-1 in Cancer: Rationale and Insight for Future Therapeutic Testing. Cancer Res. 2015, 75, 2969–2974. [Google Scholar] [CrossRef] [Green Version]
- Herz, J.; Strickland, D.K. LRP: A multifunctional scavenger and signaling receptor. J. Clin. Invest. 2001, 108, 779–784. [Google Scholar] [CrossRef]
- Nykjær, A.; Conese, M.; Christensen, E.I.; Olson, D.; Cremona, O.; Gliemann, J.; Blasi, F. Recycling of the urokinase receptor upon internalization of the uPA:serpin complexes. EMBO J. 1997, 16, 2610–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochran, B.J.; Croucher, D.R.; Lobov, S.; Saunders, D.N.; Ranson, M. Dependence on endocytic receptor binding via a minimal binding motif underlies the differential prognostic profiles of SerpinE1 and SerpinB2 in cancer. J. Biol. Chem. 2011, 286, 24467–24475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubala, M.H.; DeClerck, Y.A. The plasminogen activator inhibitor-1 paradox in cancer: A mechanistic understanding. Cancer Metastasis Rev. 2019, 38, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Croucher, D.R.; Saunders, D.N.; Stillfried, G.E.; Ranson, M. A structural basis for differential cell signalling by PAI-1 and PAI-2 in breast cancer cells. Biochem. J. 2007, 408, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Harbeck, N.; Schmitt, M.; Meisner, C.; Friedel, C.; Untch, M.; Schmidt, M.; Sweep, C.G.; Lisboa, B.W.; Lux, M.P.; Beck, T.; et al. Ten-year analysis of the prospective multicentre Chemo-N0 trial validates American Society of Clinical Oncology (ASCO)-recommended biomarkers uPA and PAI-1 for therapy decision making in node-negative breast cancer patients. Eur. J. Cancer 2013, 49, 1825–1835. [Google Scholar] [CrossRef]
- Behrendt, N.; List, K.; Andreasen, P.A.; Danø, K. The pro-urokinase plasminogen-activation system in the presence of serpin-type inhibitors and the urokinase receptor: Rescue of activity through reciprocal pro-enzyme activation. Biochem. J. 2003, 371 Pt 2, 277–287. [Google Scholar] [CrossRef]
- Petersen, L.C.; Lund, L.R.; Nielsen, L.S.; Danø, K.; Skriver, L. One-chain urokinase-type plasminogen activator from human sarcoma cells is a proenzyme with little or no intrinsic activity. J. Biol. Chem. 1988, 263, 11189–11195. [Google Scholar] [CrossRef]
- Almholt, K.; Wang, J.; Pass, J.; Røder, G.; Padkjær, S.B.; Hebsgaard, J.B.; Xia, W.; Yang, L.; Forsell, J.; Breinholt, V.M.; et al. Identification and preclinical development of an anti-proteolytic uPA antibody for rheumatoid arthritis. J. Mol. Med. 2020, 98, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Dow, M.P.D. Gonadotrophin surge-induced upregulation of mRNA for plasminogen activator inhibitors 1 and 2 within bovine periovulatory follicular and luteal tissue. Reproduction 2002, 123, 711–719. [Google Scholar] [CrossRef]
- Xing, R.H.; Rabbani, S.A. Regulation of urokinase production by androgens in human prostate cancer cells: Effect on tumor growth and metastases in vivo. Endocrinology 1999, 140, 4056–4064. [Google Scholar] [CrossRef]
- Long, B.J.; Rose, D. Invasive capacity and regulation of urokinase-type plasminogen activator in estrogen receptor (ER)-negative MDA-MB-231 human breast cancer cells, and a transfectant (S30) stably expressing ER. Cancer Lett. 1996, 99, 209–215. [Google Scholar] [CrossRef]
- Casslén, B.; Nordengren, J.; Gustavsson, B.; Nilbert, M.; Lund, L.R. Progesterone stimulates degradation of urokinase plasminogen activator (u-PA) in endometrial stromal cells by increasing its inhibitor and surface expression of the u-PA receptor. J. Clin. Endocrinol. Metab. 1995, 80, 2776–2784. [Google Scholar] [PubMed]
- Korczak, B.; Kerbel, R.S.; Dennis, J.W. Autocrine and paracrine regulation of tissue inhibitor of metalloproteinases, transin, and urokinase gene expression in metastatic and nonmetastatic mammary carcinoma cells. Cell Growth Differ. 1991, 2, 335–341. [Google Scholar]
- Moriyama, T.; Kataoka, H.; Hamasuna, R.; Yoshida, E.; Sameshima, T.; Iseda, T.; Yokogami, K.; Nakano, S.; Koono, M.; Wakisaka, S. Simultaneous up-regulation of urokinase-type plasminogen activator (uPA) and uPA receptor by hepatocyte growth factor/scatter factor in human glioma cells. Clin. Exp. Metastasis 1999, 17, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, J.F.; Iglesias, M.; Frontelo, P.; Martínez, J.; Quintanilla, M. Involvement of the Ras/MAPK signaling pathway in the modulation of urokinase production and cellular invasiveness by transforming growth factor-beta(1) in transformed keratinocytes. Biochem. Biophys. Res. Commun. 2000, 273, 521–527. [Google Scholar] [CrossRef]
- Kirchheimer, J.C.; Nong, Y.H.; Remold, H.G. IFN-gamma, tumor necrosis factor-alpha; urokinase regulate the expression of urokinase receptors on human monocytes. J. Immunol. 1988, 141, 4229–4234. [Google Scholar] [PubMed]
- Kroon, M.E.; Koolwijk, P.; van der Vecht, B.; van Hinsbergh, V.W. Urokinase receptor expression on human microvascular endothelial cells is increased by hypoxia: Implications for capillary-like tube formation in a fibrin matrix. Blood 2000, 96, 2775–2783. [Google Scholar] [CrossRef]
- Bell, S.M.; Brackenbury, R.W.; Leslie, N.D.; Degen, J.L. Plasminogen activator gene expression is induced by the src oncogene product and tumor promoters. J. Biol. Chem. 1990, 265, 1333–1338. [Google Scholar] [CrossRef]
- Johnson, M.D.; Torri, J.A.; Lippman, M.E.; Dickson, R.B. Regulation of motility and protease expression in PKC-mediated induction of MCF-7 breast cancer cell invasiveness. Exp. Cell Res. 1999, 247, 105–113. [Google Scholar] [CrossRef]
- Niiya, K.; Ozawa, T.; Tsuzawa, T.; Ueshima, S.; Matsuo, O.; Sakuragawa, N. Transcriptional regulation of urokinase-type plasminogen activator receptor by cyclic AMP in PL-21 human myeloid leukemia cells: Comparison with the regulation by phorbol myristate acetate. Thromb. Haemost. 1998, 79, 574–578. [Google Scholar]
- Myohanen, H.; Vaheri, A. Regulation and interactions in the activation of cell-associated plasminogen. Cell Mol. Life Sci. 2004, 61, 2840–2858. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Choi, E.; Kim, M.; Kim, K.; Jang, B.; Kim, S.; Kim, S.; Song, S.; Kim, J.-R. Inhibition of histone deacetylase activity down-regulates urokinase plasminogen activator and matrix metalloproteinase-9 expression in gastric cancer. Mol. Cell Biochem. 2010, 343, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Kuga, H.; Morisaki, T.; Nakamura, K.; Onishi, H.; Noshiro, H.; Uchiyama, A.; Tanaka, M.; Katano, M. Interferon-gamma suppresses transforming growth factor-beta-induced invasion of gastric carcinoma cells through cross-talk of Smad pathway in a three-dimensional culture model. Oncogene 2003, 22, 7838–7847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, X.; Su, L.; Zhu, Z.; Wu, W.; Zhou, Y. Suppressive effects on cell proliferation and motility in gastric cancer SGC-7901 cells by introducing ulinastatin in vitro. Anticancer Drugs 2016, 27, 651–659. [Google Scholar] [CrossRef]
- Gong, M.; Lu, Z.; Fang, G.; Bi, J.; Xue, X. A small interfering RNA targeting osteopontin as gastric cancer therapeutics. Cancer Lett. 2008, 272, 148–159. [Google Scholar] [CrossRef]
- Lee, K.H.; Choi, E.Y.; Koh, S.A.E.; Kim, M.K.; Jang, B.I.; Kim, S.W.; Kim, J.-R. IL-1beta-stimulated urokinase plasminogen activator expression through NF-kappaB in gastric cancer after HGF treatment. Oncol Rep. 2014, 31, 2123–2130. [Google Scholar] [CrossRef] [Green Version]
- Onwuegbusi, B.; Rees, J.; Lao-Sirieix, P.; Fitzgerald, R. Selective loss of TGFbeta Smad-dependent signalling prevents cell cycle arrest and promotes invasion in oesophageal adenocarcinoma cell lines. PLoS ONE 2007, 2, e177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Ma, M.; Zhang, S. EGF-induced urokinase plasminogen activator receptor promotes epithelial to mesenchymal transition in human gastric cancer cells. Oncol. Rep. 2017, 38, 2325–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyung Hee, L.; Eun Young, C.; Min Kyoung, K.; Myung Soo, H.; Byung Ik, J.; Tae Nyeun, K.; Sang Woon, K.; Sun Kyo, S.; Jung Hye, K.; Jae-Ryong, K. Regulation of hepatocyte growth factor-mediated urokinase plasminogen activator secretion by MEK/ERK activation in human stomach cancer cell lines. Exp. Mol. Med. 2006, 38, 27–35. [Google Scholar]
- Lian, S.; Xia, Y.; Ung, T.T.; Khoi, P.N.; Yoon, H.J.; Lee, S.G.; Kim, K.K.; Jung, Y.D. Prostaglandin E2 stimulates urokinase-type plasminogen activator receptor via EP2 receptor-dependent signaling pathways in human AGS gastric cancer cells. Mol. Carcinog. 2017, 56, 664–680. [Google Scholar] [CrossRef] [PubMed]
- Shin, V.Y.; Wu, W.K.K.; Chu, K.-M.; Wong, H.P.S.; Lam, E.K.Y.; Tai, E.K.K.; Koo, M.W.L.; Cho, C.-H. Nicotine induces cyclooxygenase-2 and vascular endothelial growth factor receptor-2 in association with tumor-associated invasion and angiogenesis in gastric cancer. Mol. Cancer Res. 2005, 3, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Yang, Y.; Lee, S.J.; Kim, K.-Y.; Koo, T.H.; Shin, S.M.; Song, K.S.; Lee, Y.H.; Kim, Y.-J.; Lee, J.J.; et al. Macrophage inhibitory cytokine-1 induces the invasiveness of gastric cancer cells by up-regulating the urokinase-type plasminogen activator system. Cancer Res. 2003, 63, 4648–4655. [Google Scholar] [PubMed]
- Li, P.-Y.; Lv, J.; Qi, W.-W.; Zhao, S.-F.; Sun, L.-B.; Liu, N.; Sheng, J.; Qiu, W.-S. Tspan9 inhibits the proliferation, migration and invasion of human gastric cancer SGC7901 cells via the ERK1/2 pathway. Oncol. Rep. 2016, 36, 448–454. [Google Scholar] [CrossRef] [Green Version]
- Kyung Hee, L.; Eun Young, C.; Sung Ae, K.; Min Kyoung, K.; Kyeong Ok, K.; Si Hyung, L.; Byung Ik, J.; Se Won, K.; Sang Woon, K.; Sun Kyo, S.; et al. Down-regulation of survivin suppresses uro-plasminogen activator through transcription factor JunB. Exp. Mol. Med. 2011, 43, 501–509. [Google Scholar]
- Liu, L.; Sun, L.; Zhao, P.; Yao, L.; Jin, H.; Liang, S.; Wang, Y.; Zhang, D.; Pang, Y.; Shi, Y.; et al. Hypoxia promotes metastasis in human gastric cancer by up-regulating the 67-kDa laminin receptor. Cancer Sci. 2010, 101, 1653–1660. [Google Scholar] [CrossRef]
- Jin, H.; Pan, Y.; He, L.; Zhai, H.; Li, X.; Zhao, L.; Sun, L.; Liu, J.; Hong, L.; Song, J.; et al. p75 neurotrophin receptor inhibits invasion and metastasis of gastric cancer. Mol. Cancer Res. 2007, 5, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Hewin, D.F.; Savage, P.B.; Alderson, D.; Vipond, M.N. Plasminogen activators in oesophageal carcinoma. Br. J. Surg. 1996, 83, 1152–1155. [Google Scholar] [CrossRef]
- Hourihan, R.N.; O’Sullivan, G.C.; Morgan, J.G. Transcriptional gene expression profiles of oesophageal adenocarcinoma and normal oesophageal tissues. Anticancer Res. 2003, 23, 161–165. [Google Scholar]
- Gouri, A.; Dekaken, A.; El Bairi, K.; Aissaoui, A.; Laabed, N.; Chefrour, M.; Ciccolini, J.; Milano, G.; Benharkat, S. Plasminogen Activator System and Breast Cancer: Potential Role in Therapy Decision Making and Precision Medicine. Biomark Insights 2016, 11, 105–111. [Google Scholar] [CrossRef]
- Umehara, Y.; Kimura, T.; Yoshida, M.; Oba, N.; Harada, Y. Relationship between plasminogen activators and stomach carcinoma stage. Acta Oncol 1991, 30, 815–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, Y.-S.; Yu, J.; Kim, B.C.; Choi, B.; Han, T.-S.; Ahn, H.S.; Kong, S.-H.; Lee, H.-J.; Kim, W.H.; Yang, H.-K. Overexpression of Plasminogen Activator Inhibitor-1 in Advanced Gastric Cancer with Aggressive Lymph Node Metastasis. Cancer Res. Treat. 2015, 47, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Chung, Y.; Sawada, T.; Ogawa, Y.; Onoda, N.; Nakata, B.; Kato, Y.; Sowa, M. Combined evaluation of urokinase-type plasminogen activator and plasminogen activator inhibitor-2 expression in gastric carcinoma. Int. J. Oncol. 1996, 8, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Konno, H.; Tanaka, T.; Maruo, Y.; Nishino, N.; Aoki, K.; Baba, S.; Sakaguchi, S.; Takada, Y.; Takada, A. Possible role of plasminogen activator inhibitor 2 in the prevention of the metastasis of gastric cancer tissues. Thromb. Res. 1992, 65, 709–719. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Zhang, H.; Zhong, M.; Zhou, Z.; Zhuang, Z.; Yin, H.; Wang, X.; Zhu, Z. Clinical significance of the uPA system in gastric cancer with peritoneal metastasis. Eur. J. Med. Res. 2013, 18, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Zhang, H.; Lu, A.; Zhou, Z.; Zhong, M.; Shen, D.; Wang, X.; Zhu, Z. Effect of urokinase-type plasminogen activator system in gastric cancer with peritoneal metastasis. Oncol. Lett. 2016, 11, 4208–4216. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, N.; Matsuoka, T.; Yashiro, M.; Hirakawa, K.; Olden, K.; Roberts, J.D. Plasminogen activator inhibitor 1 RNAi suppresses gastric cancer metastasis in vivo. Cancer Sci. 2012, 103, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Heiss, M.M.; Babic, R.; Allgayer, H.; Gruetzner, K.U.; Jauch, K.W.; Loehrs, U.; Schildberg, F.W. Tumor-associated proteolysis and prognosis: New functional risk factors in gastric cancer defined by the urokinase-type plasminogen activator system. J. Clin. Oncol. 1995, 13, 2084–2093. [Google Scholar] [CrossRef]
- Brungs, D.; Lochhead, A.; Iyer, A.; Illemann, M.; Colligan, P.; Hirst, N.G.; Splitt, A.; Liauw, W.; Vine, K.L.; Pathmanandavel, S.; et al. Expression of cancer stem cell markers is prognostic in metastatic gastroesophageal adenocarcinoma. Pathology 2019, 51, 474–480. [Google Scholar] [CrossRef]
- Plava, J.; Cihova, M.; Burikova, M.; Matuskova, M.; Kucerova, L.; Miklikova, S. Recent advances in understanding tumor stroma-mediated chemoresistance in breast cancer. Mol. Cancer 2019, 18, 67. [Google Scholar] [CrossRef] [PubMed]
- Valkenburg, K.C.; de Groot, A.E.; Pienta, K.J. Targeting the tumour stroma to improve cancer therapy. Nat. Rev. Clin. Oncol. 2018, 15, 366–381. [Google Scholar] [CrossRef]
- Bussard, K.M.; Mutkus, L.; Stumpf, K.; Gomez-Manzano, C.; Marini, F.C. Tumor-associated stromal cells as key contributors to the tumor microenvironment. Breast Cancer Res. 2016, 18, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sounni, N.E.; Noel, A. Targeting the tumor microenvironment for cancer therapy. Clin. Chem. 2013, 59, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambardella, V.; Castillo, J.; Tarazona, N.; Gimeno-Valiente, F.; Martínez-Ciarpaglini, C.; Cabeza-Segura, M.; Roselló, S.; Roda, D.; Huerta, M.; Cervantes, A.; et al. The role of tumor-associated macrophages in gastric cancer development and their potential as a therapeutic target. Cancer Treat. Rev. 2020, 86, 102015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, B.S.; Sehested, M.; Duun, S.; Rank, F.; Timshel, S.; Rygaard, J.; Johnsen, M.; Danø, K. Urokinase plasminogen activator is localized in stromal cells in ductal breast cancer. Lab. Invest. 2001, 81, 1485–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyke, C.; Graem, N.; Ralfkiaer, E.; Ronne, E.; Hoyerhansen, G.; Brunner, N.; Dano, K. Receptor for Urokinase Is Present in Tumor-Associated Macrophages in Ductal Breast-Carcinoma. Cancer Res. 1993, 53, 1911–1915. [Google Scholar]
- Bianchi, E.; Cohen, R.L.; Thor, A.T.; Todd, R.F.; Mizukami, I.F.; Lawrence, D.A.; Ljung, B.M.; Shuman, M.A.; Smith, H.S. The urokinase receptor is expressed in invasive breast cancer but not in normal breast tissue. Cancer Res. 1994, 54, 861–866. [Google Scholar]
- Bianchi, E.; Cohen, R.L.; Dai, A.; Thor, A.T.; Shuman, M.A.; Smith, H.S. Immunohistochemical localization of the plasminogen activator inhibitor-1 in breast cancer. Int. J. Cancer 1995, 60, 597–603. [Google Scholar] [CrossRef]
- Costantini, V.; Sidoni, A.; Deveglia, R.; Cazzato, O.A.; Bellezza, G.; Ferri, I.; Bucciarelli, E.; Nenci, G.G. Combined overexpression of urokinase, urokinase receptor; plasminogen activator inhibitor-1 is associated with breast cancer progression-An immunohistochemical comparison of normal, benign; malignant breast tissues. Cancer 1996, 77, 1079–1088. [Google Scholar] [CrossRef]
- Ohtani, H.; Pyke, C.; Dan Ø, K.; Nagura, H. Expression of urokinase receptor in various stromal-cell populations in human colon cancer: Immunoelectron microscopical analysis. Int. J. Cancer 1995, 62, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Pyke, C.; Kristensen, P.; Ralfkiaer, E.; Grondahlhansen, J.; Eriksen, J.; Blasi, F.; Dano, K. Urokinase-Type Plasminogen-Activator Is Expressed in Stromal Cells and Its Receptor in Cancer-Cells at Invasive Foci in Human Colon Adenocarcinomas. Am. J. Pathol. 1991, 138, 1059–1067. [Google Scholar]
- Nakstad, B.; Lyberg, T. Immunohistochemical localization of coagulation, fibrinolytic and antifibrinolytic markers in adenocarcinoma of the lung. APMIS 1991, 99, 981–988. [Google Scholar] [CrossRef]
- Noruzi, S.; Azizian, M.; Mohammadi, R.; Hosseini, S.A.; Rashidi, B.; Mohamadi, Y.; Nesaei, A.; Seiri, P.; Sahebkar, A.; Salarinia, R.; et al. Micro-RNAs as critical regulators of matrix metalloproteinases in cancer. J. Cell Biochem. 2018, 119, 8694–8712. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.S.; Wang, Y.Y.; Ye, Z.Y.; Tao, H.Q. Prognostic value of tumor-related molecular expression in gastric carcinoma. Pathol. Oncol. Res. 2009, 15, 589–596. [Google Scholar] [CrossRef]
- Allgayer, H.; Babic, R.; Beyer, B.C.; Grützner, K.U.; Tarabichi, A.; Schildberg, F.W.; Heiss, M.M. Prognostic relevance of MMP-2 (72-kD collagenase IV) in gastric cancer. Oncology 1998, 55, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Ji, F.; Chen, Y.L.; Jin, E.Y.; Wang, W.L.; Yang, Z.L.; Li, Y.M. Relationship between matrix metalloproteinase-2 mRNA expression and clinicopathological and urokinase-type plasminogen activator system parameters and prognosis in human gastric cancer. World J. Gastroenterol. 2005, 11, 3222–3226. [Google Scholar] [CrossRef]
- Mijanović, O.; Branković, A.; Panin, A.N.; Savchuk, S.; Timashev, P.; Ulasov, I.; Lesniak, M.S. Cathepsin B: A sellsword of cancer progression. Cancer Lett. 2019, 449, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Gondi, C.S.; Rao, J.S. Cathepsin B as a cancer target. Expert Opin Ther Targets 2013, 17, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Moniwa, N.; Sugimura, M.; Shinohara, H.; Ohi, H.; Terao, T. Effects of membrane-associated cathepsin B on the activation of receptor-bound prourokinase and subsequent invasion of reconstituted basement membranes. Biochim. Biophys. Acta 1993, 1178, 55–62. [Google Scholar] [CrossRef]
- Herszényi, L.; István, G.; Cardin, R.; De Paoli, M.; Plebani, M.; Tulassay, Z.; Farinati, F. Serum cathepsin B and plasma urokinase-type plasminogen activator levels in gastrointestinal tract cancers. Eur. J. Cancer Prev. 2008, 17, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Kita, Y.; Fukagawa, T.; Mimori, K.; Kosaka, Y.; Ishikawa, K.; Aikou, T.; Natsugoe, S.; Sasako, M.; Mori, M. Expression of uPAR mRNA in peripheral blood is a favourite marker for metastasis in gastric cancer cases. Br. J. Cancer 2009, 100, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Vidal, Ó.; Metges, J.P.; Elizalde, I.; Valentíni, M.; Volant, A.; Molina, R.; Castells, A.; Pera, M. High preoperative serum vascular endothelial growth factor levels predict poor clinical outcome after curative resection of gastric cancer. Br. J. Surg. 2009, 96, 1443–1451. [Google Scholar] [CrossRef]
- Rao, J.S.; Gujrati, M.; Chetty, C. Tumor-associated soluble uPAR-directed endothelial cell motility and tumor angiogenesis. Oncogenesis 2013, 2, e53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolff, H.C.; Christensen, I.J.; Svendsen, L.B.; Wilhelmsen, M.; Lund, I.K.; Thurison, T.; Høyer-Hansen, G.; Illemann, M.; Nielsen, H.J. The concentration of the cleaved suPAR forms in pre- and postoperative plasma samples improves the prediction of survival in colorectal cancer: A nationwide multicenter validation and discovery study. J. Surg. Oncol. 2019, 120, 1404–1411. [Google Scholar] [CrossRef]
- Olingy, C.E.; Dinh, H.Q.; Hedrick, C.C. Monocyte heterogeneity and functions in cancer. J. Leukoc. Biol. 2019, 106, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodges, G.; Lyngbaek, S.; Selmer, C.; Ahlehoff, O.; Theilade, S.; Sehestedt, T.B.; Abildgaard, U.; Eugen-Olsen, J.; Galloe, A.M.; Hansen, P.R.; et al. SuPAR is associated with death and adverse cardiovascular outcomes in patients with suspected coronary artery disease. Scand. Cardiovasc. J. 2020, 54, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Hamie, L.; Daoud, G.; Nemer, G.; Nammour, T.; El Chediak, A.; Uthman, I.W.; Kibbi, A.G.; Eid, A.; Kurban, M. SuPAR, an emerging biomarker in kidney and inflammatory diseases. Postgrad. Med. J. 2018, 94, 517–524. [Google Scholar] [CrossRef]
- Ho, C.H.; Chao, Y.; Lee, S.D.; Chau, W.K.; Wu, C.W.; Liu, S.M. Diagnostic and prognostic values of plasma levels of fibrinolytic markers in gastric cancer. Thromb. Res. 1998, 91, 23–27. [Google Scholar] [CrossRef]
- Banys-Paluchowski, M.; Witzel, I.; Aktas, B.; Fasching, P.A.; Hartkopf, A.; Janni, W.; Kasimir-Bauer, S.; Pantel, K.; Schon, G.; Rack, B.; et al. The prognostic relevance of urokinase-type plasminogen activator (uPA) in the blood of patients with metastatic breast cancer. Sci. Rep. 2019, 9, 2318. [Google Scholar] [CrossRef] [Green Version]
- Piironen, T.; Laursen, B.; Pass, J.; List, K.; Gårdsvoll, H.; Ploug, M.; Danø, K.; Høyer-Hansen, G. Specific Immunoassays for Detection of Intact and Cleaved Forms of the Urokinase Receptor. Clin. Chem. 2004, 50, 2059–2068. [Google Scholar] [CrossRef] [Green Version]
- Loosen, S.H.; Tacke, F.; Binnebosel, M.; Leyh, C.; Vucur, M.; Heitkamp, F.; Schoening, W.; Ulmer, T.F.; Alizai, P.H.; Trautwein, C.; et al. Serum levels of soluble urokinase plasminogen activator receptor (suPAR) predict outcome after resection of colorectal liver metastases. Oncotarget 2018, 9, 27027–27038. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.Z.; Sheng, H.Y.; Zheng, X.; Ying, L.S.; Wu, L.; Liu, D.; Liu, G. Upregulation of circulating cytokeratin 20, urokinase plasminogen activator and C-reactive protein is associated with poor prognosis in gastric cancer. Mol. Clin. Oncol. 2015, 3, 1213–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, K.; Yang, S.; Zheng, L.; Wang, S.; Xiong, B. Prognostic Role of the Circulating Tumor Cells Detected by Cytological Methods in Gastric Cancer: A Meta-Analysis. Biomed. Res. Int. 2016, 2016, 2765464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.W.; Kim, G.H.; Jeon, H.K.; Park, S.J. Clinical Application of Circulating Tumor Cells in Gastric Cancer. Gut Liver 2019, 13, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Po, J.W. Importance and Detection of Epithelial-to-Mesenchymal Transition (EMT) Phenotype in CTCs, in Tumour Metastasis; Xu, K., Ed.; IntechOpen: London, UK, 2016. [Google Scholar]
- Jo, M.; Lester, R.D.; Montel, V.; Eastman, B.; Takimoto, S.; Gonias, S.L. Reversibility of epithelial-mesenchymal transition (EMT) induced in breast cancer cells by activation of urokinase receptor-dependent cell signaling. J. Biol. Chem. 2009, 284, 22825–22833. [Google Scholar] [CrossRef] [Green Version]
- Brungs, D.; Lynch, D.; Luk, A.W.; Minaei, E.; Ranson, M.; Aghmesheh, M.; Vine, K.L.; Carolan, M.; Jaber, M.; de Souza, P.; et al. Cryopreservation for delayed circulating tumor cell isolation is a valid strategy for prognostic association of circulating tumor cells in gastroesophageal cancer. World J. Gastroenterol. 2018, 24, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Xu, L.; Yu, S.; Hong, W.; Huang, M.; Xu, P. Therapeutics targeting the fibrinolytic system. Exp. Mol. Med. 2020, 52, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Guo, Z.; Yu, S.; Jiang, L.; Huang, M. Development of inhibitors for uPAR: Blocking the interaction of uPAR with its partners. Drug Discov. Today 2021, 26, 1076–1085. [Google Scholar] [CrossRef]
- Belfiore, L.; Saunders, D.N.; Ranson, M.; Vine, K.L. N-Alkylisatin-Loaded Liposomes Target the Urokinase Plasminogen Activator System in Breast Cancer. Pharmaceutics 2020, 12, 641. [Google Scholar] [CrossRef]
- Vine, K.L.; Lobov, S.; Indira Chandran, V.; Harris, N.L.; Ranson, M. Improved pharmacokinetic and biodistribution properties of the selective urokinase inhibitor PAI-2 (SerpinB2) by site-specific PEGylation: Implications for drug delivery. Pharm. Res. 2015, 32, 1045–1054. [Google Scholar] [CrossRef]
- Vine, K.L.; Indira Chandran, V.; Locke, J.M.; Matesic, L.; Lee, J.; Skropeta, D.; Bremner, J.B.; Ranson, M. Targeting urokinase and the transferrin receptor with novel, anti-mitotic N-alkylisatin cytotoxin conjugates causes selective cancer cell death and reduces tumor growth. Curr. Cancer Drug Targets 2012, 12, 64–73. [Google Scholar] [CrossRef]
- Stutchbury, T.K.; Al-Ejeh, F.; Stillfried, G.E.; Croucher, D.R.; Andrews, J.; Irving, D.; Links, M.; Ranson, M. Preclinical evaluation of 213Bi-labeled plasminogen activator inhibitor type 2 in an orthotopic murine xenogenic model of human breast carcinoma. Mol. Cancer Ther. 2007, 6, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Hang, M.T.N.; Ranson, M.; Saunders, D.N.; Liang, X.M.; Bunn, C.L.; Baker, M.S. Pharmacokinetics and biodistribution of recombinant human plasminogen activator inhibitor type 2 (PAI-2) in control and tumour xenograft-bearing mice. Fibrinolysis Proteolysis 1998, 12, 145–154. [Google Scholar] [CrossRef]
- Buckley, B.J.; Aboelela, A.; Minaei, E.; Jiang, L.X.; Xu, Z.; Ali, U.; Fildes, K.; Cheung, C.Y.; Cook, S.M.; Johnson, D.C.; et al. 6-Substituted Hexamethylene Amiloride (HMA) Derivatives as Potent and Selective Inhibitors of the Human Urokinase Plasminogen Activator for Use in Cancer. J. Med. Chem. 2018, 61, 8299–8320. [Google Scholar] [CrossRef]
- Buckley, B.J.; Majed, H.; Aboelela, A.; Minaei, E.; Jiang, L.; Fildes, K.; Cheung, C.Y.; Johnson, D.; Bachovchin, D.; Cook, G.M.; et al. 6-Substituted amiloride derivatives as inhibitors of the urokinase-type plasminogen activator for use in metastatic disease. Bioorg. Med. Chem. Lett. 2019, 29, 126753. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, C. Quercetin Has Antimetastatic Effects on Gastric Cancer Cells via the Interruption of uPA/uPAR Function by Modulating NF-κb, PKC-δ, ERK1/2; AMPKα. Integr. Cancer Ther. 2017, 17, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Zhang, H.; Zhou, Z.; Zhong, M.; Chen, Q.; Wang, X.; Zhu, Z. u-PA inhibitor amiloride suppresses peritoneal metastasis in gastric cancer. World J. Surg. Oncol. 2012, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinemann, V.; Ebert, M.P.; Laubender, R.P.; Bevan, P.; Mala, C.; Boeck, S. Phase II randomised proof-of-concept study of the urokinase inhibitor upamostat (WX-671) in combination with gemcitabine compared with gemcitabine alone in patients with non-resectable, locally advanced pancreatic cancer. Br. J. Cancer 2013, 108, 766–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
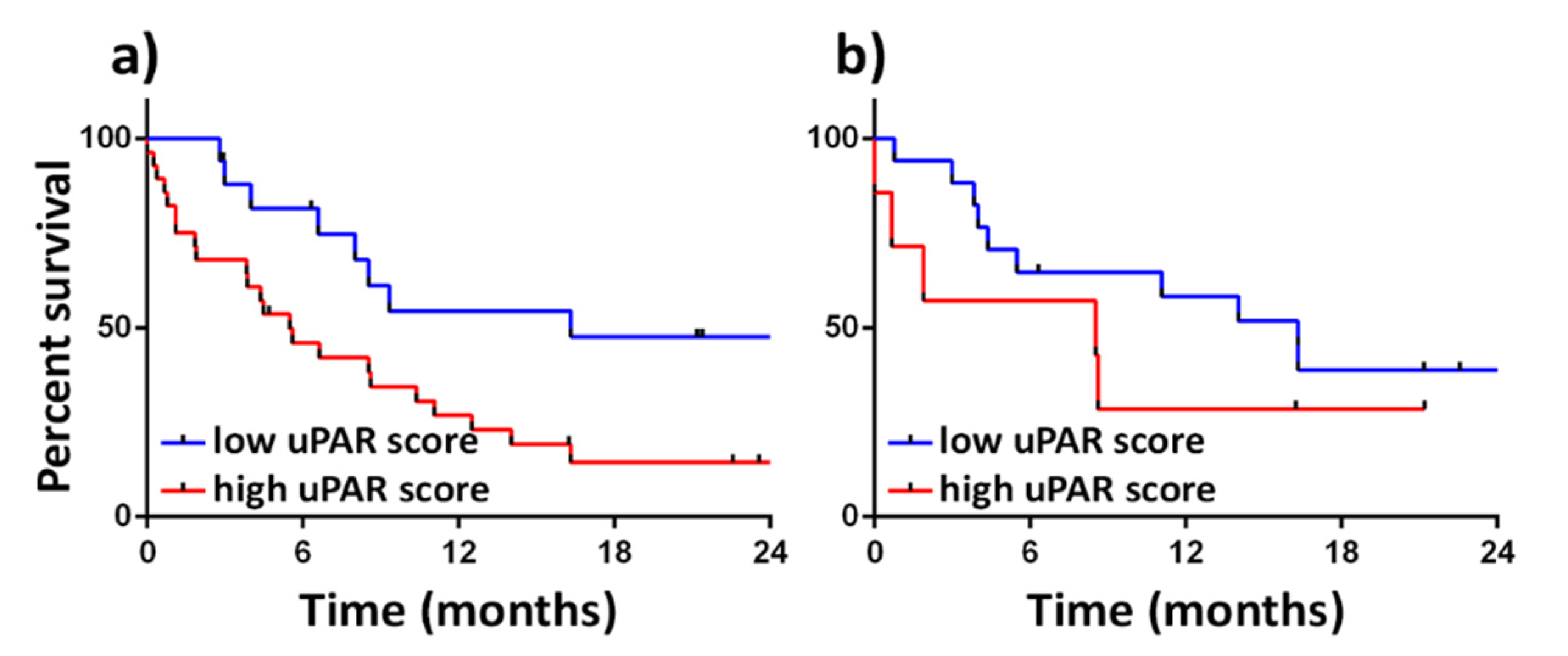{kind=link}
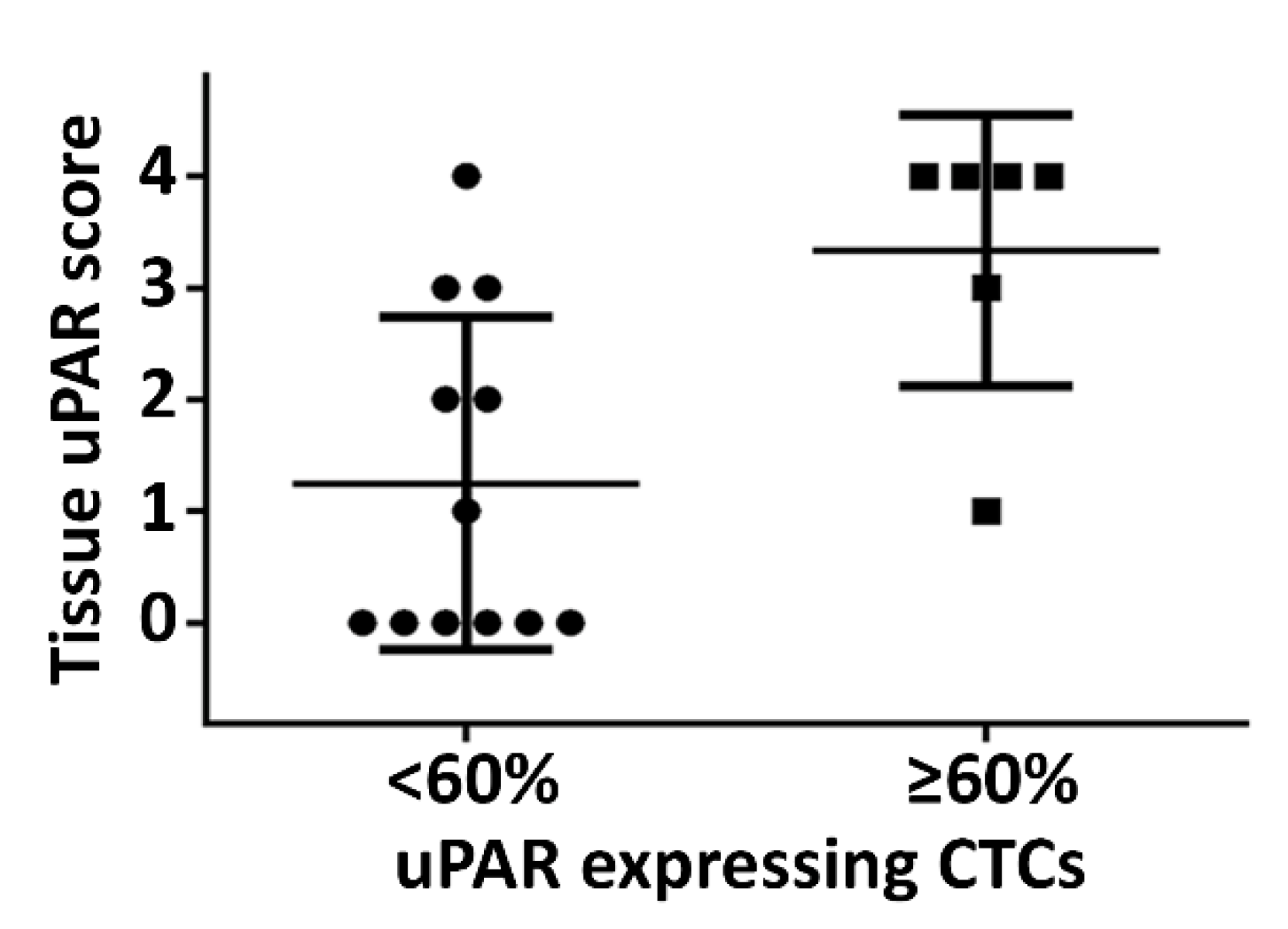{kind=link}
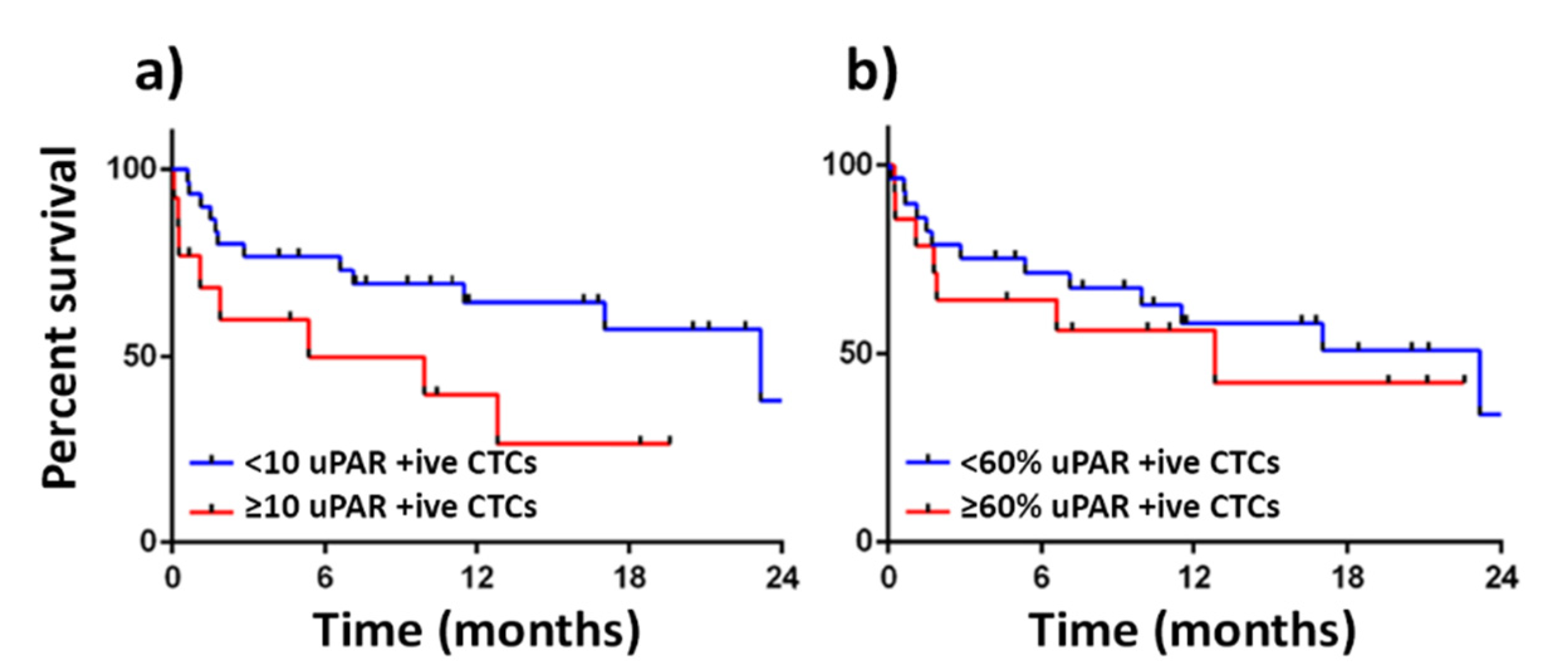{kind=link}
| Molecule, Pathway | Derived Cell Line 1 | Effect on uPAS | Reference |
|---|---|---|---|
| Upregulators | |||
| Epidermal Growth Factor (EGF) Extracellular cell signalling transducer | SGC-7901 BGC-823 | Exogenous EGF increased uPAR mRNA Blockade of ERK1/2 reduced uPAR mRNA expression | Wang, P., et al., 2017 [72] |
| Reduced uPA mRNA seen in ERK blockade | Wang, J., et al., 2016 [68] | ||
| Osteopontin Linked to PI3K/NFkB/IKK pathways | SGC-7901 BGC-823 | siRNA against Osteopontin resulted in reduced uPA mRNA levels. Xenograft model showing reduced tumour growth | Gong, M., et al., 2008 [69] |
| Hepatocyte Growth Factor (HGF)/cMET pathway Extracellular cell signalling transducer | NUGC-3 MKN-28 | Exogenous HGF exposure increased uPA and uPAR protein levels. Blockade of uPAR with antibody or siRNA resulted in reduced wound invasion, which could not be overcome with exogenous HGF. | Kyung Hee, L., et al., 2006 [73] |
| HGF signal transduction occurs via JunB/survivin pathway. Survivin inhibition resulted in reduced uPA protein expression and reduced cell invasion. | Kyung Hee, L., et al., 2011 [78] | ||
| MEK inhibition resulted in reduced uPA protein levels, whilst PI3K inhibition showed no change in uPA level. Suggesting uPA activation by HGF via ERK pathway. | Lee, K., et al., 2014 [70] | ||
| HGF signal transduction via PKC/PKD pathway can release HDAC5; HDAC5 increased uPA and MMP-9 activity. Blockade of HDAC5 (even in presence of exogenous HGF) resulted in reduced uPA protein levels. HDAC5-inhibited cells showed reduced cell invasion. | Lee, K., et al., 2010 [66] | ||
| COX-PGE2 pathway | AGS | Exogenous prostaglandin E2 resulted in increased levels of uPA and uPAR (protein and mRNA). | Lian, S., et al., 2017 [74] |
| Nicotine exposure increased PGE2 resulting in increased uPA and uPAR protein expression | Shin, V., et al., 2005 [75] | ||
| Laminin receptor (67LR) | SGC-7901 MKN-45 | Downregulation of 67LR resulted in reduced cell line uPA protein expression. | Liu, L., et al., 2010 [79] |
| TGF-β pathway | SNU-216 | Exogenous macrophage inhibitory cytokine 1 (MIC-1; a member of the TGF-β superfamily) resulted in increased uPA and uPAR (mRNA and protein); PAI-1 (mRNA) unaltered. | Lee, D., et al., 2003 [76] |
| Interferon gamma inhibition resulted in TGF-B downregulation via smad 2/3 pathway with downregulation of uPA protein expression. | Kuga, H., et al., 2003 [67] | ||
| OE33 FLOW | Increased PAI-1 mRNA levels on exposure via downstream activation of PI3K, ERK and JNK pathways on TGF-β activation. | Onwuegbusi, B., et al., 2007 [71] | |
| Downregulators | |||
| p75NTR, NF-κB signalling pathway | SGC7901 MKN45 | Upregulation of p75NTR protein caused reduced protein levels of uPA. | Jin, H., et al., 2005 [80] |
| Tspan9, ERK1/2 pathway | SGC7901 | Reduced protein levels of uPA through ERK1/2 blockade. | Li, P. et al., 2016 [77] |
| Clinicopathological Feature | uPA | uPAR | PAI-1 |
|---|---|---|---|
| Key References | |||
| T stage | [22,23,24] | [21,23,26] | [21,23,24,25,26] |
| Lymph nodes | [22,23,24] | [21,23,26] | [21,23,24,25,26] |
| Distant metastasis | [22,23,24] | [24] | |
| Vascular invasion | [22,23,24] | [21,23,26] | [21,23,25,26] |
| Lymphatic invasion | [22,23,24] | [21,23,26] | [21,23,25,26] |
| Peritoneal disease 1 | [22,23] | ||
| Serosal involvement 1 | [22,23] | ||
| Depth of invasion | [21,23,26] | [21,23,24,25,26] | |
| Histological grade | [21,23,26] | [24] | |
| Recurrence-Free Survival HR (95% CI) | Overall Survival HR (95% CI) | |
|---|---|---|
| uPA | 1.90 (1.16–3.11, p = 0.01) 3 studies, 467 patients | 2.21 (1.74–2.80, p < 0.0001) 12 studies, 1094 patients |
| uPAR | 2.69 (NR, p = 0.03) 1 study, 203 patients | 2.19 (1.80–2.66, p < 0.0001) 11 studies, 1036 patients |
| PAI-1 | 1.96 (1.07–3.58, p = 0.03) 3 studies, 467 patients | 1.84 (1.28–2.64, p < 0.0001) 10 studies, 839 patients |
| PAI-2 | NR no studies | 0.97 (0.48–1.94, p = 0.92) 2 studies, 145 patients |
| Tissue | Cell Type | uPA | uPAR | PAI-1 | PAI-2 |
|---|---|---|---|---|---|
| Breast, ductal | Macrophages | + [98] | + [99,100] | + [101] | |
| Fibroblasts | + [98,102] | Weak [102] | Weak [102] | ||
| Colon | Macrophages | + [103] | |||
| Fibroblasts | + [104] | + [103] | |||
| Lung | Macrophages | + [105] | + [105] | + [105] | |
| Fibroblasts |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tincknell, G.; Piper, A.-K.; Aghmesheh, M.; Becker, T.; Vine, K.L.; Brungs, D.; Ranson, M. Experimental and Clinical Evidence Supports the Use of Urokinase Plasminogen Activation System Components as Clinically Relevant Biomarkers in Gastroesophageal Adenocarcinoma. Cancers 2021, 13, 4097. https://doi.org/10.3390/cancers13164097
Tincknell G, Piper A-K, Aghmesheh M, Becker T, Vine KL, Brungs D, Ranson M. Experimental and Clinical Evidence Supports the Use of Urokinase Plasminogen Activation System Components as Clinically Relevant Biomarkers in Gastroesophageal Adenocarcinoma. Cancers. 2021; 13(16):4097. https://doi.org/10.3390/cancers13164097
Chicago/Turabian StyleTincknell, Gary, Ann-Katrin Piper, Morteza Aghmesheh, Therese Becker, Kara Lea Vine, Daniel Brungs, and Marie Ranson. 2021. "Experimental and Clinical Evidence Supports the Use of Urokinase Plasminogen Activation System Components as Clinically Relevant Biomarkers in Gastroesophageal Adenocarcinoma" Cancers 13, no. 16: 4097. https://doi.org/10.3390/cancers13164097