MTA2 as a Potential Biomarker and Its Involvement in Metastatic Progression of Human Renal Cancer by miR-133b Targeting MMP-9


 , and
, and 
Abstract
:1. Introduction
2. Results
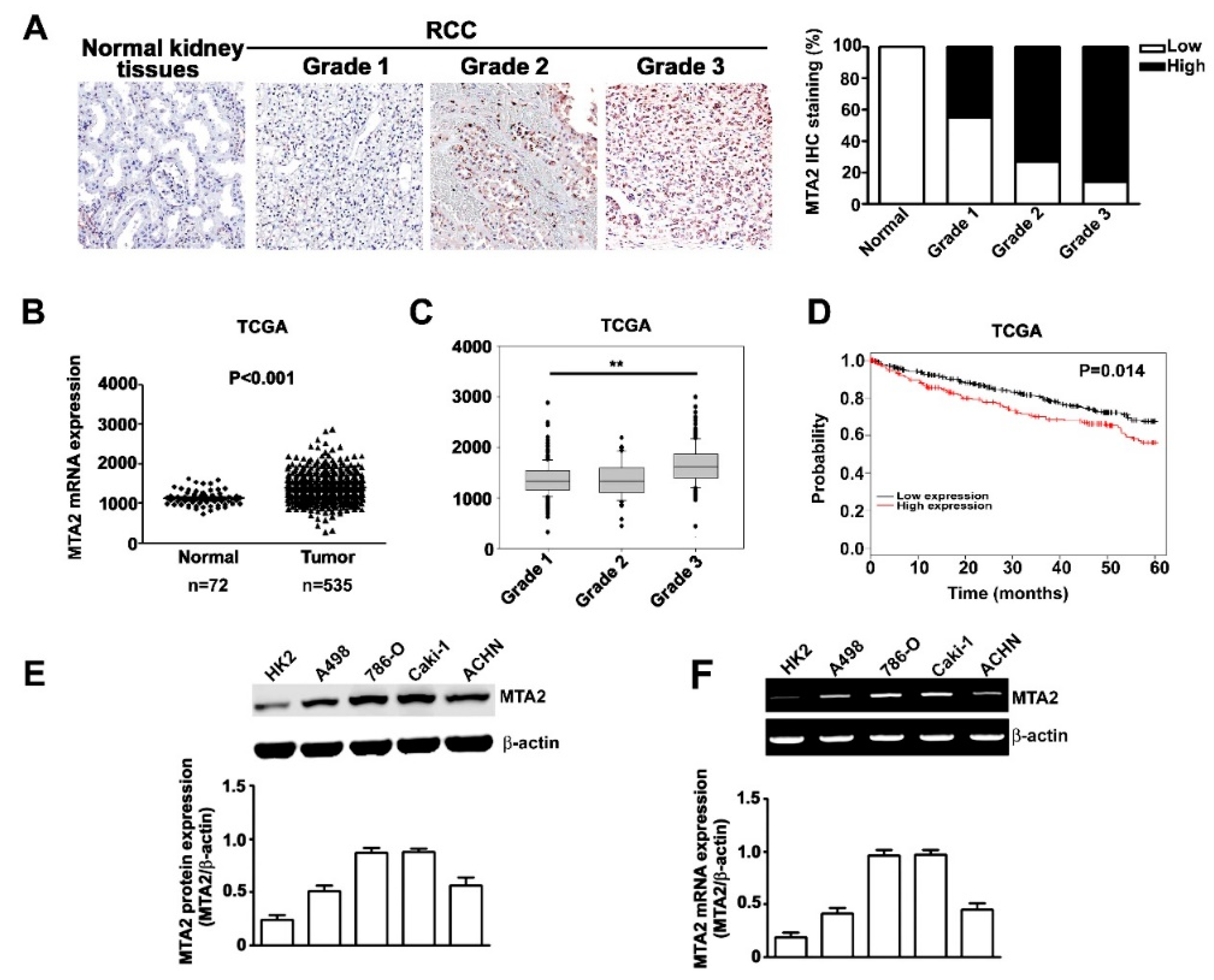
2.1. Expression and Effects of MTA2 in Human RCC and RCC Cells
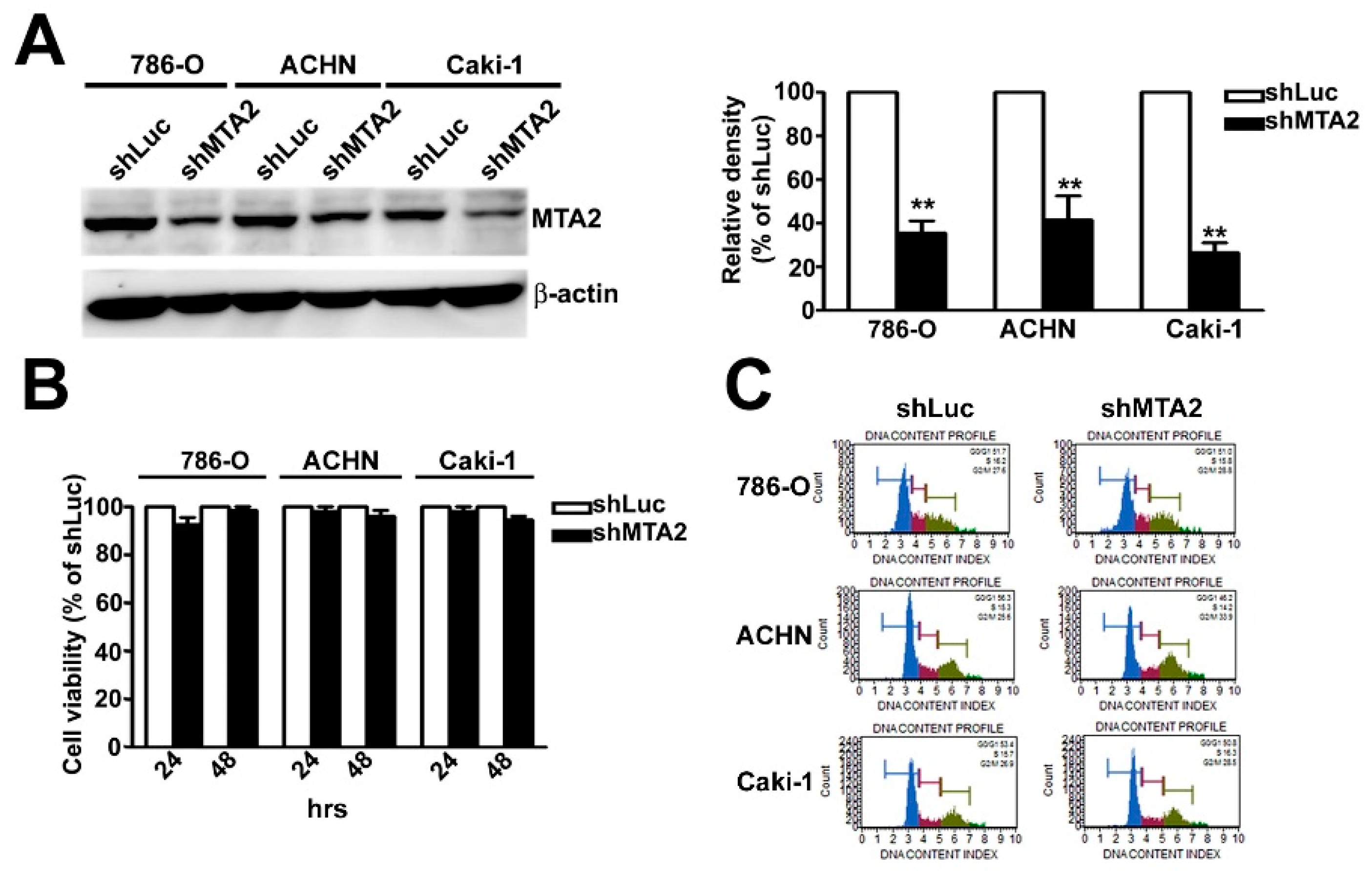
2.2. Effect of MTA2 Knockdown on RCC Cell Proliferation
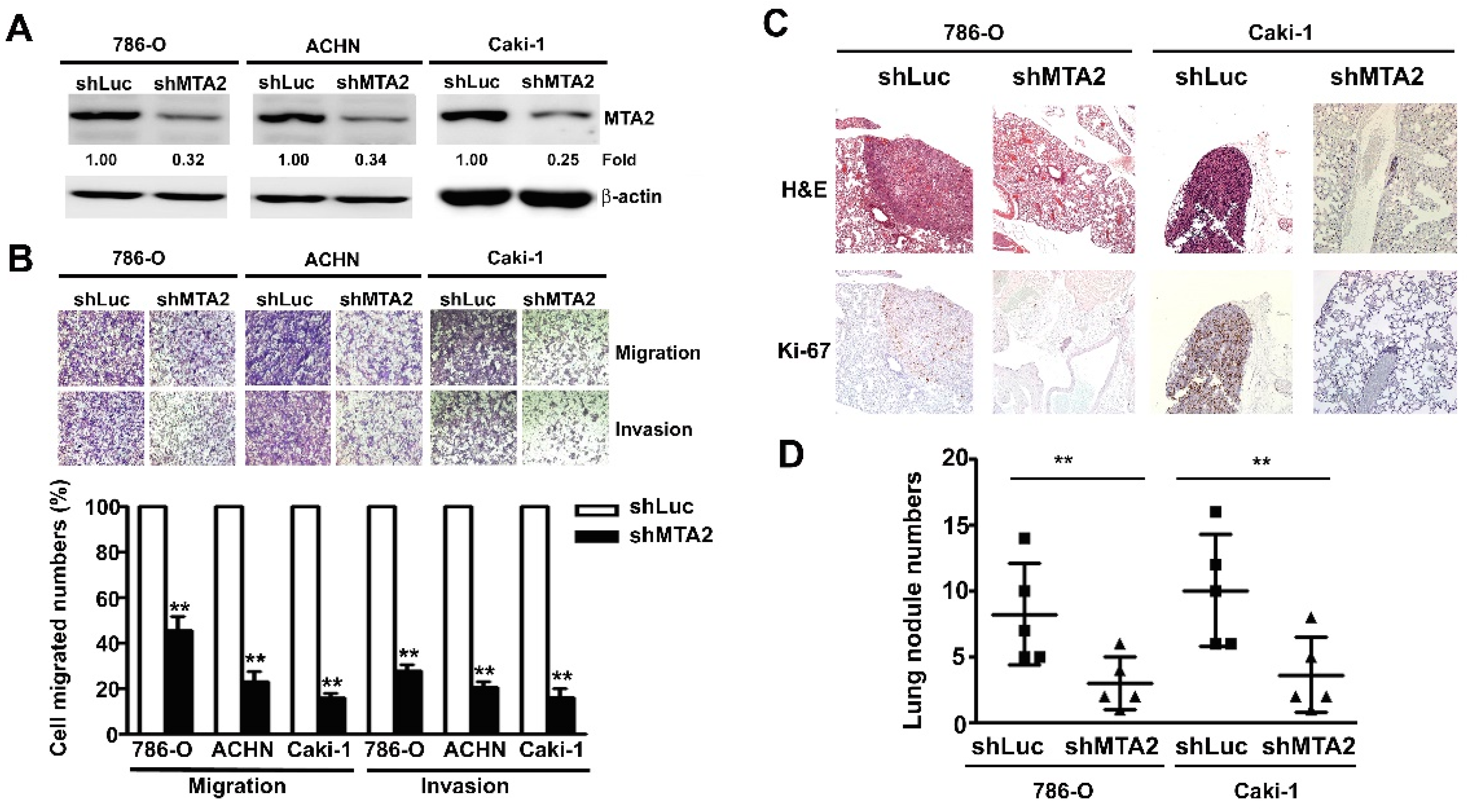
2.3. Effect of MTA2 Knockdown on RCC Cell Metastasis in Vitro and in Vivo
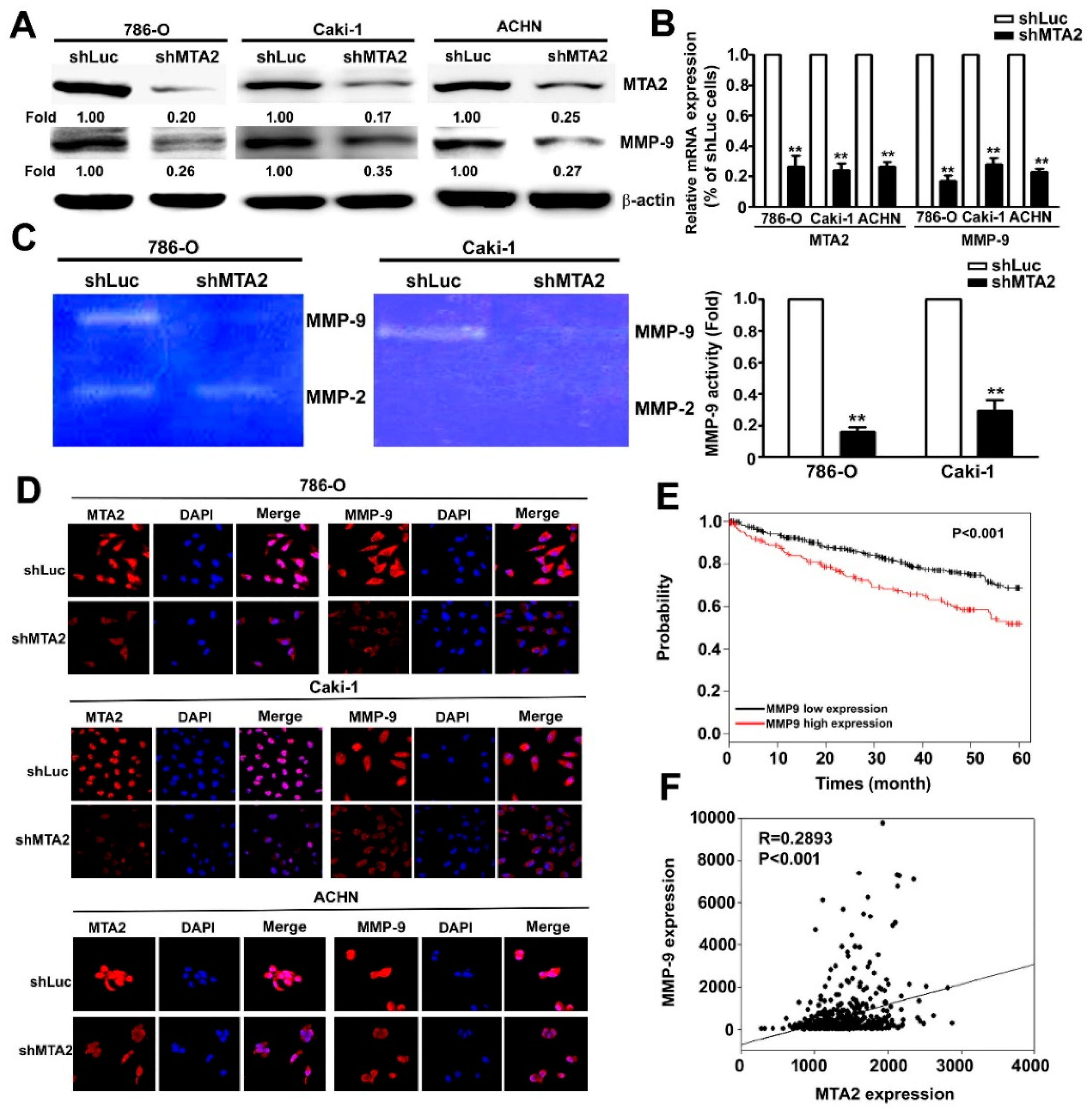
2.4. Effect of MTA2 Knockdown on MMP-9 Expression in RCC Cells
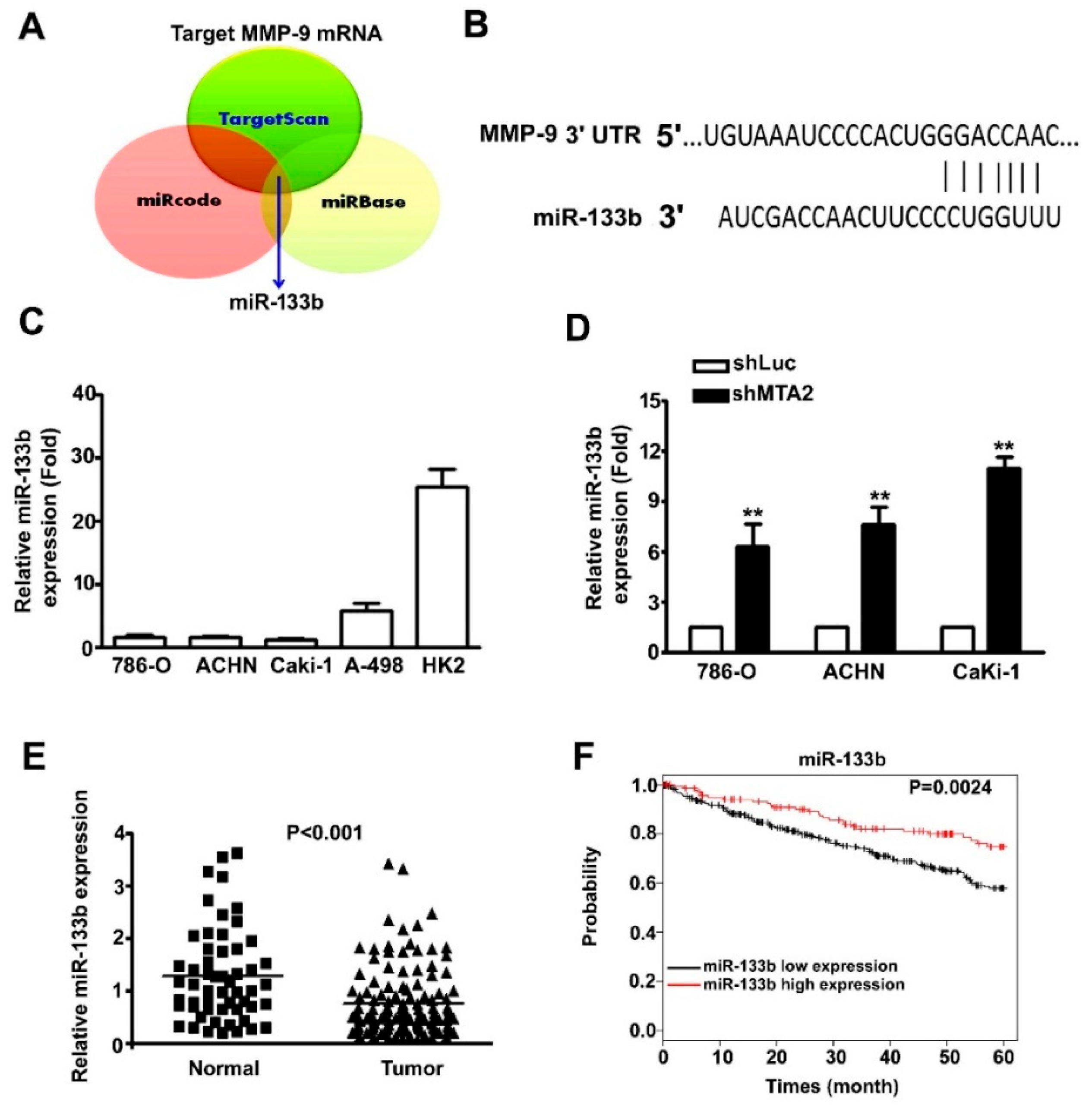
2.5. MMP-9 as the Target Gene of miR-133b and Association with Poor RCC Prognosis.
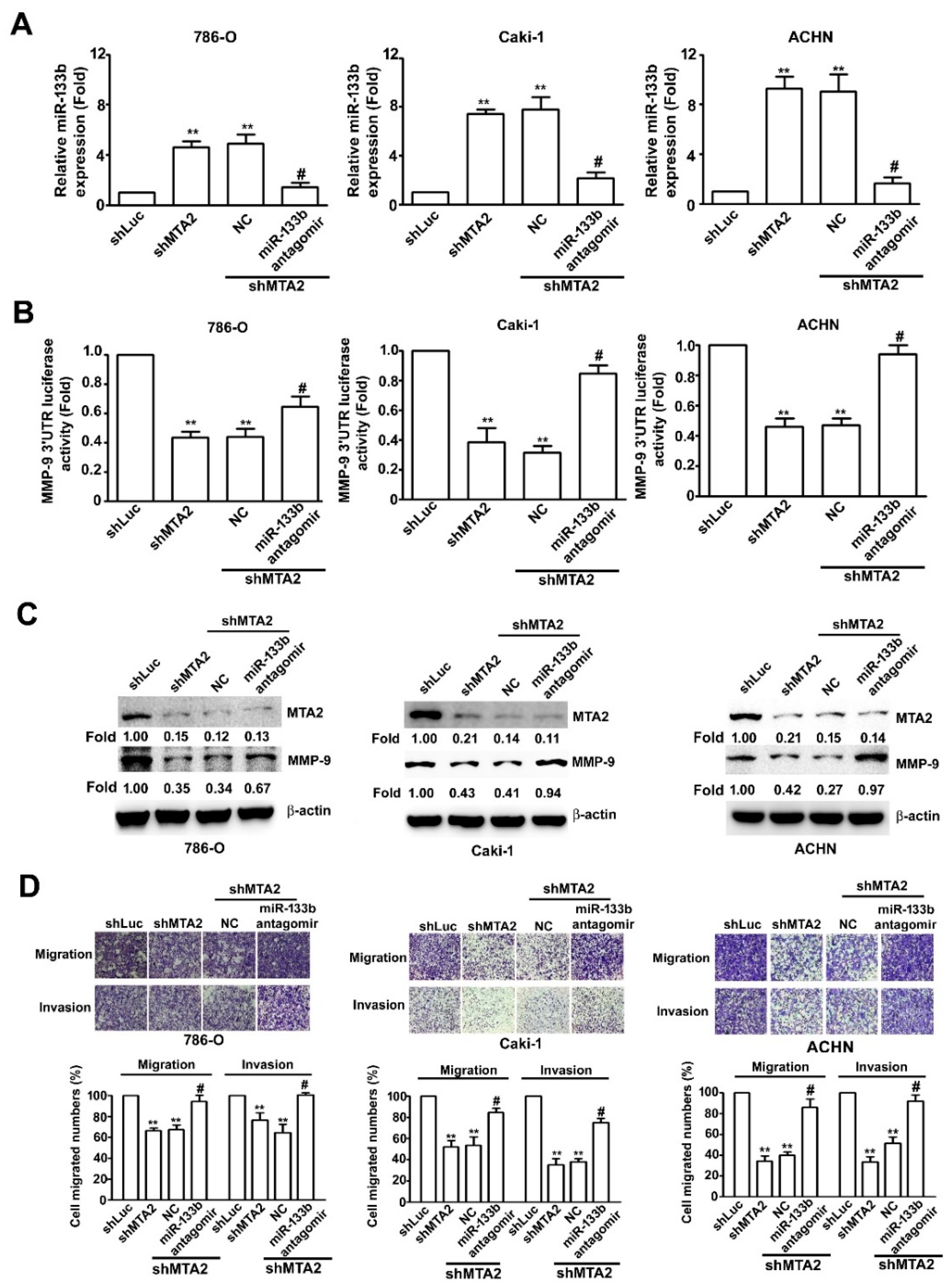
2.6. Effect of miR-133b on MMP-9 Expression Involved in Knockdown MTA2-Inhibiting RCC Cell Metastasis
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Human Kidney Clear Cell Carcinoma Tissue Array
4.3. Cell Culture and shRNA Assay
4.4. RNA Isolation, RT-PCR, and QRT-PCR
4.5. Cell Viability Assay
4.6. Cell Cycle Analysis
4.7. In Vitro Cell Migration and Invasion Assays
4.8. MiR-133b Antagomir and MTA2 Plasmid Transfection
4.9. Western Blot Analysis
4.10. Immunofluorescence Assay
4.11. Gelatine Zymography
4.12. TCGA Database and miRNA Prediction of Bioinformatic Analysis
4.13. Luciferase Reporter Assay
4.14. In Vivo Animal Model and Immunohistochemistry Analysis
4.15. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Znaor, A.; Lortet-Tieulent, J.; Laversanne, M.; Jemal, A.; Bray, F. International variations and trends in renal cell carcinoma incidence and mortality. Eur. Urol. 2015, 67, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.J.; Hernandez-Meza, G.; Agrawal, P.; Zhang, C.A.; Xie, L.; Gong, C.L.; Hoerner, C.R.; Srinivas, S.; Oermann, E.K.; Fan, A.C. Time on Therapy for at Least Three Months Correlates with Overall Survival in Metastatic Renal Cell Carcinoma. Cancers (Basel) 2019, 11, 1000. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, C.J.; De Cubas, A.A.; Fan, H.; Smith, C.C.; Lang, M.; Reznik, E.; Bowlby, R.; Gibb, E.A.; Akbani, R.; Beroukhim, R.; et al. The Cancer Genome Atlas Comprehensive Molecular Characterization of Renal Cell Carcinoma. Cell. Rep. 2018, 23, 313–326.e5. [Google Scholar] [CrossRef] [PubMed]
- Huilgol, D.; Venkataramani, P.; Nandi, S.; Bhattacharjee, S. Transcription Factors That Govern Development and Disease: An Achilles Heel in Cancer. Genes (Basel) 2019, 10, 794. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.; Larkin, J.; Ficarra, V. Renal cell carcinoma. Nat. Rev. Dis. Primers 2017, 3, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Hudes, G.; Carducci, M.; Tomczak, P.; Dutcher, J.; Figlin, R.; Kapoor, A.; Staroslawska, E.; Sosman, J.; McDermott, D.; Bodrogi, I.; et al. Temsirolimus, interferon alfa, or both for advanced renal-cell carcinoma. N. Engl. J. Med. 2007, 356, 2271–2281. [Google Scholar] [CrossRef]
- Motzer, R.J.; Escudier, B.; McDermott, D.F.; George, S.; Hammers, H.J.; Srinivas, S.; Tykodi, S.S.; Sosman, J.A.; Procopio, G.; Plimack, E.R.; et al. Nivolumab versus Everolimus in Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2015, 373, 1803–1813. [Google Scholar] [CrossRef]
- Ghosh, D.; Venkataramani, P.; Nandi, S.; Bhattacharjee, S. CRISPR-Cas9 a boon or bane: The bumpy road ahead to cancer therapeutics. Cancer Cell. Int. 2019, 19, 12. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Nandi, S. Rare Genetic Diseases with Defects in DNA Repair: Opportunities and Challenges in Orphan Drug Development for Targeted Cancer Therapy. Cancers (Basel) 2018, 10, 298. [Google Scholar] [CrossRef]
- Ravaud, A.; Motzer, R.J.; Pandha, H.S.; George, D.J.; Pantuck, A.J.; Patel, A.; Chang, Y.H.; Escudier, B.; Donskov, F.; Magheli, A.; et al. Adjuvant Sunitinib in High-Risk Renal-Cell Carcinoma after Nephrectomy. N. Engl. J. Med. 2016, 375, 2246–2254. [Google Scholar] [CrossRef]
- Zhou, J.; Zhan, S.; Tan, W.; Cheng, R.; Gong, H.; Zhu, Q. P300 binds to and acetylates MTA2 to promote colorectal cancer cells growth. Biochem. Biophys. Res. Commun. 2014, 444, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Covington, K.R.; Fuqua, S.A. Role of MTA2 in human cancer. Cancer Metastasis Rev. 2014, 33, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Ji, J.; Cai, Q.; Shi, M.; Chen, X.; Yu, Y.; Liu, B.; Zhu, Z.; Zhang, J. MTA2 promotes gastric cancer cells invasion and is transcriptionally regulated by Sp1. Mol. Cancer 2013, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, H.; Shen, G. Metastasis-associated protein 2 (MTA2) promotes the metastasis of non-small-cell lung cancer through the inhibition of the cell adhesion molecule Ep-CAM and E-cadherin. Jpn. J. Clin. Oncol. 2015, 45, 755–766. [Google Scholar] [CrossRef]
- Covington, K.R.; Brusco, L.; Barone, I.; Tsimelzon, A.; Selever, J.; Corona-Rodriguez, A.; Brown, P.; Kumar, R.; Hilsenbeck, S.G.; Fuqua, S.A. Metastasis tumor-associated protein 2 enhances metastatic behavior and is associated with poor outcomes in estrogen receptor-negative breast cancer. Breast Cancer Res. Treat. 2013, 141, 375–384. [Google Scholar] [CrossRef]
- Liotta, L.A.; Stetler-Stevenson, W.G. Tumor invasion and metastasis: An imbalance of positive and negative regulation. Cancer Res. 1991, 51, 5054s–5059s. [Google Scholar]
- Gaffney, J.; Solomonov, I.; Zehorai, E.; Sagi, I. Multilevel regulation of matrix metalloproteinases in tissue homeostasis indicates their molecular specificity in vivo. Matrix Biol. 2015, 44–46, 191–199. [Google Scholar] [CrossRef]
- Sato, A.; Nagase, H.; Obinata, D.; Fujiwara, K.; Fukuda, N.; Soma, M.; Yamaguchi, K.; Kawata, N.; Takahashi, S. Inhibition of MMP-9 using a pyrrole-imidazole polyamide reduces cell invasion in renal cell carcinoma. Int. J. Oncol. 2013, 43, 1441–1446. [Google Scholar] [CrossRef]
- Cho, N.H.; Shim, H.S.; Rha, S.Y.; Kang, S.H.; Hong, S.H.; Choi, Y.D.; Hong, S.J.; Cho, S.H. Increased expression of matrix metalloproteinase 9 correlates with poor prognostic variables in renal cell carcinoma. Eur. Urol. 2003, 44, 560–566. [Google Scholar] [CrossRef]
- Ghosh, D.; Nandi, S.; Bhattacharjee, S. Combination therapy to checkmate Glioblastoma: Clinical challenges and advances. Clin. Transl. Med. 2018, 7, 33. [Google Scholar] [CrossRef]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Tabouret, E.; Boudouresque, F.; Farina, P.; Barrie, M.; Bequet, C.; Sanson, M.; Chinot, O. MMP2 and MMP9 as candidate biomarkers to monitor bevacizumab therapy in high-grade glioma. Neuro. Oncol. 2015, 17, 1174–1176. [Google Scholar] [CrossRef] [PubMed]
- Schickel, R.; Boyerinas, B.; Park, S.M.; Peter, M.E. MicroRNAs: Key players in the immune system, differentiation, tumorigenesis and cell death. Oncogene 2008, 27, 5959–5974. [Google Scholar] [CrossRef] [PubMed]
- Kano, M.; Seki, N.; Kikkawa, N.; Fujimura, L.; Hoshino, I.; Akutsu, Y.; Chiyomaru, T.; Enokida, H.; Nakagawa, M.; Matsubara, H. miR-145, miR-133a and miR-133b: Tumor-suppressive miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int. J. Cancer 2010, 127, 2804–2814. [Google Scholar] [CrossRef]
- Qiu, T.; Zhou, X.; Wang, J.; Du, Y.; Xu, J.; Huang, Z.; Zhu, W.; Shu, Y.; Liu, P. MiR-145, miR-133a and miR-133b inhibit proliferation, migration, invasion and cell cycle progression via targeting transcription factor Sp1 in gastric cancer. FEBS Lett. 2014, 588, 1168–1177. [Google Scholar] [CrossRef]
- Zeng, W.; Zhu, J.F.; Liu, J.Y.; Li, Y.L.; Dong, X.; Huang, H.; Shan, L. miR-133b inhibits cell proliferation, migration and invasion of esophageal squamous cell carcinoma by targeting EGFR. Biomed. Pharmacother. 2019, 111, 476–484. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Zhou, C.X.; Zhan, M.N.; Tang, J.; Wang, C.L.; Ma, C.N.; He, M.; Chen, G.Q.; He, J.R.; Zhao, Q. MiR-133b targets Sox9 to control pathogenesis and metastasis of breast cancer. Cell Death Dis. 2018, 9, 752. [Google Scholar] [CrossRef]
- Wu, D.; Pan, H.; Zhou, Y.; Zhou, J.; Fan, Y.; Qu, P. microRNA-133b downregulation and inhibition of cell proliferation, migration and invasion by targeting matrix metallopeptidase-9 in renal cell carcinoma. Mol. Med. Rep. 2014, 9, 2491–2498. [Google Scholar] [CrossRef]
- Liu, Y.P.; Shan, B.E.; Wang, X.L.; Ma, L. Correlation between MTA2 overexpression and tumour progression in esophageal squamous cell carcinoma. Exp. Ther. Med. 2012, 3, 745–749. [Google Scholar] [CrossRef]
- Liu, S.L.; Han, Y.; Zhang, Y.; Xie, C.Y.; Wang, E.H.; Miao, Y.; Li, H.Y.; Xu, H.T.; Dai, S.D. Expression of metastasis-associated protein 2 (MTA2) might predict proliferation in non-small cell lung cancer. Target. Oncol. 2012, 7, 135–143. [Google Scholar] [CrossRef]
- Ding, W.; Hu, W.; Yang, H.; Ying, T.; Tian, Y. Prognostic correlation between MTA2 expression level and colorectal cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 7173–7180. [Google Scholar] [PubMed]
- Deryugina, E.I.; Bourdon, M.A.; Reisfeld, R.A.; Strongin, A. Remodeling of collagen matrix by human tumor cells requires activation and cell surface association of matrix metalloproteinase-2. Cancer Res. 1998, 58, 3743–3750. [Google Scholar] [PubMed]
- Narula, S.; Tandon, C.; Tandon, S. Role of Matrix Metalloproteinases in Degenerative Kidney Disorders. Curr. Med. Chem. 2018, 25, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jin, M.L. Short-hairpin RNA-mediated MTA2 silencing inhibits human breast cancer cell line MDA-MB231 proliferation and metastasis. Asian Pac. J. Cancer Prev. 2014, 15, 5577–5582. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Chou, Y.E.; Ko, C.P.; Yang, S.F.; Hsieh, S.C.; Lin, C.L.; Hsieh, Y.H.; Chen, K.C. Metastasis tumor-associated protein-2 knockdown suppresses the proliferation and invasion of human glioma cells in vitro and in vivo. J. Neurooncol. 2014, 120, 273–281. [Google Scholar] [CrossRef]
- Wu, M.; Ye, X.; Deng, X.; Wu, Y.; Li, X.; Zhang, L. Upregulation of metastasis-associated gene 2 promotes cell proliferation and invasion in nasopharyngeal carcinoma. OncoTargets Ther. 2016, 9, 1647–1656. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Fan, X.; Xu, B.; Pang, Q.; Teng, L. miR-133b acts as a tumor suppressor and negatively regulates EMP2 in glioma. Neoplasma 2018, 65, 494–504. [Google Scholar] [CrossRef]
- Huang, S.; Wa, Q.; Pan, J.; Peng, X.; Ren, D.; Li, Q.; Dai, Y.; Yang, Q.; Huang, Y.; Zhang, X.; et al. Transcriptional downregulation of miR-133b by REST promotes prostate cancer metastasis to bone via activating TGF-beta signaling. Cell Death Dis. 2018, 9, 779. [Google Scholar] [CrossRef]
- Zhao, F.; Zhou, L.H.; Ge, Y.Z.; Ping, W.W.; Wu, X.; Xu, Z.L.; Wang, M.; Sha, Z.L.; Jia, R.P. MicroRNA-133b suppresses bladder cancer malignancy by targeting TAGLN2-mediated cell cycle. J. Cell. Physiol. 2019, 234, 4910–4923. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, J.; Zhang, L.; Qu, Y.; Li, J.; Yu, B.; Yan, M.; Yu, Y.; Liu, B.; Zhu, Z. MiR-133b is frequently decreased in gastric cancer and its overexpression reduces the metastatic potential of gastric cancer cells. BMC Cancer 2014, 14, 34. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, G. MicroRNA-133b inhibits proliferation and invasion of ovarian cancer cells through Akt and Erk1/2 inactivation by targeting epidermal growth factor receptor. Int. J. Clin. Exp. Pathol. 2015, 8, 10605–10614. [Google Scholar] [PubMed]
- Zhou, W.; Bi, X.; Gao, G.; Sun, L. miRNA-133b and miRNA-135a induce apoptosis via the JAK2/STAT3 signaling pathway in human renal carcinoma cells. Biomed. Pharmacother. 2016, 84, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.Y.; Chen, C.M.; Hsu, W.H.; Hsieh, Y.H.; Liu, C.J. Overexpression of Endothelial Cell-Specific Molecule 1 Correlates with Gleason Score and Expression of Androgen Receptor in Prostate Carcinoma. Int. J. Med. Sci. 2017, 14, 1263–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, T.Y.; Chiou, H.L.; Lin, C.W.; Chen, Y.S.; Hsu, L.S.; Lee, C.H.; Hsieh, Y.H. Repression of metastasis-associated protein 2 for inhibiting metastasis of human oral cancer cells by promoting the p-cofilin-1/LC3-II expression. J. Oral. Pathol. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chiang, K.C.; Lai, C.Y.; Chiou, H.L.; Lin, C.L.; Chen, Y.S.; Kao, S.H.; Hsieh, Y.H. Timosaponin AIII inhibits metastasis of renal carcinoma cells through suppressing cathepsin C expression by AKT/miR-129-5p axis. J. Cell. Physiol. 2019, 234, 13332–13341. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Number of Patients (%) | p Value | |
|---|---|---|---|
| MTA2 Staining | |||
| Negative | Positive | ||
| Total Number of Patients | 40 (40.4) | 59 (59.5) | − |
| Age (Year) | |||
| <59 | 20 (31.3) | 26 (68.7) | 0.580 |
| ≥59 | 20 (32.1) | 33 (67.9) | |
| Gender | |||
| Male | 13 (43.3) | 17 (56.7) | 0.347 |
| Female | 27 (39.1) | 42 (60.9) | |
| Tumor Grade | |||
| 1 | 29 (54.7) | 24 (45.3) | 0.002 |
| 2 + 3 | 11 (23.9) | 35 (76.1) | |
| Tumor Stage | |||
| Ι | 27 (36.5) | 47 (63.5) | 0.129 |
| II + III | 2 (10) | 18 (90) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-S.; Hung, T.-W.; Su, S.-C.; Lin, C.-L.; Yang, S.-F.; Lee, C.-C.; Yeh, C.-F.; Hsieh, Y.-H.; Tsai, J.-P. MTA2 as a Potential Biomarker and Its Involvement in Metastatic Progression of Human Renal Cancer by miR-133b Targeting MMP-9. Cancers 2019, 11, 1851. https://doi.org/10.3390/cancers11121851
Chen Y-S, Hung T-W, Su S-C, Lin C-L, Yang S-F, Lee C-C, Yeh C-F, Hsieh Y-H, Tsai J-P. MTA2 as a Potential Biomarker and Its Involvement in Metastatic Progression of Human Renal Cancer by miR-133b Targeting MMP-9. Cancers. 2019; 11(12):1851. https://doi.org/10.3390/cancers11121851
Chicago/Turabian StyleChen, Yong-Syuan, Tung-Wei Hung, Shih-Chi Su, Chia-Liang Lin, Shun-Fa Yang, Chu-Che Lee, Chang-Fang Yeh, Yi-Hsien Hsieh, and Jen-Pi Tsai. 2019. "MTA2 as a Potential Biomarker and Its Involvement in Metastatic Progression of Human Renal Cancer by miR-133b Targeting MMP-9" Cancers 11, no. 12: 1851. https://doi.org/10.3390/cancers11121851