1. Introduction
The glycolysis product L-lactate (lactate) in the brain has been receiving increasing attention as a growing number of studies suggest its roles in the modulation of neuronal circuit function as diverse as memory, decision making, response to environmental novelty, depression, central arousal, or autonomic regulation [
1,
2,
3,
4,
5,
6,
7]. Likewise, where the mechanisms of lactate actions have been investigated, these, too, are diverse and are discussed in the context of meeting metabolic demands by intercellular energy substrate shuttling, or receptor-mediated signalling, or modulation of gene expression [
4,
8,
9,
10,
11].
Astrocytes can be considered the main source of brain extracellular lactate since they are endowed with the machinery to import glucose from the periphery, store energy in the form of glycogen, and dynamically shuttle energy substrates throughout the astrocytic network and to the extracellular space [
12]. Various studies have suggested that astrocytes favour the glycolytic pathway over mitochondrial oxidative phosphorylation, resulting in excess production and release of lactate [
13]. The concentration gradient of lactate, as demonstrated for the cortex, is directed from the astrocytic towards the neuronal compartment [
7,
14].
In spite of considerable progress and interest in the subject, many questions remain open. As an intermediate product in equilibrium-based enzymatic processes that exits its cells of origin in a gradient-driven but also partially regulated manner, the flux of lactate through the cellular brain compartments in vivo has proven difficult to match up quantitively with its specific actions on cellular targets and brain circuits. We, therefore, aimed to develop a set of molecular tools to support studies into the roles of astrocyte-derived lactate in the brain. The rationale was to harness the activity of bacterial enzymes that metabolise lactate, and to express them in astrocytes. Our aim was to decrease intra-astrocytic lactate pools that can be released to the extracellular space, constitutively or in response to local signals, and consequently to reduce lactate action. We recently reported that viral vector-mediated expression of bacterial lactate oxidase (LOx) in hippocampal astrocytes modulates behaviour in the context of environmental novelty in mice [
5]. Here we expand the novel toolset by two further constructs, lactate 2-monooxygenase (LMO) and D-lactate dehydrogenase (DLDH) and compare the properties between these and LOx when expressed in rodent astrocytes.
LMO, like LOx, irreversibly oxidises lactate into pyruvate at the expense of flavin cofactor reduction, which in turn is re-oxidised by molecular oxygen in a 2-step reaction [
15]. In LOx, pyruvate is released very rapidly from the enzymatic complex, a feature that accounts for the fast reaction kinetics of this enzyme [
15]. As a result, molecular oxygen, which functions as an electron acceptor, forms hydrogen peroxide. LMO activity, on the other hand, exhibits higher kinetic stability, which makes pyruvate dissociation from the enzymatic complex a significantly slower process [
15]. Thus, the reaction of the pyruvate-enzymatic complex with molecular oxygen culminates in pyruvate decarboxylation, yielding acetate and water (
Figure 1). We investigated potential differences in the time course of action and reactive oxygen species-related effects as these criteria could favour the use of one tool over the other for specific applications.
DLDH catalyses the reversible stereospecific NAD-dependent conversion of pyruvate into D-lactate (DL;
Figure 1). This was predicted to raise the DL/lactate ratio in astrocytes and, subsequently, in the extracellular space. For many lactate-dependent processes, DL is a weak substrate and may therefore indirectly impact on lactate-dependent processes but, in certain scenarios, e.g., lactate action in the
locus coeruleus, DL can act as lactate antagonist [
4].
Here we demonstrate the enzymatic activity of viral vector-based expression of LMO and DLDH in rat astrocytes in vitro and suggest that the use of these constructs may facilitate future studies into the functional roles of lactate in the CNS.
2. Materials and Methods
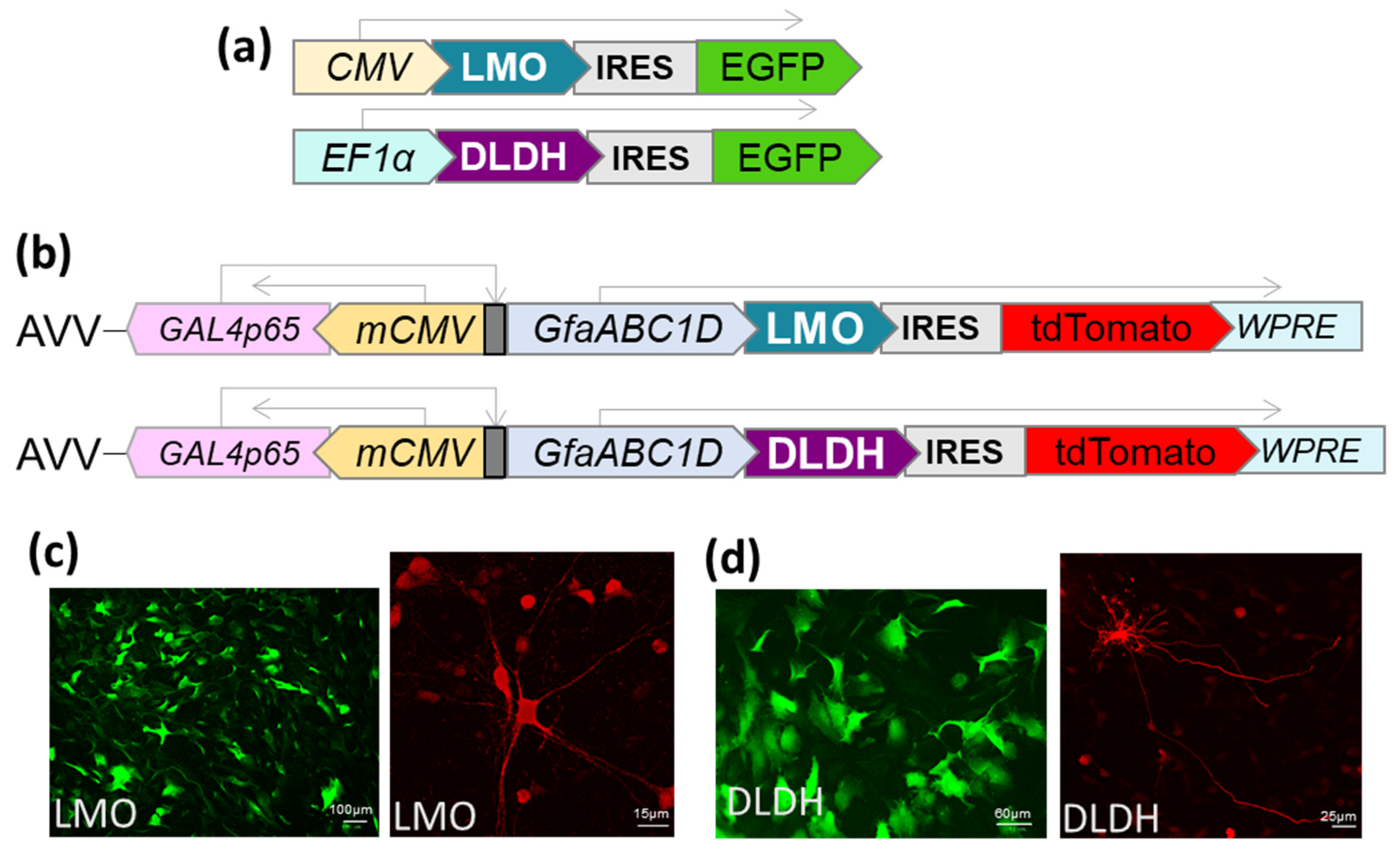
2.1. Vector Construction
The LMO (E.C. 1.13.12.4) coding sequence from
Mycobacterium smegmatis (GenBank: AIU22323.1) and the DLDH (E.C. 1.1.1.28) sequence from
Lactobacillus delbrueckii subsp.
bulgaricus (GenBank: WP013438907) were optimised for mammalian codon usage and synthesised by Invitrogen (Life Technologies, Paisley, UK). Expression cassettes were cloned to be driven by either CMV, EF1-α or the astrocyte-selective transcriptionally-enhanced sGFAP promoter [
16]. In addition, an internal ribosomal entry site (IRES), followed by a fluorescent marker (EGFP or tdTomato), was included downstream of the LMO or DLDH sequence (
Figure 2). The cassettes were transferred to an adenoviral vector (AVV) shuttle plasmid (pXCX) and vectors derived from type 5 adenovirus were produced as described previously [
17]. For experiments in HEK293, expression plasmids were transfected using TransIT-293 (Mirus, MIR 2700).
To enable assessment of changes in intracellular lactate or pyruvate, AVVs for CMV-driven expression of the FRET-based reporters for lactate and pyruvate, Laconic and Pyronic, respectively, were used [
5,
18,
19].
2.2. Astrocyte Cultures
Wistar rat pups were sacrificed in accordance with Schedule 1 of the UK Home Office (Scientific Procedures) Act (1986) and as approved by the University of Bristol ethics committees. Primary dissociated astrocytes and organotypic slice cultures were prepared as previously described [
5,
20], and AVVs were added to the culture media. The multiplicity of infection (MOI) was determined from the AVV titer (transforming units/ml) as a ratio between transforming units added and cell count per culture well. Organotypic cultures were transduced with AVV at the time of plating and used 8–12 days later; dissociated cultures were transduced at least 2 days before experimentation.
2.3. Cell Viability Assays
For estimation of cell viability with the Trypan Blue exclusion assay, astrocytes were trypsinised, harvested, and stained with trypan blue 0.4% (Sigma-Aldrich, T8154, Dorset, UK). The percentage of blue non-viable cells in the suspension was determined in a hemocytometer. For the XTT cell viability assay (Cell Signaling Technology, 9095, Beverly, MA, USA), the formazan formation reaction was performed for 2 h. The relative metabolic viability of the cells was derived from the sample’s absorbance at 450 nm using a microplate reader (Tecan Infinite M200 PRO, Labtech, Heathfield, UK).
2.4. Immunohistochemistry
Primary cultured astrocytes seeded on glass coverslips or brainstem organotypic slices excised from the culture membrane were fixed for 15 min in ice-cold 4% paraformaldehyde (PFA) in phosphate buffer (PBS; pH 7.4). After rinsing in PBS, the samples were incubated for 1 h at room temperature in a solution of 0.3% Triton X-100 (Sigma-Aldrich, T8787) and 10% serum. Samples were incubated overnight at 4 °C with anti-RFP antibody (Rockland Immunochemicals, 600-401-379S; 1:200, Limerick, PA, USA), rinsed in PBS and incubated for 1 h at room temperature with Alexa Fluor 594 antibody (Thermo Fisher Scientific, R37117, Waltham, MA, USA).
2.5. Western Blot
After 72 h of AVV transduction or 3 h of staurosporine (5 µM) treatment, respectively, astrocytes were washed twice with cold PBS, treated with RIPA buffer containing protease and phosphatase inhibitors (Sigma-Aldrich) and harvested. After lysis, cells debris was centrifuged at 12,000× g and frozen at −80 °C until use. Protein extracts were prepared for SDS-PAGE with sample buffer (NuPAGE™ LDS Sample Buffer 4×, Invitrogen) and reducing agent (NuPAGE™ Sample Reducing Agent 10×, Invitrogen), then heated for 5 min to 95 °C. Following protein quantification using Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific), equal amounts of protein were run on 10% acrylamide gel (NuPAGE™ Bis-Tris, Invitrogen), then transferred to a nitrocellulose membrane (Amersham™ Hybond™, Thermo Fisher Scientific) using Trans-Blot Cell (Bio-Rad, Watford, UK) and visualised with Ponceau staining (Sigma-Aldrich). Membranes were blocked with 2% of ECL blocking reagent in PBS-0.1% Tween 20 (PBS-T) for 1 h at room temperature, before being incubated overnight at 4 °C with the primary antibodies: LDHA (1:2000, Cell Signaling), LDHB (1:1000, Abcam, Cambridge, UK), or cleaved caspase 3 antibody (1:1000, Cell Signaling). The same membranes were later incubated with Actin antibody (1:20,000, Sigma). Following three 5 min washing steps in PBS-T, membranes were incubated for 1 h at room temperature with secondary HRP-linked antibodies: Swine anti-rabbit IgG HRP (1:10,000, Agilent, Stockport, UK) or goat anti-mouse IgG HRP (1:10,000, Dako). Enhanced Chemiluminescence EZ ECL kit (Geneflow Limited, Lichfield, UK) was used for immunodetection. Images were taken with G:BOX (Syngene, Cambridge, UK) operated with GeneSys 1.6.9 software. The quantification of immunoblots was performed with Image-J software. Total levels of protein were calculated as the ratio of the LDHA or LDHB with actin from the same membrane.
2.6. Imaging of Intracellular Lactate and Pyruvate
Primary cultured astrocytes on glass coverslips expressing Laconic or Pyronic and either LMO or DLDH with tdTomato were transferred to a recording chamber on an upright confocal microscope (Leica SP1 or SP5) and superfused with HEPES-buffered solution (HBS; in mM: NaCl 137, KCl 5.4 or 3, Na2HPO4 0.34, KH2PO4 0.44, CaCl2 1.6, MgSO4 0.8, NaHCO3 4.2, HEPES 10, Glucose 5.5 or 2; pH 7.4; 32.5 ± 1 °C). Laconic or Pyronic were excited with the 458 nm line of an Argon ion laser and the emission of mTFP and Venus was detected at wavelength 465–500 nm and 515–595 nm, respectively. Images were taken every 3 s. Regions of interest were selected and the FRET ratio (mTFP/Venus) was calculated and normalized to the baseline.
2.7. Determination of Lactate and DL Release from Dissociated Cultures
Lactate or DL levels in conditioned media from dissociated cells expressing LMO or DLDH were determined with fluorometric assays (BioAssay Systems, EFLLC-100 and EFDLC-100, Hayward, CA, USA). HEK293 cells or astrocytes were exposed to serum-free culture media for 2 or 6 h, respectively. Samples used for quantification of DL underwent deproteination using a 10 kDa spin filter to deplete any potential activity of endogenous LDH enzyme. Samples were then processed according to the manufacturer’s instructions and fluorescence intensity was measured in a microplate reader (λexcitation: 530 nm, λemission: 585 nm). Lactate and DL concentrations were calibrated to a standard curve.
2.8. Determination of Extracellular Lactate in Organotypic Brainstem Slices
Amperometric lactate biosensors (Sarissa Biomedical, SBS-LAC-05-50, SBI-NUL-05-50, Coventry, UK) were used for real-time recording of extracellular lactate dynamics in organotypic brainstem slices. Slices were transferred to a recording chamber continuously superfused with HBS (see above). The lactate electrode was placed in contact with the surface of the slice and allowed to stabilise for at least 30 min. The null electrode was positioned on the contralateral side of the slice and its readout was subtracted from the current generated by the lactate sensor.
The potential at the electrode surface was controlled by a potentiostat (Duo-Stat ME200+) and the signal was acquired and analysed using a 1401 interface and Spike 2 software (Cambridge Electronic Design, Cambridge, UK). Known amounts of lactate were applied to the chamber at the beginning and at the end of each recording to calibrate the electrodes.
2.9. Statistical Analysis
Comparison between two experimental groups was made using two-tailed t-tests, paired or unpaired, as indicated. Data from three or more groups were compared using ANOVA followed by post hoc Tukey’s test or non-parametric Wilcoxon Mann–Whitney U test, as indicated. The bars in the scatter plot graphs represent the mean and the error bars the standard deviation. Statistical tests were performed using GraphPad Prism software. Differences with p < 0.05 between the experimental groups were considered significant.
4. Discussion
Studies of lactate-mediated metabolic and signalling interactions between astrocytes and neurons have lately been held back by the lack of experimental tools to chronically and selectively manipulate the lactate release from astrocytes. To address this issue, we developed viral vector-based molecular tools that take advantage of the superior capacity of bacteria to metabolise lactate.
Both LMO and DLDH are active in many microorganisms where they are involved in glycolysis and fermentation. We selected the LMO sequence from
Mycobacterium smegmatis and the DLDH from
Lactobacillus delbrueckii subsp.
bulgaricus based on their better documented enzymatic activities [
15,
22]. When adapted to the eukaryotic expression machinery and expressed in mammalian cells, the activity of the foreign enzymes was confirmed.
A range of factors, such as non-specific side effects of AVV transduction, potential trafficking problems associated with the expression of foreign gene products, interference with glycolytic pathways, as well as by-products of additional enzymatic processes, can all potentially lead to harmful consequences for cells. However, we found that within a normal in vitro working range of AVV MOIs, the expression of LMO and DLDH led to functional enzymatic activity without causing either measurable detriment or signs of apoptotic processes in astrocytes. This was consistent with the expression of the bacterial enzyme LOx which we reported earlier, even though LOx enzymatic activity results in hydrogen peroxide as a by-product, whereas LMO produces acetate, which is expected to be efficiently metabolised further by astrocytes (
Figure 1; [
5]). In fact, since the data shown here and in [
5] were obtained in parallel and are based on the same control experiments, results from the expression of LMO, DLDH, and LOx are directly comparable. Higher levels of LMO, but not DLDH, expression suggested deleterious effects, which may highlight the importance of maintaining basal lactate or NADH levels for astrocytes. The impact of altered astrocytic lactate metabolism on surrounding brain cells, including neurons, should be assessed in future studies.
4.1. Lactate Breakdown by LMO
Many studies have pointed out the importance of lactate for CNS metabolism and also, more recently, for brain information processing. Even if it were possible, it would be deleterious for cell and tissue function to eliminate lactate pools too drastically. What our intervention aimed for and achieved was a modulatory limitation that would decrease the amplitude of lactate spikes in response to triggers of astrocytic lactate release [
7,
14]. These are more likely to play relevant roles in activity-dependent lactate signalling.
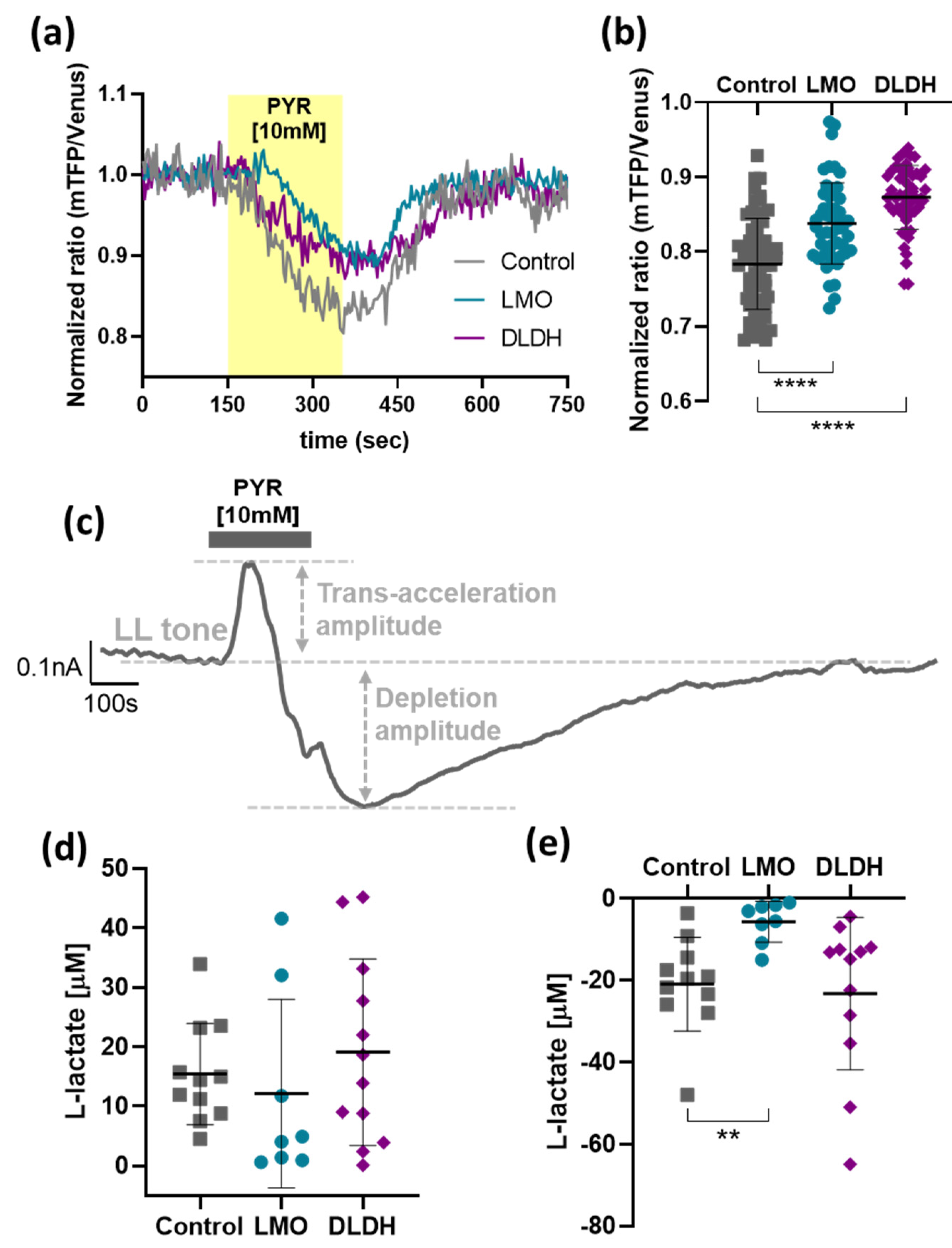
As predicted, LMO expression decreased intracellular lactate levels, and this was reflected by a reduction of extracellular lactate concentrations in unstimulated conditions, as well as reduced pool sizes detected with forced pyruvate-induced extrusion. Therefore, in organotypic brainstem slices, we would have expected a reduction in peak lactate release upon trans-acceleration but were not able to detect a significant change. It is, however, important to keep in mind that under these conditions, lactate release was forced from all MCT-expressing cells in the slice culture - including neurons - and LMO was only expressed selectively in astrocytes. Like LMO, LOx also significantly decreased the lactate tone in organotypic slices [
5]. In contrast to LMO, however, we observed also a decrease in lactate release upon pyruvate trans-acceleration [
5]. Since LMO and LOx exhibit different kinetic properties, with LOx breaking down lactate at approximately a 7000-fold faster rate than LMO, this observation may reflect the faster kinetics of LOx [
15]. The astrocyte-selective LMO expression in organotypic slices still significantly affected the maximal drop in lactate release after forced extrusion, and this was the case for LOx expression, as well [
5].
We further examined how LMO expression affected the balance of pyruvate and lactate in astrocytes. As expected, when one of the main routes of constitutive lactate export, MCT1, was blocked, lactate accumulated in the cells before pyruvate levels started rising (not shown). This would apply back-pressure on the LDH reaction until an equilibrium between pyruvate and lactate was re-established, consistent with the plateau in the recorded traces. Since the increased lactate levels under these conditions were difficult to quantify accurately, we evaluated the rate of pyruvate accumulation as an indirect measure of lactate build-up when constitutive lactate release was inhibited. The rates of pyruvate accumulation were significantly decreased in LMO-, DLDH- and also LOx-expressing astrocytes and, considering that LDH expression itself remained unchanged, these results are consistent with LMO, LOx, and DLDH activity as an additional lactate sink in the cell.
4.2. DLDH
Direct recording of DL production in astrocytes was limited as we had no access to specific biosensors. Laconic is insensitive to DL [
19]. We were able to measure DL in conditioned media and indirectly assess the implications of DLDH activity for astrocytic lactate handling via pyruvate detection. Expression of DLDH significantly raised DL release from cells, although the difference with the current version of the construct was modest. Through improvements to the expression system, the yield can potentially be increased in the future.
In unstimulated conditions, in dissociated as well as organotypic slice cultures, lactate release was not significantly affected by DLDH activity in astrocytes. Transient disruption of the lactate/pyruvate/DL equilibrium during trans-acceleration, however, indicated that by diverting pyruvate towards DL synthesis, DLDH had an impact on the maintenance of the steady-state lactate pool.
Endogenous DL production in the brain may occur through the glyoxalase pathway, so the brain must be adequately equipped to process this metabolite. Toxic levels as associated with short bowel syndrome or in the context of diabetic complications cause DL encephalopathy and can interfere severely with brain function [
23]. Interestingly, DL has also been suggested as a neuroprotective agent in ischemia [
24].
DL shares various molecular targets, such as transporters and receptors, with lactate. For example, MCT1 pumps DL, albeit with a lower affinity and at a slower rate than lactate, or DL activates the lactate receptor HCA1 but with a lower potency than lactate [
10,
25,
26]. To our knowledge, a neurophysiological signalling role of DL has not been described, although DL is known to interfere with memory, possibly through inhibition of cellular respiration or through a receptor-dependent mechanism [
27,
28,
29].
With low levels of DL production by astrocytic DLDH expression, we would, therefore, not expect any significant actions of DL via the above targets. We have, however, previously described DL as a potent antagonist which inhibited lactate-induced stimulation of noradrenaline release from locus coeruleus neurons via an unidentified receptor-mediated mechanism [
4]. For this target, even moderately increased DL levels, as seen with our DLDH expression system, could potentially have a significant effect.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






