
Transcriptome Profiling and Metagenomic Analysis Help to Elucidate Interactions in an Inflammation-Associated Cancer Mouse Model


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. DSS/AOM-Induced Enteritis/Carcinogenesis Mouse Model
2.2. Ion Torrent Sequencing Using the AmpliSeq Transcriptome Gene Expression Assay and Transcriptome Analysis
2.3. Gene Selection, Pathway Analysis and Gene Set Enrichment Analysis (GSEA)
2.4. 16S Metagenomic Sequencing and Analysis
2.5. Immunohistochemical Staining
3. Results
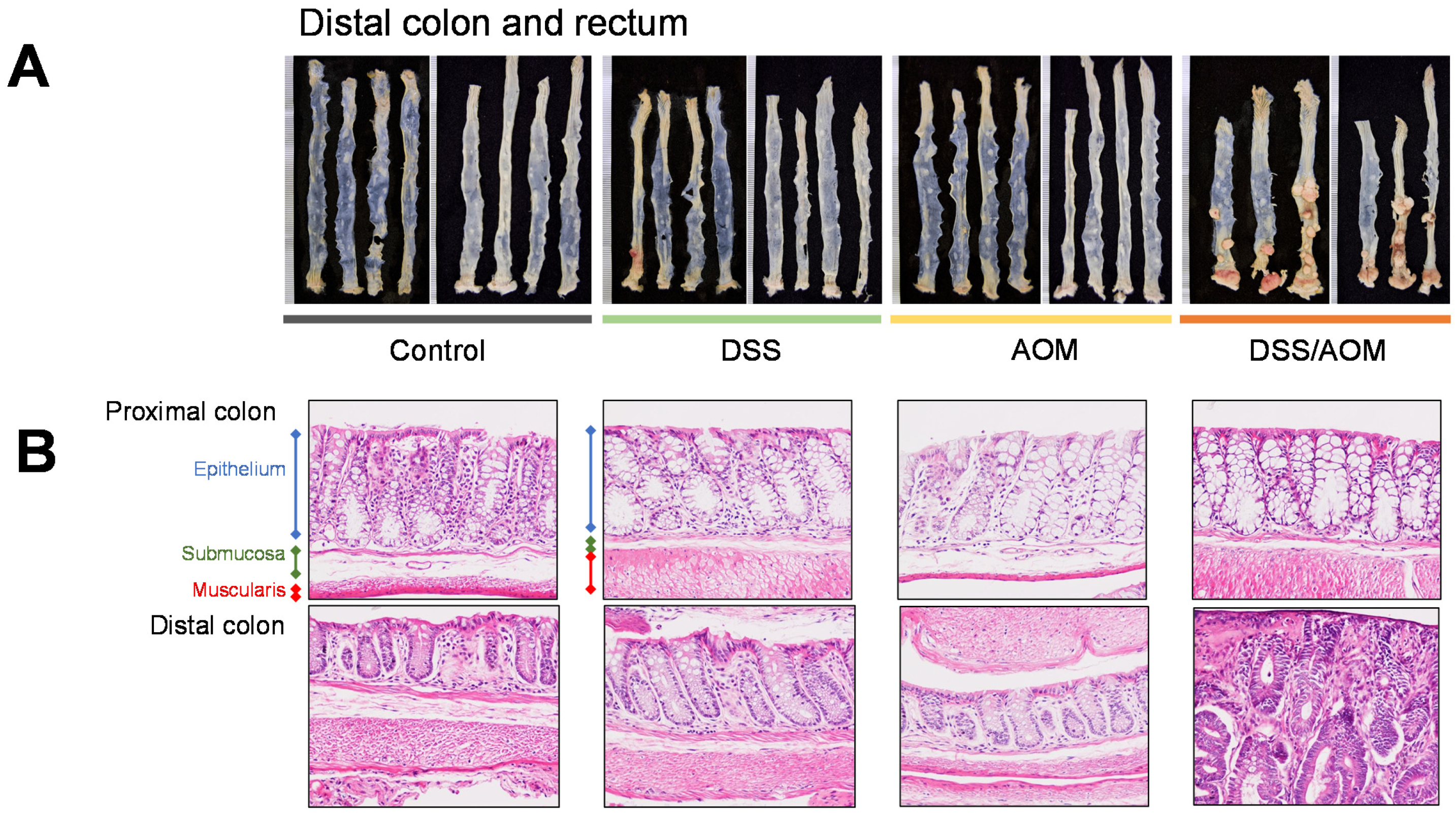
3.1. Evaluation of the DSS/AOM-Induced Colon Cancer Model
3.2. Whole Transcriptome Profiling
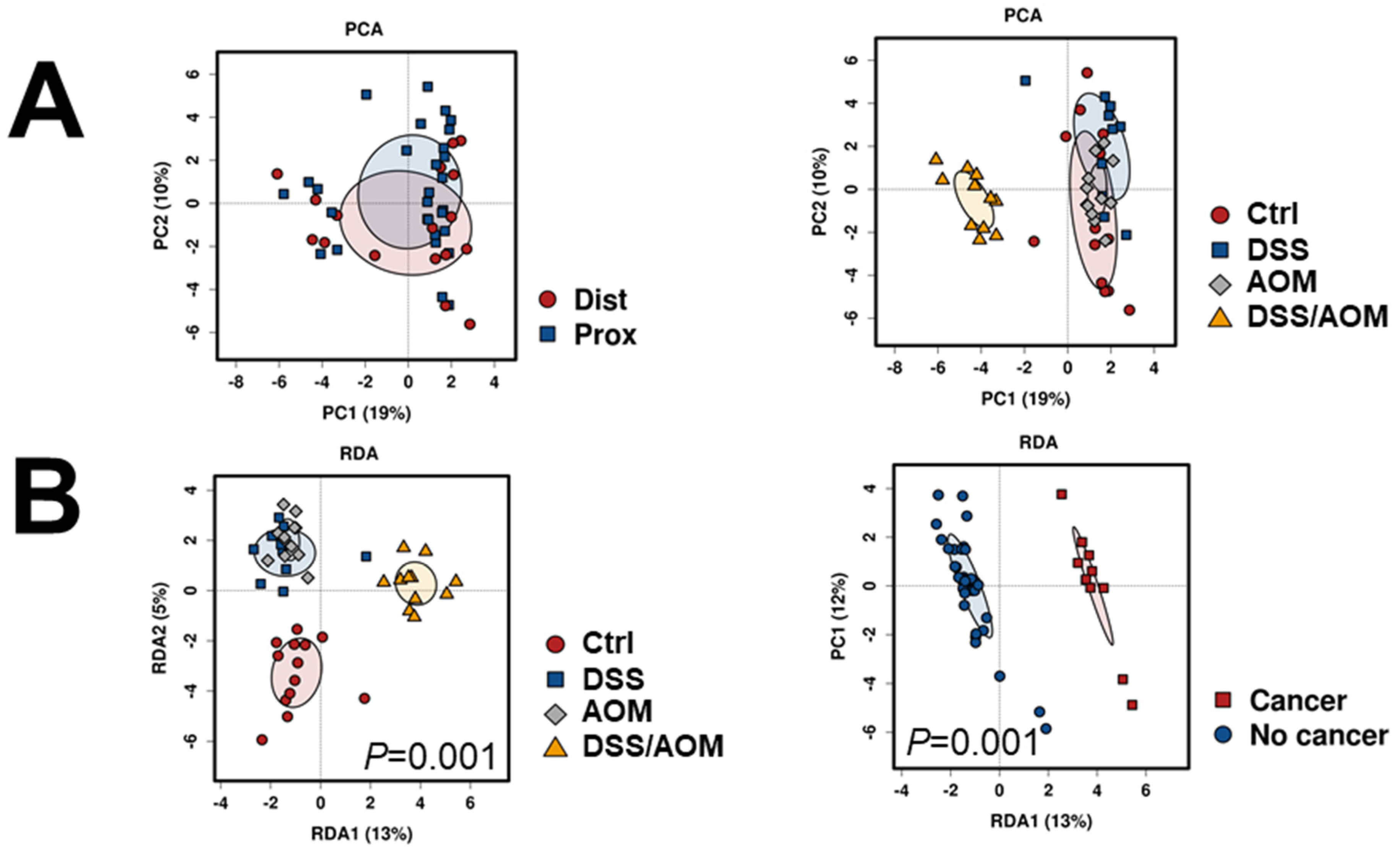
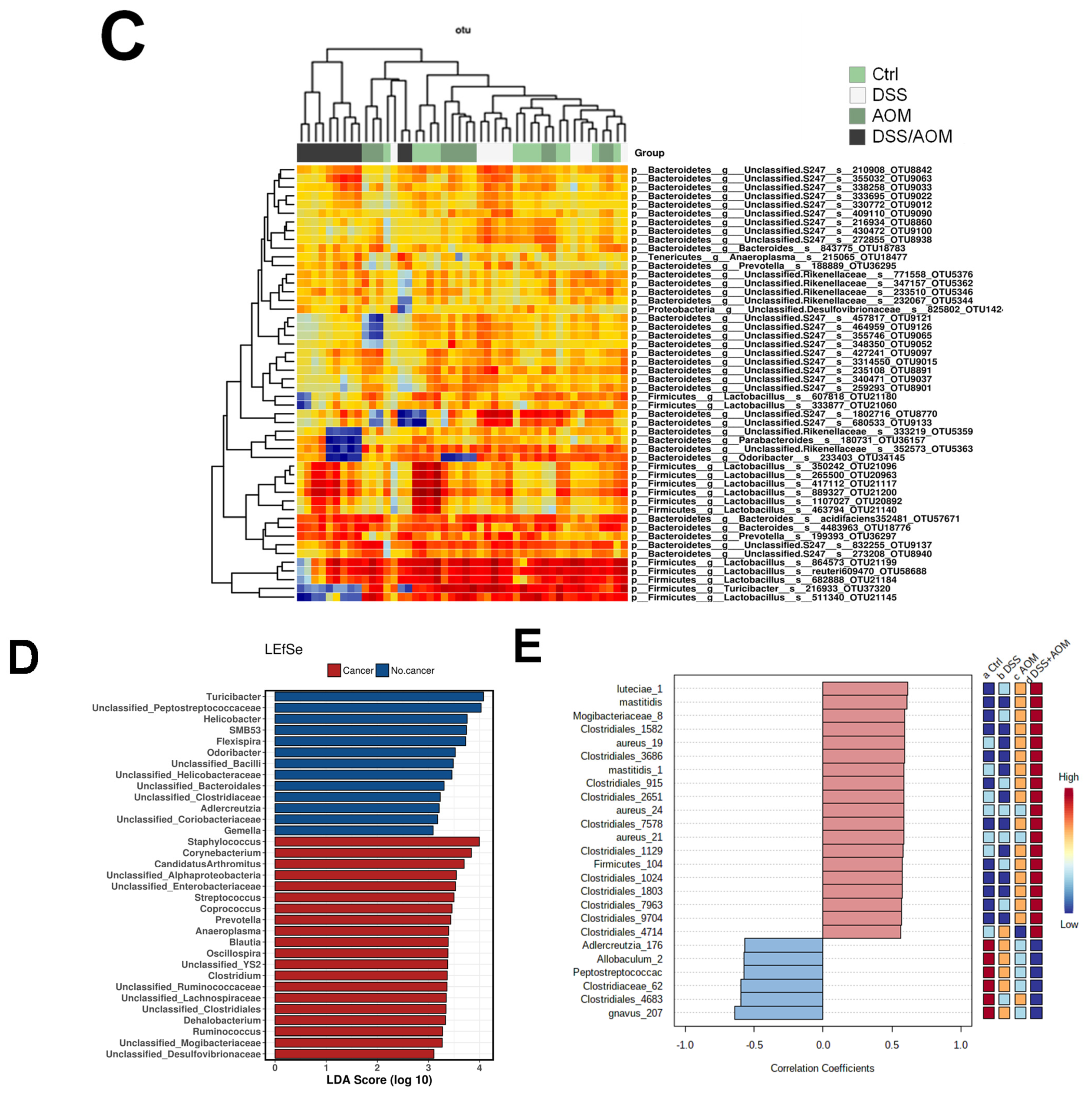
3.3. 16S Metagenomic Sequencing
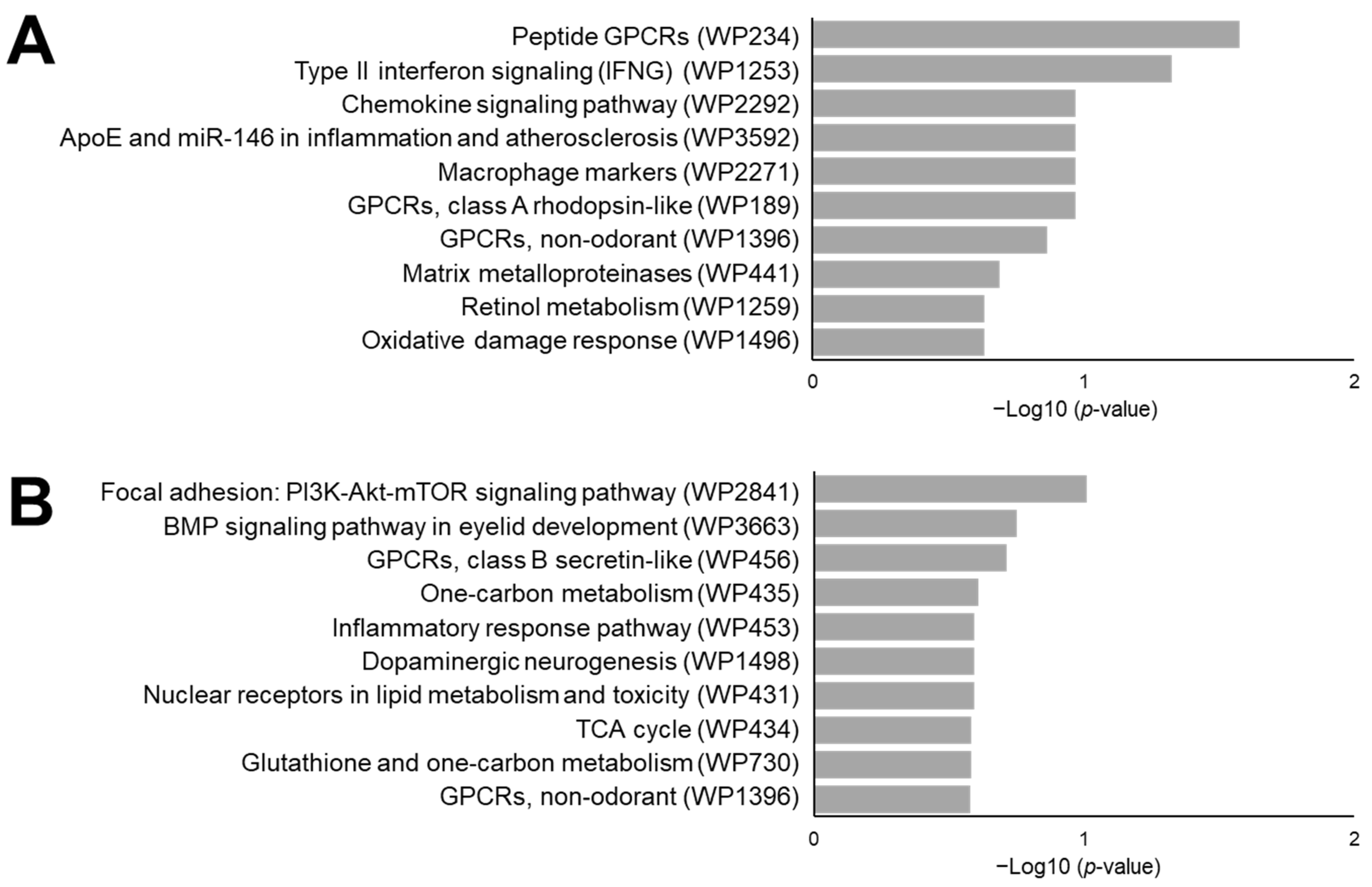
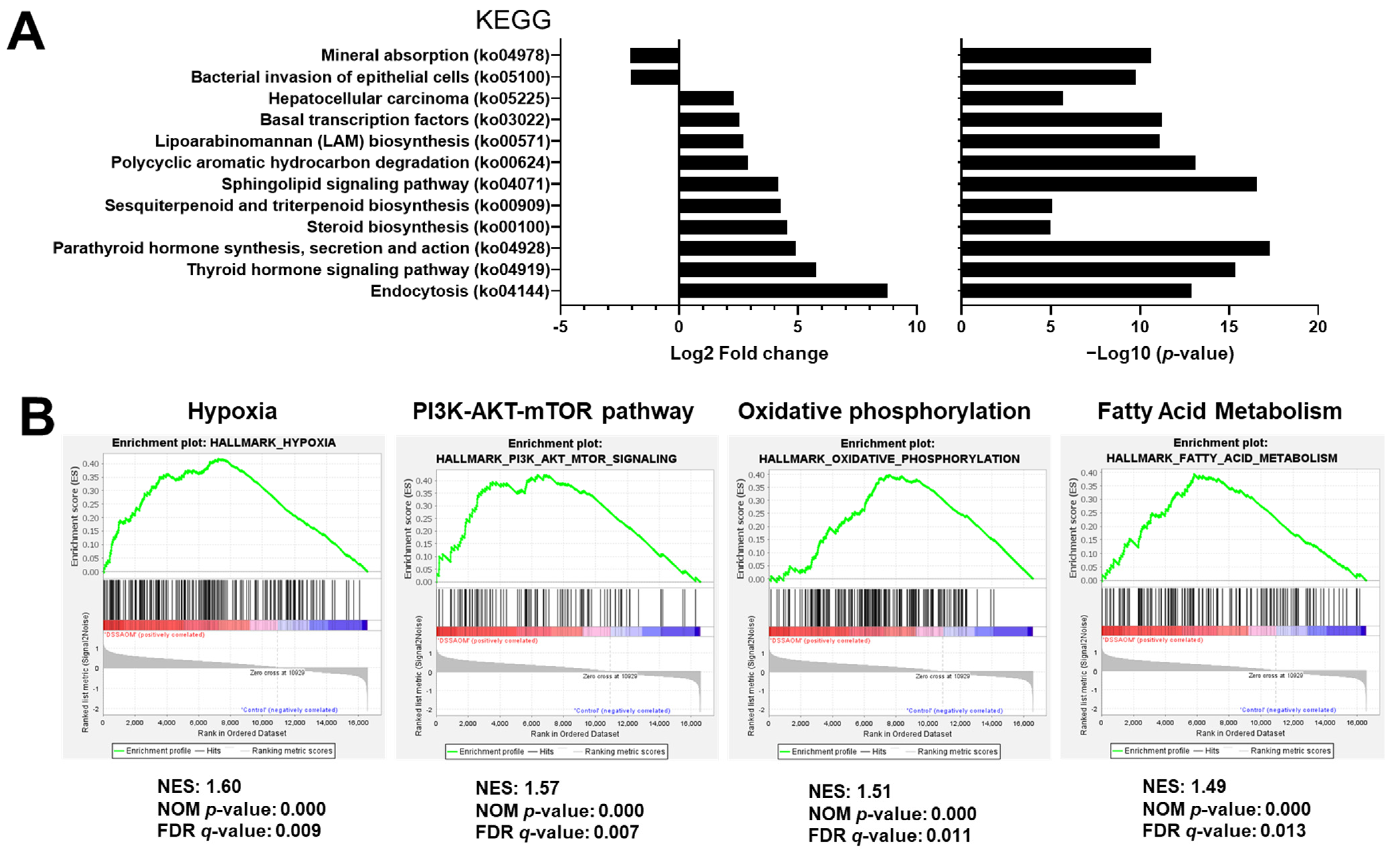
3.4. Functional Prediction Analysis Comparing Inflammation Induced Colon Cancer and Normal Mucosa
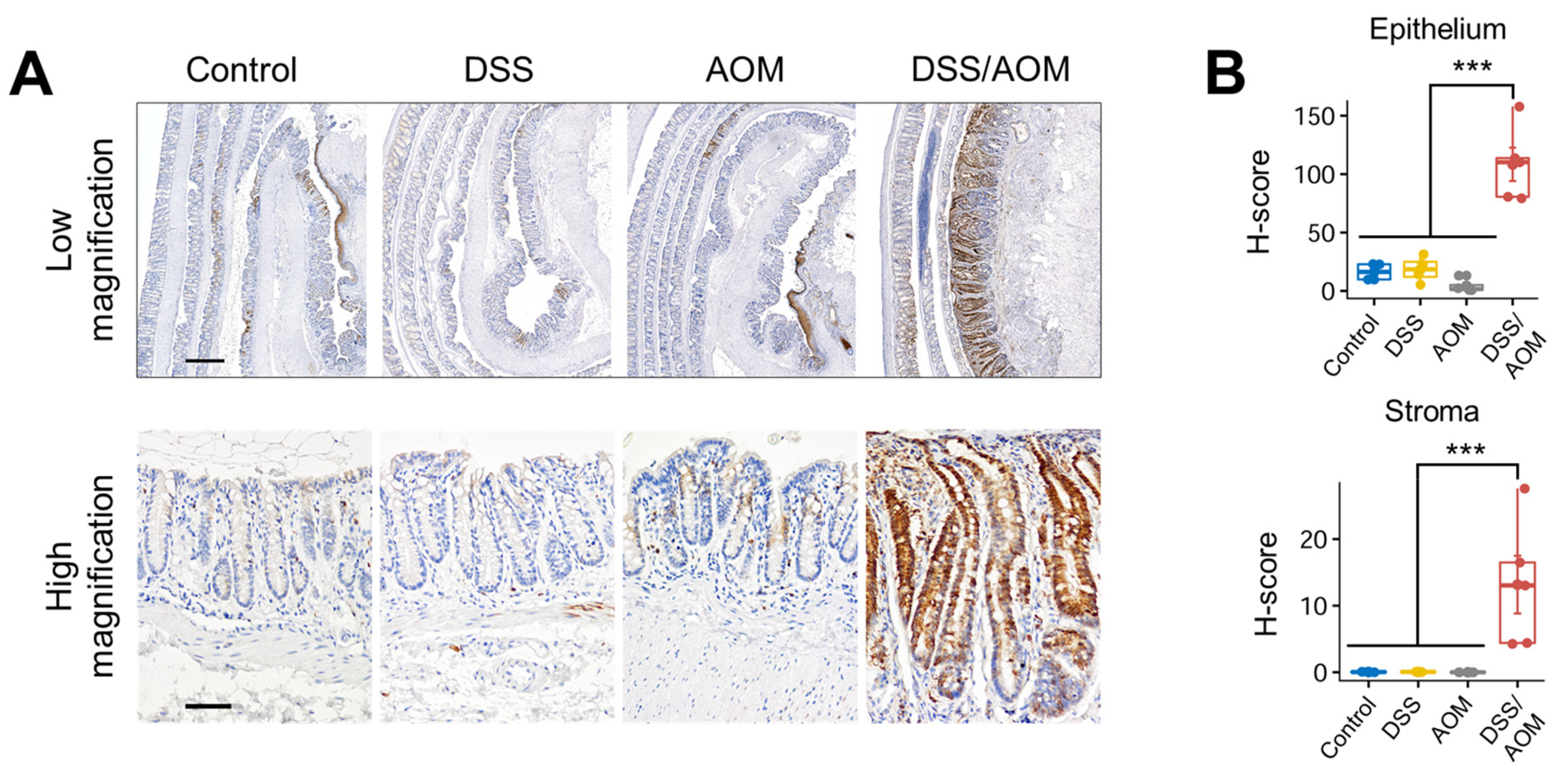
3.5. Immunohistochemical Staining of Phosphorylated-S6 Ribosomal Protein
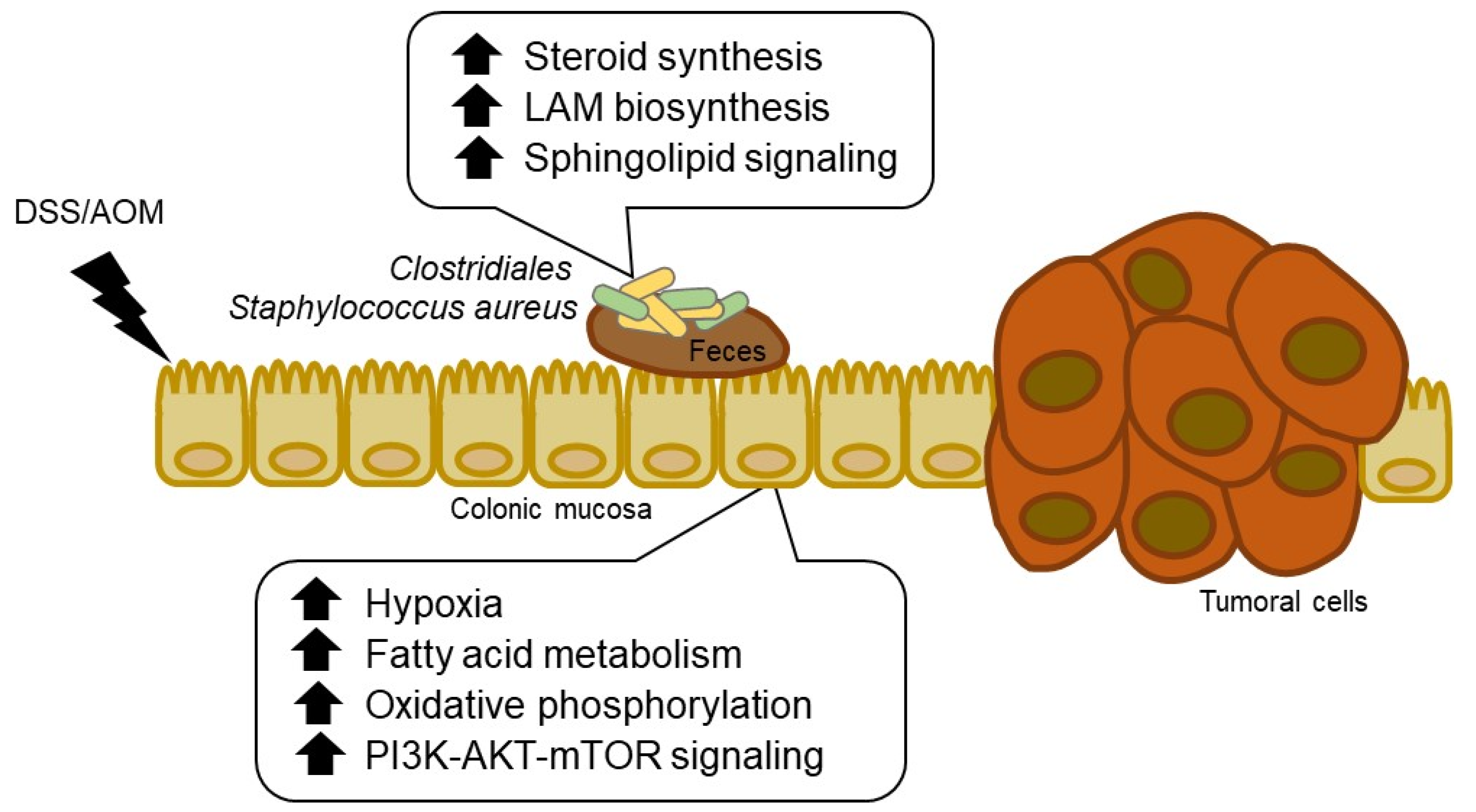
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bakken, T.; Braaten, T.; Olsen, A.; Hjartaker, A.; Lund, E.; Skeie, G. Milk and risk of colorectal, colon and rectal cancer in the Norwegian Women and Cancer (NOWAC) Cohort Study. Br. J. Nutr. 2018, 119, 1274–1285. [Google Scholar] [CrossRef] [Green Version]
- Hemeryck, L.Y.; Rombouts, C.; Hecke, T.V.; Van Meulebroek, L.; Bussche, J.V.; De Smet, S.; Vanhaecke, L. In vitro DNA adduct profiling to mechanistically link red meat consumption to colon cancer promotion. Toxicol. Res. 2016, 5, 1346–1358. [Google Scholar] [CrossRef] [Green Version]
- Eaden, J.A.; Abrams, K.R.; Mayberry, J.F. The risk of colorectal cancer in ulcerative colitis: A meta-analysis. Gut 2001, 48, 526–535. [Google Scholar] [CrossRef] [Green Version]
- Jess, T.; Rungoe, C.; Peyrin-Biroulet, L. Risk of colorectal cancer in patients with ulcerative colitis: A meta-analysis of population-based cohort studies. Clin. Gastroenterol. Hepatol. 2012, 10, 639–645. [Google Scholar] [CrossRef]
- Zhou, Q.; Shen, Z.F.; Wu, B.S.; Xu, C.B.; He, Z.Q.; Chen, T.; Shang, H.T.; Xie, C.F.; Huang, S.Y.; Chen, Y.G.; et al. Risk of Colorectal Cancer in Ulcerative Colitis Patients: A Systematic Review and Meta-Analysis. Gastroenterol. Res. Pract. 2019, 2019, 5363261. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Kian, R.; Moradi, S.; Ghorbian, S. Role of components of microRNA machinery in carcinogenesis. Exp. Oncol. 2018, 40, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Cortez, M.A.; Anfossi, S.; Ramapriyan, R.; Menon, H.; Atalar, S.C.; Aliru, M.; Welsh, J.; Calin, G.A. Role of miRNAs in immune responses and immunotherapy in cancer. Genes Chromosomes Cancer 2019, 58, 244–253. [Google Scholar] [CrossRef]
- Han, J.; Liu, Z.; Wang, N.; Pan, W. MicroRNA-874 inhibits growth, induces apoptosis and reverses chemoresistance in colorectal cancer by targeting X-linked inhibitor of apoptosis protein. Oncol. Rep. 2016, 36, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ju, H.; Zhang, L.; Lu, H.; Jie, K. microRNA-577 suppresses tumor growth and enhances chemosensitivity in colorectal cancer. J. Biochem. Mol. Toxicol. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Zhu, H.F.; Zhang, Z.Y.; Ren, F.; Hu, Y.H. MiR-384 inhibits the proliferation of colorectal cancer by targeting AKT3. Cancer Cell Int. 2018, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; Malesci, A.; Vetrano, S. Colitis-associated cancer: The dark side of inflammatory bowel disease. Gut 2011, 60, 1609–1610. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; Mantovani, A. Inflammatory bowel disease and intestinal cancer: A paradigm of the Yin-Yang interplay between inflammation and cancer. Oncogene 2010, 29, 3313–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzic, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Fumery, M.; Dulai, P.S.; Gupta, S.; Prokop, L.J.; Ramamoorthy, S.; Sandborn, W.J.; Singh, S. Incidence, Risk Factors, and Outcomes of Colorectal Cancer in Patients With Ulcerative Colitis With Low-Grade Dysplasia: A Systematic Review and Meta-analysis. Clin. Gastroenterol. Hepatol. 2017, 15, 665–674.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakatos, L.; Mester, G.; Erdelyi, Z.; David, G.; Pandur, T.; Balogh, M.; Fischer, S.; Vargha, P.; Lakatos, P.L. Risk factors for ulcerative colitis-associated colorectal cancer in a Hungarian cohort of patients with ulcerative colitis: Results of a population-based study. Inflamm. Bowel. Dis. 2006, 12, 205–211. [Google Scholar] [CrossRef]
- Thaker, A.I.; Shaker, A.; Rao, M.S.; Ciorba, M.A. Modeling colitis-associated cancer with azoxymethane (AOM) and dextran sulfate sodium (DSS). J. Vis. Exp. 2012. [Google Scholar] [CrossRef] [Green Version]
- Demsar, J.; Curk, T.; Erjavec, A.; Gorup, C.; Hocevar, T.; Milutinovic, M.; Mozina, M.; Polajnar, M.; Toplak, M.; Staric, A.; et al. Orange: Data Mining Toolbox in Python. J. Mach. Learn Res. 2013, 14, 2349–2353. [Google Scholar]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; Miller, R.A.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting communities. Nucleic Acids Res. 2021, 49, D613–D621. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.N.; Wang, Z.C.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstrale, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahan, M.E.; Bowman, S.; Chevalier, R.; Rossi, R.; Nelson, M.; Roffey, P.; Xu, B.; Power, D.; McNevin, D. Bacillus species at the Canberra Airport: A comparison of real-time polymerase chain reaction and massively parallel sequencing for identification. Forensic. Sci. Int. 2019, 295, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome-environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csala, A.; Hof, M.H.; Zwinderman, A.H. Multiset sparse redundancy analysis for high-dimensional omics data. Biometrical J. 2019, 61, 406–423. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Iwai, S.; Weinmaier, T.; Schmidt, B.L.; Albertson, D.G.; Poloso, N.J.; Dabbagh, K.; DeSantis, T.Z. Piphillin: Improved Prediction of Metagenomic Content by Direct Inference from Human Microbiomes. PLoS ONE 2016, 11, e0166104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.; Torrelles, J.B. Mannose-capped lipoarabinomannan in Mycobacterium tuberculosis pathogenesis. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Dzik, K.P.; Kaczor, J.J. Mechanisms of vitamin D on skeletal muscle function: Oxidative stress, energy metabolism and anabolic state. Eur. J. Appl. Physiol. 2019, 119, 825–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, A.F.; Sande, O.J.; Tomechko, S.E.; Ding, X.; Li, M.; Maxwell, S.; Ewing, R.M.; Harding, C.V.; Rojas, R.E.; Chance, M.R.; et al. Proteomics and Network Analyses Reveal Inhibition of Akt-mTOR Signaling in CD4(+) T Cells by Mycobacterium tuberculosis Mannose-Capped Lipoarabinomannan. Proteomics 2017, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, T.; Hirayama, D.; Wagatsuma, K.; Yamakawa, T.; Yokoyama, Y.; Nakase, H. Immunological Mechanisms in Inflammation-Associated Colon Carcinogenesis. Int. J. Mol. Sci. 2020, 21, 3062. [Google Scholar] [CrossRef] [PubMed]
- Swafford, D.; Shanmugam, A.; Ranganathan, P.; Manoharan, I.; Hussein, M.S.; Patel, N.; Sifuentes, H.; Koni, P.A.; Prasad, P.D.; Thangaraju, M.; et al. The Wnt-beta-Catenin-IL-10 Signaling Axis in Intestinal APCs Protects Mice from Colitis-Associated Colon Cancer in Response to Gut Microbiota. J. Immunol. 2020, 205, 2265–2275. [Google Scholar] [CrossRef]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fecteau, M.E.; Pitta, D.W.; Vecchiarelli, B.; Indugu, N.; Kumar, S.; Gallagher, S.C.; Fyock, T.L.; Sweeney, R.W. Dysbiosis of the Fecal Microbiota in Cattle Infected with Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2016, 11, e0160353. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermudez-Humaran, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [Green Version]
- Simren, M.; Barbara, G.; Flint, H.J.; Spiegel, B.M.; Spiller, R.C.; Vanner, S.; Verdu, E.F.; Whorwell, P.J.; Zoetendal, E.G.; Rome Foundation, C. Intestinal microbiota in functional bowel disorders: A Rome foundation report. Gut 2013, 62, 159–176. [Google Scholar] [CrossRef]
- Babolewska, E.; Witczak, P.; Pietrzak, A.; Brzezinska-Blaszczyk, E. Different potency of bacterial antigens TLR2 and TLR4 ligands in stimulating mature mast cells to cysteinyl leukotriene synthesis. Microbiol. Immunol. 2012, 56, 183–190. [Google Scholar] [CrossRef]
- Ma, J.; Gulbins, E.; Edwards, M.J.; Caldwell, C.C.; Fraunholz, M.; Becker, K.A. Staphylococcus aureus alpha-Toxin Induces Inflammatory Cytokines via Lysosomal Acid Sphingomyelinase and Ceramides. Cell Physiol. Biochem. 2017, 43, 2170–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glaser, U.G.; Fandrey, J. Sphingolipids in inflammatory hypoxia. Biol Chem 2018, 399, 1169–1174. [Google Scholar] [CrossRef] [PubMed]
- Obinata, H.; Hla, T. Sphingosine 1-phosphate and inflammation. Int. Immunol. 2019, 31, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Maiti, D.; Bhattacharyya, A.; Basu, J. Lipoarabinomannan from Mycobacterium tuberculosis promotes macrophage survival by phosphorylating Bad through a phosphatidylinositol 3-kinase/Akt pathway. J. Biol. Chem. 2001, 276, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Goude, R.; Amin, A.G.; Chatterjee, D.; Parish, T. The critical role of embC in Mycobacterium tuberculosis. J. Bacteriol. 2008, 190, 4335–4341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallenius, G.; Correia-Neves, M.; Buteme, H.; Hamasur, B.; Svenson, S.B. Lipoarabinomannan, and its related glycolipids, induce divergent and opposing immune responses to Mycobacterium tuberculosis depending on structural diversity and experimental variations. Tuberculosis 2016, 96, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Rausch, P.; Ruhlemann, M.; Hermes, B.M.; Doms, S.; Dagan, T.; Dierking, K.; Domin, H.; Fraune, S.; von Frieling, J.; Hentschel, U.; et al. Comparative analysis of amplicon and metagenomic sequencing methods reveals key features in the evolution of animal metaorganisms. Microbiome 2019, 7, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MSigDB Hallmark Pathway | NES | NOM p-Value | FDR q-Value |
|---|---|---|---|
| Protein secretion | 1.75 | 0 | 0.005 |
| TGF-β signaling | 1.73 | 0.001 | 0.003 |
| mTORC1 signaling | 1.7 | 0 | 0.005 |
| Cholesterol homeostasis | 1.68 | 0.001 | 0.006 |
| G2M checkpoint | 1.67 | 0 | 0.005 |
| UV response down-regulated | 1.6 | 0 | 0.01 |
| Hypoxia | 1.6 | 0 | 0.009 |
| E2F targets | 1.58 | 0 | 0.009 |
| Glycolysis | 1.57 | 0 | 0.009 |
| Hedgehog signaling | 1.57 | 0.008 | 0.008 |
| PI3K-Akt-mTOR signaling | 1.57 | 0 | 0.007 |
| Mitotic spindle | 1.55 | 0 | 0.009 |
| Interferon-α response | 1.54 | 0.004 | 0.009 |
| Oxidative phosphorylation | 1.51 | 0 | 0.011 |
| Reactive oxygen species pathway | 1.51 | 0.008 | 0.011 |
| Epithelial mesenchymal transition | 1.5 | 0 | 0.012 |
| Fatty acid metabolism | 1.49 | 0 | 0.013 |
| UV response up-regulated | 1.43 | 0.002 | 0.025 |
| Interferon-γ response | 1.43 | 0.002 | 0.024 |
| Androgen response | 1.42 | 0.013 | 0.026 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakai, K.; De Velasco, M.A.; Kura, Y.; Nishio, K. Transcriptome Profiling and Metagenomic Analysis Help to Elucidate Interactions in an Inflammation-Associated Cancer Mouse Model. Cancers 2021, 13, 3683. https://doi.org/10.3390/cancers13153683
Sakai K, De Velasco MA, Kura Y, Nishio K. Transcriptome Profiling and Metagenomic Analysis Help to Elucidate Interactions in an Inflammation-Associated Cancer Mouse Model. Cancers. 2021; 13(15):3683. https://doi.org/10.3390/cancers13153683
Chicago/Turabian StyleSakai, Kazuko, Marco A. De Velasco, Yurie Kura, and Kazuto Nishio. 2021. "Transcriptome Profiling and Metagenomic Analysis Help to Elucidate Interactions in an Inflammation-Associated Cancer Mouse Model" Cancers 13, no. 15: 3683. https://doi.org/10.3390/cancers13153683