TCam-2 Cells Deficient for SOX2 and FOXA2 Are Blocked in Differentiation and Maintain a Seminoma-Like Cell Fate In Vivo

 ,
,
Abstract
:1. Introduction
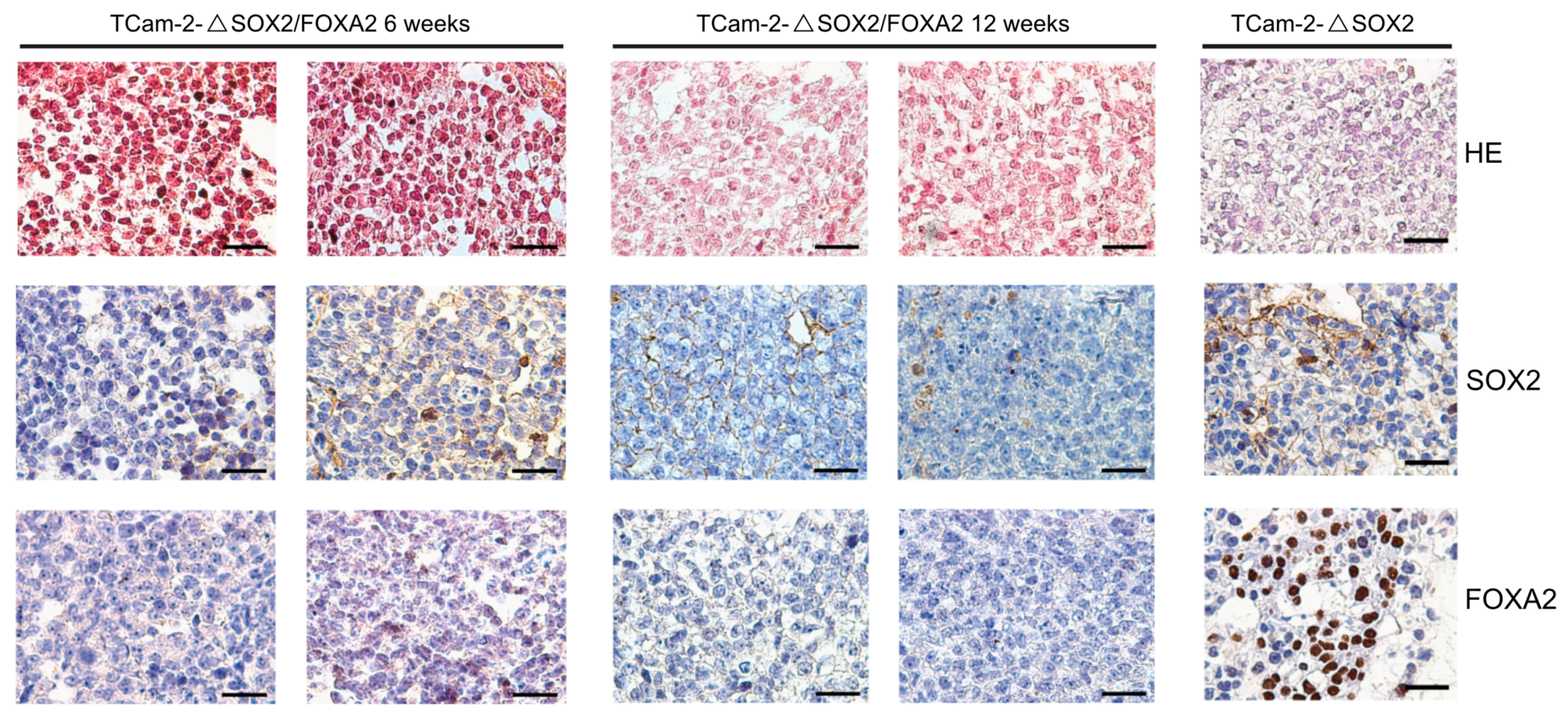
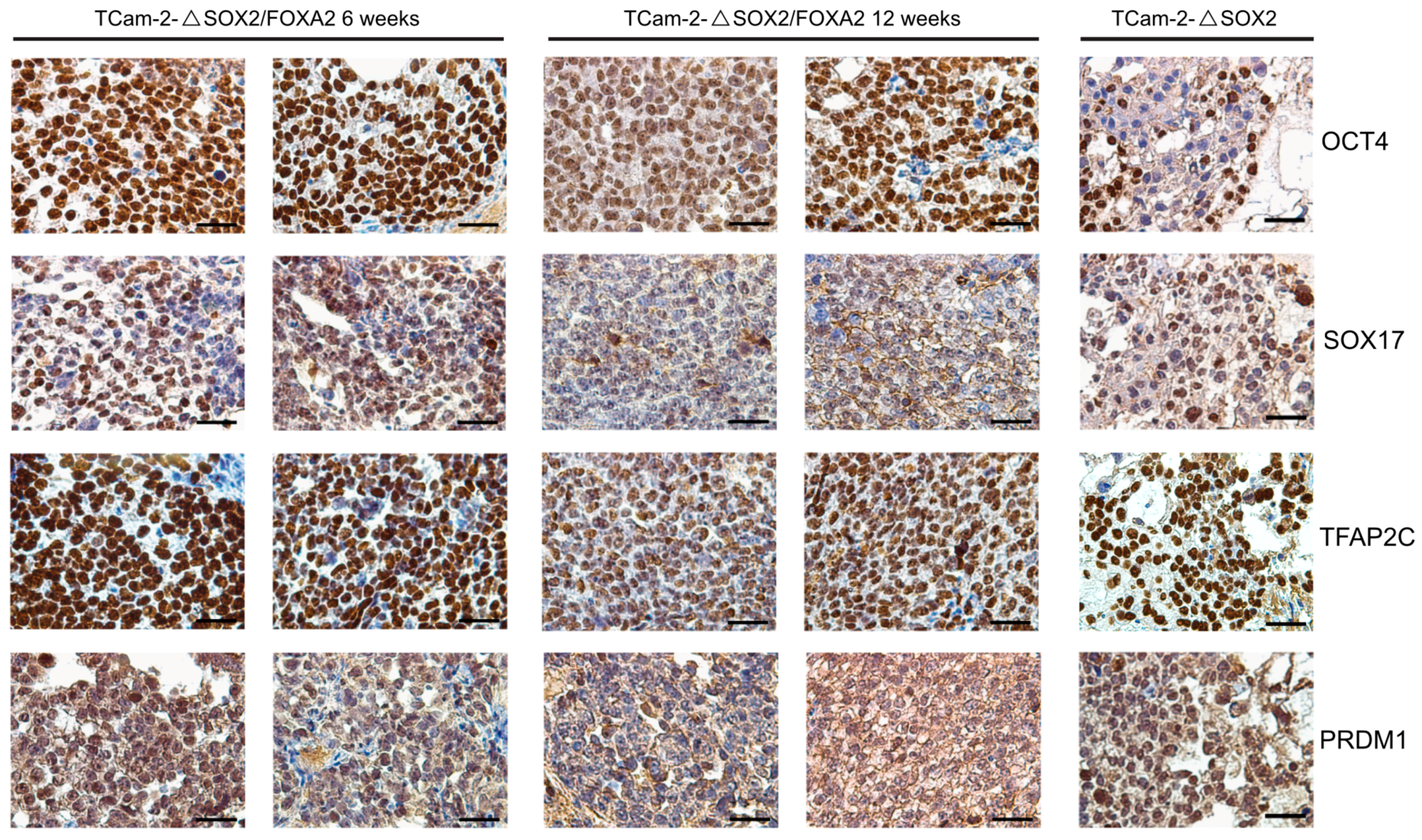
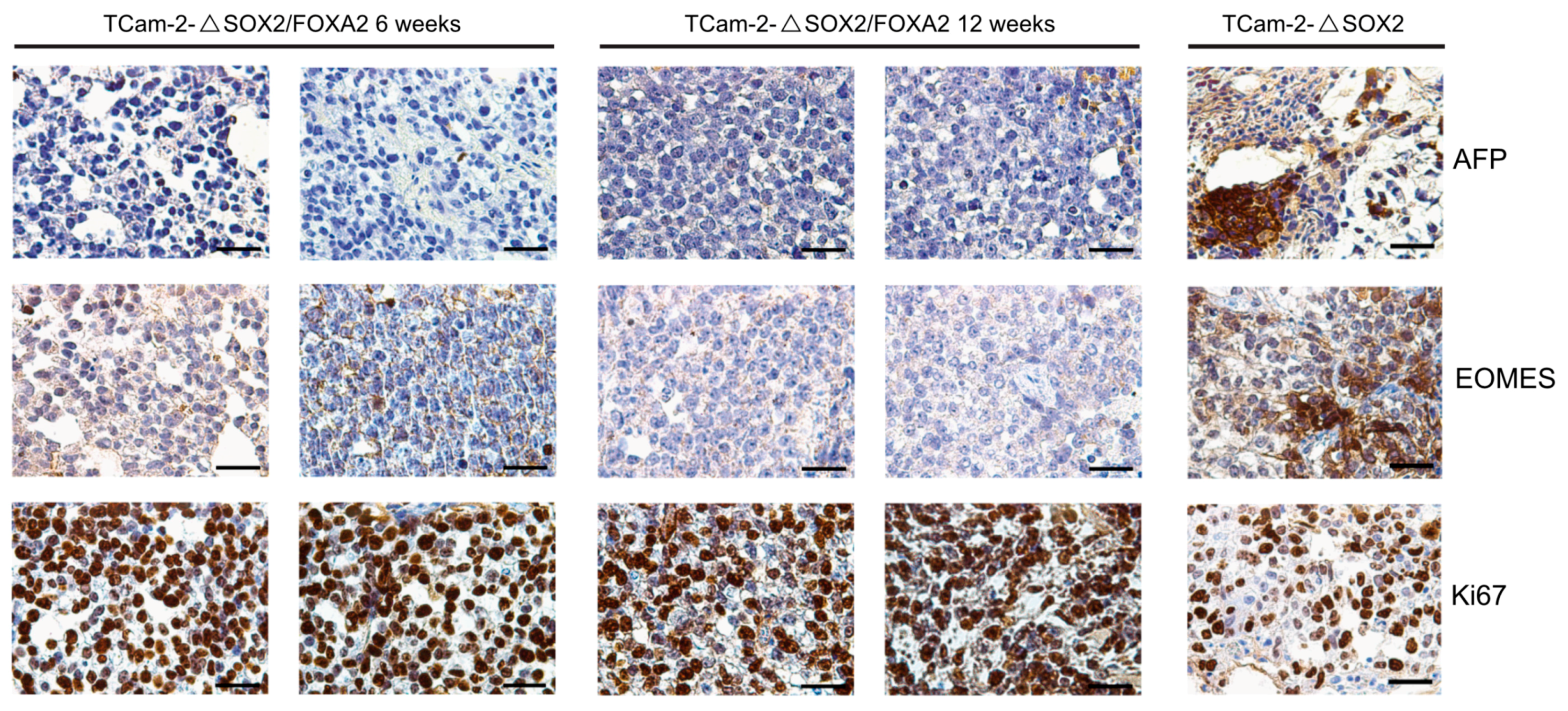
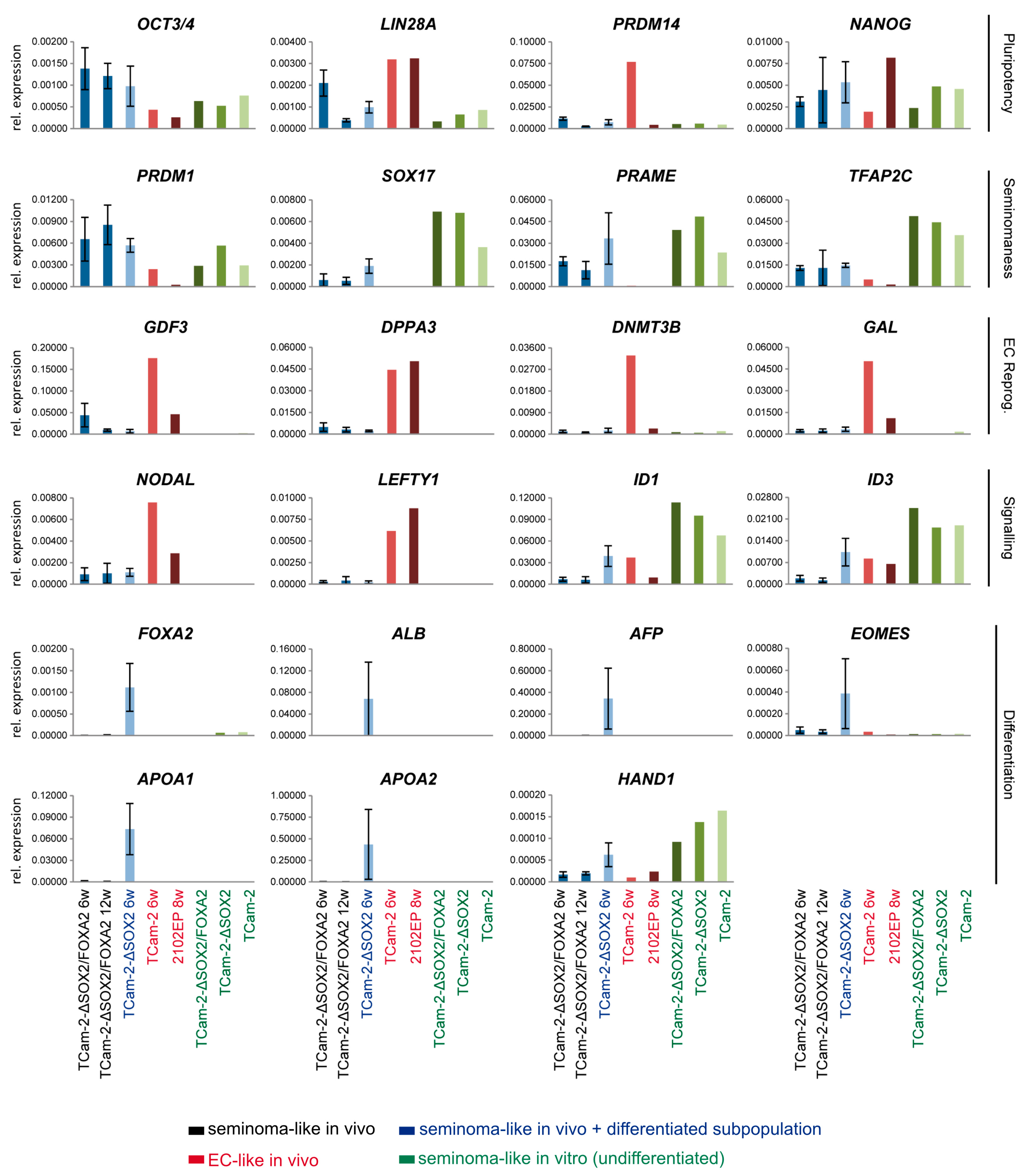
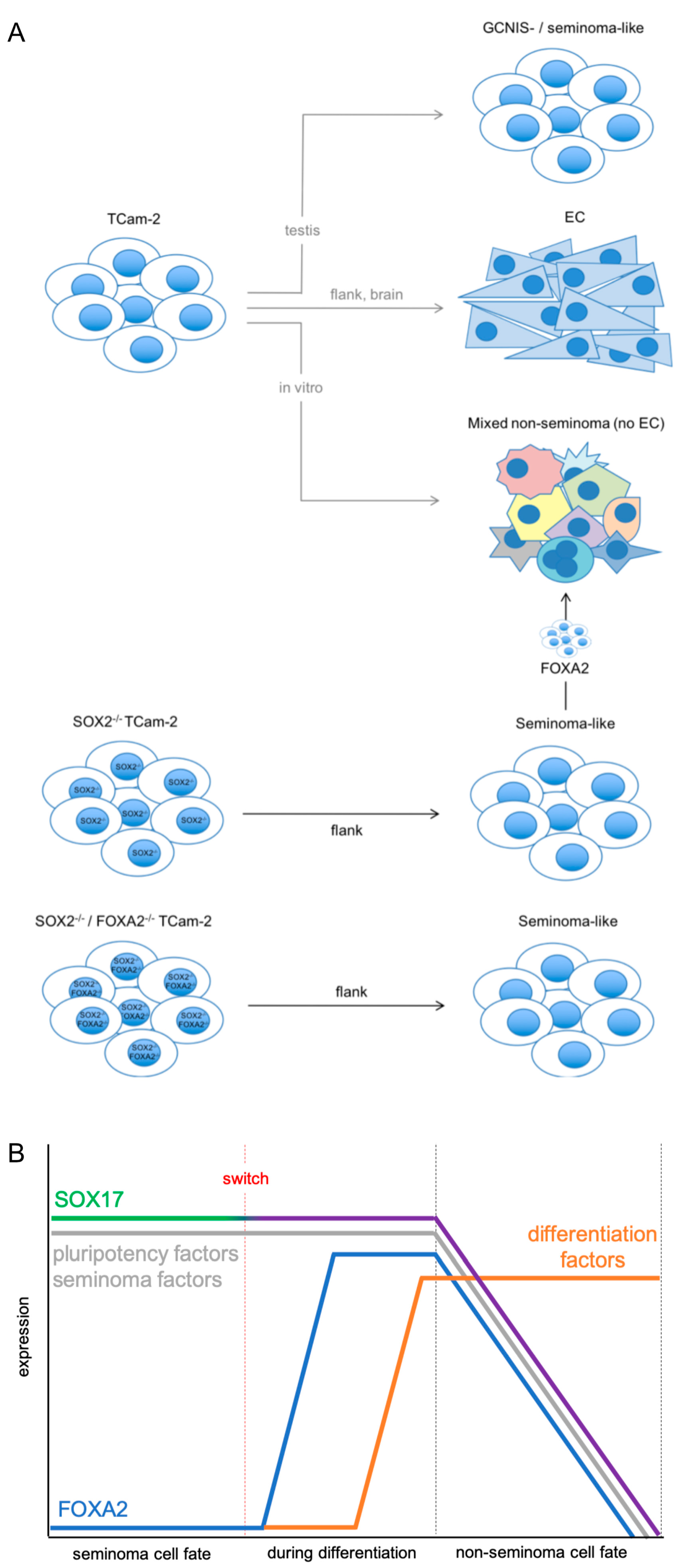
2. Results
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Cell Culture
4.3. Generation of FOXA2-Deficient TCam-2 Cells
4.4. DNA, RNA and Protein Isolation
4.5. Western Blot
4.6. Hematoxylin and Eosin Staining
4.7. Immunohistochemistry
4.8. Quantitative RT-PCR
4.9. Measurement of Proliferation Rates
4.10. Xenotransplantation
4.11. Illumina HT-12v4 and Affymetrix Expression Arrays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oosterhuis, J.W.; Looijenga, L.H.J. Testicular germ-cell tumours in a broader perspective. Nat. Rev. Cancer 2005, 5, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Berney, D.M.; Looijenga, L.H.; Idrees, M.; Oosterhuis, J.W.; Rajpert-De Meyts, E.; Ulbright, T.M.; Skakkebaek, N.E. Germ cell neoplasia in situ (GCNIS): Evolution of the current nomenclature for testicular pre-invasive germ cell malignancy. Histopathology 2016, 69, 7–10. [Google Scholar] [CrossRef]
- Eckert, D.; Nettersheim, D.; Heukamp, L.C.; Kitazawa, S.; Biermann, K.; Schorle, H. TCam-2 but not JKT-1 cells resemble seminoma in cell culture. Cell Tissue Res. 2008, 331, 529–538. [Google Scholar] [CrossRef]
- Nettersheim, D.; Westernströer, B.; Haas, N.; Leinhaas, A.; Brüstle, O.; Schlatt, S.; Schorle, H. Establishment of a versatile seminoma model indicates cellular plasticity of germ cell tumor cells. Genes Chromosom. Cancer 2012, 51, 717–726. [Google Scholar] [CrossRef] [PubMed]
- De Jong, J.; Stoop, H.; Gillis, A.J.; Hersmus, R.; Van Gurp, R.J.; Van De Geijn, G.J.; Van Drunen, E.; Beverloo, H.B.; Schneider, D.T.; Sherlock, J.K.; et al. Further characterization of the first seminoma cell line TCam-2. Genes Chromosom. Cancer 2008, 47, 185–196. [Google Scholar] [CrossRef]
- Irie, N.; Weinberger, L.; Tang, W.W.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 is a critical specifier of human primordial germ cell fate. Cell 2015, 160, 253–268. [Google Scholar] [CrossRef]
- Nettersheim, D.; Jostes, S.; Sharma, R.; Schneider, S.; Hofmann, A.; Ferreira, H.J.; Hoffmann, P.; Kristiansen, G.; Esteller, M.B.; Schorle, H. BMP Inhibition in Seminomas Initiates Acquisition of Pluripotency via NODAL Signaling Resulting in Reprogramming to an Embryonal Carcinoma. PLoS Genet. 2015, 11, e1005415. [Google Scholar] [CrossRef]
- Nettersheim, D.; Heukamp, L.C.; Fronhoffs, F.; Grewe, M.J.; Haas, N.; Waha, A.; Honecker, F.; Waha, A.; Kristiansen, G.; Schorle, H. Analysis of TET Expression/Activity and 5mC oxidation during normal and malignant germ cell development. PLoS ONE 2013, 8, e82881. [Google Scholar] [CrossRef] [PubMed]
- Nettersheim, D.; Heimsoeth, A.; Jostes, S.; Schneider, S.; Fellermeyer, M.; Hofmann, A.; Schorle, H. SOX2 is essential for in vivo reprogramming of seminoma-like TCam-2 cells to an embryonal carcinoma-like fate. Oncotarget 2016, 7, 47095–47110. [Google Scholar] [CrossRef] [PubMed]
- Kaestner, K.H. The FoxA factors in organogenesis and differentiation. Curr. Opin. Genet. Dev. 2010, 20, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerschner, J.L.; Gosalia, N.; Leir, S.H.; Harris, A. Chromatin remodeling mediated by the FOXA1/A2 transcription factors activates CFTR expression in intestinal epithelial cells. Epigenetics 2014, 9, 557–565. [Google Scholar] [CrossRef]
- Friedman, J.R.; Kaestner, K.H. The Foxa family of transcription factors in development and metabolism. Cell. Mol. Life Sci. 2006, 63, 2317–2328. [Google Scholar] [CrossRef]
- Sekiya, T.; Muthurajan, U.M.; Luger, K.; Tulin, A.V.; Zaret, K.S. Nucleosome-binding affinity as a primary determinant of the nuclear mobility of the pioneer transcription factor FoxA. Genes Dev. 2009, 23, 804–809. [Google Scholar] [CrossRef] [Green Version]
- Gosalia, N.; Yang, R.; Kerschner, J.L.; Harris, A. FOXA2 regulates a network of genes involved in critical functions of human intestinal epithelial cells. Physiol. Genom. 2015, 47, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, J.A.; Zhang, C.; Klein-Szanto, A.J.; Kormish, J.D.; Fu, J.; Zhang, M.Q.; Zaret, K.S. Study of foxA pioneer factor at silent genes reveals Rfx-repressed enhancer at Cdx2 and a potential indicator of esophageal adenocarcinoma development. PLoS Genet. 2011, 7, e1002277. [Google Scholar] [CrossRef]
- Park, J.W.; Lee, J.K.; Witte, O.N.; Huang, J. FOXA2 is a sensitive and specific marker for small cell neuroendocrine carcinoma of the prostate. Mod. Pathol. 2017, 30, 1262–1272. [Google Scholar] [CrossRef]
- Nettersheim, D.; Gillis, A.J.M.; Looijenga, L.H.J.; Schorle, H. TGF-β1, EGF and FGF4 synergistically induce differentiation of the seminoma cell line TCam-2 into a cell type resembling mixed non-seminoma. Int. J. Androl. 2011, 34, e189–e203. [Google Scholar] [CrossRef] [PubMed]
- Sinner, D.; Kordich, J.J.; Spence, J.R.; Opoka, R.; Rankin, S.; Lin, S.-C.J.; Jonatan, D.; Zorn, A.M.; Wells, J.M. Sox17 and Sox4 Differentially Regulate-Catenin/T-Cell Factor Activity and Proliferation of Colon Carcinoma Cells. Mol. Cell Biol. 2007, 27, 7802–7815. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Rodriguez, R.T.; Wang, J.; Ghodasara, A.; Kim, S.K. Targeting SOX17 in human embryonic stem cells creates unique strategies for isolating and analyzing developing endoderm. Cell Stem Cell 2011, 8, 335–346. [Google Scholar] [CrossRef]
- Aksoy, I.; Jauch, R.; Chen, J.; Dyla, M.; Divakar, U.; Bogu, G.K.; Teo, R.; Leng Ng, C.K.; Herath, W.; Lili, S.; et al. Oct4 switches partnering from Sox2 to Sox17 to reinterpret the enhancer code and specify endoderm. EMBO J. 2013, 32, 938–953. [Google Scholar] [CrossRef] [Green Version]
- de Jong, J.; Stoop, H.; Gillis, A.J.; van Gurp, R.J.; van de Geijn, G.J.; Boer, M.; Hersmus, R.; Saunders, P.T.; Anderson, R.A.; Oosterhuis, J.W.; et al. Differential expression of SOX17 and SOX2 in germ cells and stem cells has biological and clinical implications. J. Pathol. 2008, 215, 21–30. [Google Scholar] [CrossRef]
- Nettersheim, D.; Jostes, S.; Schorle, H. Xenografting of cancer cell lines for in vivo screening of the therapeutic potential of HDAC inhibitors. Methods Mol. Biol. 2017, 1510, 211–215. [Google Scholar] [PubMed]
- Nettersheim, D.; Jostes, S.; Fabry, M.; Honecker, F.; Schumacher, V.; Kirfel, J.; Kristiansen, G.; Schorle, H. A signaling cascade including ARID1A, GADD45B and DUSP1 induces apoptosis and affects the cell cycle of germ cell cancers after romidepsin treatment. Oncotarget 2016, 7, 74931–74946. [Google Scholar] [CrossRef] [PubMed]
- Jostes, S.; Nettersheim, D.; Fellermeyer, M.; Schneider, S.; Hafezi, F.; Honecker, F.; Schumacher, V.; Geyer, M.; Kristiansen, G.; Schorle, H. The bromodomain inhibitor JQ1 triggers growth arrest and apoptosis in testicular germ cell tumours in vitro and in vivo. J. Cell Mol. Med. 2017, 21, 1300–1314. [Google Scholar] [CrossRef]
- Nettersheim, D.; Arndt, I.; Sharma, R.; Riesenberg, S.; Jostes, S.; Schneider, S.; Hölzel, M.; Kristiansen, G.; Schorle, H. The cancer/testis-antigen PRAME supports the pluripotency network and represses somatic and germ cell differentiation programs in seminomas. Br. J. Cancer 2016, 115, 454–464. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| AFP | GTGGTCAGTTTGCAGCATTC | GCAGAGGAGATGTGCTGGAT |
| ALB | TCAGCCATTTCACCATAGGTT | TGCTGATGAGTCAGCTGAAA |
| APOA1 | CCCAGTTGTCAAGGAGCTTT | TGGATGTGCTCAAAGACAGC |
| APOA2 | AGTTCCGTTCCAGCCTTCTT | GACCGTGACTGACTATGGCA |
| DNMT3B | CCAGCTCTTACCTTACCATC | CAGACATAGCCTGTCGCTTG |
| DPPA3 | TCAACGTCTCGGAGGAGATT | CAACCTACATCCCAGGGTCT |
| EOMES | CACATTGTAGTGGGCAGTGG | CGCCACCAAACTGAGATGAT |
| FOXA2 | TACGTGTTCATGCCGTTCAT | CGACTGGAGCAGCTACTATGC |
| GAL | CTGGTGAGGCCATTCTTGTC | AAGGAAAAACGAGGCTGGA |
| GAPDH | TGCCAAATATGATGACATCAAGA | GGAGTGGGTGTCGCTGTTG |
| GDF3 | CAGGAGGAAGCTGGGAAAT | TGCTACGTAAAGGAGCTGGG |
| ID1 | TCCAGCACGTCATCGACTAC | TCAGCGACACAAGATGCG |
| ID3 | TCAGCTTAGCCAGGTGGAAATC | TGGCTCGGCCAGGACTAC |
| LEFTY1 | TTGGGGACTATGGAGCTCAG | TCAAGTCCCTCGATGGCTAC |
| LIN28A | ACCCTTCCATGTGCAGCTTA | TGTAAGTGGTTCAACGTGCG |
| NANOG | ATGGAGGAGGGAAGAGGAGA | GATTTGTGGGCCTGAAGAAA |
| NODAL | ATGCCAGATCCTCTTGTTGG | AGACATCATCCGCAGCCTAC |
| OCT3/4 | GGGAGATTGATAACTGGTGTGTT | GTGTATATCCCAGGGTGATCCTC |
| PRAME | CGTAGACTCCTCCTCTCCCACAT | TGGGCGATATACTGCTCTTCCT |
| PRDM1 | GGGTGCAGCCTTTATGAGTC | CCTTGTTCATGCCCTGAGAT |
| PRDM14 | TCCACACAGGGGGTGTACTT | GAGCCTTCAGGTCACAGAGC |
| SOX17 | GGCGCAGCAGAATCCAGA | CCACGACTTGCCCAGCAT |
| TFAP2C | GGCCCAGCAACTGTGTAAAGA | GCAGTTCTGTATGTTCGTCTCCA |
| FOXA2 genotyping 1 | CCAGGGAGAGAGAGGGAGT | CCTCGGGCTCTGCATAGTAG |
| FOXA2 genotyping 2 | CTCGCTCTCCTTCAACGACT | TCTTCTCCCTTGCGTCTCTG |
| FOXA2 genotyping 3 | TTAAACTGCCATGCACTCGG | GGGAGTACACCCCCTGGTAG |
| FOXA2 guideRNA1 | AAGGGCACGAGCCGTCCGAC | |
| FOXA2 guideRNA2 | GTAGTGCATCACCTGTTCGT | |
| FOXA2 guideRNA3 | CATGAACATGTCGTCGTACG |
| Primary Antibodies | |||||
| Target | Company | Number | Species | IHC | Western Blot |
| AFP | Dako | A0008 | Rabbit | 1:250 | - |
| β-Actin | Merck | A5441 | Mouse | - | 1:25000 |
| EOMES | Abcam | ab23345 | Rabbit | 1:200 | - |
| FOXA2 | R&D systems | AF2400 | Goat | 1:200 | 1:500 |
| Ki67 | Zytomed Systems | MSK018 | Mouse | 1:500 | - |
| OCT3/4 | Santa Cruz | C-10 | Mouse | 1:200 | - |
| PRAME | Santa Cruz | H-10 | Mouse | - | 1:400 |
| PRDM1 | H.M. Jäck | - | Rabbit | 1:200 | - |
| SOX2 | R&D systems | MAB2018 | Mouse | 1:200 | 1:200 |
| SOX17 | Abcam | ab84990 | Mouse | 1:400 | - |
| SOX17 | R&D systems | AF1924 | Goat | - | 1:1000 |
| TFAP2C | Santa Cruz | 6E4/4 | Mouse | 1:200 | - |
| Secondary Antibodies | |||||
| Target | Company | Number | Species | IHC | Western Blot |
| Rabbit Anti-Goat HRP | Dako | P0160 | Rabbit | - | 1:2000 |
| Rabbit Anti-Mouse HRP | Dako | P0260 | Rabbit | - | 1:1000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nettersheim, D.; Vadder, S.; Jostes, S.; Heimsoeth, A.; Schorle, H. TCam-2 Cells Deficient for SOX2 and FOXA2 Are Blocked in Differentiation and Maintain a Seminoma-Like Cell Fate In Vivo. Cancers 2019, 11, 728. https://doi.org/10.3390/cancers11050728
Nettersheim D, Vadder S, Jostes S, Heimsoeth A, Schorle H. TCam-2 Cells Deficient for SOX2 and FOXA2 Are Blocked in Differentiation and Maintain a Seminoma-Like Cell Fate In Vivo. Cancers. 2019; 11(5):728. https://doi.org/10.3390/cancers11050728
Chicago/Turabian StyleNettersheim, Daniel, Saskia Vadder, Sina Jostes, Alena Heimsoeth, and Hubert Schorle. 2019. "TCam-2 Cells Deficient for SOX2 and FOXA2 Are Blocked in Differentiation and Maintain a Seminoma-Like Cell Fate In Vivo" Cancers 11, no. 5: 728. https://doi.org/10.3390/cancers11050728