
Neurofeedback-Augmented Mindfulness Training Elicits Distinct Responses in the Subregions of the Insular Cortex in Healthy Adolescents

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
1.1. The Role of Interoception in Mindfulness and Its Key Hub Insula
1.2. Real-Time Functional Magnetic Resonance Imaging Neurofeedback-Augmented MT Targeting the Posterior Cingulate Cortex
1.3. The Present Study
2. Materials and Methods
2.1. Participants
2.2. Experimental Procedures
2.2.1. Neurofeedback-Augmented Mindfulness Training Task (NAMT)
2.2.2. Psychological Measurements
2.2.3. Data Acquisition
2.2.4. Data Processing and Analysis
2.2.5. Data and Code Availability Statement
3. Results
3.1. Demographic, Task, and Clinical Characteristics
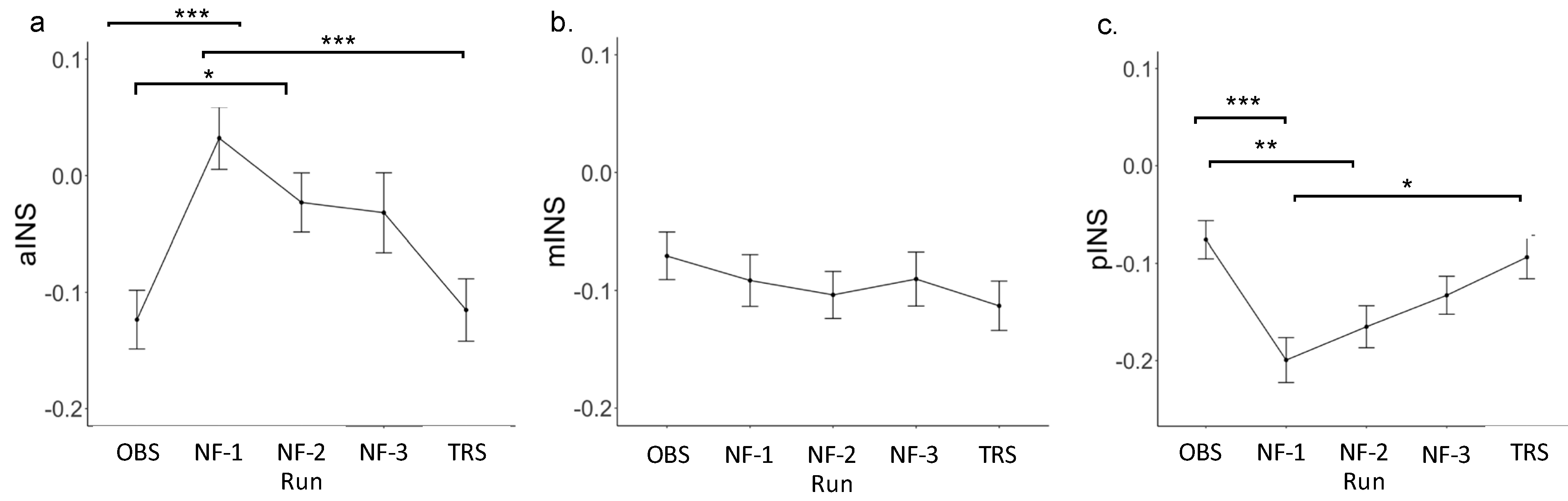
3.2. Insula Region of Interest (ROI) Results
3.3. Self-Reported Questionnaire Responses and Insula ROI Results
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Neurofeedback-Augmented Mindfulness Training Task (NAMT)
Appendix A.2. Real-Time fMRI Processing
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Domain | Item # | Checklist Item | Reported on Page # |
|---|---|---|---|
| Pre-experiment | |||
| 1a | Pre-register experimental protocol and planned analyses | n/a | |
| 1b | Justify sample size | 4 | |
| Control groups | |||
| 2a | Employ control group(s) or control condition(s) | 5 | |
| 2b | When leveraging experimental designs where a double-blind is possible, use a double-blind | n/a | |
| 2c | Blind those who rate the outcomes, and when possible, the statisticians involved | n/a | |
| 2d | Examine to what extent participants and experimenters remain blinded | n/a | |
| 2e | In clinical efficacy studies, employ a standard-of-care intervention group as a benchmark for improvement | n/a | |
| Control measures | |||
| 3a | Collect data on psychosocial factors | 5–6 | |
| 3b | Report whether participants were provided with a strategy | 5 | |
| 3c | Report the strategies participants used | n/a | |
| 3d | Report methods used for online-data processing and artifact correction | 6–7 | |
| 3e | Report condition and group effects for artifacts | n/a | |
| Feedback specifications | |||
| 4a | Report how the online-feature extraction was defined | 6 | |
| 4b | Report and justify the reinforcement schedule | n/a | |
| 4c | Report the feedback modality and content | 4–5 | |
| 4d | Collect and report all brain activity variable(s) and/or contrasts used for feedback, as displayed to experimental participants | 4–5 | |
| 4e | Report the hardware and software used | 6–9 | |
| Outcome measures | |||
| Brain | 5a | Report neurofeedback regulation success based on the feedback signal | n/a |
| 5b | Plot within-session and between-session regulation blocks of feedback variable(s), as well as pre-to-post resting baselines or contrasts | n/a | |
| 5c | Statistically compare the experimental condition/group to the control condition(s)/group(s) (not only each group to baseline measures) | n/a | |
| Behavior | 6a | Include measures of clinical or behavioral significance, defined a priori, and describe whether they were reached | n/a |
| 6b | Run correlational analyses between regulation success and behavioral outcomes | n/a | |
| Data storage | |||
| 7a | Upload all materials, analysis scripts, code, and raw data used for analyses, as well as final values, to an open access data repository, when feasible | n/a | |
| Gyrus | ROI | Label ID.L | Label ID.R | Anatomical and Modified Cyto-Architectonic Descriptions | Left Hemisphere MNI Coordinates | Right Hemisphere MNI Coordinates |
|---|---|---|---|---|---|---|
| Insular Gyrus | Anterior Insula | 165 | 166 | vIa, ventral agranular insula | −32, 14, −13 | 33, 14, −13 |
| 167 | 168 | dIa, dorsal agranular insula | −34, 18, 1 | 36, 18, 1 | ||
| Mid-Insula | 169 | 170 | vId/vIg, ventral dysgranular and granular insula | −38, −4, −9 | 39, −2, −9 | |
| 173 | 174 | dId, dorsal dysgranular insula | −38, 5, 5 | 38, 5, 5 | ||
| Posterior Insula | 163 | 164 | G, hypergranular insula | −36, −20, 10 | 37, −18, 8 | |
| 171 | 172 | dIg, dorsal granular insula | −38, −8, 8 | 39, −7, 8 |
| Run | Mean | SD | Estimate | SE | t | p | Cohen’s d |
|---|---|---|---|---|---|---|---|
| Anterior insula | |||||||
| OBS | −0.12 | 0.17 | |||||
| NF-1 | 0.03 | 0.18 | 0.16 | 0.04 | 4.40 | <0.001 | 0.74 |
| NF-2 | −0.02 | 0.17 | 0.10 | 0.04 | 2.74 | <0.01 | 0.46 |
| NF-3 | −0.03 | 0.23 | 0.09 | 0.04 | 2.59 | <0.05 | 0.43 |
| TRS | −0.12 | 0.18 | 0.01 | 0.04 | 0.23 | 0.81 | 0.04 |
| Mid-insula | |||||||
| OBS | −0.07 | 0.14 | |||||
| NF-1 | −0.09 | 0.15 | −0.02 | 0.03 | −0.75 | 0.45 | −0.13 |
| NF-2 | −0.10 | 0.14 | −0.03 | 0.03 | −1.22 | 0.23 | −0.20 |
| NF-3 | −0.09 | 0.16 | −0.02 | 0.03 | −0.71 | 0.48 | −0.12 |
| TRS | −0.11 | 0.14 | −0.04 | 0.03 | −1.53 | 0.13 | −0.26 |
| Posterior insula | |||||||
| OBS | −0.08 | 0.13 | |||||
| NF-1 | −0.20 | 0.16 | 0.03 | 0.03 | 1.11 | 0.27 | 0.19 |
| NF-2 | −0.17 | 0.15 | 0.07 | 0.03 | 2.25 | <0.05 | 0.38 |
| NF-3 | −0.13 | 0.13 | 0.12 | 0.03 | 4.16 | <0.001 | 0.70 |
| TRS | −0.09 | 0.15 | 0.11 | 0.03 | 3.56 | <0.001 | 0.60 |
| Run | Estimate | Std. Error | z Statistic | p Value |
|---|---|---|---|---|
| Anterior insular cortex (aINS) | ||||
| NF-1:OBS | 0.16 | 0.04 | 4.40 | <0.001 |
| NF-2:OBS | 0.10 | 0.04 | 2.74 | 0.05 |
| NF-3:OBS | 0.09 | 0.04 | 2.59 | 0.07 |
| TR:OBS | 0.01 | 0.04 | 0.23 | 1.00 |
| NF-2:NF-1 | −0.06 | 0.04 | −1.63 | 0.48 |
| NF-3:NF-1 | −0.06 | 0.04 | −1.81 | 0.37 |
| TR:NF-1 | −0.15 | 0.04 | −4.17 | <0.001 |
| NF-3:NF-2 | −0.01 | 0.04 | −0.17 | 1.00 |
| TR:NF-2 | −0.09 | 0.04 | −2.50 | 0.09 |
| TR:NF-3 | −0.08 | 0.04 | −2.36 | 0.13 |
| Mid-insular cortex (mINS) | ||||
| NF-1:OBS | −0.02 | 0.03 | −0.75 | 0.94 |
| NF-2:OBS | −0.03 | 0.03 | −1.22 | 0.74 |
| NF-3:OBS | −0.02 | 0.03 | −0.71 | 0.96 |
| TR:OBS | −0.04 | 0.03 | −1.53 | 0.54 |
| NF-2:NF-1 | −0.01 | 0.03 | −0.47 | 0.99 |
| NF-3:NF-1 | 0.00 | 0.03 | 0.04 | 1.00 |
| TR:NF-1 | −0.02 | 0.03 | −0.78 | 0.94 |
| NF-3:NF-2 | 0.01 | 0.03 | 0.52 | 0.99 |
| TR:NF-2 | −0.01 | 0.03 | −0.30 | 1.00 |
| TR:NF-3 | −0.02 | 0.03 | −0.82 | 0.92 |
| Posterior insular cortex (pINS) | ||||
| NF-1:OBS | −0.12 | 0.03 | −4.16 | <0.001 |
| NF-2:OBS | −0.09 | 0.03 | −3.02 | <0.05 |
| NF-3:OBS | −0.06 | 0.03 | −1.92 | 0.31 |
| TR:OBS | −0.02 | 0.03 | −0.60 | 0.98 |
| NF-2:NF-1 | 0.03 | 0.03 | 1.11 | 0.80 |
| NF-3:NF-1 | 0.07 | 0.03 | 2.25 | 0.16 |
| TR:NF-1 | 0.11 | 0.03 | 3.56 | <0.01 |
| NF-3:NF-2 | 0.03 | 0.03 | 1.12 | 0.80 |
| TR:NF-2 | 0.07 | 0.03 | 2.43 | 0.11 |
| TR:NF-3 | 0.04 | 0.03 | 1.32 | 0.68 |
| Neurofeedback Runs | Observe Run | Transfer Run | |||||||
|---|---|---|---|---|---|---|---|---|---|
| aINS | mINS | pINS | aINS | mINS | pINS | aINS | mINS | pINS | |
| PROMIS Positive Affect | −0.2 | −0.15 | −0.1 | −0.24 | −0.16 | −0.19 | 0.08 | 0.15 | 0.16 |
| PROMIS Meaning and Purpose | −0.23 | −0.24 | −0.06 | −0.23 | −0.09 | −0.19 | 0.07 | 0.09 | 0.22 |
| PROMIS Life Satisfaction | −0.37 * | −0.32 | −0.19 | −0.21 | −0.2 | −0.17 | 0.06 | 0.00 | 0.11 |
| PROMIS Pain Interference | 0.2 | 0.18 | 0.29 | 0.12 | 0.03 | 0.15 | 0.15 | 0.09 | 0.12 |
| PROMIS Pain Behavior | 0.18 | 0.32 | 0.33 * | −0.09 | −0.02 | −0.04 | 0.13 | 0.20 | 0.18 |
| PROMIS Fatigue | 0.28 | 0.3 | 0.23 | 0.27 | 0.21 | 0.29 | 0.06 | 0.05 | 0.10 |
| Task Ratings Current Feeling | −0.13 | −0.22 | −0.25 | 0.01 | −0.25 | −0.18 | −0.43 ** | −0.37 * | −0.19 |
| Task Ratings Mind Wander | −0.31 | −0.36 | −0.24 | −0.11 | −0.24 | −0.34 * | −0.20 | −0.28 | −0.22 |
| Task Ratings Focus-on-Breath | −0.03 | −0.14 | −0.18 | −0.14 | −0.17 | −0.04 | 0.11 | 0.26 | 0.16 |



References
- Kabat-Zinn, J. Mindfulness. Mindfulness 2015, 6, 1481–1483. [Google Scholar] [CrossRef]
- Tomlinson, E.R.; Yousaf, O.; Vittersø, A.D.; Jones, L. Dispositional Mindfulness and Psychological Health: A Systematic Review. Mindfulness 2018, 9, 23–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Langberg, J.M.; Becker, S.P. Dispositional mindfulness moderates the relation between brooding rumination and sleep problems in adolescents. Sleep Med. 2022, 90, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Khoury, B.; Sharma, M.; Rush, S.E.; Fournier, C. Mindfulness-Based stress reduction for healthy individuals: A meta-analysis. J. Psychosom. Res. 2015, 78, 519–528. [Google Scholar] [CrossRef]
- Guo, D.; Sun, L.; Yu, X.; Liu, T.; Wu, L.; Sun, Z.; Zhang, F.; Zhou, Y.; Shen, M.; Liu, W. Mindfulness-based stress reduction improves the general health and stress of Chinese military recruits: A pilot study. Psychiatry Res. 2019, 281, 112571. [Google Scholar] [CrossRef]
- Blanck, P.; Perleth, S.; Heidenreich, T.; Kröger, P.; Ditzen, B.; Bents, H.; Mander, J. Effects of mindfulness exercises as stand-alone intervention on symptoms of anxiety and depression: Systematic review and meta-analysis. Behav. Res. Ther. 2018, 102, 25–35. [Google Scholar] [CrossRef]
- Goldberg, S.B.; Tucker, R.P.; Greene, P.A.; Davidson, R.J.; Wampold, B.E.; Kearney, D.J.; Simpson, T.L. Mindfulness-Based interventions for psychiatric disorders: A systematic review and meta-analysis. Clin. Psychol. Rev. 2018, 59, 52–60. [Google Scholar] [CrossRef]
- Raj, S.; Ghosh, D.; Verma, S.K.; Singh, T. The mindfulness trajectories of addressing suicidal behaviour: A systematic review. Int. J. Soc. Psychiatry 2021, 67, 507–519. [Google Scholar] [CrossRef]
- Essau, C.A.; Lewinsohn, P.M.; Lim, J.X.; Moon-ho, R.H.; Rohde, P. Incidence, recurrence and comorbidity of anxiety disorders in four major developmental stages. J. Affect. Disord. 2018, 228, 248–253. [Google Scholar] [CrossRef] [Green Version]
- Keyes, K.M.; Gary, D.; O’Malley, P.M.; Hamilton, A.; Schulenberg, J. Recent increases in depressive symptoms among US adolescents: Trends from 1991 to 2018. Soc. Psychiatry Psychiatr. Epidemiol. 2019, 54, 987–996. [Google Scholar] [CrossRef]
- Burstein, B.; Agostino, H.; Greenfield, B. Suicidal attempts and ideation among children and adolescents in US emergency departments, 2007–2015. JAMA Pediatrics 2019, 173, 598–600. [Google Scholar] [CrossRef] [PubMed]
- Sanger, K.L.; Thierry, G.; Dorjee, D. Effects of school-based mindfulness training on emotion processing and well-being in adolescents: Evidence from event-related potentials. Dev. Sci. 2018, 21, e12646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.B.; Martin, G. Mind full or mindful: A report on mindfulness and psychological health in healthy adolescents. Int. J. Adolesc. Youth 2016, 21, 64–74. [Google Scholar] [CrossRef]
- Gibson, J. Mindfulness, Interoception, and the Body: A Contemporary Perspective. Front. Psychol. 2019, 10, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, L.; Thom, N.J.; Shukla, A.; Davenport, P.W.; Simmons, A.N.; Stanley, E.A.; Paulus, M.P.; Johnson, D.C. Mindfulness-based training attenuates insula response to an aversive interoceptive challenge. Soc. Cogn. Affect. Neurosci. 2015, 11, 182–190. [Google Scholar] [CrossRef] [Green Version]
- De Jong, M.; Lazar, S.W.; Hug, K.; Mehling, W.E.; Hölzel, B.K.; Sack, A.T.; Peeters, F.; Ashih, H.; Mischoulon, D.; Gard, T. Effects of mindfulness-based cognitive therapy on body awareness in patients with chronic pain and comorbid depression. Front. Psychol. 2016, 7, 967. [Google Scholar] [CrossRef] [Green Version]
- Bornemann, B.; Singer, T. Taking time to feel our body: Steady increases in heartbeat perception accuracy and decreases in alexithymia over 9 months of contemplative mental training. Psychophysiology 2017, 54, 469–482. [Google Scholar] [CrossRef]
- Khalsa, S.S.; Adolphs, R.; Cameron, O.G.; Critchley, H.D.; Davenport, P.W.; Feinstein, J.S.; Feusner, J.D.; Garfinkel, S.N.; Lane, R.D.; Mehling, W.E.; et al. Interoception and Mental Health: A Roadmap. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 501–513. [Google Scholar] [CrossRef]
- Hölzel, B.K.; Lazar, S.W.; Gard, T.; Schuman-Olivier, Z.; Vago, D.R.; Ott, U. How does mindfulness meditation work? Proposing mechanisms of action from a conceptual and neural perspective. Perspect. Psychol. Sci. 2011, 6, 537–559. [Google Scholar] [CrossRef]
- Farb, N.; Daubenmier, J.; Price, C.J.; Gard, T.; Kerr, C.; Dunn, B.D.; Klein, A.C.; Paulus, M.P.; Mehling, W.E. Interoception, contemplative practice, and health. Front. Psychol. 2015, 6, 763. [Google Scholar] [CrossRef] [Green Version]
- Mehling, W.E.; Price, C.; Daubenmier, J.J.; Acree, M.; Bartmess, E.; Stewart, A. The multidimensional assessment of interoceptive awareness (MAIA). PLoS ONE 2012, 7, e48230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehling, W.E.; Chesney, M.A.; Metzler, T.J.; Goldstein, L.A.; Maguen, S.; Geronimo, C.; Agcaoili, G.; Barnes, D.E.; Hlavin, J.A.; Neylan, T.C. A 12-week integrative exercise program improves self-reported mindfulness and interoceptive awareness in war veterans with posttraumatic stress symptoms. J. Clin. Psychol. 2018, 74, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Fissler, M.; Winnebeck, E.; Schroeter, T.; Gummersbach, M.; Huntenburg, J.M.; Gaertner, M.; Barnhofer, T. An investigation of the effects of brief mindfulness training on self-reported interoceptive awareness, the ability to decenter, and their role in the reduction of depressive symptoms. Mindfulness 2016, 7, 1170–1181. [Google Scholar] [CrossRef]
- Fogel, A. The Psychophysiology of Self-Awareness: Rediscovering the Lost Art of Body Sense; WW Norton & Co.: New York, NY, USA, 2009. [Google Scholar]
- Craig, A.D.; Craig, A.D. How do you feel? Interoception: The sense of the physiological condition of the body. Nat. Rev. Neurosci. 2002, 3, 655–666. [Google Scholar] [CrossRef]
- Critchley, H.D.; Wiens, S.; Ohman, P.R.A.; Dolan, R.J. Neural systems supporting interoceptive awareness. Psychol. Sect. Karolinska Hosp. 2004, 7, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogolla, N. The insular cortex. Curr. Biol. 2017, 27, R580–R586. [Google Scholar] [CrossRef] [Green Version]
- Khalsa, S.S.; Rudrauf, D.; Feinstein, J.S.; Tranel, D. The pathways of interoceptive awareness. Nat. Neurosci. 2009, 12, 1494–1496. [Google Scholar] [CrossRef] [Green Version]
- Khalsa, S.S.; Rudrauf, D.; Damasio, A.R.; Davidson, R.J.; Lutz, A.; Tranel, D. Interoceptive awareness in experienced meditators. Psychophysiology 2008, 45, 671–677. [Google Scholar] [CrossRef] [Green Version]
- Paulus, M.P.; Stein, M.B. An insular view of anxiety. Biol. Psychiatry 2006, 60, 383–387. [Google Scholar] [CrossRef]
- Ives-Deliperi, V.L.; Solms, M.; Meintjes, E.M. The neural substrates of mindfulness: An fMRI investigation. Soc. Neurosci. 2011, 6, 231–242. [Google Scholar] [CrossRef]
- Kabat-Zinn, J. Full Catastrophe Living, Revised Edition: How to Cope With Stress, Pain and Illness Using Mindfulness Meditation; Hachette: Paris, France, 2013. [Google Scholar]
- Gotink, R.A.; Meijboom, R.; Vernooij, M.W.; Smits, M.; Hunink, M.G. 8-Week Mindfulness Based Stress Reduction induces brain changes similar to traditional long-term meditation practice—A systematic review. Brain Cogn. 2016, 108, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Young, K.S.; van der Velden, A.M.; Craske, M.G.; Pallesen, K.J.; Fjorback, L.; Roepstorff, A.; Parsons, C.E. The impact of Mindfulness-Based Interventions on brain activity: A systematic review of functional magnetic resonance imaging studies. Neurosci. Biobehav. Rev. 2017, 84, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, B.K.; Ott, U.; Hempel, H.; Hackl, A.; Wolf, K.; Stark, R.; Vaitl, D. Differential engagement of anterior cingulate and adjacent medial frontal cortex in adept meditators and non-meditators. Neurosci. Lett. 2007, 421, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Tomasino, B.; Fabbro, F. Increases in the right dorsolateral prefrontal cortex and decreases the rostral prefrontal cortex activation after-8 weeks of focused attention based mindfulness meditation. Brain Cogn. 2016, 102, 46–54. [Google Scholar] [CrossRef]
- Zeidan, F.; Emerson, N.M.; Farris, S.R.; Ray, J.N.; Jung, Y.; McHaffie, J.G.; Coghill, R.C. Mindfulness meditation-based pain relief employs different neural mechanisms than placebo and sham mindfulness meditation-induced analgesia. J. Neurosci. 2015, 35, 15307–15325. [Google Scholar] [CrossRef]
- Tang, Y.-Y.; Hölzel, B.K.; Posner, M.I. The neuroscience of mindfulness meditation. Nat. Rev. Neurosci. 2015, 16, 213. [Google Scholar] [CrossRef]
- Kabat-Zinn, J. Wherever You Go, There You Are: Mindfulness Meditation in Everyday Life; Hachette Books: New York, NY, USA, 2009. [Google Scholar]
- Zsadanyi, S.E.; Kurth, F.; Luders, E. The Effects of Mindfulness and Meditation on the Cingulate Cortex in the Healthy Human Brain: A Review. Mindfulness 2021, 12, 2371–2387. [Google Scholar] [CrossRef]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Whitfield-Gabrieli, S.; Moran, J.M.; Nieto-Castañón, A.; Triantafyllou, C.; Saxe, R.; Gabrieli, J.D. Associations and dissociations between default and self-reference networks in the human brain. Neuroimage 2011, 55, 225–232. [Google Scholar] [CrossRef]
- Brewer, J.A.; Worhunsky, P.D.; Gray, J.R.; Tang, Y.-Y.; Weber, J.; Kober, H. Meditation experience is associated with differences in default mode network activity and connectivity. Proc. Natl. Acad. Sci. USA 2011, 108, 20254–20259. [Google Scholar] [CrossRef] [Green Version]
- Gusnard, D.A.; Akbudak, E.; Shulman, G.L.; Raichle, M.E. Medial prefrontal cortex and self-referential mental activity: Relation to a default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 4259–4264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longarzo, M.; D’Olimpio, F.; Chiavazzo, A.; Santangelo, G.; Trojano, L.; Grossi, D. The relationships between interoception and alexithymic trait. The Self-Awareness Questionnaire in healthy subjects. Front. Psychol. 2015, 6, 1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossi, D.; Di Vita, A.; Palermo, L.; Sabatini, U.; Trojano, L.; Guariglia, C. The brain network for self-feeling: A symptom-lesion mapping study. Neuropsychologia 2014, 63, 92–98. [Google Scholar] [CrossRef]
- Garrison, K.A.; Scheinost, D.; Worhunsky, P.D.; Elwafi, H.M.; Thornhill, T.A.; Thompson, E.; Saron, C.; Desbordes, G.; Kober, H.; Hampson, M. Real-Time fMRI links subjective experience with brain activity during focused attention. Neuroimage 2013, 81, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Kirlic, N.; Cohen, Z.P.; Tsuchiyagaito, A.; Misaki, M.; McDermott, T.J.; Aupperle, R.L.; Stewart, J.L.; Singh, M.K.; Paulus, M.P.; Bodurka, J. Self-Regulation of the Posterior Cingulate Cortex with Real-Time fMRI Neurofeedback in Healthy Adolescents. Cogn. Affect. Behav. Neurosci. 2021; in press. [Google Scholar]
- Takesian, A.E.; Hensch, T.K. Balancing plasticity/stability across brain development. Prog. Brain Res. 2013, 207, 3–34. [Google Scholar]
- Fuhrmann, D.; Knoll, L.J.; Blakemore, S.-J. Adolescence as a sensitive period of brain development. Trends Cogn. Sci. 2015, 19, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Tanay, G.; Bernstein, A. State Mindfulness Scale (SMS): Development and initial validation. Psychol. Assess. 2013, 25, 1286. [Google Scholar] [CrossRef] [Green Version]
- Cella, D.; Riley, W.; Stone, A.; Rothrock, N.; Reeve, B.; Yount, S.; Amtmann, D.; Bode, R.; Buysse, D.; Choi, S. The Patient-Reported Outcomes Measurement Information System (PROMIS) developed and tested its first wave of adult self-reported health outcome item banks: 2005–2008. J. Clin. Epidemiol. 2010, 63, 1179–1194. [Google Scholar] [CrossRef] [Green Version]
- Forrest, C.B.; Bevans, K.B.; Filus, A.; Devine, J.; Becker, B.D.; Carle, A.C.; Teneralli, R.E.; Moon, J.; Ravens-Sieberer, U. Assessing children’s eudaimonic well-being: The PROMIS pediatric meaning and purpose item banks. J. Pediatr. Psychol. 2019, 44, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Forrest, C.B.; Ravens-Sieberer, U.; Devine, J.; Becker, B.D.; Teneralli, R.E.; Moon, J.; Carle, A.C.; Tucker, C.A.; Bevans, K.B. Development and evaluation of the PROMIS® pediatric positive affect item bank, child-report and parent-proxy editions. J. Happiness Stud. 2018, 19, 699–718. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, N.R.; Kashikar-Zuck, S.; Mara, C.; Goldschneider, K.R.; Revicki, D.A.; Dampier, C.; Sherry, D.D.; Crosby, L.; Carle, A.; Cook, K.F. Development and validation of the self-reported PROMIS pediatric pain behavior item bank and short form scale. Pain 2017, 158, 1323. [Google Scholar] [CrossRef]
- Amtmann, D.; Cook, K.F.; Jensen, M.P.; Chen, W.-H.; Choi, S.; Revicki, D.; Cella, D.; Rothrock, N.; Keefe, F.; Callahan, L. Development of a PROMIS item bank to measure pain interference. Pain 2010, 150, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.-S.; Stucky, B.D.; Thissen, D.; Varni, J.W.; DeWitt, E.M.; Irwin, D.E.; Yeatts, K.B.; DeWalt, D.A. Development and psychometric properties of the PROMIS® pediatric fatigue item banks. Qual. Life Res. 2013, 22, 2417–2427. [Google Scholar] [CrossRef] [Green Version]
- Rudkin, E.; Medvedev, O.N.; Siegert, R.J. The five-facet mindfulness questionnaire: Why the observing subscale does not predict psychological symptoms. Mindfulness 2018, 9, 230–242. [Google Scholar] [CrossRef]
- Desmond, J.E.; Glover, G.H. Estimating sample size in functional MRI (fMRI) neuroimaging studies: Statistical power analyses. J. Neurosci. Methods 2002, 118, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Kelley, W.M.; Macrae, C.N.; Wyland, C.L.; Caglar, S.; Inati, S.; Heatherton, T.F. Finding the self? An event-related fMRI study. J. Cogn. Neurosci. 2002, 14, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Ros, T.; Enriquez-Geppert, S.; Zotev, V.; Young, K.D.; Wood, G.; Whitfield-Gabrieli, S.; Wan, F.; Vuilleumier, P.; Vialatte, F.; Van De Ville, D. Consensus on the Reporting and Experimental Design of Clinical and Cognitive-Behavioural Neurofeedback Studies (CRED-nf Checklist); Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Pruessmann, K.P.; Weiger, M.; Scheidegger, M.B.; Boesiger, P. SENSE: Sensitivity encoding for fast MRI. Magn. Reson. Med. Off. J. Int. Soc. Magn. Reson. Med. 1999, 42, 952–962. [Google Scholar] [CrossRef]
- Bellgowan, P.S.; Bandettini, P.A.; van Gelderen, P.; Martin, A.; Bodurka, J. Improved BOLD detection in the medial temporal region using parallel imaging and voxel volume reduction. Neuroimage 2006, 29, 1244–1251. [Google Scholar] [CrossRef]
- Bodurka, J.; Ye, F.; Petridou, N.; Murphy, K.; Bandettini, P.A. Mapping the MRI voxel volume in which thermal noise matches physiological noise—Implications for fMRI. Neuroimage 2007, 34, 542–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, R.W. AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput. Biomed. Res. Int. J. 1996, 29, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Glover, G.H.; Li, T.Q.; Ress, D. Image-Based method for retrospective correction of physiological motion effects in fMRI: RETROICOR. Magn. Reson. Med. Off. J. Int. Soc. Magn. Reson. Med. 2000, 44, 162–167. [Google Scholar] [CrossRef]
- Birn, R.M.; Smith, M.A.; Jones, T.B.; Bandettini, P.A. The respiration response function: The temporal dynamics of fMRI signal fluctuations related to changes in respiration. Neuroimage 2008, 40, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Avants, B.B.; Epstein, C.L.; Grossman, M.; Gee, J.C. Symmetric diffeomorphic image registration with cross-correlation: Evaluating automated labeling of elderly and neurodegenerative brain. Med. Image Anal. 2008, 12, 26–41. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.J.; Saad, Z.S.; Simmons, W.K.; Milbury, L.A.; Cox, R.W. Mapping sources of correlation in resting state FMRI, with artifact detection and removal. Neuroimage 2010, 52, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, M.A.; Geuter, S.; Wager, T.D.; Caffo, B.S. Modular preprocessing pipelines can reintroduce artifacts into fMRI data. Hum. Brain Mapp. 2019, 40, 2358–2376. [Google Scholar] [CrossRef] [Green Version]
- Misaki, M.; Bodurka, J. The impact of real-time fMRI denoising on online evaluation of brain activity and functional connectivity. J. Neural Eng. 2021, 18, 046092. [Google Scholar] [CrossRef]
- Fan, L.; Li, H.; Zhuo, J.; Zhang, Y.; Wang, J.; Chen, L.; Yang, Z.; Chu, C.; Xie, S.; Laird, A.R. The human brainnetome atlas: A new brain atlas based on connectional architecture. Cereb. Cortex 2016, 26, 3508–3526. [Google Scholar] [CrossRef]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Revelle, W.R. Psych: Procedures for Personality and Psychological Research; Northwestern University Press: Evanston, IL, USA, 2017. [Google Scholar]
- Bates, D.; Sarkar, D.; Bates, M.D.; Matrix, L. The lme4 Package. R Package Version. 2007, Volume 2, p. 74. Available online: https://cran.r-project.org/web/packages/lme4/lme4.pdf (accessed on 2 March 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. J. Math. Methods Biosci. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Kurth, F.; Zilles, K.; Fox, P.T.; Laird, A.R.; Eickhoff, S.B. A link between the systems: Functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct. Funct. 2010, 214, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Dietz, M.; Blair, K.S.; van Beek, M.; Rees, G.; Vestergaard-Poulsen, P.; Lutz, A.; Roepstorff, A. Cognitive-Affective neural plasticity following active-controlled mindfulness intervention. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 15601–15610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, U.; Gu, X.; Sharp, C.; Hula, A.; Fonagy, P.; Montague, P.R. Mindfulness training increases cooperative decision making in economic exchanges: Evidence from fMRI. NeuroImage 2016, 138, 274–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevinc, G.; Hölzel, B.K.; Hashmi, J.; Greenberg, J.; McCallister, A.; Treadway, M.; Schneider, M.L.; Dusek, J.A.; Carmody, J.; Lazar, S.W. Common and dissociable neural activity after mindfulness-based stress reduction and relaxation response programs. Psychosom. Med. 2018, 80, 439. [Google Scholar] [CrossRef] [Green Version]
- Holzel, B.K.; Hoge, E.A.; Greve, D.N.; Gard, T.; Creswell, J.D.; Brown, K.W.; Barrett, L.F.; Schwartz, C.; Vaitl, D.; Lazar, S.W. Neural mechanisms of symptom improvements in generalized anxiety disorder following mindfulness training. Neuroimage Clin. 2013, 2, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Haase, L.; May, A.C.; Falahpour, M.; Isakovic, S.; Simmons, A.N.; Hickman, S.D.; Liu, T.T.; Paulus, M.P. A pilot study investigating changes in neural processing after mindfulness training in elite athletes. Front. Behav. Neurosci. 2015, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Azzalini, D.; Rebollo, I.; Tallon-Baudry, C. Visceral signals shape brain dynamics and cognition. Trends Cogn. Sci. 2019, 23, 488–509. [Google Scholar] [CrossRef] [Green Version]
- Uddin, L.Q.; Nomi, J.S.; Hébert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and function of the human insula. J. Clin. Neurophysiol. Off. Publ. Am. Electroencephalogr. Soc. 2017, 34, 300. [Google Scholar] [CrossRef]
- Kleckner, I.R.; Zhang, J.; Touroutoglou, A.; Chanes, L.; Xia, C.; Simmons, W.K.; Quigley, K.S.; Dickerson, B.C.; Barrett, L.F. Evidence for a large-scale brain system supporting allostasis and interoception in humans. Nat. Hum. Behav. 2017, 1, 0069. [Google Scholar] [CrossRef] [Green Version]
- Sitaram, R.; Ros, T.; Stoeckel, L.; Haller, S.; Scharnowski, F.; Lewis-Peacock, J.; Weiskopf, N.; Blefari, M.L.; Rana, M.; Oblak, E. Closed-Loop brain training: The science of neurofeedback. Nat. Rev. Neurosci. 2017, 18, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Vassena, E.; Silvetti, M.; Boehler, C.N.; Achten, E.; Fias, W.; Verguts, T. Overlapping neural systems represent cognitive effort and reward anticipation. PLoS ONE 2014, 9, e91008. [Google Scholar] [CrossRef] [PubMed]
- Westbrook, A.; Braver, T.S. Cognitive effort: A neuroeconomic approach. Cogn. Affect. Behav. Neurosci. 2015, 15, 395–415. [Google Scholar] [CrossRef] [PubMed]
- Krause, F.; Kogias, N.; Krentz, M.; Luhrs, M.; Goebel, R.; Hermans, E.J. Self-Regulation of stress-related large-scale brain network balance using real-time fMRI neurofeedback. Neuroimage 2021, 243, 118527. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Macdonald, A.M.; Botvinick, M.; Ross, L.L.; Stenger, V.A.; Noll, D.; Cohen, J.D. Parsing executive processes: Strategic vs. evaluative functions of the anterior cingulate cortex. Proc. Natl. Acad. Sci. USA 2000, 97, 1944–1948. [Google Scholar] [CrossRef] [Green Version]
- Kerns, J.G.; Cohen, J.D.; MacDonald, A.W.; Cho, R.Y.; Stenger, V.A.; Carter, C.S. Anterior cingulate conflict monitoring and adjustments in control. Science 2004, 303, 1023–1026. [Google Scholar] [CrossRef] [Green Version]
- Craig, A.D. How do you feel—Now? The anterior insula and human awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef]
- Ninaus, M.; Kober, S.; Witte, M.; Koschutnig, K.; Stangl, M.; Neuper, C.; Wood, G. Neural substrates of cognitive control under the belief of getting neurofeedback training. Front. Hum. Neurosci. 2013, 7, 914. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, R.B.; Skandali, N.; Dayan, P.; Dolan, R.J. A computational and neural model of momentary subjective well-being. Proc. Natl. Acad. Sci. USA 2014, 111, 12252–12257. [Google Scholar] [CrossRef] [Green Version]
- Nardo, D.; Högberg, G.; Flumeri, F.; Jacobsson, H.; Larsson, S.; Hällström, T.; Pagani, M. Self-Rating scales assessing subjective well-being and distress correlate with rCBF in PTSD-sensitive regions. Psychol. Med. 2011, 41, 2549–2561. [Google Scholar] [CrossRef]
- Cabeen, R.P.; Toga, A.W.; Allman, J.M. Frontoinsular cortical microstructure is linked to life satisfaction in young adulthood. Brain Imaging Behav. 2021, 15, 2775–2789. [Google Scholar] [CrossRef]
- Li, R.; Zhu, X.; Zheng, Z.; Wang, P.; Li, J. Subjective well-being is associated with the functional connectivity network of the dorsal anterior insula. Neuropsychologia 2020, 141, 107393. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.G.; Rapport, L.J.; Hanks, R.A.; Lichtenberg, P.A.; Telmet, K. Cognitive and psychosocial predictors of subjective well-being in urban older adults. Clin. Neuropsychol. 2003, 17, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.J.; Kanai, R.; Rees, G.; Bates, T.C. Neural correlates of the ‘good life’: Eudaimonic well-being is associated with insular cortex volume. Soc. Cogn. Affect. Neurosci. 2014, 9, 615–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cash, M.; Whittingham, K. What facets of mindfulness contribute to psychological well-being and depressive, anxious, and stress-related symptomatology? Mindfulness 2010, 1, 177–182. [Google Scholar] [CrossRef]
- Farb, N.; Segal, Z.; Anderson, A. Mindfulness meditation training alters cortical representations of interoceptive attention. Soc. Cogn. Affect. Neurosci. 2013, 8, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Segerdahl, A.R.; Mezue, M.; Okell, T.W.; Farrar, J.T.; Tracey, I. The dorsal posterior insula is not an island in pain but subserves a fundamental role-Response to:“Evidence against pain specificity in the dorsal posterior insula” by Davis et al. F1000Research 2015, 4, 1207. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, D.; Obaid, S.; Fournier-Gosselin, M.P.; Bouthillier, A.; Nguyen, D.K. Deep Brain Stimulation of the Posterior Insula in Chronic Pain: A Theoretical Framework. Brain Sci. 2021, 11, 639. [Google Scholar] [CrossRef]
- Farb, N.; Segal, Z.V.; Mayberg, H.; Bean, J.; McKeon, D.; Fatima, Z.; Anderson, A.K. Attending to the present: Mindfulness meditation reveals distinct neural modes of self-reference. Soc. Cogn. Affect. Neurosci. 2007, 2, 313–322. [Google Scholar] [CrossRef]
- Grant, J.A.; Courtemanche, J.; Rainville, P. A non-elaborative mental stance and decoupling of executive and pain-related cortices predicts low pain sensitivity in Zen meditators. Pain 2011, 152, 150–156. [Google Scholar] [CrossRef]
- Gard, T.; Hölzel, B.K.; Sack, A.T.; Hempel, H.; Lazar, S.W.; Vaitl, D.; Ott, U. Pain attenuation through mindfulness is associated with decreased cognitive control and increased sensory processing in the brain. Cereb. Cortex 2012, 22, 2692–2702. [Google Scholar] [CrossRef] [Green Version]
- Kirlic, N.; Aupperle, R.L.; Rhudy, J.L.; Misaki, M.; Kuplicki, R.; Sutton, A.; Alvarez, R.P. Latent variable analysis of negative affect and its contributions to neural responses during shock anticipation. Neuropsychopharmacology 2019, 44, 695–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulus, M.P.; Khalsa, S. When you don’t feel right inside: Homeostatic dysregulation and the mid-insular cortex in psychiatric disorders. Am. Psychiatric. Assoc. 2021, 178, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Jakab, A.; Molnár, P.P.; Bogner, P.; Béres, M.; Berényi, E.L. Connectivity-Based parcellation reveals interhemispheric differences in the insula. Brain Topogr. 2012, 25, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Ghaziri, J.; Tucholka, A.; Girard, G.; Houde, J.-C.; Boucher, O.; Gilbert, G.; Descoteaux, M.; Lippé, S.; Rainville, P.; Nguyen, D.K. The corticocortical structural connectivity of the human insula. Cereb. Cortex 2017, 27, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Cerliani, L.; Thomas, R.M.; Jbabdi, S.; Siero, J.C.; Nanetti, L.; Crippa, A.; Gazzola, V.; D’Arceuil, H.; Keysers, C. Probabilistic tractography recovers a rostrocaudal trajectory of connectivity variability in the human insular cortex. Hum. Brain Mapp. 2012, 33, 2005–2034. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Supekar, K.S.; Ryali, S.; Menon, V. Dynamic reconfiguration of structural and functional connectivity across core neurocognitive brain networks with development. J. Neurosci. 2011, 31, 18578–18589. [Google Scholar] [CrossRef]
- Apkarian, A.V.; Bushnell, M.C.; Treede, R.-D.; Zubieta, J.-K. Human brain mechanisms of pain perception and regulation in health and disease. Eur. J. Pain 2005, 9, 463–484. [Google Scholar] [CrossRef]
- Taylor, K.S.; Seminowicz, D.A.; Davis, K.D. Two systems of resting state connectivity between the insula and cingulate cortex. Hum. Brain Mapp. 2009, 30, 2731–2745. [Google Scholar] [CrossRef]
- Vogt, B.A. Cingulate cortex in the three limbic subsystems. Handb. Clin. Neurol. 2019, 166, 39–51. [Google Scholar]
- Vogt, B.A.; Vogt, L.; Farber, N.B.; Bush, G. Architecture and neurocytology of monkey cingulate gyrus. J. Comp. Neurol. 2005, 485, 218–239. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, I.L.; de Matos, V.P.; Cuesta, T.A.C.; Herbet, G.; Destrieux, C. The human cingulum: From the limbic tract to the connectionist paradigm. Neuropsychologia 2020, 144, 107487. [Google Scholar] [CrossRef] [PubMed]
- Gehrlach, D.A.; Dolensek, N.; Klein, A.S.; Chowdhury, R.R.; Matthys, A.; Junghänel, M.; Gaitanos, T.N.; Podgornik, A.; Black, T.D.; Vaka, N.R. Aversive state processing in the posterior insular cortex. Nat. Neurosci. 2019, 22, 1424–1437. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.R.; Lau, M.; Shapiro, S.; Carlson, L.; Anderson, N.D.; Carmody, J.; Segal, Z.V.; Abbey, S.; Speca, M.; Velting, D.; et al. Mindfulness: A Proposed Operational Definition. Clin. Psychol. Sci. Pract. 2004, 11, 230–241. [Google Scholar] [CrossRef]
- Kabat-Zinn, J. An outpatient program in behavioral medicine for chronic pain patients based on the practice of mindfulness meditation: Theoretical considerations and preliminary results. Gen. Hosp. Psychiatry 1982, 4, 33–47. [Google Scholar] [CrossRef]
- Lindsay, E.K.; Creswell, J.D. Mechanisms of mindfulness training: Monitor and Acceptance Theory (MAT). Clin. Psychol. Rev. 2017, 51, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Landtblom, A.-M.; Lindehammar, H.; Karlsson, H. Insular cortex activation in a patient with “sensed presence”/ecstatic seizures. Epilepsy Behav. 2011, 20, 714–718. [Google Scholar] [CrossRef]
- Chambers, R.; Gullone, E.; Allen, N.B. Mindful emotion regulation: An integrative review. Clin. Psychol. Rev. 2009, 29, 560–572. [Google Scholar] [CrossRef]
- Chen, W.G.; Schloesser, D.; Arensdorf, A.M.; Simmons, J.M.; Cui, C.; Valentino, R.; Gnadt, J.W.; Nielsen, L.; Hillaire-Clarke, C.S.; Spruance, V. The emerging science of interoception: Sensing, integrating, interpreting, and regulating signals within the self. Trends Neurosci. 2021, 44, 3–16. [Google Scholar] [CrossRef]
- Li, D.; Zucker, N.L.; Kragel, P.A.; Covington, V.E.; LaBar, K.S. Adolescent development of insula-dependent interoceptive regulation. Dev. Sci. 2017, 20, e12438. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.; Brewer, R.; Catmur, C.; Bird, G. Interoception and psychopathology: A developmental neuroscience perspective. Dev. Cogn. Neurosci. 2017, 23, 45–56. [Google Scholar] [CrossRef]
- May, A.C.; Stewart, J.L.; Paulus, M.P.; Tapert, S.F. The effect of age on neural processing of pleasant soft touch stimuli. Front. Behav. Neurosci. 2014, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Van Hoorn, J.; Crone, E.A.; Van Leijenhorst, L. Hanging out with the right crowd: Peer influence on risk-taking behavior in adolescence. J. Res. Adolesc. 2017, 27, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.G.; Xiao, L.; Bechara, A. Decision making in children and adolescents: Impaired Iowa Gambling Task performance in early adolescence. Dev. Psychol. 2012, 48, 1180. [Google Scholar] [CrossRef]
- Northoff, G.; Heinzel, A.; De Greck, M.; Bermpohl, F.; Dobrowolny, H.; Panksepp, J. Self-Referential processing in our brain—A meta-analysis of imaging studies on the self. Neuroimage 2006, 31, 440–457. [Google Scholar] [CrossRef]
- Misaki, M.; Barzigar, N.; Zotev, V.; Phillips, R.; Cheng, S.; Bodurka, J. Real-Time fMRI processing with physiological noise correction—Comparison with off-line analysis. J. Neurosci. Methods 2015, 256, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Taren, A.A.; Smith, D.V. Functional Parcellation of the Default Mode Network: A Large-Scale Meta-Analysis. Sci. Rep. 2020, 10, 16096. [Google Scholar] [CrossRef]
- Zotev, V.; Phillips, R.; Yuan, H.; Misaki, M.; Bodurka, J. Self-Regulation of human brain activity using simultaneous real-time fMRI and EEG neurofeedback. NeuroImage 2014, 85, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-K.; Zotev, V.; Misaki, M.; Phillips, R.; Luo, Q.; Bodurka, J. Automatic EEG-assisted retrospective motion correction for fMRI (aE-REMCOR). Neuroimage 2016, 129, 133. [Google Scholar] [CrossRef] [Green Version]
- Birn, R.M.; Diamond, J.B.; Smith, M.A.; Bandettini, P.A. Separating respiratory-variation-related fluctuations from neuronal-activity-related fluctuations in fMRI. Neuroimage 2006, 31, 1536–1548. [Google Scholar] [CrossRef]
- Chang, C.; Cunningham, J.P.; Glover, G.H. Influence of heart rate on the BOLD signal: The cardiac response function. Neuroimage 2009, 44, 857–869. [Google Scholar] [CrossRef] [Green Version]
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in behavior made easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [PubMed] [Green Version]
- Zotev, V.; Krueger, F.; Phillips, R.; Alvarez, R.P.; Simmons, W.K.; Bellgowan, P.; Drevets, W.C.; Bodurka, J. Self-Regulation of amygdala activation using real-time fMRI neurofeedback. PLoS ONE 2011, 6, e24522. [Google Scholar] [CrossRef] [PubMed]


| Demographic | % |
|---|---|
| Race | |
| White | 71 |
| Black | 2 |
| Asian | 5 |
| American Indian/Alaska Native | 11 |
| Biracial/Multiracial | 11 |
| Education | |
| 7th grade | 18 |
| 8th grade | 38 |
| 9th grade | 18 |
| 10th grade | 16 |
| 11th grade | 7 |
| 13th grade | 3 |
| Family Income | |
| $0–$49,999 | 7 |
| $50,000–$99,999 | 38 |
| $100,000–$149,999 | 21 |
| $150,000–$199,999 | 17 |
| >$200,000 | 17 |
| Task Ratings | Mean | SD | Estimate | SE | t | p | Cohen’s d |
|---|---|---|---|---|---|---|---|
| Focus-on-Breath | |||||||
| OBS | 7.09 | 1.54 | |||||
| NF-1 | 6.56 | 1.67 | −0.51 | 0.36 | −1.42 | 0.16 | −0.25 |
| NF-2 | 6.45 | 1.70 | −0.63 | 0.36 | −1.72 | 0.09 | −0.30 |
| NF-3 | 6.70 | 2.04 | −0.37 | 0.35 | −1.05 | 0.30 | −0.18 |
| TRS | 6.53 | 1.61 | −0.50 | 0.35 | −1.42 | 0.16 | −0.25 |
| Mind Wander | |||||||
| OBS | 5.23 | 2.12 | |||||
| NF-1 | 4.85 | 1.86 | −0.47 | 0.36 | −1.31 | 0.19 | −0.23 |
| NF-2 | 5.21 | 2.09 | −0.11 | 0.36 | −0.31 | 0.76 | −0.05 |
| NF-3 | 5.83 | 1.83 | 0.10 | 0.35 | 0.29 | 0.77 | 0.05 |
| TRS | 5.75 | 1.84 | 0.45 | 0.36 | 1.26 | 0.21 | 0.22 |
| Current Feeling | |||||||
| OBS | 2.89 | 1.53 | |||||
| NF-1 | 3.41 | 1.76 | 0.38 | 0.29 | 1.30 | 0.20 | 0.22 |
| NF-2 | 3.06 | 1.69 | 0.15 | 0.29 | 0.51 | 0.61 | 0.09 |
| NF-3 | 3.03 | 1.78 | 0.05 | 0.29 | 0.16 | 0.87 | 0.03 |
| TRS | 3.06 | 2.04 | 0.10 | 0.29 | 0.34 | 0.74 | 0.06 |
| Measure | Mean | SD | Estimate | SE | t | p | Cohen’s d |
| State Mindfulness Scale (SMS) | |||||||
| T1 | 71.22 | 14.09 | |||||
| T2 | 74.68 | 12.69 | 3.46 | 1.43 | 2.41 | <0.05 | 0.53 |
| INS Subregions | Run | PCC_OBS | PCC_NF-1 | PCC_NF-2 | PCC_NF-3 | PCC_TRS |
|---|---|---|---|---|---|---|
| aINS | OBS | 0.65 *** | - | - | - | - |
| NF-1 | - | 0.12 | - | - | - | |
| NF-2 | - | - | 0.35 * | - | - | |
| NF-3 | - | - | - | 0.37 * | - | |
| TRS | - | - | - | - | 0.34 * | |
| mINS | OBS | 0.69 *** | - | - | - | - |
| NF-1 | - | 0.09 | - | - | - | |
| NF-2 | - | - | 0.36 * | - | - | |
| NF-3 | - | - | - | 0.42 ** | - | |
| TRS | - | - | - | - | 0.35 * | |
| pINS | OBS | 0.71 *** | - | - | - | - |
| NF-1 | - | 0.45 ** | - | - | - | |
| NF-2 | - | - | 0.56 *** | - | - | |
| NF-3 | - | - | - | 0.53 *** | - | |
| TRS | - | - | - | - | 0.42 ** |
| Run | Mean | SD | Estimate | SE | t | p | Cohen’s d |
|---|---|---|---|---|---|---|---|
| Anterior insular cortex (aINS) | |||||||
| OBS | −0.16 | 0.19 | |||||
| NF-1 | −0.01 | 0.18 | 0.16 | 0.04 | 3.81 | <0.001 | 0.63 |
| NF-2 | −0.02 | 0.21 | 0.14 | 0.04 | 3.26 | <0.01 | 0.54 |
| NF-3 | −0.05 | 0.28 | 0.11 | 0.04 | 2.75 | <0.01 | 0.46 |
| TRS | −0.12 | 0.24 | 0.05 | 0.04 | 1.16 | <0.001 | 0.19 |
| Mid-insular cortex (mINS) | |||||||
| OBS | 0 | 0.18 | |||||
| NF-1 | −0.06 | 0.17 | −0.06 | 0.03 | −1.70 | 0.09 | −0.28 |
| NF-2 | −0.05 | 0.15 | −0.05 | 0.03 | −1.47 | 0.15 | −0.24 |
| NF-3 | −0.08 | 0.2 | −0.08 | 0.03 | −2.42 | 0.05 | −0.40 |
| TRS | −0.06 | 0.19 | −0.06 | 0.03 | −1.63 | 0.11 | −0.27 |
| Posterior insular cortex (pINS) | |||||||
| OBS | 0.01 | 0.17 | |||||
| NF-1 | −0.17 | 0.14 | −0.18 | 0.03 | −5.25 | <0.001 | −0.88 |
| NF-2 | −0.16 | 0.15 | −0.17 | 0.03 | −4.91 | <0.001 | −0.82 |
| NF-3 | −0.17 | 0.18 | −0.18 | 0.03 | −5.27 | <0.001 | −0.88 |
| TRS | −0.05 | 0.17 | −0.06 | 0.03 | −1.72 | 0.09 | −0.29 |
| Run | Estimate | Std. Error | z Statistic | p Value |
|---|---|---|---|---|
| Anterior insular cortex (aINS) | ||||
| NF-1:OBS | 0.16 | 0.04 | 3.81 | <0.01 |
| NF-2:OBS | 0.14 | 0.04 | 3.26 | <0.05 |
| NF-3:OBS | 0.11 | 0.04 | 2.75 | <0.05 |
| TR:OBS | 0.05 | 0.04 | 1.16 | 0.78 |
| NF-2:NF-1 | −0.02 | 0.04 | −0.52 | 0.98 |
| NF-3:NF-1 | −0.04 | 0.04 | −1.06 | 0.83 |
| TR:NF-1 | −0.11 | 0.04 | −2.66 | 0.06 |
| NF-3:NF-2 | −0.02 | 0.04 | −0.53 | 0.98 |
| TR:NF-2 | −0.09 | 0.04 | −2.11 | 0.22 |
| TR:NF-3 | −0.07 | 0.04 | −1.60 | 0.50 |
| Mid-insular cortex (mINS) | ||||
| NF-1:OBS | −0.06 | 0.03 | −1.70 | 0.43 |
| NF-2:OBS | −0.05 | 0.03 | −1.47 | 0.59 |
| NF-3:OBS | −0.08 | 0.03 | −2.42 | 0.11 |
| TR:OBS | −0.06 | 0.03 | −1.63 | 0.48 |
| NF-2:NF-1 | 0.01 | 0.03 | 0.22 | 1.00 |
| NF-3:NF-1 | −0.02 | 0.03 | −0.72 | 0.95 |
| TR:NF-1 | 0.00 | 0.03 | 0.07 | 1.00 |
| NF-3:NF-2 | −0.03 | 0.03 | −0.93 | 0.89 |
| TR:NF-2 | −0.01 | 0.03 | −0.15 | 1.00 |
| TR:NF-3 | 0.03 | 0.03 | 0.79 | 0.93 |
| Posterior insular cortex (pINS) | ||||
| NF-1:OBS | −0.18 | 0.03 | −5.25 | <0.001 |
| NF-2:OBS | −0.17 | 0.03 | −4.91 | <0.001 |
| NF-3:OBS | −0.18 | 0.03 | −5.27 | <0.001 |
| TR:OBS | −0.06 | 0.03 | −1.72 | 0.42 |
| NF-2:NF-1 | 0.01 | 0.03 | 0.30 | 1.00 |
| NF-3:NF-1 | 0.00 | 0.03 | −0.02 | 1.00 |
| TR:NF-1 | 0.12 | 0.03 | 3.52 | <0.01 |
| NF-3:NF-2 | −0.01 | 0.03 | −0.32 | 1.00 |
| TR:NF-2 | 0.11 | 0.03 | 3.20 | <0.05 |
| TR:NF-3 | 0.12 | 0.03 | 3.55 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Cohen, Z.P.; Tsuchiyagaito, A.; Cochran, G.; Aupperle, R.L.; Stewart, J.L.; Singh, M.K.; Misaki, M.; Bodurka, J.; Paulus, M.P.; et al. Neurofeedback-Augmented Mindfulness Training Elicits Distinct Responses in the Subregions of the Insular Cortex in Healthy Adolescents. Brain Sci. 2022, 12, 363. https://doi.org/10.3390/brainsci12030363
Yu X, Cohen ZP, Tsuchiyagaito A, Cochran G, Aupperle RL, Stewart JL, Singh MK, Misaki M, Bodurka J, Paulus MP, et al. Neurofeedback-Augmented Mindfulness Training Elicits Distinct Responses in the Subregions of the Insular Cortex in Healthy Adolescents. Brain Sciences. 2022; 12(3):363. https://doi.org/10.3390/brainsci12030363
Chicago/Turabian StyleYu, Xiaoqian, Zsofia P. Cohen, Aki Tsuchiyagaito, Gabriella Cochran, Robin L. Aupperle, Jennifer L. Stewart, Manpreet K. Singh, Masaya Misaki, Jerzy Bodurka, Martin P. Paulus, and et al. 2022. "Neurofeedback-Augmented Mindfulness Training Elicits Distinct Responses in the Subregions of the Insular Cortex in Healthy Adolescents" Brain Sciences 12, no. 3: 363. https://doi.org/10.3390/brainsci12030363