
The CRISPR/Cas9 Minipig—A Transgenic Minipig to Produce Specific Mutations in Designated Tissues

 ,
,
Abstract
:Simple Summary
Abstract
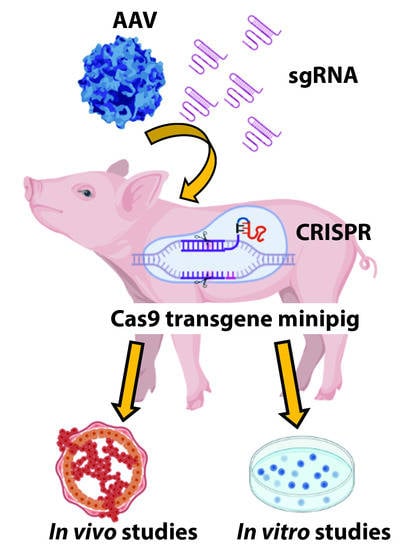
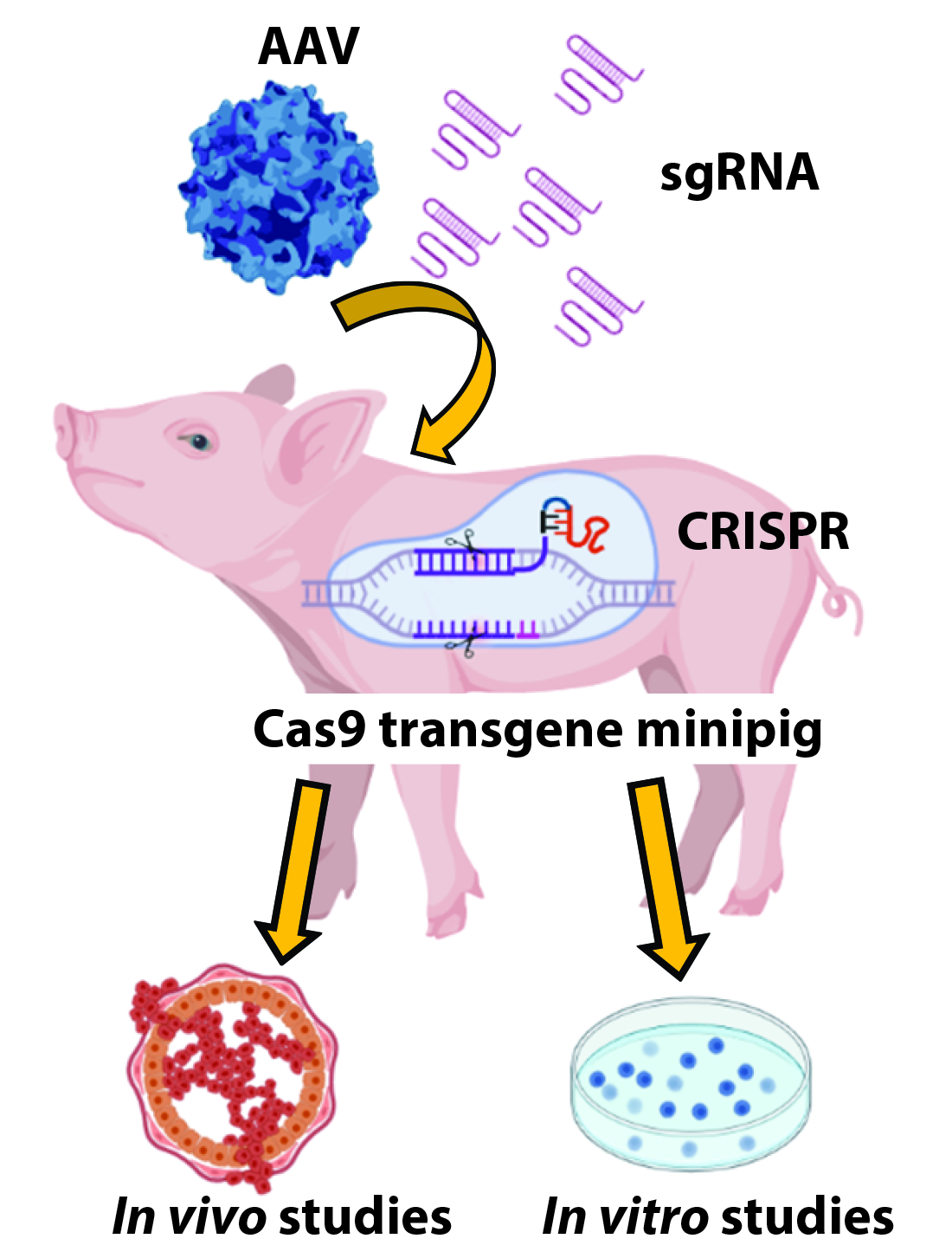{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vector Construction
2.3. Single Guide RNA (sgRNA) Design and Validation
2.4. ICE Analysis
2.5. Handmade Cloning (HMC), Culture and Transfer of Embryos
2.6. Cell Work
2.7. Air–Liquid Interface (ALI) Culture of Porcine Tracheobronchial Epithelial Cells
2.8. In Vivo Imaging Systems Analysis
2.9. Fluorescence-Assisted Cell Sorting
2.10. Southern Blot Analysis
2.11. AAV Production
2.12. AAV Delivery to Pig Lungs
2.13. Construct Delivery to the Skin of the Pigs
2.14. Β-galactosidase Staining
2.15. Western Blotting
2.16. Histochemical Analysis
2.17. Real-Time Quantitative PCR
2.18. PCR
2.19. 18F-Fluorodeoxyglucose Positron Emission Tomography–Computed Tomograph (PET/CT) Scanning
2.20. Statistics
3. Results
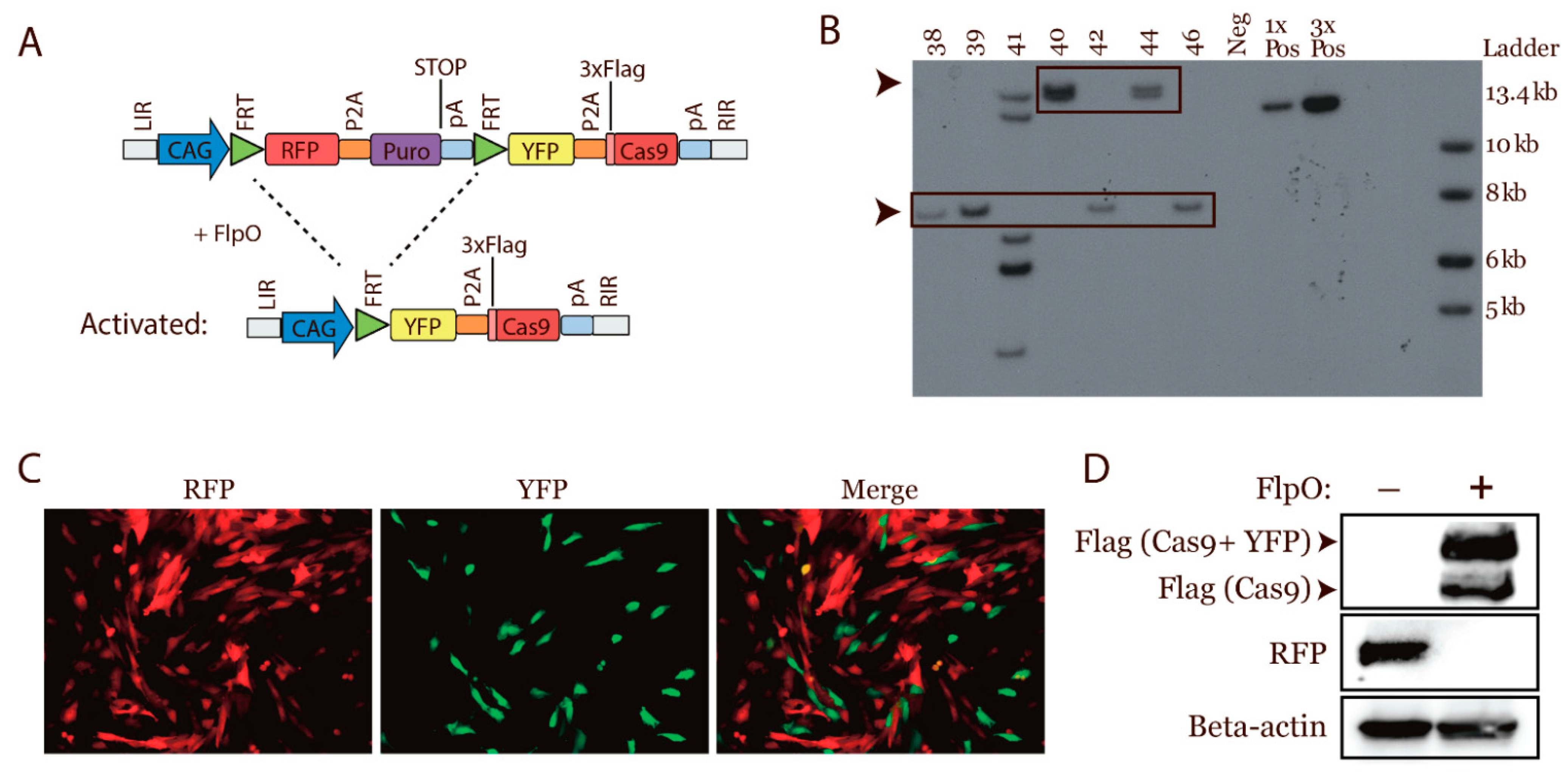
3.1. Design of a Conditional Cas9 Expression Construct
3.2. Generation of a Transgenic Cas9 Minipig by Cloning
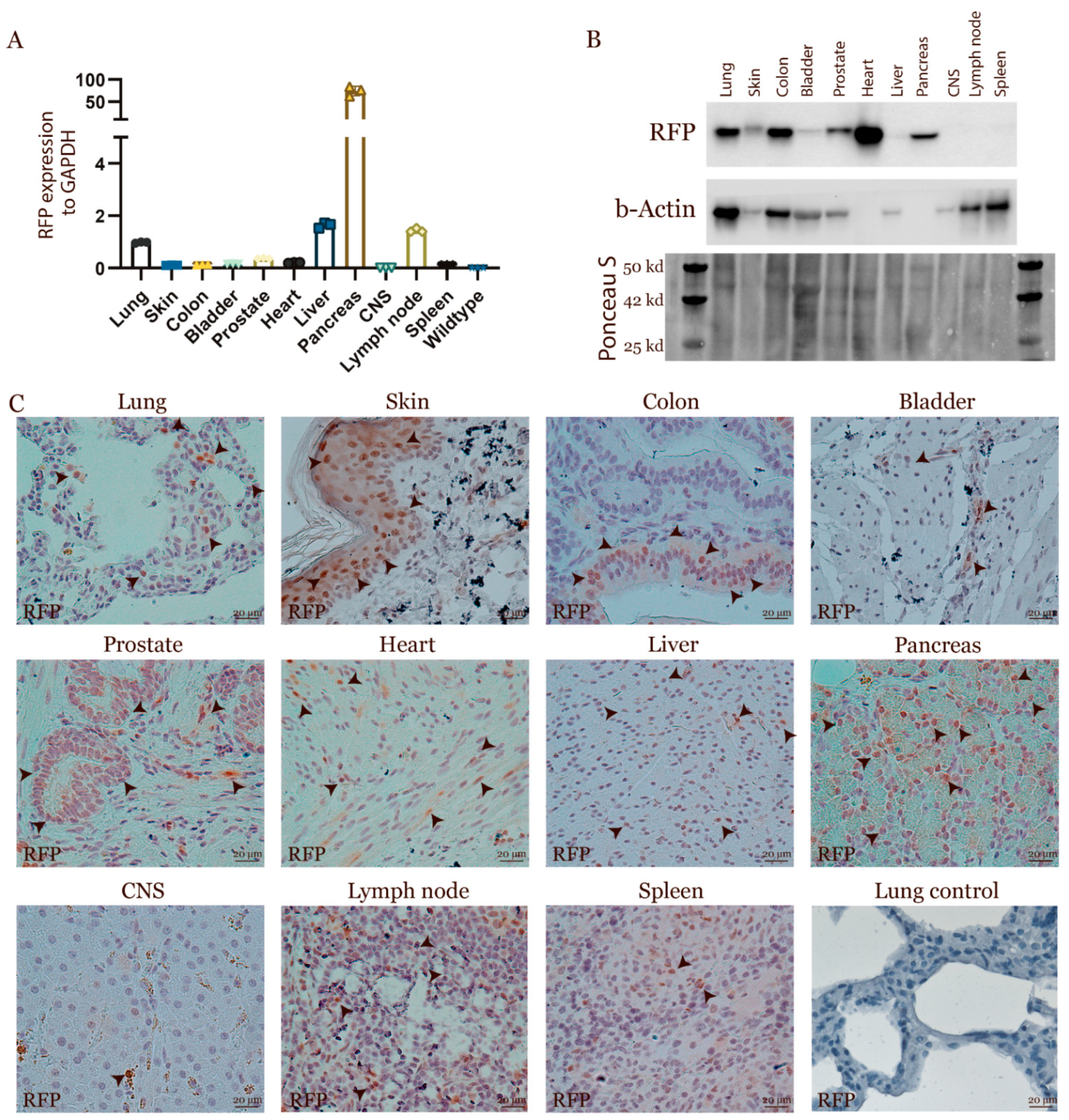
3.3. Characterization of the Transgenic Expression
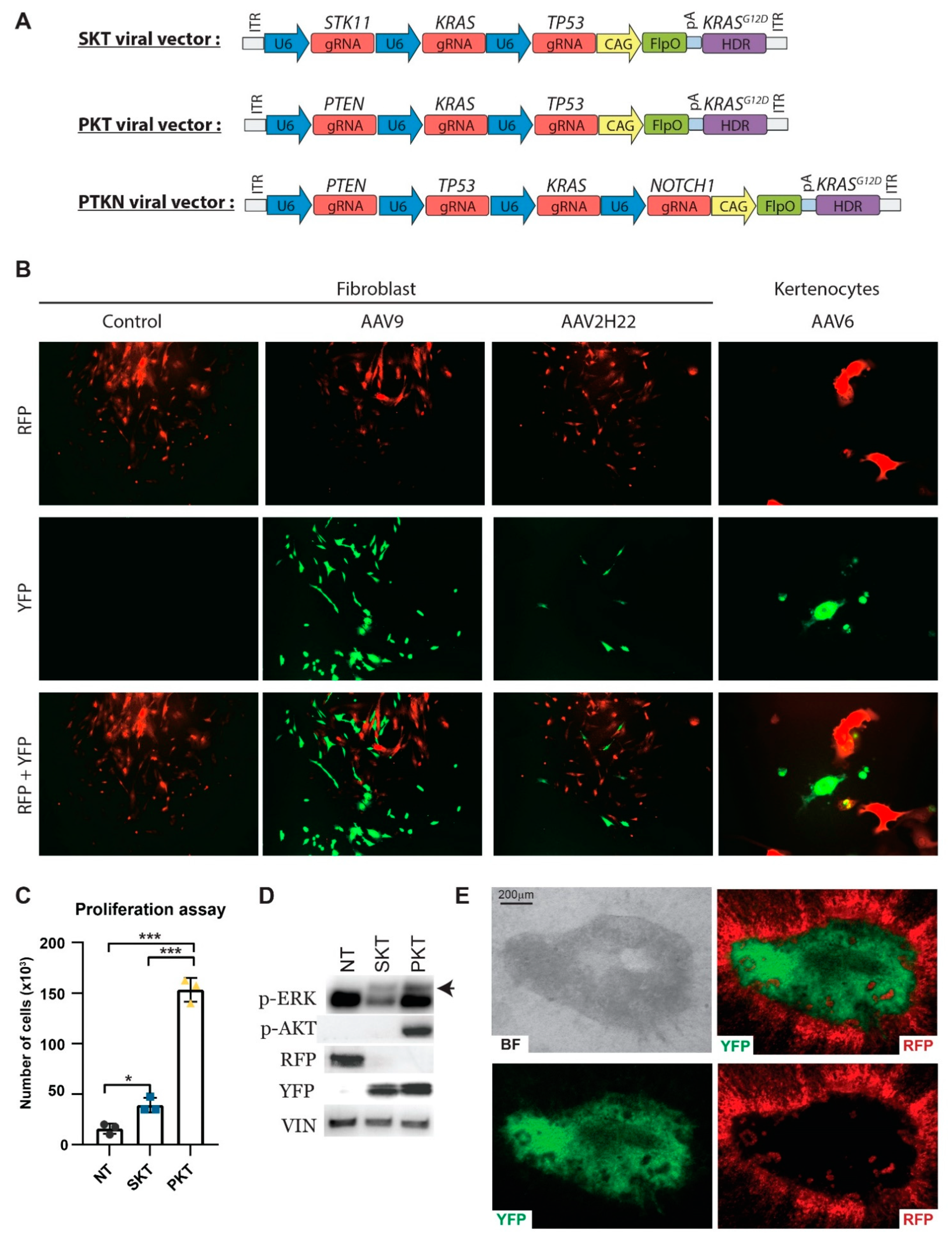
3.4. In Vitro Validation of CRISPR/Cas9-Induced Gene Alterations
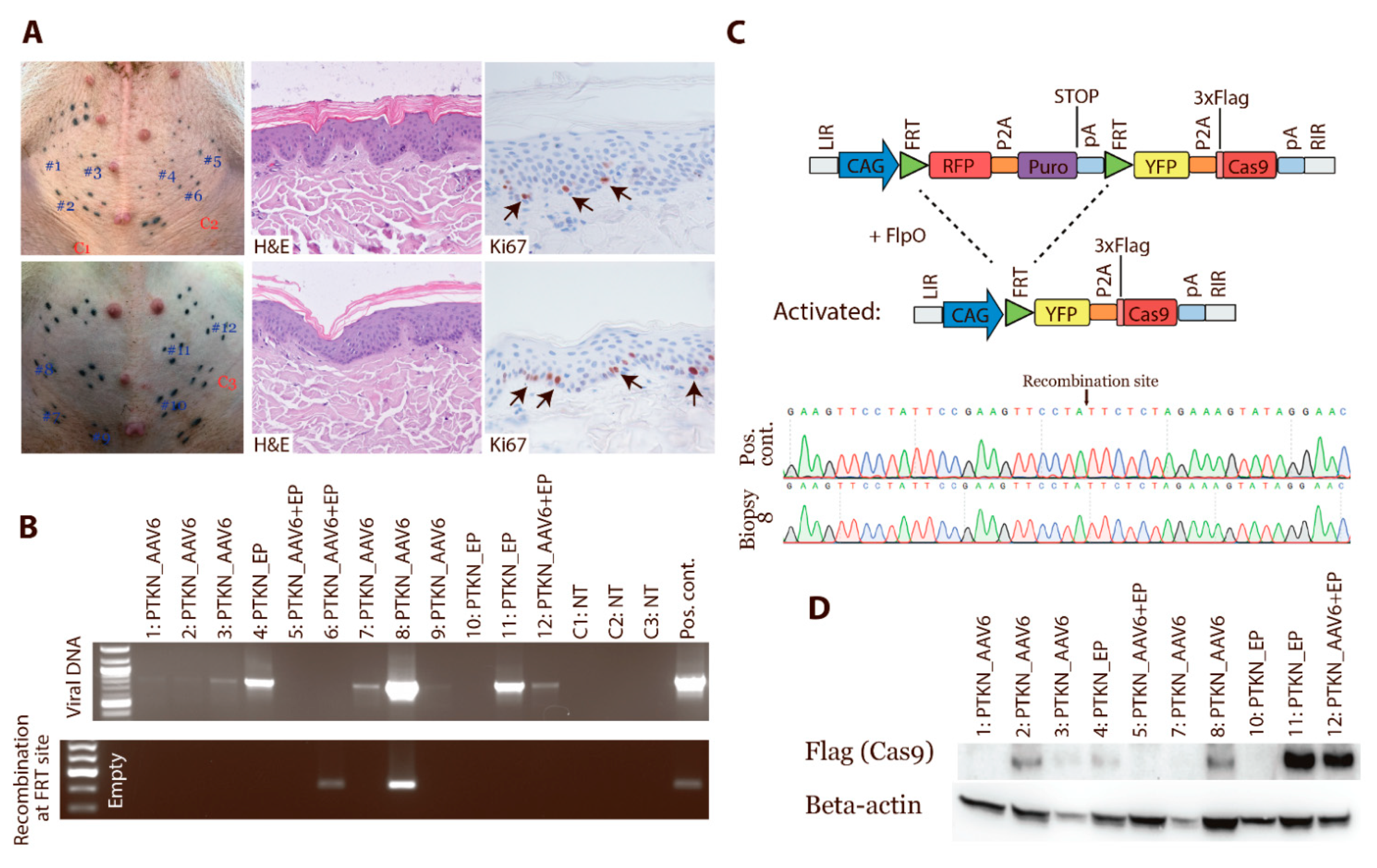
3.5. In Vivo Activation of Cas9 Expression by FlpO-Mediated Recombination
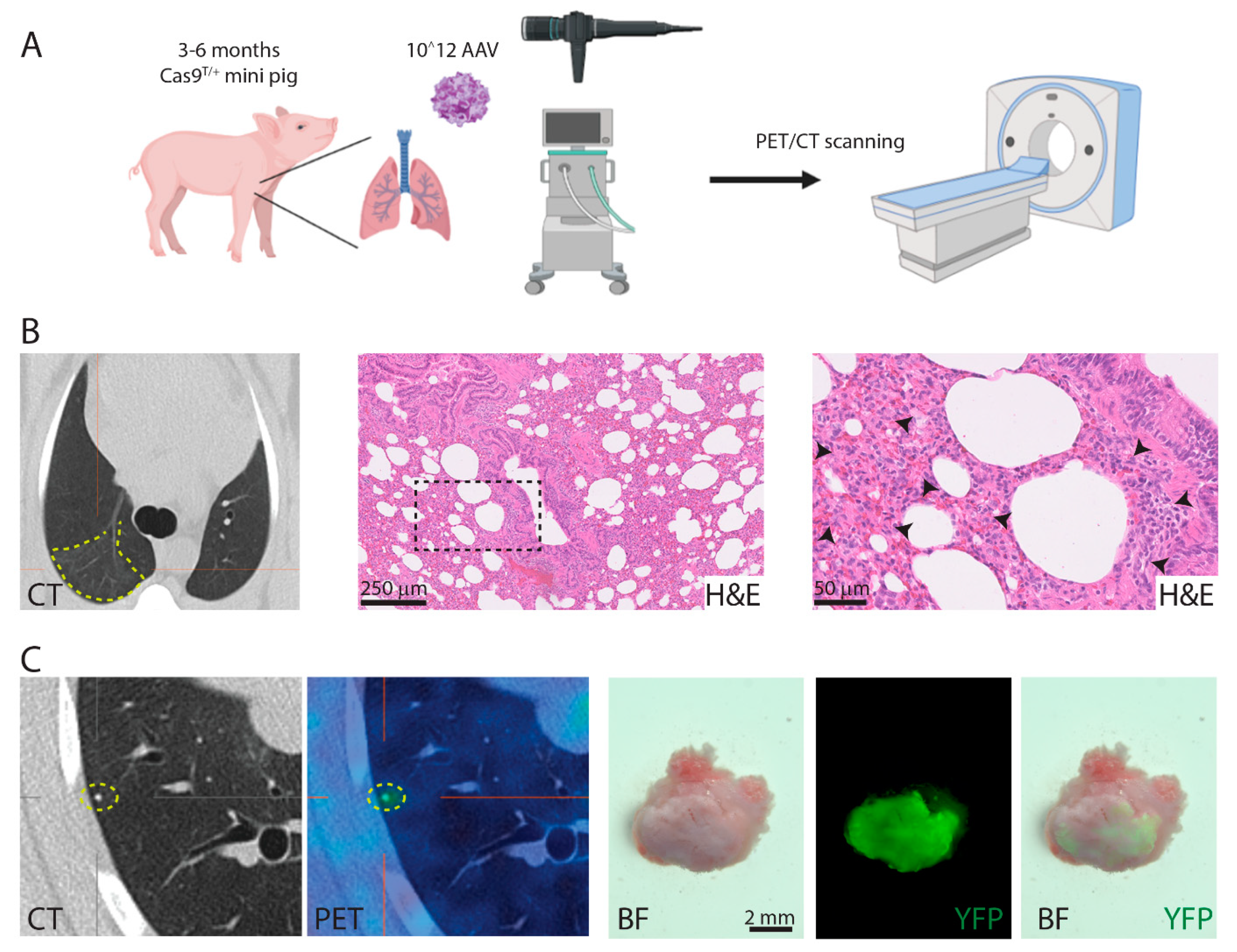
3.6. Induction of Lung Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kellar, A.; Egan, C.; Morris, D. Preclinical Murine Models for Lung Cancer: Clinical Trial Applications. BioMed Res. Int. 2015, 2015, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, I.W.Y.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Res. 2014, 6, 114–118. [Google Scholar] [PubMed]
- Tollis, M.; Boddy, A.M.; Maley, C.C. Peto’s Paradox: How has evolution solved the problem of cancer prevention? BMC Biol. 2017, 15, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, A.L.; Carlson, D.F.; Largaespada, D.A.; Hackett, P.B.; Fahrenkrug, S.C. Engineered Swine Models of Cancer. Front. Genet. 2016, 7, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Wolf, E.; Kind, A.; Aigner, B.; Schnieke, A. Genetically Engineered Large Animals in Biomedicine. In Animal Biotechnology 2; Springer International Publishing: Cham, Switzerland, 2018; pp. 169–214. [Google Scholar]
- Gupta, R.C. Biomarkers in Toxicology, 2nd ed.; Elsevier/Academic Press: Waltham, MA, USA, 2019; p. xxiv. 1222p. [Google Scholar]
- Kalla, D.; Kind, A.; Schnieke, A. Genetically Engineered Pigs to Study Cancer. Int. J. Mol. Sci. 2020, 21, 488. [Google Scholar] [CrossRef]
- Flisikowska, T.; Kind, A.; Schnieke, A. Pigs as models of human cancers. Theriogenology 2016, 86, 433–437. [Google Scholar] [CrossRef]
- Robertson, N.; Schook, L.B.; Schachtschneider, K.M. Porcine cancer models: Potential tools to enhance cancer drug trials. Expert Opin. Drug Discov. 2020, 15, 893–902. [Google Scholar] [CrossRef]
- Schachtschneider, K.M.; Liu, Y.; Mäkeläinen, S.; Madsen, O.; Rund, L.A.; Groenen, M.A.M.; Schook, L.B. Oncopig soft-tissue sarcomas recapitulate key transcriptional features of human sarcomas. Sci. Rep. 2017, 7, 2624. [Google Scholar] [CrossRef] [Green Version]
- Leuchs, S.; Saalfrank, A.; Merkl, C.; Flisikowska, T.; Edlinger, M.; Durkovic, M.; Rezaei, N.; Kurome, M.; Zakhartchenko, V.; Kessler, B.; et al. Inactivation and Inducible Oncogenic Mutation of p53 in Gene Targeted Pigs. PLoS ONE 2012, 7, e43323. [Google Scholar] [CrossRef] [Green Version]
- Sieren, J.C.; Meyerholz, D.K.; Wang, X.J.; Davis, B.T.; Newell, J.D.; Hammond, E.; Rohret, J.A.; Rohret, F.A.; Struzynski, J.T.; Goeken, J.A.; et al. Development and translational imaging of a TP53 porcine tumorigenesis model. J. Clin. Investig. 2014, 124, 4052–4066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callesen, M.M.; Árnadóttir, S.S.; Lyskjær, I.; Ørntoft, M.B.W.; Høyer, S.; Dagnæs-Hansen, F.; Liu, Y.; Li, R.; Callesen, H.; Rasmussen, M.H.; et al. A genetically inducible porcine model of intestinal cancer. Mol. Oncol. 2017, 11, 1616–1629. [Google Scholar] [CrossRef] [Green Version]
- Berthelsen, M.F.; Callesen, M.M.; Østergaard, T.S.; Liu, Y.; Li, R.; Callesen, H.; Dagnæs-Hansen, F.; Hamilton-Dutoit, S.; Jakobsen, J.E.; Thomsen, M.K. Pancreas specific expression of oncogenes in a porcine model. Transgenic Res. 2017, 26, 603–612. [Google Scholar] [CrossRef]
- Niu, G.; Hellmuth, I.; Flisikowska, T.; Pausch, H.; Rieblinger, B.; Carrapeiro, A.; Schade, B.; Bohm, B.; Kappe, E.; Fischer, K.; et al. Porcine model elucidates function of p53 isoform in carcinogenesis and reveals novel circTP53 RNA. Oncogene 2021, 40, 1896–1908. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Riedel, M.; Berthelsen, M.F.; Cai, H.; Haldrup, J.; Borre, M.; Paludan, S.R.; Hager, H.; Vendelbo, M.H.; Wagner, E.F.; Bakiri, L.; et al. In vivo CRISPR inactivation of Fos promotes prostate cancer progression by altering the associated AP-1 subunit Jun. Oncogene 2021. [Google Scholar] [CrossRef]
- Riedel, M.; Berthelsen, M.F.; Bakiri, L.; Wagner, E.F.; Thomsen, M.K. Virus Delivery of CRISPR Guides to the Murine Prostate for Gene Alteration. J. Vis. Exp. 2018, 2018, e57525. [Google Scholar] [CrossRef]
- Berthelsen, M.F.; Leknes, S.L.; Riedel, M.; Pedersen, M.A.; Joseph, J.V.; Hager, H.; Vendelbo, M.H.; Thomsen, M.K. Comparative Analysis of Stk11/Lkb1 versus Pten Deficiency in Lung Adenocarcinoma Induced by CRISPR/Cas9. Cancers 2021, 13, 974. [Google Scholar] [CrossRef]
- Platt, R.J.; Chen, S.; Zhou, Y.; Yim, M.J.; Swiech, L.; Kempton, H.R.; Dahlman, J.E.; Parnas, O.; Eisenhaure, T.M.; Jovanovic, M.; et al. CRISPR-Cas9 Knockin Mice for Genome Editing and Cancer Modeling. Cell 2014, 159, 440–455. [Google Scholar] [CrossRef] [Green Version]
- Hammer, R.E.; Pursel, V.G.; Rexroad, C.E.; Wall, R.J.; Bolt, D.J.; Ebert, K.M.; Palmiter, R.D.; Brinster, R.L. Production of transgenic rabbits, sheep and pigs by microinjection. Nature 1985, 315, 680–683. [Google Scholar] [CrossRef] [Green Version]
- Vajta, G.; Kragh, P.M.; Mtango, N.R.; Callesen, H. Hand-made cloning approach: Potentials and limitations. Reprod. Fertil. Dev. 2005, 17, 97–112. [Google Scholar] [CrossRef]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef]
- Bollen, P.; Ellegaard, L. The Göttingen minipig in pharmacology and toxicology. Pharmacol. Toxicol. 1997, 80 (Suppl. 2), 3–4. [Google Scholar] [CrossRef]
- Kragh, P.M.; Nielsen, A.L.; Li, J.; Du, Y.; Lin, L.; Schmidt, M.; Bøgh, I.B.; Holm, I.E.; Jakobsen, J.E.; Johansen, M.G.; et al. Hemizygous minipigs produced by random gene insertion and handmade cloning express the Alzheimer’s disease-causing dominant mutation APPsw. Transgenic Res. 2009, 18, 545–558. [Google Scholar] [CrossRef]
- Schmidt, M.; Kragh, P.M.; Li, J.; Du, Y.; Lin, L.; Liu, Y.; Bøgh, I.B.; Winther, K.D.; Vajta, G.; Callesen, H. Pregnancies and piglets from large white sow recipients after two transfer methods of cloned and transgenic embryos of different pig breeds. Theriogenology 2010, 74, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Raymond, C.S.; Soriano, P. High-efficiency FLP and PhiC31 site-specific recombination in mammalian cells. PLoS ONE 2007, 2, e162. [Google Scholar] [CrossRef]
- Jakobsen, J.E.; Li, J.; Kragh, P.M.; Moldt, B.; Lin, L.; Liu, Y.; Schmidt, M.; Winther, K.D.; Schyth, B.D.; Holm, I.E.; et al. Pig transgenesis by Sleeping Beauty DNA transposition. Transgenic Res. 2011, 20, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Shcherbo, D.; Murphy, C.S.; Ermakova, G.V.; Solovieva, E.A.; Chepurnykh, T.V.; Shcheglov, A.S.; Verkhusha, V.V.; Pletnev, V.Z.; Hazelwood, K.L.; Roche, P.M.; et al. Far-red fluorescent tags for protein imaging in living tissues. Biochem. J. 2009, 418, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Ibata, K.; Park, E.S.; Kubota, M.; Mikoshiba, K.; Miyawaki, A. A variant of yellow fluorescent protein with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 2002, 20, 87–90. [Google Scholar] [CrossRef]
- Szymczak-Workman, A.L.; Vignali, K.M.; Vignali, D.A.A. Design and Construction of 2A Peptide-Linked Multicistronic Vectors. Cold Spring Harb. Protoc. 2012, 2012, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Alexopoulou, A.N.; Couchman, J.R.; Whiteford, J.R. The CMV early enhancer/chicken beta actin (CAG) promoter can be used to drive transgene expression during the differentiation of murine embryonic stem cells into vascular progenitors. BMC Cell Biol. 2008, 9, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steines, B.; Dickey, D.D.; Bergen, J.; Excoffon, K.J.; Weinstein, J.R.; Li, X.; Yan, Z.; Abou Alaiwa, M.H.; Shah, V.S.; Bouzek, D.C.; et al. CFTR gene transfer with AAV improves early cystic fibrosis pig phenotypes. JCI Insight 2016, 1, e88728. [Google Scholar] [CrossRef]
- Koerber, J.T.; Klimczak, R.; Jang, J.H.; Dalkara, D.; Flannery, J.G.; Schaffer, D.V. Molecular evolution of adeno-associated virus for enhanced glial gene delivery. Mol. Ther. 2009, 17, 2088–2095. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K. Advances in swine biomedical model genomics. Int. J. Biol. Sci. 2007, 3, 179–184. [Google Scholar] [CrossRef]
- Wang, K.; Jin, Q.; Ruan, D.; Yang, Y.; Liu, Q.; Wu, H.; Zhou, Z.; Ouyang, Z.; Liu, Z.; Zhao, Y.; et al. Cre-dependent Cas9-expressing pigs enable efficient in vivo genome editing. Genome Res. 2017, 27, 2061–2071. [Google Scholar] [CrossRef]
- Silver, D.P.; Livingston, D.M. Self-excising retroviral vectors encoding the cre recombinase overcome Cre-mediated cellular toxicity. Mol. Cell 2001, 8, 233–243. [Google Scholar] [CrossRef]
- Takata, Y.; Kondo, S.; Goda, N.; Kanegae, Y.; Saito, I. Comparison of efficiency between FLPe and Cre for recombinase-mediated cassette exchange in vitro and in adenovirus vector production. Genes Cells 2011, 16, 765–777. [Google Scholar] [CrossRef]
- Khan, A.N.; Al-Jahdali, H.H.; Allen, C.M.; Irion, K.L.; Al Ghanem, S.; Koteyar, S.S. The calcified lung nodule: What does it mean? Ann. Thorac. Med. 2010, 5, 67–79. [Google Scholar] [CrossRef]
- McGavin, M.D.; Carlton, W.; Zachary, J.F.; Thomson, R.G. Thomson’s Special Veterinary Pathology, 3rd ed.; Mosby: St. Louis, MO, USA, 2001; p. xi. 755p. [Google Scholar]
- Overgaard, N.H.; Principe, D.R.; Schachtschneider, K.M.; Jakobsen, J.T.; Rund, L.A.; Grippo, P.J.; Schook, L.B.; Jungersen, G. Genetically induced tumors in the oncopig model invoke an antitumor immune response dominated by cytotoxic CD8β+ T cells and differentiated γδ T cells alongside a regulatory response mediated by foxp3+ t cells and immunoregulatory molecules. Front. Immunol. 2018, 9, 1301. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berthelsen, M.F.; Riedel, M.; Cai, H.; Skaarup, S.H.; Alstrup, A.K.O.; Dagnæs-Hansen, F.; Luo, Y.; Jensen, U.B.; Hager, H.; Liu, Y.; et al. The CRISPR/Cas9 Minipig—A Transgenic Minipig to Produce Specific Mutations in Designated Tissues. Cancers 2021, 13, 3024. https://doi.org/10.3390/cancers13123024
Berthelsen MF, Riedel M, Cai H, Skaarup SH, Alstrup AKO, Dagnæs-Hansen F, Luo Y, Jensen UB, Hager H, Liu Y, et al. The CRISPR/Cas9 Minipig—A Transgenic Minipig to Produce Specific Mutations in Designated Tissues. Cancers. 2021; 13(12):3024. https://doi.org/10.3390/cancers13123024
Chicago/Turabian StyleBerthelsen, Martin Fogtmann, Maria Riedel, Huiqiang Cai, Søren H. Skaarup, Aage K. O. Alstrup, Frederik Dagnæs-Hansen, Yonglun Luo, Uffe B. Jensen, Henrik Hager, Ying Liu, and et al. 2021. "The CRISPR/Cas9 Minipig—A Transgenic Minipig to Produce Specific Mutations in Designated Tissues" Cancers 13, no. 12: 3024. https://doi.org/10.3390/cancers13123024