Electrochemical Response of Saccharomyces cerevisiae Corresponds to Cell Viability upon Exposure to Dioclea reflexa Seed Extracts and Antifungal Drugs

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Methods
2.1. Growth Medium and Cell Culturing
2.2. Seed Drying and Extraction and Drug Acquisition
2.3. Solvent Extraction
2.4. Cell Viability Measurements Using Trypan Blue Based Assay
3. Results
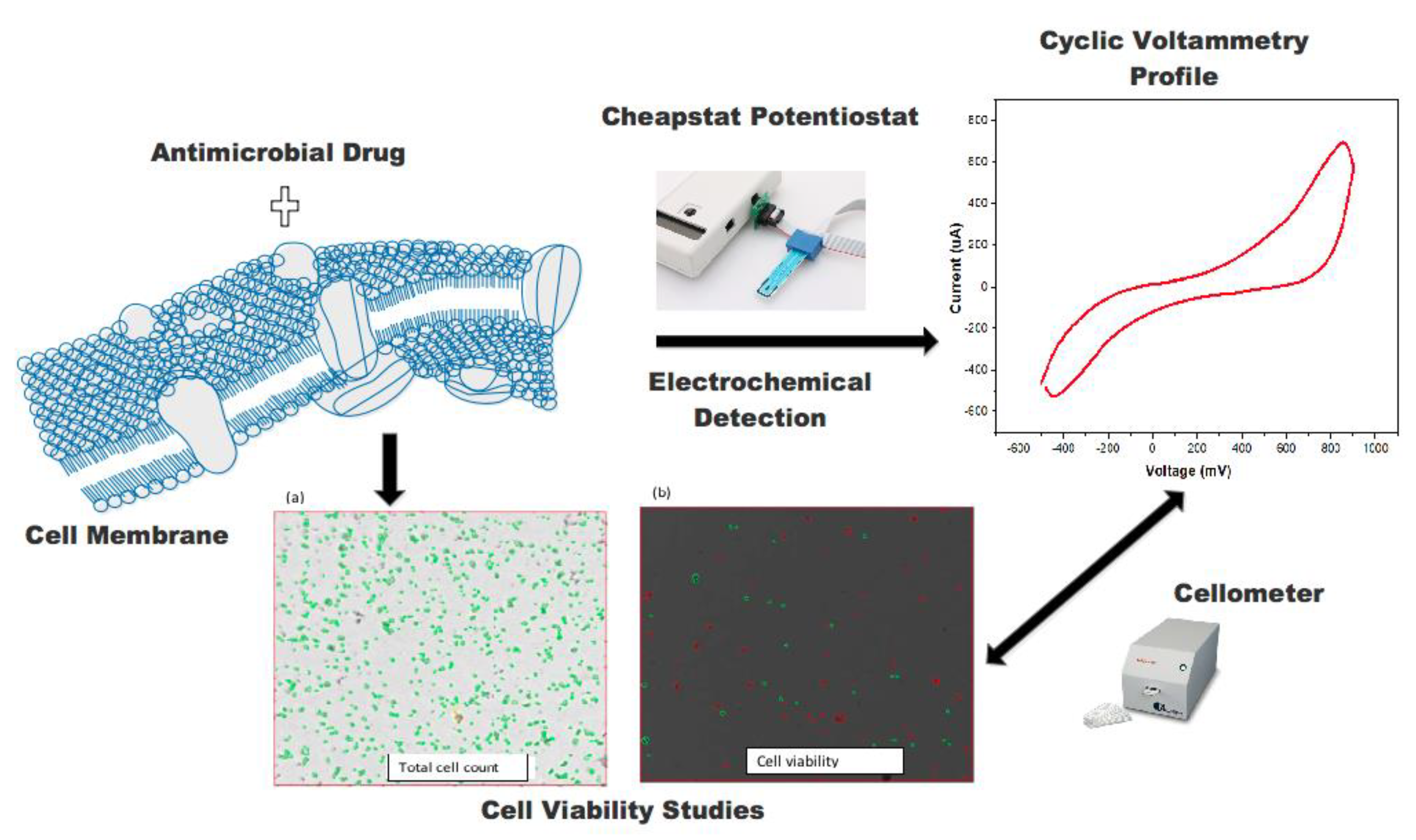
3.1. Structure of the Antifungal Drugs and Schematic of the Study
3.2. UV-VIS Spectrophotometry Studies
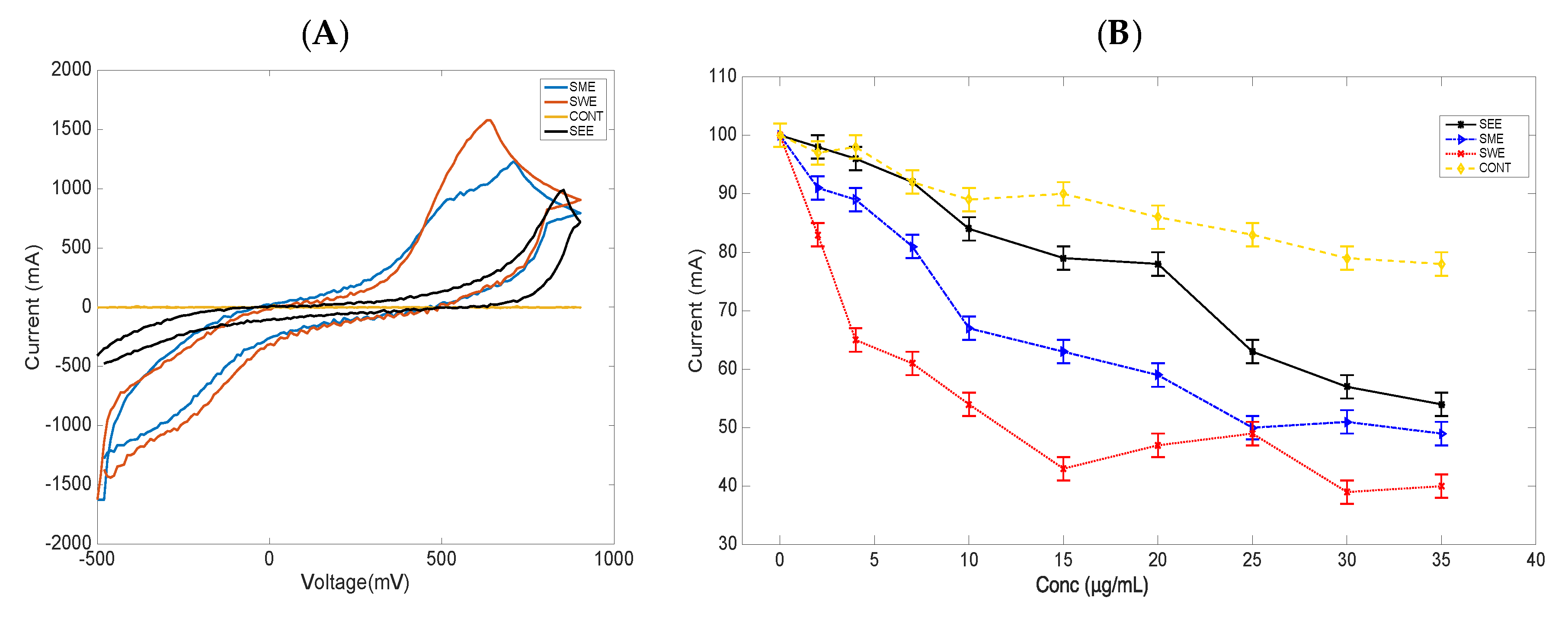
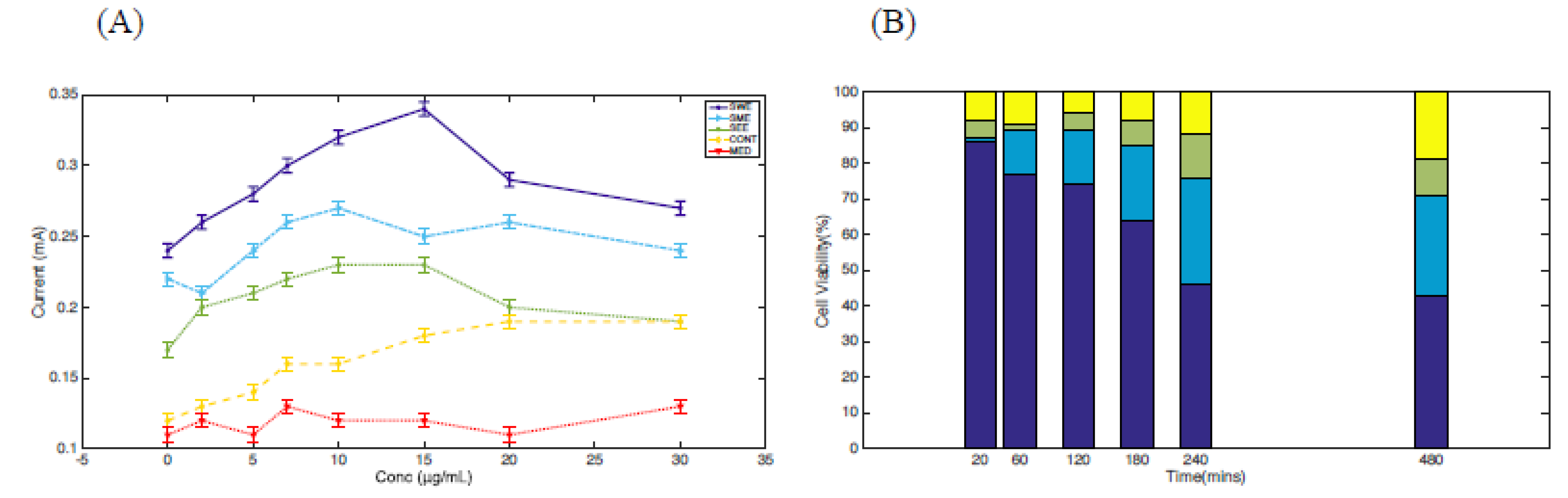
3.3. Cyclic Voltammetry and Cell Viability Studies of S. cerevisiae Cells Treated with Extracts
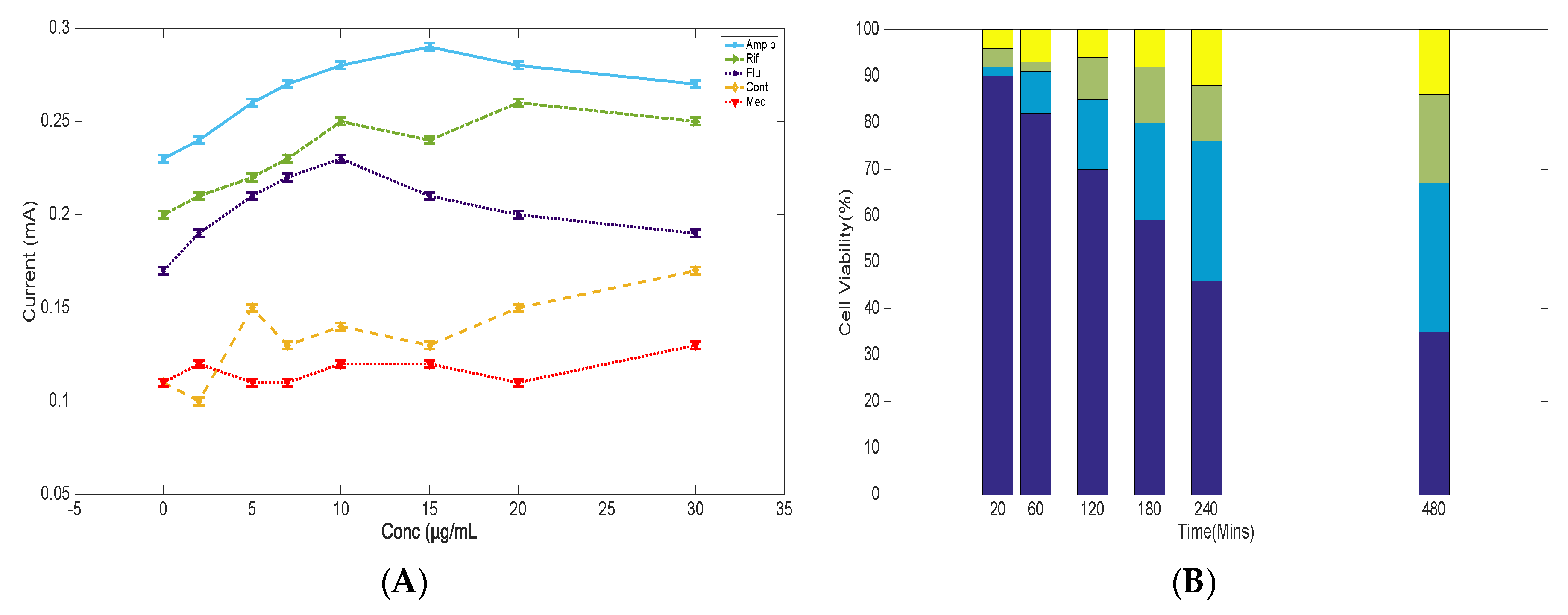
3.4. Cyclic Voltammetry Studies of S. cerevisiae in the Presence of Antifungal Drugs
3.5. Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nikitin, M.; Deych, K.; Grevtseva, I.; Girsova, N.; Kuznetsova, M.; Pridannikov, M.; Dzhavakhiya, V.; Statsyuk, N.; Golikov, A. Preserved Microarrays for Simultaneous Detection and Identification of Six Fungal Potato Pathogens with the Use of Real-Time PCR in Matrix Format. Biosensors 2018, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Antunes, R.S.; Ferraz, D.; Garcia, L.F.; Thomaz, D.V.; Luque, R.; Lobon, G.S.; Gil, E.S.; Lopes, F.M. Development of a Polyphenol Oxidase Biosensor from Jenipapo Fruit Extract (Genipa americana L.) and Determination of Phenolic Compounds in Textile Industrial Effluents. Biosensors 2018, 8, 47. [Google Scholar] [CrossRef]
- Anderson, D.E.; Balapangu, S.; Fleischer, H.N.A.; Viade, R.A.; Krampa, F.D.; Kanyong, P.; Awandare, G.A.; Tiburu, E.K. Investigating the Influence of Temperature on the Kaolinite-Base Synthesis of Zeolite and Urease Immobilization for the Potential Fabrication of Electrochemical Urea Biosensors. Sensors 2017, 17, 1831. [Google Scholar] [CrossRef]
- Lee, W.; Lee, D.G. A novel mechanism of fluconazole: Fungicidal activity through dose-dependent apoptotic responses in Candida albicans. Microbiology 2018, 164, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Durnas, B.; Wnorowska, U.; Pogoda, K.; Deptula, P.; Watek, M.; Piktel, E.; Gluszek, S.; Gu, X.; Savage, P.B.; Niemirowicz, K.; et al. Candidacidal Activity of Selected Ceragenins and Human Cathelicidin LL-37 in Experimental Settings Mimicking Infection Sites. PLoS ONE 2016, 11, e0157242. [Google Scholar] [CrossRef] [PubMed]
- Palanisami, A.; Mercier, G.T.; Fang, J.; Miller, J.H., Jr. Nonlinear impedance of whole cells near an electrode as a probe of mitochondrial activity. Biosensors 2011, 1, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.C.; Gunning, C.; Oliveira, C.M.; Couto, J.A.; Teixeira, J.A.; Martins, R.C.; Ferreira, A.C. Saccharomyces cerevisiae oxidative response evaluation by cyclic voltammetry and gas chromatography-mass spectrometry. J. Agric. Food Chem. 2012, 60, 7252–7261. [Google Scholar] [CrossRef]
- Joseph-Horne, T.; Wood, P.M.; Wood, C.K.; Moore, A.L.; Headrick, J.; Hollomon, D. Characterization of a split respiratory pathway in the wheat “take-all” fungus, Gaeumannomyces graminis var. tritici. J. Biol. Chem. 1998, 273, 11127–11133. [Google Scholar] [CrossRef]
- Rawson, F.J.; Downard, A.J.; Baranian, K.H. Electrochemcal Deection of Intracellular and Cell Membrane Redox System in Saccharomyces cerevisiae. Sci. Rep. 2014, 4, 5216–5223. [Google Scholar] [CrossRef]
- Tiburu, E.K.; Mutocheluh, M.; Arthur, P.K.; Narkwa, P.W.; Salifu, A.A.; Agyei, M.A.; Yeboah, R.; Fleischer, H.N.A.; Zhuang, A.G. Antifungal and Anti-Proliferative Effects of Zeolites A and X on Yeast Pathogenic and Cancer Cells In Vitro. J. Biomater. Tissue Eng. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Koehn, F.E. High impact technologies for natural products screening. Prog. Drug Res. 2008, 65, 175–210. [Google Scholar]
- Rios, J.L.; Recio, M.C.; Villar, A. Screening methods for natural products with antimicrobial activity: A review of the literature. J. Ethnopharmacol. 1988, 23, 127–149. [Google Scholar] [CrossRef]
- Fang, J.; Palanisami, A.; Rajapakshe, K.; Widger, W.R.; Miller, J.H. Nonlinear dielectric spectroscopy as an indirect probe of metabolic activity in thylakoid membrane. Biosensors 2011, 1, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Oladele Oladimeji, A.; Adebayo Oladosu, I.; Shaiq Ali, M.; Lateef, M. Dioclins A and B, new antioxidant flavonoids from Dioclea reflexa. Nat. Prod. Res. 2018, 32, 2017–2024. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Junior, V.R.; Correia, J.L.; Pereira, R.I.; Pereira-Junior, F.N.; Santiago, M.Q.; Osterne, V.J.; Madeira, J.C.; Cajazeiras, J.B.; Nagano, C.S.; Delatorre, P.; et al. Purification and molecular characterization of a novel mannose-specific lectin from Dioclea reflexa hook seeds with inflammatory activity. J. Mol. Recognit. 2016, 29, 134–141. [Google Scholar] [CrossRef]
- Hassan, R.Y.; Bilitewski, U. Direct electrochemical determination of Candida albicans activity. Biosens. Bioelectron. 2013, 49, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro Rde, A.; de Oliveira, J.S.; de Castelo-Branco, D.S.; Teixeira, C.E.; Marques, F.J.; Bittencourt, P.V.; Carvalho, V.L.; de Bandeira, T.J.; Brilhante, R.S.; Moreira, J.L.; et al. Candida tropicalis isolates obtained from veterinary sources show resistance to azoles and produce virulence factors. Med. Mycol. 2015, 53, 145–152. [Google Scholar] [CrossRef]
- Lord, A.T.; Mohandas, K.; Somanath, S.; Ambu, S. Multidrug resistant yeasts in synanthropic wild birds. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 11. [Google Scholar] [CrossRef]
- Montineri, A.; Iacobello, C.; Larocca, L.; La Rosa, R.; Nigro, L.; Fatuzzo, F. [Saccharomyces cerevisiae fungemia associated with multifocal pneumonia in a patient with alcohol-related hepatic cirrhosis]. Infez. Med. 2008, 16, 227–229. [Google Scholar] [PubMed]
- Campbell, E.A.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural mechanism for rifampicin inhibition of bacterial rna polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Bolard, J.; Legrand, P.; Heitz, F.; Cybulska, B. One-sided action of amphotericin B on cholesterol-containing membranes is determined by its self-association in the medium. Biochemistry 1991, 30, 5707–5715. [Google Scholar] [CrossRef] [PubMed]
- Gray, K.C.; Palacios, D.S.; Dailey, I.; Endo, M.M.; Uno, B.E.; Wilcock, B.C.; Burke, M.D. Amphotericin primarily kills yeast by simply binding ergosterol. Proc. Natl. Acad. Sci. USA 2012, 109, 2234–2239. [Google Scholar] [CrossRef] [Green Version]
- Brito, A.; Ramirez, J.E.; Areche, C.; Sepulveda, B.; Simirgiotis, M.J. HPLC-UV-MS Profiles of Phenolic Compunds and Antioxidant Activity of Fruits from Three Citrus Species Consumed in Northern Chile. Molecules 2014, 10, 17400–17421. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, Z.; Han, X.; Tang, J.; Wang, J.; Dong, S.; Wang, E. Ion channel behavior of amphotericin B in sterol-free and cholesterol- or ergosterol-containing supported phosphatidylcholine bilayer model membranes investigated by electrochemistry and spectroscopy. Biophys. J. 2002, 83, 3245–3255. [Google Scholar] [CrossRef]
- Ibragimova, V.; Alieva, I.; Kasumov, K.; Khutorsky, V. Transient permeability induced by alkyl derivatives of amphotericin B in lipid membranes. Biochim. Biophys. Acta 2006, 1758, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dymond, A.W.; So, K.; Martin, P.; Huang, Y.; Severin, P.; Mathews, D.; Lisbon, E.; Mariani, G. Effects of cytochrome P450 (CYP3A4 and CYP2C19) inhibition and induction on the exposure of selumetinib, a MEK1/2 inhibitor, in healthy subjects: Results from two clinical trials. Eur. J. Clin. Pharmacol. 2017, 73, 175–184. [Google Scholar] [CrossRef]
- Serhan, G.; Stack, C.M.; Perrone, G.G.; Morton, C.O. The polyene antifungals, amphotericin B and nystatin, cause cell death in Saccharomyces cerevisiae by a distinct mechanism to amphibian-derived antimicrobial peptides. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Delattin, N.; Cammue, B.P.; Thevissen, K. Reactive oxygen species-inducing antifungal agents and their activity against fungal biofilms. Future Med. Chem. 2014, 6, 77–90. [Google Scholar] [CrossRef]
- Enache-Angoulvant, A.; Hennequin, C. Invasive Saccharomyces infection: A comprehensive review. Clin. Infect. Dis. 2005, 41, 1559–1568. [Google Scholar] [CrossRef]
- Sessa, G.; Weissmann, G. Effect of polyene antibiotics of phospholipid spherules containing varying amounts of charged components. Biochim. Biophys. Acta 1967, 135, 416–426. [Google Scholar] [CrossRef]
- Stoodley, R.; Shepherd, J.; Wasan, K.M.; Bizzotto, D. Amphotericin B interactions with a DOPC monolayer. Electrochemical investigations. Biochim. Biophys. Acta 2002, 1564, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Segneanu, A.E.; Grozescu, I.; Cziple, F.; Berki, D.; Damian, D.; Niculite, C.M.; Florea, A.; Leabu, M. Helleborus purpurascens-Amino Acid and Peptide Analysis Linked to the Chemical and Antiproliferative Properties of the Extracted Compounds. Molecules 2015, 20, 22170–22187. [Google Scholar] [CrossRef] [PubMed]
- Salamifar, S.E.; Lai, R.Y. Use of combined scanning electrochemical and fluorescence microscopy for detection of reactive oxygen species in prostate cancer cells. Anal. Chem. 2013, 85, 9417–9421. [Google Scholar] [CrossRef]
- Rawson, F.J.; Cole, M.T.; Hicks, J.M.; Aylott, J.W.; Milne, W.I.; Collins, C.M.; Jackson, S.K.; Silman, N.J.; Mendes, P.M. Electrochemical communication with the inside of cells using micro-patterned vertical carbon nanofibre electrodes. Sci. Rep. 2016, 6, 37672. [Google Scholar] [CrossRef] [Green Version]
- Hashkavayi, A.B.; Raoof, J.B.; Ojani, R.; Kavoosian, S. Ultrasensitive electrochemical aptasensor based on sandwich architecture for selective label-free detection of colorectal cancer (CT26) cells. Biosens. Bioelectron. 2017, 92, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Haupt, A.; Campetelli, A.; Bonazzi, D.; Piel, M.; Chang, F.; Minc, N. Electrochemical regulation of budding yeast polarity. PLoS Biol. 2014, 12, e1002029. [Google Scholar] [CrossRef] [PubMed]
- Hudcova, K.; Trnkova, L.; Kejnovska, I.; Vorlickova, M.; Gumulec, J.; Kizek, R.; Masarik, M. Novel biophysical determination of miRNAs related to prostate and head and neck cancers. Eur. Biophys. J. 2015, 44, 131–138. [Google Scholar] [CrossRef]
- Nguyen, H.V.; Richtera, L.; Moulick, A.; Xhaxhiu, K.; Kudr, J.; Cernei, N.; Polanska, H.; Heger, Z.; Masarik, M.; Kopel, P.; et al. Electrochemical sensing of etoposide using carbon quantum dot modified glassy carbon electrode. Analyst 2016, 141, 2665–2675. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.C.; Trevijano-Contador, N.; Roman, E.; Sanchez-Fresneda, R.; Casas, C.; Herrero, E.; Arguelles, J.C.; Pla, J.; Cuenca-Estrella, M.; Zaragoza, O. The production of reactive oxygen species is a universal action mechanism of Amphotericin B against pathogenic yeasts and contributes to the fungicidal effect of this drug. Antimicrob. Agents Chemother. 2014, 58, 6627–6638. [Google Scholar] [CrossRef] [PubMed]
- Speiser, B. From cyclic voltammetry to scanning elec- trochemical microscopy: Modern electroanalytical methods to study organic compounds, materials and reactions. Curr. Org. Chem. 1999, C3, 171–191. [Google Scholar]
- Heatley, N.G. A method for the assay of penicillin. Biochem. J. 1944, 38, 61–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acworth, I.N.; Bailey, B.A.; Maher, T.J. The use of HPLC with electrochemical detection to monitor reactive oxygen and nitrogen species, markers of oxidative damage and antioxidants: Application to the neurosciences. Prog. HPLC-HPCE 1998, 7, 3–56. [Google Scholar]
- Shuker, S.B.; Hajduk, P.J.; Meadows, R.P.; Fesik, S.W. Discovering high-affinity ligands for proteins: SAR by NMR. Science 1996, 274, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Hsu, C.H.; Wen, Z.H.; Lin, C.S. Recent advances of fluorescent technologies for drug discovery and development. Curr. Pharm. Des. 2009, 15, 3552–3570. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Strategy | Sample Size | Robustness | Incubation Time (h) | Disadvantages |
|---|---|---|---|---|---|
| The current Method | Uses Interdigitated electrode and Cellometer | microliters | Fast and Sensitive | 0.3–8 | Preliminary Target not known |
| Fluorescence techniques | Based on staining and counter staining | milliliters | Complex architecture | long | phototoxic-unclear images [44] |
| High Performance Liquid Chromatography-Electrochemical Detection (HPLC-ECD) | A separation technique | microliters | selective and sensitive | Relatively fast | Expensive [44] |
| Structural Activity Relationship by NMR (SAR by NMR) | nuclear magnetic resonance (NMR)-based | microliters | target-directed drug research | Short | Focuses on hit validation studies [43] |
| Diffusion Methods | Agar plates are inoculated and compared to standard | large | Simplicity and low cost | 16–24 | Inaccuracies [43] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arthur, P.K.; Yeboah, A.B.; Issah, I.; Balapangu, S.; Kwofie, S.K.; Asimeng, B.O.; Foster, E.J.; Tiburu, E.K. Electrochemical Response of Saccharomyces cerevisiae Corresponds to Cell Viability upon Exposure to Dioclea reflexa Seed Extracts and Antifungal Drugs. Biosensors 2019, 9, 45. https://doi.org/10.3390/bios9010045
Arthur PK, Yeboah AB, Issah I, Balapangu S, Kwofie SK, Asimeng BO, Foster EJ, Tiburu EK. Electrochemical Response of Saccharomyces cerevisiae Corresponds to Cell Viability upon Exposure to Dioclea reflexa Seed Extracts and Antifungal Drugs. Biosensors. 2019; 9(1):45. https://doi.org/10.3390/bios9010045
Chicago/Turabian StyleArthur, Patrick Kobina, Anthony Boadi Yeboah, Ibrahim Issah, Srinivasan Balapangu, Samuel K. Kwofie, Bernard O. Asimeng, E. Johan Foster, and Elvis K. Tiburu. 2019. "Electrochemical Response of Saccharomyces cerevisiae Corresponds to Cell Viability upon Exposure to Dioclea reflexa Seed Extracts and Antifungal Drugs" Biosensors 9, no. 1: 45. https://doi.org/10.3390/bios9010045